
Abstract
Forests are major sources of terrestrial CH4 and CO2 fluxes but not all surfaces within forests have been measured and accounted for. Stem respiration is a well-known source of CO2, but more recently tree stems have been shown to be sources of CH4 in wetlands and upland habitats. A study transect was established along a natural moisture gradient, with one end anchored in a forested wetland, the other in an upland forest and a transitional zone at the midpoint. Stem and soil fluxes of CH4 and CO2 were measured using static chambers during the 2013 and 2014 growing seasons, from May to October. Mean stem CH4 emissions were 68.8 ± 13.0 (mean ± standard error), 180.7 ± 55.2 and 567.9 ± 174.5 µg m−2 h−1 for the upland, transitional and wetland habitats, respectively. Mean soil methane fluxes in the upland, transitional and wetland were − 64.8 ± 6.2, 7.4 ± 25.0 and 190.0 ± 123.0 µg m−2 h−1, respectively. Measureable CH4 fluxes from tree stems were not always observed, but every individual tree in our experiment released measureable CH4 flux at some point during the study period. These results indicate that tree stems represent overlooked sources of CH4 in forested habitats and warrant investigation to further refine CH4 budgets and inventories.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Atmospheric methane (CH4) concentrations have increased from 700 ppb to over 1800 ppb since the beginning of the industrial revolution and presently contribute 0.7 W m−2 or 25% of radiative forcing (IPCC 2013). Although the net balance of CH4 sources and sinks is well constrained compared to trace gases other than CO2, the relative contributions of individual sources and sinks are less certain (Kirschke et al. 2013; Saunois et al. 2016). Such uncertainty has made it difficult to explain phenomena such as changes in the globally averaged atmospheric growth rate and isotopic concentration of methane (Aydin et al. 2011; Nisbet et al. 2016) and exposed the limits of our current mechanistic understanding of CH4 cycling. In response, the past decade has been a period of prospecting for novel CH4 sinks and sources that might better describe CH4 cycle dynamics.
Wetlands have always been considered a source of CH4, which is emitted across both the soil–atmosphere interface and plant surfaces (Dacey and Klug 1979). Emission from herbaceous wetland plants is facilitated by aerenchyma tissue which allows rapid rates of gas exchange between soils and the atmosphere, supporting aerobic respiration but also diffusion and mass flow of CH4 past oxic zones at the soil surface.
While whole-ecosystem (plant and soil) CH4 emissions have been measured extensively in wetlands dominated by herbaceous plants (Conrad 2007; Dacey and Klug 1979), such data are generally lacking from woody plants such as trees because their large stature makes plant flux measurements difficult. Early studies demonstrated CH4 emissions from woody wetland tree roots (Pulliam 1992; Rusch and Rennenberg 1998), and seedlings (Garnet et al. 2005; Vann and Megonigal 2003), but field measurements to quantify tree CH4 emissions were conducted only in the last decade (Gauci et al. 2010; Pangala et al. 2015; Terazawa et al. 2007). The results indicate that tree-mediated CH4 emissions have been overlooked and may account for 60–87% of total CH4 efflux in tropical wetlands (Pangala et al. 2013) and 20% in temperate wetlands (Gauci et al. 2010). Given that forested wetlands represent 53% of total wetlands (Fung et al. 1987), these numbers are significant, yet have not been included global earth systems models and budgets (Saunois et al. 2016).
Upland forests have been generally considered net sinks of CH4 based upon the assumption that the only surface in a forest that interacts with CH4 is the soil. Studies have shown that CH4 can be produced inside upland trees (Bushong 1907; Covey et al. 2012; Zeikus and Ward 1974), however, few in situ direct measurements have attempted to quantify net fluxes from trees (Machacova et al. 2016; Maier et al. 2017; Pitz and Megonigal 2017; Wang et al. 2016; Warner et al. 2017). Wang et al. (2016) estimated that tree CH4 flux was sufficient to offset the soil sink by 5–10% on an annual basis. Machacova et al. (2016) suggested that depending on soil moisture, Scots pine CH4 emission account up to 35% of soil uptake. Clearly, tree–atmosphere trace gas interactions cannot be ignored and have to be included in calculating CH4 budgets in forested ecosystems.
The processes by which CH4 is produced and emitted to the atmosphere through trees are poorly understood. Data are insufficient for developing a generalized conceptual model of tree CH4 emissions that captures the wide range of species, ecosystems, and conditions. Studies in wetland forests generally show a positive relationship between tree emission rates and water table depth (Gauci et al. 2010; Pangala et al. 2015; Terazawa et al. 2007), suggesting that CH4 produced under saturated, anaerobic conditions in groundwater becomes entrained in the transpiration stream or diffuses into plant tissue, where it is transported and eventually emitted to the atmosphere. In contrast, a more diverse set of mechanisms have been proposed in upland forests, including non-soil CH4 sources such as UV-driven aerobic production (Keppler et al. 2008; Vigano et al. 2008) and anaerobic biological production in trunks associated with heart rot (Covey et al. 2012) or non-structural carbohydrates (Covey et al. 2016). Megonigal and Guenther (2008) hypothesized that groundwater is also a source of CH4 emitted by upland forest tree species. In this case, deep roots growing in CH4-rich groundwater or anoxic soil microsites could entrain CH4 and transport it to the atmosphere, bypassing the oxic soil horizons where it would otherwise be consumed by methanotrophs. A growing list of studies indicates that a variety of plant-mediated CH4 sources exist and that all ecosystems contain some surfaces that have the potential to emit CH4.
Gradient studies can reveal insights into some of these processes as they vary across wetland and upland forests, but studies reported thus far have focused on only one habitat. In this study, we conducted the first CH4 flux measurement along a soil moisture gradient from wetland to upland at the same location. The close proximity of upland and wetland allows potentially confounding variables such as climate, past land use, and, to some extent, plant community composition to be kept constant. We directly measured CH4 fluxes from soils and stems from a variety of tree species that are common in mid-Atlantic deciduous forests.
The goals of this study were to: (1) quantify CH4 emissions from trees growing across a soil moisture gradient in a temperate forest ecosystem, (2) compare the relative contribution of soils and trees in upland and wetland forests, and (3) relate these fluxes to environmental factors. We also report stem and soil CO2 fluxes because CO2 emissions from stem respiration are relatively well understood and thus help to interpret the pattern of CH4 fluxes.
Methods
Study site
The study was conducted at the Smithsonian Environmental Research Center (SERC), a property of 1072 hectares (2650 acres) on the western shore of Chesapeake Bay in Maryland. Much of the site is forested with smaller areas of brackish tidal wetlands and farmland. Forests have been recovering for 70–150 years from different land use and disturbance histories such as logging, wind damage, and agricultural abandonment (Higman 1968; Yesilonis et al. 2016), with small patches that have no known history of land use. Our main study plot was in an upland forest that was most likely grazed before the Civil War and then abandoned (Higman 1968). Today the forest is dominated by Tulip poplar (Liriodendron tulipifera), American beech (Fagus grandfolia), and several species of oaks (Quercus spp.), and hickories Carya spp. The species composition is typical of the mature stage of a Tulip poplar association (Brown and Parker 1994; Brush et al. 1980) with a closed canopy and very little understory. Mean rainfall is 1146 mm and mean annual temperature is 13.0 °C (Correll, Jordan, and Duls, unpublished data). The mean annual maximum temperature is 19.0 °C and the mean minimum temperature is 8.0 °C (NCDC database, Annapolis Police Bar Station).
Soils at SERC are predominately fine sandy loams or sandy loams. Physical and chemical characteristics of surface soils reflect past land use history, forest age and non-native earthworm activity (Yesilonis et al. 2016). Our transect crossed three soil associations, with the upland and transitional sections in the Collington–Annapolis series and the Collington–Wist–Westphalia series, respectively. Soils in the wetland section transect were in the Widewater–Issue series (Natural-Resources-Conservation-Service 2016).
We established an approximately 150 m long transect along a soil moisture gradient (location 38.8878, − 76.5624). The elevation difference between the two end points of the transect was approximately 6 m. Based upon soil characteristics, elevation, and water table depth we divided the transect into three habitat types: upland (100 m), transitional (25 m) and wetland (25 m). Thirty-two trees selected for the study belonged to nine species (Table 1). Based upon stem counts, these nine species make up 80% of the mature stand adjacent to the transect (Parker and Tibbs 2004). Liquidambar styraciflua (sweetgum) occurred in all three habitat types, with the remaining species present in one or two habitats.
Stem and soil chambers, and flux measurements
Tree and soil measurements were made between May and November in 2013 and between May and September in 2014 using the closed chamber technique. A total of 32 trees were fitted with opaque rectangular chambers modified from Ryan (1990), originally designed to measure stem respiration. In 2013, 21 chambers were installed across the transect. In 2014, 10 additional trees were fitted with chambers in order to expand the upland section of the transect. Each tree was paired with a soil gas flux chamber placed within 1 m of the base. Sampling rounds differed among habitats with the upland habitat sampled more frequently in 2014 than the other two habitats, because our main interest was to quantify upland methane stem fluxes. Rectangular stem chambers were constructed of acrylic, permanently fixed to stems 30–60 cm above the soil, and were secured to the stem using elastic shock cord. Each chamber was 28 cm in height with varying depths and widths depending on the tree size. To create an airtight seal, closed-cell neoprene foam was placed between the chamber edge and the stem, and sealed with dental mold to create a non-VOC seal (Examix™, GC America, Alsip, IL, USA). Soil flux chambers were constructed out of 30.5 cm-diameter (12′′) schedule 80 PVC pipe, machined into a 10-cm high ring and placed 5 cm into the soil surface. All chambers were in place for a minimum of 1 week before taking flux measurements, and once mounted they remained in place for the duration of the study.
In the 2 years of study we used two different instruments to analyze the gas samples from the headspace of the flux chambers. Gas samples from the headspace of the flux chambers were analyzed by gas chromatography (GC) in 2013 and by a more accurate and precise portable off-axis integrated cavity output spectroscope (OA-ICOS) (Ultra-Portable Greenhouse Gas Analyzer, Los Gatos Research, Mountain View, CA, USA) in 2014. The two instruments provided different amounts of data for each flux which required a different approach, described below, for the statistical analysis.
In 2013, gas concentrations in air samples were determined using a gas chromatograph. After closing the chamber lid, 12 mL samples were withdrawn by syringe at 0, 15, 30, 45, and 60 min. The air samples were immediately transferred from the syringe to a 12 mL, nitrogen-flushed exetainers (Labco, UK). The gas samples were analyzed for CH4 and CO2 on a Varian GC-450, equipped with a flame ionization detector (FID) and a thermal conductivity detector (TCD). The FID had a precision of 0.120 ppm for CH4 and the TCD had a precision 5 ppm for CO2.
In 2014, gas concentrations were measured using a portable OA-ICOS. The instrument is capable of measuring CH4 within a range of 0.01–100 ppm with a precision of 0.002 ppm at 0.5 Hz. The OA-ICOS can also measure CO2 in a range of 200–20,000 ppm with a precision of 0.3 ppm. The OA-ICOS was used as a closed system: headspace gas was drawn from the chamber, measured non-destructively for CH4 and CO2 concentration, and returned to the flux chamber as described in Baird et al. (2010). Changes in CH4 and CO2 concentration were measured over periods of approximately 5–30 min, during which the system generated ≥ 150 observations.
In 2013, five concentration data points were collected for each flux measurement. We occasionally dropped a concentration data point for two reasons; soil CH4 ebullition or poor quality data from the GC. If a data point was dropped due to poor quality data from the GC, both CH4 and CO2 data had to support that conclusion. The slope of gas concentration change over time was determined by linear regression (SAS® procedure Proc Reg). In every case, the slope was based on ≥ 4 observations. For a slope to be determined as a quality data point, the R2 had to be greater than 0.90. If the R2 of a slope was less than 0.90, then the slope and flux was considered to be zero.
In 2014, the OA-ICOS provided concentration data at a rate of 0.5 Hz which required us to use a slightly different treatment of the data. When calculating the slope of gas concentration change over time, we ignored the first 20% of the observations, as those may produce false readings associated with closing the lid. The slope of gas concentration change was determined as described above (SAS® procedure Proc Reg). In every case, the slope was based on ≥ 120 observations.
Gas flux was calculated using the following equation:
where F is the flux in μg m−2 h−1, P is atmospheric pressure, T is temperature, R is the universal gas constant, A is the collar surface area and V is the volume of the air enclosed by the chamber. Air temperature was measured by the OA-ICOS unit on gas circulating between the unit and the chamber. Atmospheric pressure was based on a nearby weather station (< 1 km). Flux units are reported in µg m−2 h−1 or mg m−2 h−1 to allow for direct comparison with stem flux data published by others.
Environmental data
Soil moisture was measured using a FieldScout TDR (Spectrum Technologies Inc., Aurora, Illinois, USA). Soil temperature was measured with a digital thermometer at 10 cm. Weather data was collected on site from the SERC weather station (Campbell Scientific, Utah, USA).
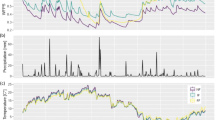
Water table depth or groundwater elevation along the transect was monitored using several monitoring wells. Four 5.08 cm diameter wells were installed (three in 2013, and one was added in 2014). One was installed in the wetland, one in the transitional zone, and two in the upland. The fourth well was added to the upland in 2014 when new trees were added to the transect. Wells were constructed of 5.08 cm PVC and screened with 152 cm sections of PVC with 0.25 mm slot size. Number #1 sand was used as a screen pack. Water table depth was recorded manually during each sampling event using a water level meter (Model 102 Water Level Meter, Solinist, Georgetown, Ontario, Canada). Well #2 in the transitional was monitored during the growing seasons in 2013 and 2014 (Fig. 1) using a groundwater elevation logger (Aqua Troll 200, In-Situ Inc., Fort Collins, Colorado, USA).

Daily mean air temperature (a), daily precipitation (b) and groundwater elevation (c) at the SERC study site during 2013 and 2014. A continuous groundwater elevation data logger was placed in a well (Well 2) within the transitional habitat; groundwater elevation in the three other wells located within the study site were recorded manually (see “Methods”). The water table elevation data logger was removed for 3 months in the winter of 2014
Statistical analysis
All statistical analyses on flux data were conducted using R v3.1.2 (R Core Team 2014), except gas fluxes that were calculated using SAS. Methane flux rates were Box–Cox transformed after increasing all values until the minimum value in the data set was 10 µg m−2 h−1 to avoid negative values after log transformation. Means and standard errors presented in the text, figures, and tables were calculated using non-transformed data. P values below 0.05 were considered significant; those between 0.05 and 0.1 were considered marginally significant. In this paper we report data on CH4 and CO2 fluxes from three habitat types (upland, transitional, and wetland) in 2013 and 2014. Upland stem and soil CH4 flux data from 2014 were reported in Pitz and Megonigal (2017) but were combined in the present study for statistical analysis.
The mean CH4 and CO2 fluxes were calculated for each tree and the respective soil chambers and analyzed for the effects of habitats using analysis of variance (ANOVA) followed by the Tukey’s HSD test for multiple comparisons. Mixed effect models were conducted using the lme4 package to evaluate the correlations between environmental factors and stem CH4 flux, soil CH4 flux, stem CO2 flux, and soil CO2 flux. Sampling round and tree identity were treated as random effects. Fixed effects included were habitat for all analyses, tree species and diameter at breast height (DBH) for stem fluxes, DTW, soil moisture and soil temperature for CH4 fluxes, and soil moisture and soil temperature for CO2 fluxes. These factors were evaluated in the models following the above order. Likelihood ratio tests were used to access significant differences between nested models, and were followed by the Tukey’s HSD test for multiple comparisons using the multcomp package (Hothorn et al. 2008).
Results
A total of 470 flux measurements were made during the study, 235 measurements of tree stems and 235 of soils. Every tree stem emitted measurable CH4 at least once during the two seasons of monitoring. Net CH4 production was detected in all but two stem CH4 measurements. Stem CH4 fluxes ranged from − 8.1 to 1900 µg m−2 h−1 in the upland habitat, − 16.4 to 2146.2 µg m−2 h−1 in the transitional habitat and − 157.1 to 3757.6 µg m−2 h−1 in the wetland habitat of the transect (Fig. 2a). Soil was generally a CH4 sink in the upland where 97% of the measurements showed net consumption, and a source in the wetland where 80% of the measurements showed net production. The transitional segment of the transect fell in between, with 74% of the measurements demonstrating net consumption (Figs. 2a). Soil CH4 fluxes ranged from − 309.3 to 68.1 µg m−2 h−1 in the upland habitat, − 167.4 to 651.4 µg m−2 h−1 in the transitional habitat and − 33.3 to 4726.9 µg m−2 h−1 in the wetland habitat of the transect (Figs. 2a, 5b). Neither stem nor soil CH4 fluxes exhibited a seasonal trend; rather high fluxes compared to average values were few and episodic (Fig. 2a).

Temporal changes of stem and soil CH4 (a) and CO2 (b) fluxes in the three habitat types. Data for 2013–2014 are combined. 2013 fluxes are represented by circles and 2014 fluxes are represented by triangles. Error bars are standard error. Note the different scales
Stem and soil CO2 fluxes were consistently positive and three orders of magnitude higher than CH4 fluxes (Fig. 2b). Both stem and soil CO2 fluxes showed a seasonal trend with high values in the growing season and gradually declining values in the fall (Fig. 2b).
Combining all stem CH4 measurements by habitat yielded mean (± SE) rates of 68.8 ± 13.0 µg m−2 h−1 (upland), 180.7 ± 55.2 µg m−2 h−1 (transitional) and 567.9 ± 174.5 µg m−2 h−1 (wetland). Stem CH4 fluxes were significantly different in the three habitat types (F2, 29 = 5.80, P = 0.0076), and were higher in wetlands than in uplands (P = 0.006, Tukey’s HSD test) (Fig. 3a). Mean soil CH4 fluxes were − 64.8 ± 6.2 µg m−2 h−1 (upland), 7.4 ± 25.0 µg m−2 h−1 (transitional), and 190.0 ± 123.0 µg m−2 h−1 (wetland). ANOVA showed significant differences in soil CH4 flux among the three habitats types (F2, 29 = 20.08, P < 0.001), and significantly higher fluxes in wetland and transitional (P < 0.001 and P = 0.012, respectively) than in upland (Fig. 3a).

Mean (± SE) stem and soil flux for CH4 (a) and CO2 (b) in the three habitat types at the SERC study site. Means with different letters are significantly different (Tukey’s HSD, P < 0.05). Tests for stem and soil were run separately, and differences are indicated by upper and low case letters, respectively. Note the different units for CH4 and CO2 flux
Stem CO2 fluxes were only marginally affected by habitat types (F2, 29 = 2.88, P = 0.073). Contrary to CH4, stem and soil CO2 fluxes showed opposite trends across habitats. Soil CO2 fluxes were significantly affected by habitat (F2, 29 = 4.90, P = 0.015), and were higher in upland than in wetland and transitional habitats (P = 0.011) (Fig. 3b). The significant effect of habitat was further supported by the mixed effect models (Table 2).
Stem CH4 fluxes were marginally affected by tree species (Fig. 4), positively related to DBH and soil temperature, and negatively related to depth to water table (DTW) (Table 2). Soil CH4 fluxes were negatively associated with DTW and positively associated with soil temperature. Both soil temperature and moisture showed significant positive relationships with stem and soil CO2 fluxes (Table 2).

Species and habitat effects on stem methane fluxes. a Four species in one habitat (upland); b One species (Liquidambar styraciflua, sweetgum) in three habitats. The species codes are LT (Liriodendron tulipifera, tulip poplar), FG (Fagus grandifolia, beech), LS (Liquidambar styraciflua, sweetgum), and Qsp (Quercus sp., oak). Weighted mean (± SE) flux is shown (see “Methods”). Number of trees is shown on top of each column
The range of stem CH4 fluxes varied greatly by tree species (Online Resource 2). We detected the highest emissions from green ash and sweetgum, and the lowest from oak and ironwood (Carpinus caroliniana). Tree species identity was significant even after taking into account the confounding effect of habitat (Table 2), and comparing stem fluxes only in the upland habitat On the other hand, habitat effects are clearly shown for stem CH4 fluxes for sweetgum, the only tree species found in all three habitat types (Fig. 4b).
Although there was a positive correlation between soil moisture and CH4 flux in the upland soil (Fig. 5b), soil moisture in general was not a significant variable in the mixed effects model (Table 2). Soil moisture co-varies with habitat and DTW, and was added to the model as the last of these three variables.

Correlation between methane flux and soil moisture in the three habitats at SERC. Data for 2013–2014 are combined. Left panels: stem fluxes; right panels: soil fluxes. Note the different scales on the y axes
Discussion
The present study is the first that simultaneously explores stem and soil trace gas fluxes along a soil moisture gradient (wetland, transitional and upland habitats) in close proximity to one-another. Habitat type is clearly the main driver of both CO2 and CH4 fluxes, but appears to more strongly affect the latter. To tease out the subset of abiotic and biotic factors that locally determines trace gas fluxes remains challenging.
We found large differences in the amount of CH4 emitted from individual trees and from different species. Our data support other studies that report species differences in wetland (Pangala et al. 2015) and upland forests (Pitz and Megonigal 2017; Wang et al. 2016). Aspects of wood anatomy and tree physiology such as wood vessel structure, wood specific density, lenticel density, transpiration rates and sap flow rates may contribute to species level differences. While it has been shown that wood specific density and lenticel density affect stem CH4 fluxes in wetland trees (Pangala et al. 2014, 2013), it is unclear how these other factors contribute to interspecific CH4 flux differences in upland trees. Among the four tree species we examined in the upland ecosystem, Quercus spp. have ring-porous vessel structure while the other three species are diffuse-porous. We did not detect any patterns that distinguish these morphological types. Future studies should incorporate these potential drivers into the experimental design.
The mixed effect model found stem CH4 emissions to be positively related to DBH. This contrasts with Pangala et al. (2013) and Pangala et al. (2015) who found small diameter wetland trees emitted more CH4 per unit area of stem than larger trees. There was little overlap between the DBH range of those studies (7.5–19.8 cm) and ours (16.1–92.6 cm). The mean stem diameter across all habitats in our study was 42.3 cm (Table 1), with only four trees out of 31 having DBH < 20 cm. Differences in root morphology and biomass between small and large trees may explain the contrasting results. Older and bigger trees have larger and deeper root systems and more likely have deep roots that tap into anaerobic soils or groundwater, both being potential sources of CH4. Other potential CH4 sources such as heart-rot and non- structural carbohydrates would result in a positive relationship between tree diameter and fluxes (Covey et al. 2012, 2016). Regardless of the mechanism, the data highlight the importance of tree size especially when scaling up plot-level studies to ecosystem-level estimates (Covey et al. 2016; Pangala et al. 2015). Future studies should address multiple tree size classes on a given species within one study site.
In the temperate region, stem CH4 flux from trees has been shown to be related to temperature-dependent CH4 production processes. In wetland forests, Pangala et al. (2015) reported a correlation between stem CH4 emission and temperature, water-table depth, and CH4 concentration in pore water. They concluded that CH4 flux was driven by CH4 production in water-logged wetland soil. By sampling a nearby spring, Wang et al. (2016) concluded that groundwater was not a source of CH4 in an upland forest; rather, they attributed stem CH4 flux to in situ CH4 production in the heartwood. The trees in the present study were arranged along an upland-to-wetland gradient of groundwater depth. Our results showed a strong negative correlation between stem CH4 flux and the depth to groundwater, and is consistent with a belowground CH4 source hypothesis previously proposed for wetland trees (Pangala et al. 2015). We measured high stem CH4 fluxes in the wetland habitat where soil CH4 production is consistently high and lower but still positive stem CH4 fluxes from upland trees even when adjacent soils are net CH4 consumers. The observation that co-located upland soils act as CH4 sinks while trees act as sources suggests that trees may provide a flux pathway through the woody tissue that bypasses the methanotrophs in the oxic soil layers (Megonigal and Guenther 2008).
Methane transport in plants can be via diffusion or transpiration (Megonigal and Guenther 2008; Pangala et al. 2014). The latter process has a strong diurnal pattern, suggesting that transpiration-driven CH4 fluxes should exhibit a diel cycle. Pangala et al. (2014) reported only a weak positive relationship between emissions and transpiration, and Terazawa et al. (2015) did not observe diurnal patterns. Both studies concluded that diffusion is the major driver of stem CH4 emissions. In an upland forest stand near our transect, Pitz and Megonigal (2017) demonstrated a strong diurnal pattern in stem CH4 flux for American beech and tulip poplar, with a two-fold diurnal difference for the latter. This indicates that transpiration may play a significant role in stem emissions. Clearly, high frequency measurements for longer periods are necessary to reveal the relative importance of transpiration and diffusion in different habitats.
Soil moisture is considered a major driver of belowground biogeochemical processes, and is thus often reported as an abiotic variable. Usually soil moisture measurements reflect only the surface conditions. In a wetland, such data reflects conditions throughout the soil profile, but in the upland, surface conditions do not represent the steeper, more variable, vertical soil moisture gradient. Soil moisture was not a significant variable in our model of stem CH4 fluxes (Table 2), which is consistent with root uptake from groundwater and transpiration as a mechanism for gas transport.
Soil CO2 efflux decreased from upland to wetland, while stem respiration showed the opposite pattern (Fig. 3). Biological and physical processes may simultaneously explain this result. Lower soil CO2 emissions in the wetland coincided with near-continuous flooding, which can be explained by several mechanisms. Flooding can decrease CO2 respiration of stems by suppressing overall tree growth, and from roots by lowering both growth and root:shoot ratio (Megonigal et al. 1997). Similarly, flooding decreases microbial respiration both by suppressing overall microbial respiration and by lowering the CO2:CH4 ratio (Megonigal et al. 2004; Yu et al. 2008). Because stem CO2 emissions were similar in the wetland and transitional zones, the most likely biological explanation for lower soil CO2 emissions in the wetland is a decrease in microbial respiration as opposed to lower plant respiration. A purely physical explanation is that high soil moisture in the transitional and wetland habitats reduced gas diffusion rates through soil pore spaces (Davidson et al. 1998; Moyano et al. 2013; Suseela et al. 2012) favoring diffusion through woody tissues. These explanations are not mutually exclusive and may both have a role in explaining the cross-habitat patterns of soil and stem CO2 emissions.
Tree CH4 emission rates have now been measured in a variety of forest ecosystems (Table 3). Although the number of studies are few, it is noteworthy that during the growing season, the highest mean emissions from upland [190 µg m−2 h−1 (Covey et al. 2012)] and wetland stems (567.9 µg m−2 h−1 in the present study) are within the same order of magnitude. The observation that upland trees have the potential to emit CH4 at rates comparable to wetland trees suggests the need for detailed mechanistic studies of CH4 sources and sinks across all forest ecosystems.
Soil trace gas fluxes generally show high spatio-temporal variability and are often cited as examples for biogeochemical hotspots and hot moments (Hagedorn and Bellamy 2011; McClain et al. 2003; Ullah and Moore 2011). In such situations a single measurement can be several orders of magnitude higher than the mean (Kuzyakov and Blagodatskaya 2015). Our data also show high variability, especially in the wetland where soils and stems consistently emit CH4. In this habitat, the coefficient of variation for soil flux (1.94) is twice as much as stem flux (0.92). Traditional soil trace gas measurement methods capture processes only in a small area, with large variations among individual measurements due to the existence of biogeochemical hot spots in a highly heterogeneous landscape. For a single tree, soil conditions around individual roots are highly heterogeneous, the entire root system spreads over a much larger area, and thus integrates widely varying conditions both vertically and horizontally (Schenk and Jackson 2002). Across all habitats pairwise measurements of stem CH4 fluxes consistently exceeded soil CH4 fluxes on an area basis, suggesting that the base of tree stems are a localized hot spot relative to the soil. In a few instances, we also recorded hot moments with extremely high (an order of magnitude higher than the mean) stem CH4 fluxes (Fig. 2). It is important not to discard such data as outliers, even though data analysis becomes more challenging. Recent advances of continuous field trace gas measurement technology will allow high frequency sampling and thus to better characterize the sources, sinks and drivers of CH4 fluxes in forests.
Despite the heightened interest following Keppler et al. (2006), to date only a handful studies convincingly documented trees as CH4 sources (Table 3). Six of those, including our study, attempted to quantify CH4 flux from live upland trees, and five directly measured stem fluxes in the field. Different studies report their data using incompatible approaches, including different methodologies to scale up data. Studies have been conducted under different climatic conditions, from tropical rainforests to boreal forests. Chambers were closed from 6 min to 6 h for each measurement. The position of chambers above the ground, which has been shown to affect flux rates (Pangala et al. 2013), varied from 15 to 200 cm. One issue that we address here is the variations in stem flux chamber design used in direct field measurements. Some studies (Terazawa et al. 2007, 2015), ours included, employed a chamber that is mounted one side of a tree (i.e., partial-circumference design). The advantages of this design are that it allows measurements on large trees, maximizes the surface to volume ratio, allowing for relatively short measurement times. The design suited our goal of measuring emissions from a wide range of tree sizes and detecting small fluxes in upland forests. However, this type of chamber does not capture radially variation in emissions around the circumference of the tree and is difficult to fit on small trees. Other studies (Gauci et al. 2010; Pangala et al. 2013) have used a chamber that completely encloses the stem (i.e., full-circumference design). This chamber style results in a lower surface to volume ratio and may result in longer measurement times but it is well suited for small diameter stems. Recently Siegenthaler et al. (2016) developed a full-circumference design using low permeability, flexible material that significantly improves surface to volume ratios. This chamber style may ideally address some field, habitat, and tree diameter constraints. A systematic comparison between chamber styles and recommendations of standardized chamber designs and protocols will allow for interpretation and synthesis of stem flux measurements.
Conclusion
Forests cover about 30% of the global land surface (FAO 2016) and are important sinks of atmospheric carbon (IPCC 2013). The present and other studies indicate that upland trees emit CH4 and thus have to be incorporated into forest carbon cycling models. Our data support a below ground CH4 source, but this pathway probably works simultaneously with other mechanisms, such as CH4 derived from internal microbial sources. Our study highlighted the many biotic and abiotic factors that influence tree mediated CH4 fluxes. Future studies should focus on teasing apart the roles of tree size and species identity (physiology, wood structure, rooting depth), and a multitude of above- and below-ground environmental factors. Using high frequency measurements (Pitz and Megonigal 2017) will help determine drivers on diurnal, seasonal and annual scales, and identify hot moments. Considering evidence that the global warming potential of CH4 is dramatically higher when being consumed from the atmosphere than emitted (sustained global warming potential of uptake = 203 vs emission = 45 over 100 years) (Neubauer and Megonigal 2015), systems such as upland forests where CH4 is simultaneously produced and consumed are particularly important to evaluate from a whole-system perspective.
References
Aydin M et al (2011) Recent decreases in fossil-fuel emissions of ethane and methane derived from firn air. Nature 476:198–201. https://doi.org/10.1038/Nature10352
Baird AJ, Stamp I, Heppell CM, Green SM (2010) CH4 flux from peatlands: a new measurement method. Ecohydrology 3:360–367. https://doi.org/10.1002/eco.109
Brown MJ, Parker GG (1994) Canopy light transmittance in a chronosequence of mixed-species deciduous forests. Can J For Res 24:1694–1703. https://doi.org/10.1139/X94-219
Brush GS, Lenk C, Smith J (1980) The natural forests of Maryland - an explanation of the vegetation map of Maryland. Ecol Monogr 50:77. https://doi.org/10.2307/2937247
Bushong FW (1907) Composition of gas from cottonwood trees. Trans Kans Acad Sci 21:53. https://doi.org/10.2307/3624516
Conrad R (2007) Microbial ecology of methanogens and methanotrophs. Adv Agron 96:1–63. https://doi.org/10.1016/S0065-2113(07)96005-8
Covey KR, Wood SA, Warren RJ, Lee X, Bradford MA (2012) Elevated methane concentrations in trees of an upland forest. Geophys Res Lett. https://doi.org/10.1029/2012gl052361
Covey KR et al (2016) Greenhouse trace gases in deadwood. Biogeochemistry 130:215–226. https://doi.org/10.1007/s10533-016-0253-1
Dacey JWH, Klug MJ (1979) Methane efflux from lake-sediments through water lilies. Science 203:1253–1255. https://doi.org/10.1126/Science.203.4386.1253
Davidson EA, Belk E, Boone RD (1998) Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biol 4:217–227. https://doi.org/10.1046/j.1365-2486.1998.00128.x
FAO (2016) State of the world’s forests 2016. FAO, Rome
Fung I, Matthews E, Lerner J (1987) Atmospheric methane response to biogenic sources - results from a 3-D atmospheric tracer model. Abstr Pap Am Chem Soc 193:6-Geoc
Garnet KN, Megonigal JP, Litchfield C, Taylor GE (2005) Physiological control of leaf methane emission from wetland plants. Aquat Bot 81:141–155. https://doi.org/10.1016/j.aquabot.2004.10.003
Gauci V, Gowing DJG, Hornibrook ERC, Davis JM, Dise NB (2010) Woody stem methane emission in mature wetland alder trees. Atmos Environ 44:2157–2160. https://doi.org/10.1016/J.Atmosenv.2010.02.034
Hagedorn F, Bellamy P (2011) Hot spots and hot moments for greenhouse gas emissions from soils. Soil carbon in sensitive European ecosystems, vol 13. From science to land management. Wiley, hoboken, p 32. https://doi.org/10.1002/9781119970255
Higman D (1968) An ecologically annotated checklist of the vascular flora at the Chesapeake Bay Center for field biology, with keys. Smithsonian Institution, Washington DC
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363. https://doi.org/10.1002/bimj.200810425
IPCC (2013) Climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge and New York, NY. https://doi.org/10.1017/CBO9781107415324
Keppler F, Hamilton JTG, Brass M, Rockmann T (2006) Methane emissions from terrestrial plants under aerobic conditions. Nature 439:187–191. https://doi.org/10.1038/Nature04420
Keppler F, Hamilton JTG, McRoberts WC, Vigano I, Brass M, Rockmann T (2008) Methoxyl groups of plant pectin as a precursor of atmospheric methane: evidence from deuterium labelling studies. New Phytol 178:808–814. https://doi.org/10.1111/j.1469-8137.2008.02411.x
Kirschke S et al (2013) Three decades of global methane sources and sinks. Nat Geosci 6:813–823. https://doi.org/10.1038/Ngeo1955
Kuzyakov Y, Blagodatskaya E (2015) Microbial hotspots and hot moments in soil: concept & review. Soil Biol Biochem 83:184–199. https://doi.org/10.1016/j.soilbio.2015.01.025
Machacova K et al (2016) Pinus sylvestris as a missing source of nitrous oxide and methane in boreal forest. Sci Rep 6:23410. https://doi.org/10.1038/srep23410. http://www.nature.com/articles/srep23410#supplementary-information
Maier M, Machacova K, Lang F, Svobodova K, Urban O (2017) Combining soil and tree-stem flux measurements and soil gas profiles to understand CH4 pathways in Fagus sylvatica forests. J Plant Nutr Soil Sci. https://doi.org/10.1002/jpln.201600405
McClain ME et al (2003) Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems 6:301–312. https://doi.org/10.1007/S10021-003-0161-9
Megonigal JP, Guenther AB (2008) Methane emissions from upland forest soils and vegetation. Tree Physiol 28:491–498
Megonigal JP, Conner WH, Kroeger S, Sharitz RR (1997) Aboveground production in Southeastern floodplain forests: a test of the subsidy-stress hypothesis. Ecology 78:370–384
Megonigal JP, Hines ME, Visscher PT (2004) Anaerobic metabolism: linkages to trace gases and aerobic processes. In: Schlesinger WH (ed) Biogeochemistry, vol 8, 1st edn. Treatise on geochemistry. Elsevier, Oxford, pp 317–424
Moyano FE, Manzoni S, Chenu C (2013) Responses of soil heterotrophic respiration to moisture availability: an exploration of processes and models. Soil Biol Biochem 59:72–85. https://doi.org/10.1016/j.soilbio.2013.01.002
Natural-Resources-Conservation-Service (2016) Web soil survey. http://websoilsurvey.nrcs.usda.gov/. Accessed 15 Jan 2017
Neubauer SC, Megonigal JP (2015) Moving beyond global warming potentials to quantify the climatic role of ecosystems. Ecosystems 18:1000–1013. https://doi.org/10.1007/s10021-015-9879-4
Nisbet EG et al (2016) Rising atmospheric methane: 2007–2014 growth and isotopic shift. Glob Biogeochem Cycles 30:1356–1370. https://doi.org/10.1002/2016gb005406
Pangala SR, Moore S, Hornibrook ERC, Gauci V (2013) Trees are major conduits for methane egress from tropical forested wetlands. New Phytol 197:524–531. https://doi.org/10.1111/Nph.12031
Pangala SR, Gowing DJ, Hornibrook ERC, Gauci V (2014) Controls on methane emissions from Alnus glutinosa saplings. New Phytol 201:887–896. https://doi.org/10.1111/Nph.12561
Pangala SR, Hornibrook ERC, Gowing DJ, Gauci V (2015) The contribution of trees to ecosystem methane emissions in a temperate forested wetland. Glob Change Biol. https://doi.org/10.1111/gcb.12891
Parker GG, Tibbs DJ (2004) Structural phenology of the leaf community in the canopy of a Liriodendron tulipifera L. forest in Maryland, USA. For Sci 50:387–397
Pitz S, Megonigal JP (2017) Temperate forest methane sink diminished by tree emissions. New Phytol. https://doi.org/10.1111/nph.14559
Pulliam WM (1992) Methane emissions from cypress knees in a southeastern floodplain swamp. Oecologia 91:126–128
R Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Rusch H, Rennenberg H (1998) Black alder (Alnus glutinosa (L.) Gaertn.) trees mediate methane and nitrous oxide emission from the soil to the atmosphere. Plant Soil 201:1–7. https://doi.org/10.1023/A:1004331521059
Ryan MG (1990) Growth and maintenance respiration in stems of Pinus Contorta and Picea Engelmannii. Can J For Res 20:48–57. https://doi.org/10.1139/X90-008
Saunois M et al (2016) The global methane budget 2000–2012. Earth Syst Sci Data 8:697–751. https://doi.org/10.5194/essd-8-697-2016
Schenk HJ, Jackson RB (2002) Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. J Ecol 90:480–494. https://doi.org/10.1046/J.1365-2745.2002.00682.X
Siegenthaler A, Welch B, Pangala SR, Peacock M, Gauci V (2016) Technical note: semi-rigid chambers for methane gas flux measurements on tree stems. Biogeosciences 13:1197–1207. https://doi.org/10.5194/bg-13-1197-2016
Suseela V, Conant RT, Wallenstein MD, Dukes JS (2012) Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Glob Change Biol 18:336–348. https://doi.org/10.1111/j.1365-2486.2011.02516.x
Terazawa K, Ishizuka S, Sakatac T, Yamada K, Takahashi M (2007) Methane emissions from stems of Fraxinus mandshurica var. japonica trees in a floodplain forest. Soil Biol Biochem 39:2689–2692. https://doi.org/10.1016/J.Soilbio.2007.05.013
Terazawa K, Yamada K, Ohno Y, Sakata T, Ishizuka S (2015) Spatial and temporal variability in methane emissions from tree stems of Fraxinus mandshurica in a cool-temperate floodplain forest. Biogeochemistry 123:349–362. https://doi.org/10.1007/s10533-015-0070-y
Tiner RW, Burke DG (1995) Wetlands of maryland. US Fish and Wildlife Service, Ecological Services, Region 5, Hadley, and Maryland Department of Natural Resources, Annapolis
Ullah S, Moore TR (2011) Biogeochemical controls on methane, nitrous oxide, and carbon dioxide fluxes from deciduous forest soils in eastern Canada. J Geophys Res Biogeosci. https://doi.org/10.1029/2010jg001525
Vann CD, Megonigal JP (2003) Elevated CO2 and water depth regulation of methane emissions: comparison of woody and non-woody wetland plant species. Biogeochemistry 63:117–134. https://doi.org/10.1023/A:1023397032331
Vigano I, van Weelden H, Holzinger R, Keppler F, McLeod A, Rockmann T (2008) Effect of UV radiation and temperature on the emission of methane from plant biomass and structural components. Biogeosciences 5:937–947
Wang ZP et al (2016) Methane emissions from the trunks of living trees on upland soils. New Phytol 211:429–439. https://doi.org/10.1111/nph.13909
Warner DL, Villarreal S, McWilliams K, Inamdar S, Vargas R (2017) Carbon dioxide and methane fluxes from tree stems, coarse woody debris, and soils in an upland temperate forest. Ecosystems. https://doi.org/10.1007/s10021-016-0106-8
Yesilonis I, Szlavecz K, Pouyat R, Whigham D, Xia L (2016) Historical land use and stand age effects on forest soil properties in the Mid-Atlantic US. For Ecol Manag 370:83–92. https://doi.org/10.1016/j.foreco.2016.03.046
Yu KW, Faulkner SP, Baldwin MJ (2008) Effect of hydrological conditions on nitrous oxide, methane, and carbon dioxide dynamics in a bottomland hardwood forest and its implication for soil carbon sequestration. Glob Change Biol 14:798–812. https://doi.org/10.1111/j.1365-2486.2008.01545.x
Zeikus JG, Ward JC (1974) Methane formation in living trees: a microbial origin. Science 184:1181–1183. https://doi.org/10.1126/science.184.4142.1181
Acknowledgements
This study was supported by Grants from the Department of Energy (DE-SC0008165), the National Science Foundation (ACI-1244820, EAGER NEON EF-1550795, ERC-MIRTHE EEC-0540832), and the Department of Earth and Planetary Sciences Summer Field funds. We thank Jess Parker, Anand Gnanadesikan, Lisa Schile, and members of the GCREW Lab for their advice and useful suggestions throughout the study. We are thankful for all the help that Mike Bernard, Jacob Rode, Adam Dec, Andy Sample and Kyle King provided in the field. Anand Gnanadesikan and two anonymous reviewers provided helpful comment on earlier versions of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Jan Mulder.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pitz, S.L., Megonigal, J.P., Chang, CH. et al. Methane fluxes from tree stems and soils along a habitat gradient. Biogeochemistry 137, 307–320 (2018). https://doi.org/10.1007/s10533-017-0400-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-017-0400-3