
Abstract
Carbon accumulation and storage is a defining characteristic of peatland ecosystems. Decomposition of peat releases dissolved organic carbon (DOC) to receiving waters and can be an important fraction of the peatland carbon budget, along with being an important modifier of downstream water quality. Changes in temperature and hydrological processes under future climate scenarios are expected to impact decomposition processes in peatlands with unclear ramifications for both the quantity and the quality of the DOC released. We experimentally examined the individual and interactive effects of increased temperature, elevated atmospheric carbon dioxide concentration, and lower water table position on peatland decomposition and the quantity and quality of porewater DOC in intact, replicated peat monoliths in a full factorial design. Decomposition rates and porewater DOC concentrations significantly increased under elevated temperature conditions; however, the quality of this carbon was variable, showing signs of both increased lability and recalcitrance. Lowered water table treatments also increased decomposition rates, although the high water conditions prompted greater porewater DOC concentrations and lability. It is expected that elevated decomposition rates under future climate scenarios will alter porewater DOC quantity in peatlands; however, we suggest that contributions from the aboveground system are needed to fully understand changes in DOC quality and subsequent ecosystem dynamics.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
For peatlands in northern regions, a cool, wet climate, acidic, nutrient-poor soils, low ecosystem productivity, and low decomposition rates support high carbon storage (Yavitt et al. 1997; Rydin and Jeglum 2013). In fact, northern peatlands are the most efficient terrestrial ecosystem at storing carbon per unit area in the world (Parish et al. 2008), currently accounting for approximately 30 % of the Earth’s terrestrial carbon (Gorham 1991). Changes to the carbon balance of northern peatlands is likely under global climate change, where elevated temperature and changes to the water balance drive enhanced decomposition, increasing carbon loss as CO2, CH4 and dissolved organic carbon (DOC) (Roulet et al. 1992; Rouse et al. 1997)—potentially driving these systems toward net carbon sources, rather than sinks. Although considerable effort has gone into understanding the processes controlling the exchange of CO2 and CH4 between peatlands and the atmosphere, slightly less effort has been committed to exploring the role of porewater DOC production and export in the carbon balance of peatlands, particularly in the context of future climates. Loss of DOC, while dependent on porewater DOC concentration and hydrologic conditions (e.g. water runoff), may comprise up to 20 % of a peatland carbon balance (Koehler et al. 2011). As both decomposition processes releasing DOC into porewater and changes in hydrologic conditions within peatlands are expected to change under future climate scenarios, understanding how global change factors affects DOC production will be a critical part of understanding peatland carbon budgets.
Dissolved organic carbon compounds in peatlands are derived primarily from the decomposition of peat and plant tissue, but also through contributions from plant root exudates and microbial activity, such as extracellular enzymes (Kalbitz et al. 2000). The production of DOC through all these processes is dependent on environmental variables such as temperature and water table (Moore and Dalva 2001; Clark et al. 2009). Elevated temperature, in particular, has been shown to be an important driver of increased porewater DOC concentrations through increased aboveground litter production and leaching, as well as increased microbial extracellular enzymes production and activity belowground (Fenner et al. 2007; Kane et al. 2014). Warming, as well as increased atmospheric CO2, also enhances the production of root exudates and increases belowground plant root biomass, contributing to increased porewater DOC concentrations (Fenner et al. 2007). Peatland water table elevation is another important control on DOC production (Worrall et al. 2006; Clark et al. 2009) where lower water tables and more aerobic conditions in surface peats are linked to increased DOC production (Worrall et al. 2006) through enhanced decomposition processes. Increased aerobic conditions can also promote vegetation growth, resulting in increased litter and root exudates that add to the DOC pool (Strack et al. 2008).
The chemical composition of carbon compounds in peatland DOC ranges from simple, aliphatic carbon chains to complex, aromatic ring structures (Kalbitz et al. 2003), with simple compounds often derived from microbial material, litter leachate or root exudates, while complex compounds are predominately produced via decomposition processes. The relative proportions of these ‘labile’ versus ‘recalcitrant’ DOC compounds (sometimes referred to high and low DOC quality, respectively) is a critical component of the net carbon balance of northern peatland ecosystems under climate change. For instance, the quality of DOC determines whether DOC acts as a terminal electron acceptor for biogeochemical reactions, as a substrate for fermentative processes such as methanogenesis, or as a source of energy to both in situ and ex situ microbial metabolism (Marschner and Kalbitz 2003; Tfaily et al. 2013). Functionally, quality defines the role DOC has in biogeochemical cycles, affecting decomposition process pathways and rates, and determining the available basal energy in an ecosystem—but characterisation of DOC quality has proven to be challenging.
DOC quality has been explored using techniques such as column-based fractionation or gas chromatography-mass spectrometry (GC-MS) that elucidate the source of DOC (e.g. plant or microbial) and the stage of decomposition. Generally, these techniques have shown that peatland DOC is dominated by high molecular weight carbon compounds (Strack et al. 2011; Kiikkilä et al. 2014) rich in lignin, phenols, and polysaccharides (Kracht and Gleixner 2000; Biester et al. 2014; Treat et al. 2014). However, these methods are limited by challenging preparation, instrumentation, and interpretation of results. An alternative is specific UV absorbance (SUVA) and fluorescence spectroscopy. Specific UV absorbance at 254 nm (SUVA254) is a proxy for DOC aromaticity and has proven to be a widely adopted and straightforward measure (Weishaar et al. 2003). Fluorescence spectroscopy, conversely, uses excitation-emission spectra to make sophisticated but indirect measures of DOC quality (Cory and McKnight 2005). Ratios of specific excitation-emission values and other matrix derivatives have been compellingly linked to the source of DOC, freshness of the compounds, and degree of humification (Fellman et al. 2010); information that can indicate significant changes in carbon quality in peatland ecosystems.
When applied to peatland systems studies have linked a decrease in SUVA254 values with inputs of more labile, lower molecular weight DOC from root exudates or microbial sources under elevated temperatures due to increasing photosynthesis, metabolic rates, and/or microbial turnover (Kane et al. 2014; Lou et al. 2014). More aerobic peat conditions, conversely, are associated with higher SUVA254 indicating an increase in the aromatic fraction of DOC (Höll et al. 2009; Kiikkilä et al. 2014). In the case of emission-excitation matrices (EEMs) and their derivatives, peatland DOC has consistently and perhaps unsurprisingly been described as having a highly humified and recalcitrant character of terrestrial origin (Wickland et al. 2007; Olefeldt et al. 2014). However, to the best of our knowledge, only a limited number of studies have applied either of these techniques to specific investigations of climate change factors on peatland DOC quality (but see Kane et al. 2014; Lou et al. 2014), and fewer still have considered the interacting effects of such factors.
Here we determine the independent and interactive effects of temperature, atmospheric CO2 concentrations, and water table position on peat decomposition rates as well as DOC quantity and quality using intact, vegetated peat mesocosms and a replicated, factorial experimental design. As our treatment mesocosms transition from a Sphagnum moss to a graminoid dominated community during our study (see Dieleman et al. 2015) increased vegetative inputs from litter and root exudates were possible. Accordingly, we wanted to determine if interacting climate change factors affecting intact mesocosms enhance decomposition rates to increase DOC concentrations and recalcitrance, or alternatively if these conditions increase labile DOC production. Based on the current literature we predicted that rising temperatures, elevated atmospheric CO2, and lowered water table conditions would interact synergistically to increase DOC production. We also anticipated that the strong influence of lowered water table conditions and rising temperatures on decomposition rates would render said DOC generally more recalcitrant than that produced under ambient conditions. With such information, we can better define the trajectory of peatland DOC concentrations as well as the changes in its composition that may affect its biogeochemical properties and ultimately peatland ecosystem stability.
Methods
Mesocosms and experimental treatments
Intact, vegetated cylindrical peat monoliths (30 cm diameter × 35 cm depth) were collected in August 2012 from a Sphagnum-dominated nutrient-poor fen near White River, Ontario, Canada (48°21′N, 85°21′W) to create the 84 peatland mesocosms used in this experiment. Dieleman et al. (2015) describe the specifics of the source site, the experimental design, and the vegetative community, including the observed shift in plant composition. Briefly, the mesocosms (5 gallons, 30 cm diameter, 35 cm tall) were fitted with a 2 cm ABS barbed fitting port attached to a 16 mm diameter, 25 cm long perforated PVC pipe drain to collect DOC samples and monitor water table levels. In the University of Western Ontario’s Biotron Environmental Climate Change Research Centre, seven replicates of each lowered (25 cm below the peat surface) and elevated (5 cm below the peat surface) water table treatment mesocosms were randomly placed in six environmentally controlled, state-of-the-art greenhouses at three temperature (ambient, ambient +4 °C, ambient +8 °C) and two atmospheric CO2 conditions (430, 750 ppm). The greenhouses were divided into two replicates of the temperature treatment, one set paired with ambient atmospheric CO2 conditions and the second paired with elevated atmospheric CO2 conditions. The specific temperature and CO2 treatments were selected to bracket IPCC (2013) predictions for higher latitude northern ecosystems in the next 50–100 years, while the water table treatments reflect the predictions of Roulet et al. (1992) for northern peatlands.
Temperature, atmospheric CO2 concentrations, and relative humidity were regulated using an automated environmental control system (ARGUS Control Systems Ltd., White Rock, BC, Canada), while water table conditions were manually maintained. Relative humidity was regulated to no less than 60 %. The temperature conditions were maintained under two regimes corresponding to the growing and dormant seasons. During the growing season (May 1–October 31), ambient temperatures matched the hourly 5 year average daily temperatures for the region. During the dormant period (November 1–April 31) the experimental greenhouse temperatures were infrastructurally-limited to temperatures >10 °C so ambient temperatures were maintained at the average growing season temperature for the region, approximately 11.5 °C, while the other experimental treatments were maintained at the same +4 and +8 °C offsets. The two different water table treatments were held constant to avoid any confounding influence of temperature and/or changes in aboveground vegetation via simulated rain events biweekly at the individual mesocosm surface, and supplemental water additions below the moss layer as needed. The individual mesocosms were also randomly relocated within their individual greenhouse on a monthly basis, and transferred to a different greenhouse with replicate conditions every 6 months to address both microclimate and greenhouse effects (Hurlbert 1984).
Decomposition
Decomposition rates were quantified via mass loss of Sphagnum litter collected from destructively sampled mesocosms prior to the experiment. Three replicate decomposition bags (10 cm × 7 cm with 1 mm mesh) were inserted 10 cm below the peat surface in four of the seven mesocosms under each experimental combination (48 treatments; 144 litterbags in total) at the beginning of the experiment in December 2012. Each litterbag contained ~30 g wet weight homogenized peat (~3 g dry weight); initial absolute dry weights were determined for every litterbag. One litterbag from each mesocosm was retrieved every 4 months (4, 8, 12 months) over the course of the experiment. Collected bags were oven-dried for 72 h at 60 °C, reweighed and mass loss (%) calculated relative to the initial dry mass.
Dissolved organic carbon sampling and analyses
Porewater samples were collected monthly from the ports at the bottom of the mesocosms for all 84 mesocosms (seven replicate mesocosms for each treatment) from December 2012 to December 2013. These monthly samples were assessed for changes in carbon quantity, measured as DOC in mg L−1, and carbon quality, measured as specific ultraviolet absorbance at a wavelength of 254 nm in L mg C−1 m−1. To further quantify changes in carbon quality additional samples for excitation-emission matrix fluorescence spectroscopy analysis were collected at the experiment end in December 2013. All porewater samples were collected using pre-cleaned tubing and a peristaltic pump to collect 200 mL samples that were stored (maximum of 48 h) in 500 mL Nalgene™ bottles in the dark at 4 °C. All samples were then filtered using ashed glass filters (pore size 0.5 µm) and split into 50 mL HDPE bottles for DOC samples, and 20 mL glass vials for SUVA254 samples as well as for EEMs samples when applicable. All samples were promptly stored in the dark at 4 °C until processed.
Dissolved organic carbon concentration was determined using an iTOC Aurora 1030 (OI Analytical, College Station, TX, USA) via the persulfate wet oxidation method. The measurement of SUVA254 used methods modified from Weishaar et al. (2003). Absorbance was measured at λ = 254 nm using 200 µL sample aliquots in UV transparent 96-well plates read on a Spectramax® M2 spectrophotometer in cm−1. Absorbance values were then converted to m−1 and divided by the DOC concentration of the sample to determine the SUVA254 value. All absorbance values were corrected for potential Fe interference from low Fe concentrations associated with added simulated rainwater. Fe absorbance at λ = 254 nm were calculated based on Weishaar et al. (2003) published extinction coefficient of 0.08 and maximum Fe concentrations (0.114–1.058 mg L−1) in December 2013. Therefore, the SUVA254 may be considered conservative as in some cases they may be overcorrected for Fe interference.
Fluorescence excitation-emission matrices for DOC samples from December 2013 were used to calculate informative optical indices that are indicative of changes in carbon quality; here we focus on three common indices: the fluorescence index (FI), the humification index (HIXEM) and the ‘freshness’ index (BIX) (Fellman et al. 2010). Fluorescence excitation and emission was measured on a Horiba Aqualog® spectrofluorometer with a xenon lamp in the Watershed Hydrology Group Laboratory at McMaster University. The excitation wavelengths ranged from 240 to 600 nm with 10 nm increments and the emission wavelengths ranged from 210 to 620 nm with 3.27 nm increments, all with an integration time of 0.5 s. All samples were diluted 1:4 with ultra-pure deionised water to minimize the attenuation of light by the sample itself (inner filtering effects). To address changes in lamp intensity due to natural decay all samples were normalized using the area under the water Raman peak at an excitation of 350 nm (Lawaetz and Stedmon 2009). The produced EEMs were then used to calculate the aforementioned optical indices using R Software (R Core Team 2012) and the most recent mathematical methods outlined in Fellman et al. (2010).
Statistical analysis
Three-way repeated measures Anova (RM-Anova) with Tukey post hoc tests where necessary were used to quantify the changes in decomposition rates, porewater DOC concentration, and porewater SUVA254 values under treatment conditions (Statistica 7, StatSoft. Inc. 2004). A multivariate Anova (Manova) was utilized to test for experimental effects on the florescence index, the HIXEM, and the BIX (Statistica 7, StatSoft. Inc. 2004) at the end of the experiment. A Pearson correlation test was also conducted to determine the relationship between endpoint SUVA254 values and FI, HIXEM, and BIX.
Results
Decomposition
Sphagnum litter decomposition (expressed as percent mass loss relative to initial mass) was apparent across all experimental treatments over time (Fig. 1). The main driver of decomposition was water table, with significantly greater mass loss under the lowered water table treatment across all conditions (water table: F 1,36 = 118.68, P < 0.001). The rate of mass loss (i.e. the time × water table interaction) was also greater under a lowered water table (F 2,72 = 19.55, P < 0.001), but no other interactive effects were noted. Under both water table treatments, mass loss was positively and significantly related to temperature (temperature: F 2,36 = 25.56, P < 0.001) with mass loss approaching 40 % in some replicates under +8 °C and lowered water table conditions. Under both water table treatments the difference between +8 °C and the other temperature treatments was greater than that between ambient and +4 °C (Fig. 1). There was also a significant, but weaker relationship between percent mass loss and atmospheric CO2 treatments, with greater decomposition under ambient atmospheric CO2 conditions (F 1,36 = 4.77, P = 0.036) (ambient CO2 = 17.6 ± 1.19 %, elevated CO2 = 15.6 ± 1.06 %).

The percent mass loss of homogenized Sphagnum litter (mean ± SE) indicating the decomposition rates of experimental mesocosms under temperature and water table treatments from December 2012 to December 2013. Low water table conditions (a) indicate the water table was approximately 25 cm below the peat surface, while high water table conditions (b) dictate that the water table was approximately 5 cm from the peat surface
Dissolved organic carbon concentration
Despite placing the mesocosms in a 4 month acclimation period under ambient conditions from the time of field sampling to the initiation of the experiment (August 2012–December 2012), a steady decline in DOC concentrations (starting at quite elevated concentrations >80 mg L−1) from December 2012 to February 2013 followed by a stabilization suggests that the initial 3 months of water chemistry data were more strongly influenced by the field sampling and disturbance than experimental treatments. However, overall, total DOC concentrations were positively and significantly higher under the +8 °C treatment (temperature: F 2,61 = 39.600, P < 0.001) and continued to rise over time (temperature × time: F 22,671 = 4.578, P < 0.001), while mean concentrations under ambient and the +4 °C temperature treatments were not significantly different over the course of the experiment (Fig. 2). The pattern of DOC concentrations over time differed between the two water table treatments at +8 °C (water table × time: F 22,671 = 2.892, P < 0.001). Under low water table conditions DOC concentrations show an increase over May–July, falling in August–September then gently rising again toward the end of the experiment. The pattern under high water tables was quite different with a steady increase over the experiment, reaching the highest concentrations of any of the experimental treatments (discounting the recovery period) in December (Fig. 2). DOC concentrations were also significantly influenced by the interactive effects of elevated atmospheric CO2 and increased temperature conditions over time (CO2 × temperature × time: F22,671 = 2.118, P = 0.002), where DOC values continually increased throughout the experiment under the +8 °C/elevated atmospheric CO2 conditions (Table 1). The DOC concentration under the ambient atmospheric CO2 treatment, conversely, remained relatively static from April to October, only rising slightly near the experiment’s end and only under the +8 °C treatment.

The interactive effects of temperature and low water table (a) and high water table (b) conditions on dissolved organic carbon concentrations (mean ± SE) in northern peatland mesocosms over 1 year. Elevated values were recorded for the first 4 months due to mesocosm collection disturbance; however, a full recovery is apparent by April 2013
Specific UV absorbance
Discounting the first three sampling times as artefacts of the acclimation period, the low water table conditions across all experimental treatments had consistently higher SUVA254 values than under high water tables (water table: F 1,57 = 80.790, P < 0.001;) indicating greater DOC aromaticity. Under both water table treatments there was an early summer peak in SUVA254 in May under ambient and the +4 °C temperature treatments that were not observed under +8 °C. After this point in time, the two water table treatments behaved quite differently, with SUVA254 values being relatively stable under low water tables, while steadily declining under high water tables through to the end of the experiment (water table × time: F 11,627 = 7.790, P < 0.001) (Fig. 3). The lowest values for SUVA254 were recorded for the +8 °C treatment under high water tables. Indeed, overall SUVA254 was significantly lower under the higher temperature treatments (temperature: F 2,57 = 10.420, P < 0.001) with the post hoc test indicating SUVA254 values from the +8 °C treatment were significantly lower than SUVA254 values from both ambient and +4 °C treatments—a trend that intensified over time (temperature × time: F 22,627 = 2.180, P = 0.002). SUVA254 values also significantly decreased under elevated atmospheric CO2 conditions (SUVA254 = 4.124 ± 0.036 L mg C−1 m−1) compared to ambient CO2 conditions (SUVA254 = 4.253 ± 0.037 L mg C−1 m−1) (CO2: F 1,57 = 6.220, P = 0.016), with the greatest decreases occurring steadily under the +8 °C treatment from April to December (CO2 × time: F 22,627 = 1.860, P = 0.010) (Table 1). Finally, we found that endpoint SUVA254 values were significantly correlated with both the FI (R = −0.61, P < 0.001) and the HIXEM (R = 0.56, P < 0.001).

The trends in mesocosm porewater carbon quality as measured by specific ultraviolet absorbance at the wavelength 254 nm (SUVA254) (mean ± SE) over 1 year in response to elevated temperature conditions in conjunction with low water table (a) and high water table treatments (b)
EEMs fluorescence indices
Carbon quality, as inferred from EEMs fluorescence measurements, was influenced by both water table and temperature treatments (Table 2). Specifically, both the florescence index and the HIXEM were dominantly determined by the water table (FI: F 1,81 = 184.100, P < 0.001; HIXEM: F 1,81 = 188.900, P < 0.001), where the lowered water table treatment decreased FI values (more aromatic carbon, terrestrially sourced) while raising HIXEM values (more humified carbon). Conversely, raising temperature conditions prompted slightly higher FI (less aromatic carbon, microbially sourced) and HIXEM values (more humified carbon) under +4 and +8 °C treatment conditions when compared to ambient temperature conditions (FI: F 2,81 = 10.300, P < 0.001; HIXEM: F 2,81 = 7.400, P = 0.001). Unlike the other indices, the temperature, water table, and atmospheric CO2 treatments had no clear effects on the BIX.
Discussion
Northern peatlands are a globally relevant carbon sink (Gorham 1991), that will be affected by climate change factors such as elevated temperature, increased atmospheric CO2 concentrations, and lowered water table elevations (Bragazza et al. 2013). Of particular concern are changes in peatland decomposition rates and vegetation inputs, as the recalcitrant character of peatland vegetation paired with the unusually slow rate of decomposition in this ecosystem results in its notable carbon accumulation (Ward et al. 2015). In our experimental system we found that warming conditions increased both decomposition rates and general DOC production resulting in increased porewater DOC concentrations, but the associated quality of that DOC was dependent on other factors, such as water table elevation and atmospheric CO2 concentration.
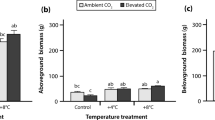
A growing number of studies have demonstrated that higher temperatures and lower water table elevation in northern peatlands increases DOC production and export (Höll et al. 2009; Hribljan et al. 2014). In fact, authors from across Europe and North America are reporting increasing DOC concentrations in systems downstream of peatlands in association with ongoing atmospheric warming as a product of warming-enhanced decomposition (Freeman et al. 2001; Evans et al. 2005; Kane et al. 2014). While our study is in line with these other observed impacts of warming on DOC production, we found that warming conditions synergistically interacted with higher water tables as well as elevated atmospheric CO2 to further increase DOC concentrations in peatland porewaters. As decomposition rates were reduced under these conditions, we suggest that warmer, wetter, and higher atmospheric CO2 conditions stimulate vegetative inputs of DOC, explaining our observed patterns. Specifically, graminoid biomass and abundance (both aboveground and belowground), have been shown to significantly increase under the aforementioned climate change conditions (Weltzin et al. 2000), a trend that was observed in other results reported from this experiment (Dieleman et al. 2016). This increased presence of graminoid biomass can enhance the availability of labile carbon through litter inputs and root exudates (Hobbie 1996; Fenner et al. 2007). In our study this is further supported through a pattern of increased DOC production during the growing season (May–October), mirroring vegetative productivity trends. This does not imply that decomposition processes are not contributing to DOC concentrations under wetter and elevated CO2 conditions, but that other sources, such as graminoids, are enhancing DOC production rates.
Plant inputs, particularly root exudates, are generally a complex mixture of carbohydrates, amino acids, and simple organic acids that tend to be labile, aliphatic compounds (Jones et al. 2009). The products of advanced decomposition, conversely, include condensed lignins and humic acids—all highly recalcitrant carbon compounds (Berg and McClaugherty 2014). The optical indices we employed to monitor changes in carbon quality indicated both the labile and recalcitrant signals increased under warming conditions, providing evidence that the DOC in our study was in fact coming from two sources. The SUVA254 data were highly suggestive of a strong vegetative influence, as the least aromatic values throughout the study were consistently observed under the same wetter and CO2 enriched conditions that are established to support enhanced graminoid productivity (Weltzin et al. 2000; Fenner et al. 2007). These plant inputs may have stimulated microbial growth or directly prompted the slight increase in FI values we observed under warming by supplying microbial-like, labile carbon compounds. Unlike SUVA254 and FI, the HIXEM values indicated an increase in recalcitrant-humified carbon production with warming, reflecting our Sphagnum litter decomposition trends. These findings pair well with other recent warming studies, such as Kane et al. (2014), who also report simultaneous increases in carbon recalcitrance and lability with warming. That said, when interpreting results like ours it is important to note that measures of carbon quality, such as EEMs, can only reveal a subset of carbon molecules that are responsive to the technique. Consequently, relatively less responsive carbon molecules, like some recalcitrant tannins compounds, may constitute a larger portion of the carbon pool than revealed. Nonetheless, our results suggest that warming conditions significantly alter carbon quality; however, the prevailing characteristics of peatland DOC are determined by indirect, temperature-driven changes in both the aboveground plant productivity and belowground microbial activity.
Unlike our warming treatments, the lowered water table treatment consistently resulted in a signature of increased carbon recalcitrance. Various studies, including our own, have demonstrated that decomposition is intensified under the more aerobic conditions lowered water table treatments produce (Bergman et al. 1999; Šantrůčková et al. 2004). These conditions increase the availability of effective electron acceptors, such as O2, increasing decomposition reaction rates, which causes the rapid break down of simple carbons, ultimately leaving mostly recalcitrant compounds in solution (Höll et al. 2009). However, as previously mentioned, high water table treatments enhances graminoid productivity and biomass in peatlands, in general (Weltzin et al. 2000), and in our specific experiment (Dieleman et al. 2016); biomass that is a source of labile carbon (Hobbie 1996; Cornelissen et al. 2007). Such inputs could increase the lability of the high water table treatment, rendering the low water table more recalcitrant by comparison. Our SUVA254 data are suggestive of just that, as its values progressively decline under the high water table conditions that enhance graminoid biomass production, while the low water table values remain relatively constant throughout most of the growing period. That said it is important to note that we did not allow for natural variation in our water table treatments, a factor that could influence how water table conditions drive carbon quality. Yet our reported trends are in accordance with a study where water tables treatments did naturally vary. Specifically, Hribljan et al. (2014) observed a decrease in peatland carbon aromaticity under more saturated conditions due to increased labile carbon production by algae. Taken together we suggest that while future water table drawdown events are likely to enhance the recalcitrant carbon signal in northern peatlands via decomposition, it is increased vegetation inputs that can strongly alter DOC quality.
Significance
We experimentally demonstrated that future climate change conditions may strongly influence dissolved carbon quantity and quality in northern peatlands. Specifically, these environmental conditions interact to enhance decomposition rates and vegetative inputs to increase DOC production and predominately increase lability. For naturally occurring low-turnover ecosystems, like peatlands, this increased organic matter cycling can cause the ecosystem to become ‘leaky’, exporting nutrients and energy to surrounding ecosystems (Wardle et al. 2004). Our findings indicate that warmer conditions in particular can accelerate material cycling, ultimately observed as increased DOC concentrations and lability in peatland porewaters. The consequences of these ecosystem changes are significant. If such DOC rich porewaters are exported, they could significantly alter the environmental conditions and ecosystem structure of receiving downstream aquatic ecosystems (Karlsson et al. 2009; Erlandsson et al. 2010). The enhanced decomposition rates could slow or reverse peat accumulation, compromising the stability of the peatland. Finally, more rapidly cycling peatlands are likely to have a diminished capacity to store carbon, and a greater potential to become a globally significant carbon source.
Abbreviations
- CO2 :
-
Carbon dioxide
- DOC:
-
Dissolved organic carbon
- EEM:
-
Excitation-emission matrix
- FI:
-
Fluorescence index
- BIX:
-
Freshness index
- HIXEM :
-
Humification index
- CH4 :
-
Methane
- SUVA:
-
Specific ultraviolet absorbance
References
Berg B, McClaugherty C (2014) Plant litter decomposition, humus formation, carbon sequestration. Springer-Verlag, Heidelberg
Bergman I, Lundberg P, Nilsson M (1999) Microbial carbon mineralisation in an acid surface peat: effects of environmental factors in laboratory incubations. Soil Biol Biochem 31:1867–1877. doi:10.1016/S0038-0717(99)00117-0
Biester H, Knorr KH, Schellekens J, Basler A, Hermanns Y-M (2014) Comparison of different methods to determine the degree of peat decomposition in peat bogs. Biogeosciences 11:2691–2707. doi:10.5194/bg-11-2691-2014
Bragazza L, Parisod J, Buttler A, Bardgett RD (2013) Biogeochemical plant-soil microbe feedback in response to climate warming in peatlands. Nat Clim Change 3:273–277. doi:10.1038/nclimate1781
Clark JM, Ashley D, Wagner M, Chapman PJ, Lane SN, Evans CD, Heathwaite AL (2009) Increased temperature sensitivity of net DOC production from ombrotrophic peat due to water table draw-down. Glob Change Biol 15:794–807. doi:10.1111/j.1365-2486.2008.01683.x
Cornelissen JHC, Van Bodegom PM, Aerts R et al (2007) Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol Lett 10:619–627. doi:10.1111/j.1461-0248.2007.01051.x
Cory RM, McKnight DM (2005) Fluorescence spectroscopy reveals ubiquitous presence of oxidized and reduced quinones in dissolved organic matter. Environ Sci Technol 39:8142–8149. doi:10.1021/es0506962
Dieleman CM, Branfireun BA, McLaughlin JW, Lindo Z (2015) Climate change drives a shift in peatland ecosystem plant community: implications for ecosystem function and stability. Glob Change Biol 21:388–395. doi:10.1111/gcb.12643
Dieleman CM, Branfireun BA, McLaughlin JW, Lindo Z (2016) Enhanced carbon release under future climate conditions in a peatland mesocosm experiment: the role of phenolic compounds. Plant Soil 400:81–91. doi:10.1007/s11104-015-2713-0
Erlandsson M, Cory N, Köhler S, Bishop K (2010) Direct and indirect effects of increasing dissolved organic carbon levels on pH in lakes recovering from acidification. J Geophys Res 115:G03004. doi:10.1029/2009JG001082
Evans CD, Monteith DT, Cooper DM (2005) Long-term increases in surface water dissolved organic carbon: observations, possible causes and environmental impacts. Environ Pollut 137:55–71. doi:10.1016/j.envpol.2004.12.031
Fellman JB, Hood E, Spencer RGM (2010) Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: a review. Limnol Oceanogr 55:2451–2462. doi:10.4319/lo.2010.55.6.2452
Fenner N, Freeman C, Lock MA, Harmens H, Reynolds B, Sparks T (2007) Interactions between elevated CO2 and warming could amplify DOC exports from peatland catchments. Environ Sci Technol 41:3146–3152. doi:10.1021/es061765v
Freeman C, Evans C, Monteith DT, Reynolds B, Fenner N (2001) Export of organic carbon from peat soils. Nature 412:785. doi:10.1038/35090628
Gorham E (1991) Northern peatlands: role in the carbon cycle and probable responses to climatic warming. Ecol Appl 1:182–195. doi:10.2307/1941811
Hobbie SE (1996) Temperature and plant species control over litter decomposition in Alaskan tundra. Ecol Monogr 66:503–522. doi:10.2307/2963492
Höll BS, Fiedler S, Jungkunst HF, Kalbitz K, Freibauer A, Drösler M, Stahr K (2009) Characteristics of dissolved organic matter following 20 years of peatland restoration. Sci Total Environ 408:78–83. doi:10.1016/j.scitotenv.2009.08.046
Hribljan JA, Kane ES, Pypker TG, Chimner RA (2014) The effect of long-term water table manipulations on dissolved organic carbon dynamics in a poor fen peatland. J Geophys Res Biogeosci 119:577–595. doi:10.1002/2013JG002527
Hurlbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:187–211. doi:10.2307/1942661
IPCC (2013) The physical science basis. In: Stocker TF, Qin D, Plattner G, Tignor MMB, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Climate change 2014: contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Jones DL, Nguyen C, Finlay RD (2009) Carbon flow in the rhizosphere: carbon trading at the soil–root interface. Plant Soil 321:5–33. doi:10.1007/s11104-009-9925-0
Kalbitz K, Solinger S, Park J-H, Michalzik B, Matzner E (2000) Controls on the dynamics of dissolved organic matter in soils: a review. Soil Sci 165:277–304
Kalbitz K, Schmerwitz J, Schwesig D, Matzner E (2003) Biodegradation of soil-derived dissolved organic matter as related to its properties. Geoderma 113:273–291. doi:10.1016/S0016-7061(02)00365-8
Kane ES, Mazzoleni LR, Kratz CJ, Hribljan JA, Johnson CP, Pypker TG, Chimner R (2014) Peat porewater dissolved organic carbon concentration and lability increase with warming: a field temperature manipulation experiment in a poor-fen. Biogeochemistry 119:161–178. doi:10.1007/s10533-014-9955-4
Karlsson J, Byström P, Ask J, Ask P, Persson L, Jansson M (2009) Light limitation of nutrient-poor lake ecosystems. Nature 460:506–509. doi:10.1038/nature08179
Kiikkilä O, Smolander A, Ukonmaanaho L (2014) Properties of dissolved organic matter in peatland: implications for water quality after harvest. Vadose Zone J 13:1–9. doi:10.2136/vzj2013.08.0155
Koehler A-K, Sottocornola M, Kiely G (2011) How strong is the current carbon sequestration of an Atlantic blanket bog? Glob Change Biol 17:309–319. doi:10.1111/j.1365-2486.2010.02180.x
Kracht O, Gleixner G (2000) Isotope analysis of pyrolysis products from Sphagnum peat and dissolved organic matter from bog water. Org Geochem 31:645–654. doi:10.1016/S0146-6380(00)00041-3
Lawaetz AJ, Stedmon CA (2009) Fluorescence intensity calibration using the Raman scatter peak of water. Appl Spectrosc 63:936–940. doi:10.1366/000370209788964548
Lou X-D, Zhai S-Q, Kang B, Hu Y-L, Hu L-L (2014) Rapid response of hydrological loss of DOC to water table drawdown and warming in Zoige peatland: results from a mesocosm experiment. PLoS One 9:e109861. doi:10.1371/journal.pone.0109861
Marschner B, Kalbitz K (2003) Controls of bioavailability and biodegradability of dissolved organic matter in soils. Geoderma 113:211–235. doi:10.1016/S0016-7061(02)00362-2
Moore TR, Dalva M (2001) Some controls on the release of dissolved organic carbon by plant tissues and soil. Soil Sci 166:38–47
Olefeldt D, Persson A, Turetsky MR (2014) Influence of the permafrost boundary on dissolved organic matter characteristics in rivers within the Boreal and Taiga plains of western Canada. Environ Res Lett 9:035005. doi:10.1088/1748-9326/9/3/035005
Parish F, Sirin A, Charman D, Joosten H, Minayeva T, Silvius M, Stringer L (2008) Assessment on peatlands, biodiversity and climate change: main report. Global Environment Centre, Kuala Lumpur
Roulet N, Moore T, Bubier J, Lafleur P (1992) Northern fens: methane flux and climatic change. Tellus B 44:100–105
Rouse WR, Douglas MSV, Hecky RE, Hershey AE, Kling GW, Lesack L, Marsh P, McDonald M, Nicholson BJ, Roulet NT, Smol JP (1997) Effects of climate change on the freshwaters of arctic and subarctic North America. Hydrol Process 11:873–902. doi:10.1002/(SICI)1099-1085(19970630)11:8<873:AID-HYP510>3.0.CO;2-6
Rydin K, Jeglum JK (2013) The biology of peatlands. Oxford University Press, Oxford
Šantrůčková H, Picek T, Tykva R, Šimek M, Pavlů B (2004) Short-term partitioning of 14C-[U]-glucose in the soil microbial pool under varied aeration status. Bio Fert Soils 40:386–392. doi:10.1007/s00374-004-0790-y
Strack M, Waddington JM, Bourbonniere RA, Buckton EL, Shaw K, Whittington P, Price JS (2008) Effect of water table drawdown on peatland dissolved organic carbon export and dynamics. Hydrol Process 22:3373–3385. doi:10.1002/hyp.6931
Strack M, Tóth K, Bourbonniere R, Waddington JM (2011) Dissolved organic carbon production and runoff quality following peatland extraction and restoration. Ecol Eng 37:1998–2008. doi:10.1016/j.ecoleng.2011.08.015
Tfaily MM, Hamdan R, Corbett JE, Chanton JP, Glaser PH, Cooper WT (2013) Investigating dissolved organic matter decomposition in northern peatlands using complimentary analytical techniques. Geochim Cosmochim Acta 112:116–129. doi:10.1016/j.gca.2013.03.002
Treat CC, Wollheim WM, Varner RK, Grandy AS, Talbot J, Frolking S (2014) Temperature and peat type control CO2 and CH4 production in Alaskan permafrost peats. Glob Change Biol 20:2674–2686. doi:10.1111/gcb.12572
Ward SE, Orwin KH, Ostle NJ, Briones MJI, Thomson BC, Griffiths RI, Oakley S, Quirk H, Bardgett RD (2015) Vegetation exerts a greater control on litter decomposition than climate warming in peatlands. Ecology 96:113–123. doi:10.1890/14-0292.1
Wardle DA, Bardgett RD, Klironomos JN, Setälä H, van der Putten WH, Wall DH (2004) Ecological linkages between aboveground and belowground biota. Science 304:1629–1633. doi:10.1126/science.1094875
Weishaar JL, Aiken GR, Bergamaschi BA, Fram MS, Fujii R, Mopper K (2003) Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ Sci Technol 37:4702–4708. doi:10.1021/es030360x
Weltzin JF, Pastor J, Harth C, Bridgham SD, Updegraff K, Chapin CT (2000) Response of bog and fen plant communities to warming and water-table manipulations. Ecology 81:3464–3478. doi:10.1890/0012-9658(2000)081[3464:ROBAFP]2.0.CO;2
Wickland KP, Neff JC, Aiken GR (2007) Dissolved organic carbon in Alaskan boreal forest: sources, chemical characteristics, and biodegradability. Ecosystems 10:1323–1340. doi:10.1007/s10021-007-9101-4
Worrall F, Burt TP, Adamson JK (2006) Trends in drought frequency—the fate of DOC export from British peatlands. Clim Change 76:339–359. doi:10.1007/s10584-006-9069-7
Yavitt JB, Williams CJ, Wieder RK (1997) Production of methane and carbon dioxide in peatland ecosystems across North America: effects of temperature, aeration, and organic chemistry of peat. Geomicrobiol J 14:299–316. doi:10.1080/01490459709378054
Acknowledgments
We are grateful to Dr. C. Dean, Dean of the Western Faculty of Science, for supporting our use of the Biotron, funding from the Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant program (ZL, BB), Canada Research Chairs program (BB), NSERC Strategic Network support to the Canada Network for Aquatic Ecosystem Services (BB), Ontario Ministry of Natural Resources’ Far North and Science and Research Branches (JM) and the Ontario Graduate Scholarship program (CD). We thank the numerous volunteers and work study students for their help, especially C. Chemali, N. Erampamoorthy, R. Doyle, J. Warner, Z. Moore, C. Carvalhal, N. Chattergoon, and L. Albano. We also thank Dr. C. Oswald (Ryerson University) for advice and aid on EEMs techniques and Dr. S. Carey (McMaster University) for access to laboratory instruments. Discussion with Dr. G Thorn and A. Asemaninejad Hassankiadeh were appreciated.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Jennifer Leah Tank.
Rights and permissions
About this article

Cite this article
Dieleman, C.M., Lindo, Z., McLaughlin, J.W. et al. Climate change effects on peatland decomposition and porewater dissolved organic carbon biogeochemistry. Biogeochemistry 128, 385–396 (2016). https://doi.org/10.1007/s10533-016-0214-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-016-0214-8