
Abstract
Downed woody debris is an important component of the forest floor, responsible for long-term storage of nutrients in many ecosystems. In large floodplain forests of the southeastern United States, wood loads are relatively low, suggesting these landscapes may promote rapid decomposition and nutrient turnover rates. The goals of this study were to identify the major factors responsible for wood decomposition and associated nutrient dynamics in an old-growth floodplain forest at Congaree National Park, South Carolina (USA). In situ decomposition and nutrient dynamics were quantified for 700 days (April 2011–March 2013) using wood from a common overstory species, red maple (Acer rubrum L.). During the study period, regional drought conditions allowed red imported fire ant (RIFA, Solenopsis invicta Buren) colonization and infestation of the study wood after 183 days in the field. Carbon (C) content decreased throughout the study, indicating net mineralization was occurring year-round. Invertebrate activity contributed to extremely fast decomposition rates (mean k 0.650 ± 0.02 year−1) and wood turnover times (mean 4.79 ± 0.15 years). In contrast to C, wood nitrogen (N) and phosphorus (P) content increased following RIFA infestation, displaying net nutrient immobilization. The rapid turnover rates measured at CONG indicate that the C storage function of woody debris may be diminished, but availability of mineralized N and P may also have increased the importance of wood stocks to support forest net primary productivity.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Woody debris is an important ecosystem component for accumulation and long-term storage of organic matter and nutrients (Currie et al. 2002; Muller 2003). Inputs and losses of woody debris in forested ecosystems are dependent upon species composition, landscape position, rates of tree disturbance, substrate quality, and localized climate (Harmon et al. 1986; Laiho and Prescott 2004). It has been suggested that woody debris is a minor pool of nutrients during all stages of forest succession when compared to soil and leaf litter (Laiho and Prescott 2004). However, most woody debris studies in North America have been conducted in northern latitude upland landscapes of the United States and Canada (Harmon et al. 1986; Tyrrell and Crow 1994; Laiho and Prescott 2004), and relatively few studies of dead wood have been completed in low gradient floodplains or wetlands (Day 1982; Rice et al. 1997; Wohl et al. 2011). Thus, current standing woody debris stocks and the processes contributing to decomposition and nutrient release from these pools remain unquantified in many large temperate and subtropical zone floodplain forests (Wohl 2013).
Measurement of in situ decomposition is more common for leaf litter than woody debris (Day 1982; Rice et al. 1997). Wood decomposition rates are highly variable between species due to differences in initial litter quality (Harmon et al. 1986; Tyrrell and Crow 1994), but in most cases wood decay is much slower than leaf litter with typical decay constants (k) < 0.100 year−1 in temperate zone climates (Day 1982; Harmon et al. 1986; Mattson et al. 1987; Schilling and Lockaby 2005). Floodplain woody debris is also a critical resource for many macroinvertebrates, acting as a source of food and shelter for various species (Braccia and Batzer 2008). Macroinvertebrates influence the litter decomposition process, typically accounting for small portions (<30 %) of total nutrient mineralization, but physical litter fragmentation may significantly increase mineralization rates by stimulating microbial activity (Hättenschwiler et al. 2005; Bastow et al. 2008). Although the importance of macroinvertebrates have been noted with regard to wood decay, separating their effects from physical and microbial mineralization processes is often difficult in field studies.
Woody debris is an important source of slow release nutrients via mineralization of stored organic carbon (C), nitrogen (N), and phosphorus (P). Nutrient mineralization-immobilization trends in woody debris are largely controlled by microbial processes and local climate conditions that promote seasonal leaching from throughfall organic matter (Day 1983, Harmon et al. 1986). Downed wood nutrient source-sink behavior is dependent on initial nutrient content as well as the level of microbial activity within the woody debris and underlying soil (Harmon et al. 1986; Currie et al. 2002). Wood decomposition results in cumulative losses of mass and volume as organic C is evolved as CO2 (Pregitzer and Euskirchen 2004) and the slow rates of wood decay make it a relatively long-term sink for C (Currie et al. 2002). Wood N and P are typically immobilized during the initial stages of decomposition and are eventually mineralized during advanced stages of decay (Harmon and Hua 1991; Chueng and Brown 1995; Rice et al. 1997). In most upland ecosystems downed woody debris is not a significant source of inorganic nutrients when compared to soil and foliar litter due to minimal initial nutrient concentrations (Laiho and Prescott 2004). However, given the vast differences in hydrology, soil properties, and vegetation types between floodplain and upland forests, this commonly held assumption may not apply in riparian ecosystems. It is evident that clarification regarding woody debris stocks, decomposition rates, and nutrient circulation are needed for wetland and riverine forests in order to better assess the importance of downed wood at an ecosystem scale.
In this study, we chose to investigate wood decomposition and nutrient dynamics in an old-growth bottomland hardwood forest located in Congaree National Park (CONG) in central South Carolina, USA. To date, relatively few decomposition studies have been completed in old-growth floodplain forests, although there has been a previous study of woody debris volume and loads within CONG (Wohl et al. 2011). This study indicated that the small tributaries within the floodplain had below average wood loads and relatively few logjams when compared to other old-growth floodplains in temperate climates. The authors concluded that the relative scarcity of wood in the floodplain was likely a result of the combined effects of high wood transport capacity and fast rates of decay. Although there is hydrological evidence that frequent spring floods facilitate wood transport at CONG, there have not been any studies regarding wood decomposition rates within the forests of the Congaree floodplain (Wohl et al. 2011). Knowledge of floodplain wood decay rates and associated nutrient dynamics would help elucidate the importance of woody debris in total ecosystem nutrient storage and circulation. To that end, the specific goals of this study were: (1) to quantify total downed woody debris mass and nutrient (C, N, P) pools; and (2) assess temporal trends of in situ woody debris decomposition and nutrient dynamics (mineralization and immobilization of C, N, P) over an approximate 2 year period. We hypothesized that wood decay rates would be greater than those reported for other temperate old-growth forests because of the effects of wetting–drying cycles associated with annual flooding at CONG and the humid subtropical climate of the region.
Methods
Study site
Congaree National Park is located in central South Carolina, approximately 30 km southeast from the city of Columbia. The Park is located within the upper Atlantic Coastal Plain and contains approximately 11,000 ha of bottomland hardwood forest associated with the Congaree River. The Congaree River drains a large portion (>21,000 km2) of the Piedmont physiographic province of central and northern South Carolina and the floodplain is classified as eutrophic because it receives nearly annual additions of silty, nutrient-rich sediment. The region has a humid subtropical climate that typically receives 1220 mm of annual precipitation and has an average annual air temperature of 17.6 °C. Our study was conducted in an old-growth bottomland forest tract near Butterfly Pond (33°46′19.33″N, 80°43′36.55″W). Five study transects (255–380 m in length) were established adjacent to the active Congaree River channel representing an alluvial soil toposequence (Fig. 1). The lateral toposequence included four major fluvial landscape units defined as the natural river levee, hardwood flat, transitional flat, and backswamp. Representative research plot sites were selected based on relative elevation with regard to the river channel and plant species characteristics. Four circular research plots were established along each transect (20 total plots, 5 m radius, 78.5 m2) to represent the major fluvial landscape units. Soils in this section of the floodplain are mapped at a series level as well drained Congaree (Fine-loamy, mixed, active, nonacid, thermic Oxyaquic Udifluvents) and poorly drained Chastain (Fine, mixed, semiactive, acid, thermic Fluvaquentic Endoaquepts) units (Soil Survey Staff 2013). Dominant overstory tree species in the study area ranged from mixed hardwoods such as sweetgum (Liquidambar styraciflua L.), sugarberry (Celtis laevigata Willd.), sycamore (Platanus occidentalis L.), and various bottomland oaks (Quercus spp.) on the river levee to baldcypress (Taxodium distichum (L.) Rich.) in the backswamp wetlands.

Overview map of study transects and sample plots. The wood harvest site depicted in the inset map was where live red maple (Acer rubrum) saplings were harvested for the decomposition study
Woody debris decomposition
Woody debris decomposition rates were assessed along the study toposequence using an in situ method similar to that described by Rice et al. (1997). Wood decay is dependent on initial litter quality, which can vary by tree species (Koch 1985; Berg and McClaugherty 2008), therefore a common species was selected to limit variability in wood quality. Red maple (Acer rubrum L.) wood was used, because this overstory species grows in both well drained natural river levees and poorly drained backwater sloughs of the region. We harvested live red maple saplings in March 2011 from a common site located approximately 5.5 km downstream of the study transects (Fig. 1). The harvest site was chosen because it has similar soil characteristics to those of the study transects (a gradient from Congaree to Chastain soil mapping units).
We cut saplings into approximately equal segments of 30 cm length and 3.0–4.5 cm diameter. Green wood segments were then air-dried to a constant weight in the laboratory (approximately 2 weeks). Initial wood volume was calculated (to the nearest mm3) from total length and mean diameter, measured at the edges and center of each segment using a hand-held caliper. Initial sample mass was recorded to the nearest 0.01 g and each segment was individually tagged for later identification. Control wood dowels (eastern white pine, Pinus strobus L.) were also cut into 30 cm lengths and initial mass was quantified in a similar manner to that of the red maple wood. Control wood dowels were used to evaluate the relative importance of landscape variability on decomposition of a substrate with homogeneous litter quality.
Red maple and control samples were placed in the field in April 2011. Tagged wood samples were placed in contact with the soil surface and tethered to trees on common strings to prevent sample loss during spring floods at CONG. Each string consisted of 10 wood segments, with duplicate red maple strings paired with a control wood string in each study plot (N = 400 maple, N = 200 control samples). Time zero maple and control wood samples were taken from each string, placed in sealed polyethylene sample bags, and transported to the laboratory in coolers. Wood samples were stored at 4 °C until processed in the laboratory. Initial samples were used to estimate initial handling mass loss, nutrient contents, and litter quality. The remaining samples were left in the field and collected after approximately 1, 2, 3, 6, 12, 15, 18, 24 months in the field to estimate decomposition and nutrient dynamics over time. During each field collection a total of 40 maple (n = 2 per plot, 10 per landscape) and 20 control (n = 1 per plot, 5 per landscape) wood samples were taken.
Initial wood samples were returned to the laboratory and oven-dried to a constant weight (approximately 7 days) at 65 °C. We corrected initial air-dried weights to oven-dried weights, and then remeasured volume and mass to calculate losses due to handling and transport. Ash weight (subsamples heated at 500 °C for 8 h) was subtracted from total wood mass to correct for any mineral additions. Corrections for mean differences in moisture, handling loss, and ash weight were applied to all sample collections. We remeasured mass and volume in a similar manner for all subsequent field collections to calculate wood decomposition rates over time.
Litter quality and nutrient dynamics
Following mass and volume measurements, nutrient concentrations were quantified to evaluate changes in litter quality and nutrient dynamics during wood decomposition. Three 2-cm disks (including bark, sapwood, and heartwood) were cut from each wood sample and ground to pass a 20-mesh (0.1 cm) screen with a laboratory Wiley mill. We measured total C and N concentration from composite stick samples using thermal combustion on a Perkin Elmer 2400 Series II CHNS/O analyzer (Perkin Elmer, Waltham, MA). Total plant tissue P was analyzed using the vanadomolybdate procedure on an HCl extract following sample-ashing at 500 °C (Jackson 1958). Lignin concentration was quantified using the acid detergent fiber method (Van Soest and Wine 1968). Nutrient concentrations were used to calculate molar C:N, C:P, N:P ratios and estimate total nutrient contents by multiplying the concentration by the wood dry mass (Rice et al. 1997). We quantified wood nutrient content after each collection and changes in nutrient dynamics (mineralization/immobilization) were assessed relative to mean time zero content.
Study site characterization and climate data
Precise soil surface heights (±2 mm vertical accuracy) were measured at each study plot with a TopCon GTS-240NW total station (Topcon Positioning Systems Inc., Livermore, CA) (Edward Schenk, personal communications). We collected multiple soil cores (n = 3, 0–10 cm depth) from each study plot for further laboratory analyses. Soil pH was measured from air dried subsamples using an 1:1 soil-deionized water slurry (Soil Survey Laboratory Staff 2004). Soil particle size distribution (texture) was quantified using the hydrometer method (Gee and Bauder 1986). Total downed wood volume and mass were estimated using modified transect sampling methods utilized by the United States Forest Inventory and Analysis program (Woodall and Williams 2005). We collected woody debris data along four 25 m parallel transects within twenty 40 × 25 m permanent plots (80 total transects). Volume of dead wood was estimated using the formula for the frustum of a right circular cone summed at the plot level (Waddell 2002). We calculated wood mass from volume estimates using standard procedures that employ decay class reduction factors and specific gravity values by species to obtain dry mass (Waddell 2002; Woodall and Williams 2005). Composite downed wood samples were collected from each study plot (n = 14 per plot) to calculate mean nutrient contents. Total woody debris C, N, and P were quantified in a similar manner to decomposition samples as indicated above. Monthly climate data including precipitation (mm), mean daily air temperature (°C), and Palmer drought severity index (PDSI) values were collected from the nearest continuous monitoring station located approximately 32 km northeast of the study plots at the Columbia Metropolitan Airport, South Carolina (NOAA, National Climatic Data Center, Asheville, NC).
Red imported fire ants
During the October 2011 field collection, we qualitatively observed significant damage to the bark and outermost sapwood of the red maple sticks retrieved from the field (control wood was not affected). During laboratory processing approximately 65 live ants were collected from the red maple sample bags and later identified as red imported fire ants (RIFA, Solenopsis invicta Buren) (Wayne Clark, personal communications). No other ant species were present at the time of sampling and wood damage was attributed to RIFA activity. We estimated percent damage using point count methods for each collection after infestation (October 2011 until March 2013) to quantify the impact of RIFA damage on wood decomposition. Photographs of the upper and lower (soil side down) sides of each individual red maple segment were taken at a fixed vertical distance from the sample (26 cm). Digital photographs (314 dpi) were downloaded and fixed at dimensions of 27 × 18 cm. Photographs were then overlain with a counting grid (2.25 cm2 cells) and an equidistant dot grid (2.2 points per cm2 density). Points falling within the photographed area of the sample stick were quantified into two categories; damaged or undamaged. Tallies were completed manually for both photographs of each sample and surface damage was quantified as a percent of total points tallied for each sample.
Statistical analysis
Statistical analyses were performed in SAS 9.2 (SAS Institute, Cary, NC). Mean comparisons between initial (time 0) and final (after 700 days) litter quality metrics including lignin content, lignin:N, and molar nutrient ratios (C:N, C:P, N:P) were completed using Student’s t-tests. Mean soil characteristics, downed woody debris standing stocks and nutrient storage, maple wood decomposition rates (mass remaining, k constants), and C, N, P content remaining were compared among the four transect landscapes (levee, flat, transitional, backswamp) using one-way analysis of variance (ANOVA) with Tukey’s honest significant difference tests (PROC GLM). Wood decomposition rates were quantified by fitting a negative exponential decay equation outlined by Olson (1963): y = e−kt, where k is the decay constant, t the time (years), and y is the fraction (mass at time t/initial mass) of original dry mass remaining at time t. Plot-specific k constants were used to calculate wood half-life (0.693/k) and 95 % decomposition (3/k) rates (Olson 1963). To evaluate the relationship between RIFA damage and wood decomposition, a logarithmic regression was fitted between surficial damage counts (% surface area) and mass remaining (% original mass). Nonlinear regressions were fitted in SAS (PROC NLIN) using multiple iterations and initial parameter estimates (β0, β1) derived from simple linear regressions (PROC REG). All statistical tests were evaluated using α = 0.05.
Results
Toposequence and climate characteristics
The study toposequence had an overall elevation gradient of 1.62 m from the river levee to backswamp positions (Table 1). Mean surface soil pH was significantly (ANOVA F3,16 = 19.16, P < 0.001) greater on the levee (5.91 ± 0.08) and flat (5.72 ± 0.17) landscapes compared to the lower transitional (5.01 ± 0.05) and backswamp (5.11 ± 0.08) positions. Clay content (0–10 cm depth) was significantly greater (F3,16 = 43.32, P < 0.001) in the backswamp and transitional areas compared to the higher elevation levee and flats. Mean standing stocks of downed woody debris were variable for all landscapes, ranging from a minimum of approximately 9 ± 3 Mg ha−1 for the flat to 24 ± 8 Mg ha−1 in the backswamp landscape. There were no significant differences in total downed woody debris mass (F3,36 = 1.05, P = 0.38) or nutrient pools (C, N, P) by landscape (Table 1). Mean air temperatures during our study were above average and monthly precipitation was below average from August 2011 until April 2012. The combination of above average temperatures and lack of precipitation resulted in moderate to severe drought conditions throughout the central South Carolina region in 2011 and early 2012 (Fig. 2).

Climatological data over the study period (April 2011–March 2013) from the Columbia South Carolina Metropolitan Airport monitoring station and corresponding Palmer drought severity index (PDSI) for the central region (Division 6) of South Carolina (data source: National Oceanic and Atmospheric Administration, National Climatic Data Center, Asheville, North Carolina, USA). PDSI values <−2 indicate drought conditions
Woody debris decomposition
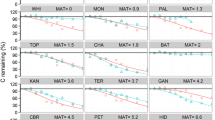
Wood decomposition rates, expressed as a percent original mass remaining, were similar across the toposequence landscapes (Fig. 3a). Control wood (milled eastern white pine) was low in initial nutrients and decayed much slower than red maple samples. After 700 days in the field, average control wood mass remaining was still approximately 90 %. By contrast, the red maple samples decomposed much faster, with an average mass remaining of 29 % after 700 days in the field. Mass remaining data were similar to wood C content and there were no statistical differences in mean C content by landscape during the study (Fig. 3b). Mean C remaining was only 29 % after 700 days in the field and showed net mineralization during all collections.

Summary of wood mass and carbon remaining over the course of the decomposition study. a Mean (n = 10 per collection, ±1 standard error) red maple (Acer rubrum) and control (Pinus strobus dowels) woody debris mass loss (ash free) by landscape from April 2011 to March 2013. b Mean (n = 10 per collection, ±1 standard error) carbon remaining in red maple woody debris by landscape from April 2011 to March 2013. Red imported fire ants (Solenopsis invicta) infested the red maple wood in September 2011, approximately 150 days into the decomposition study
Maple wood decomposition was approximately 3-times faster than control wood, suggesting litter quality was an important regulator of wood decay (Fig. 3a). Initial maple wood lignin concentrations ranged from 12.5 ± 0.3 to 13.4 ± 0.4 % and increased over the decomposition study to >24 % in all toposequence landscapes (Table 2). Initial lignin:N ratios ranged from 50.7 ± 3.7 to 69.7 ± 8.7 and decreased in all landscapes as the wood decayed. Mean initial C:N ratios were wide, ranging from 194 ± 14 in the transitional position to 252 ± 25 in the backswamp. Over the course of the study C:N ratios decreased to ≤125 in all landscapes. C:P ratios were variable, both initially and in the final wood collection (Table 2). The drier toposequence positions (levee, flat) showed net decreases in C:P with decomposition, while the wetter landscapes (transitional, backswamp) displayed increasing C:P ratios. Wood N:P ratios were initially narrow (range 6.3 ± 0.5 to 6.9 ± 0.5), however, after 700 days in the field N:P ratios widened to >10 in all landscapes.
Wood nutrient dynamics during decomposition
Wood N content displayed both mineralization periods and immobilization peaks (Fig. 4). The largest immobilization phase was observed in the 183 days collection that followed RIFA disturbance in autumn 2011. There was numerically more relative N content in the wetter toposequence landscapes (>125 %) compared to the higher elevation flat (111 ± 12.5 %) and river levee (98.8 ± 7.6 %). Following RIFA infestation and net immobilization phases, N gradually decreased below 100 % original content. Only during the 459 days collection (July 2012) was there significantly (F3,36 = 3.19, P = 0.035) more relative N content in the transitional (92.6 ± 9.2 %) position compared to the backswamp (58.5 ± 5.4 %) landscape (Fig. 4). Another immobilization peak occurred at 548 days (October 2012) followed by net mineralization of N. Mean wood N content after 700 days ranged from 50.8 ± 4.4 to 68.9 ± 6.0 %.

Mean nitrogen remaining (n = 10 per collection, ±1 standard error) in red maple woody debris by landscape from April 2011 to March 2013. Means with different letters are significantly different according to Tukey’s HSD tests
Wood P content showed less immobilization compared to N (Fig. 5). There was a significant difference (F3,36 = 3.53, P = 0.024) in P content at 59 days (June 2011), with greater P in the transitional (92.0 ± 5.0 %) compared to the levee (70.9 ± 3.4 %) positions. The following collection at 95 days indicated net P mineralization in all landscapes except the natural levee. Wood P immobilization in the levee resulted in highly significant (F3,36 = 13.82, P < 0.001) differences among landscapes with the general relationship; levee > transitional ≥ flat ≥ backswamp. RIFA infested the wood in autumn 2011 and the 183 days collection (October 2011) displayed large immobilization peaks in the flat, transitional, and backswamp landscapes (Fig. 5). There was significantly (F3,36 = 34.25, P < 0.001) more P immobilization in the backswamp (262 ± 19 %) and transitional (220 ± 12 %) landscapes compared to the flat (148 ± 10 %) and levee (95 ± 5 %). Following the RIFA damage, wood P content displayed a general trend of mineralization and there were no significant differences among the toposequence landscapes. During the final 700 days collection, wood P remaining ranged from 29 ± 2 to 38 ± 4 % of original content.

Mean phosphorus remaining (n = 10 per collection, ±1 standard error) in red maple woody debris by landscape from April 2011 to March 2013. Means with different letters are significantly different according to Tukey’s HSD tests, note the graph inset of significant differences prior to ant infestation
Red imported fire ants
RIFA were found in maple wood in October 2011 and there were significant negative, logarithmic relationships between surface damage and mass remaining in all subsequent collections (Fig. 6). Immediately after infestation (October 2011), most wood had <20 % surface damage, but there was still significantly (R2 = 0.22, F1,38 = 11.01, P < 0.01) less mass remaining in samples with greater damage. For all collections, samples displaying the most bark and outer sapwood damage decomposed faster relative to undamaged wood. As time progressed, samples displayed greater amounts of decay, although the relationship with surface damage remained significant.

Summary of significant logarithmic relationships between surface area damaged on red maple (Acer rubrum) woody debris by red imported fire ants (Solenopsis invicta) and mass remaining through time averaged over all toposequence plots
Following the final 700 days collection, exponential decay constants (k) were calculated for each study plot and landscape. Mean decay constants were statistically similar (F3,36 = 0.36, P = 0.78) among the toposequence landscapes, with a mean k value (year−1) of 0.650 ± 0.02 (Table 3). Despite differences in RIFA damage, mean exponential decay model coefficient of determination (R2) values ranged from 0.87 ± 0.03 to 0.92 ± 0.01, indicating the models fit the data fairly well. The average wood half-life and time until 95 % decay was 1.11 ± 0.03 and 4.79 ± 0.15 years, respectively.
Discussion
Woody debris pools and decomposition
Downed woody debris mass was highly variable (range 9–24 Mg ha−1) along the study toposequence. Woody debris pools in CONG were within the range of those typically reported for temperate deciduous forests (11–38 Mg ha−1; Harmon et al. 1986). Estimates of downed woody debris in CONG were also greater than those reported by Polit and Brown (1996) of 6.6 Mg ha−1 for a floodplain forest in Illinois (USA), but similar to values reported by Muller (2003) of 21.8 Mg ha−1 for an old-growth deciduous forest in Kentucky (USA). Woody debris nutrient pools (specifically N and P) were also variable across the study toposequence in CONG (Table 1). Our estimates of wood biomass and nutrient storage are a reflection of greater dead wood inputs associated with the old-growth forests at CONG and frequent landscape disturbances, such as hurricanes, in the region (Harmon et al. 1986; Putz and Sharitz 1991). However, when compared to other old-growth forests, downed woody debris stocks were relatively low at CONG, probably due to fast wood turnover times (Wohl et al. 2011).
Decay rates for red maple sticks in this study were very fast, with an average k rate constant of 0.650 ± 0.2 year−1 (Table 3). Wood decay rates at CONG were much higher than those reported for temperate forests, ranging from 0.004 to 0.520 year−1, but typically <0.100 year−1 (Harmon et al. 1986; Mattson et al. 1987). A similar decomposition study conducted in an upland landscape of Massachusetts (USA) reported that red maple sticks <5 cm diameter had 25.4 % mass remaining after 5 years (k = ~0.274 year−1, Berg and McClaugherty 2008). In comparison to data from floodplains and wetlands, red maple wood decomposition at CONG was faster than whole silver maple (Acer saccharinum Marsh.) logs in Illinois (k = 0.089 year−1, Chueng and Brown 1995) and red maple wood in the Great Dismal Swamp of North Carolina (k range 0.179–0.305 year−1, Day 1982). Rice et al. (1997) also reported average mass remaining of downed pumpkin ash (Fraxinus profunda Bush) fine woody debris to be ≥42.4 % after 2.5 years in the Atchafalaya basin of Louisiana (USA). These decomposition rates were all less than those at CONG, where <31.4 % original wood mass remained after only 1.92 years (Fig. 3a).
These data indicate that wood decay rates at CONG are faster than those in other temperate or humid subtropical zone forests, and approach values reported in tropical ecosystems (Clark et al. 2002). In tropical climates, whole bole k rates have been reported up to 0.67 year−1 (Chambers et al. 2000) and wood turnover times are typically fast, ranging from 5.9 to >20 years at the ecosystem level (Buxton 1981; Delaney et al. 1998; Clark et al. 2002). The average time required for 95 % wood decomposition at CONG was 4.79 years, which is similar to rates reported in tropical climates. In our study, we utilized fine woody debris (3.0–4.5 cm diameter) from a single overstory species and it is important to note that the high rates of decomposition for these samples may not reflect all downed wood within the riverine forests of CONG. Although data from our study and past observations (Wohl et al. 2011) suggest extremely fast wood turnover within CONG, further analyses of wood from mixed-species and varied diameter classes are necessary to quantify ecosystem level wood decomposition dynamics.
Fast rates of red maple wood decomposition observed at CONG are likely a result of multiple factors such as initial litter quality, regional humid subtropical climate, and invertebrate activity. Litter quality is extremely important to the decomposition process (Melillo et al. 1982), and litter low in lignin and high in N typically decays faster than materials with high lignin and low N content (Taylor et al. 1989). The lignin concentration of wood from CONG (Table 2) was nearly half the reported average values for red maple wood ranging from 20.6 to 24.0 % (Koch 1985; Harmon et al. 1986). Although lignin concentration in CONG wood was low, implying favorable litter quality for decomposition, mean lignin:N ratios were wide (>50) because of low N concentrations. Initial wood C:N ratios at CONG were also at the upper limits of those typically reported for fresh (green) wood ranging from 80 to 250 (Koch 1985), further suggesting low N content. An additional consideration related to initial litter quality is the wood decay class when it is added to the forest floor. In this study we used fresh cut green red maple wood to limit the variability among initial litter quality parameters. Although this is a widely used method for wood decay studies (Rice et al. 1997; Chueng and Brown 1995), it may not be representative of all wood added to the forest floor. Furthermore, green wood tends to have higher concentrations of soluble nutrients (K, N, P, S, C) and fast decaying simple organic compounds (saccharides, carbohydrates, cellulose) compared to standing dead wood (Berg and McClaugherty 2008). Therefore, green wood trends to decompose quicker when added to the forest floor, at least initially, which may have contributed to the fast decay rates we observed in this study.
Climatic factors can also be important for litter decomposition dynamics. Globally, litter decay rates are generally faster in warm and wet climates, compared to cool and dry regions (Zhang et al. 2008). However, temperatures exceeding 40 °C or below 5 °C have been shown to inhibit mesophilic microbial processes, such as C mineralization (Harmon et al. 1986). CONG is within a humid subtropical climate which rarely has average air temperatures <5 °C (Fig. 2), suggesting microbial mineralization can occur year-round. Carbon mineralization data from our study support this idea because total C mineralization occurred throughout the study (Fig. 3b). Although temperature conditions were favorable for C mineralization during our study, the region also underwent a wide-spread drought which could have slowed microbial processes by limiting moisture availability within the red maple wood (Berg and McClaugherty 2008). Climatological data indicate that while CONG did undergo moderate to severe drought conditions in 2011–2012, there was approximately 50 mm or greater precipitation during these months (Fig. 2). This rainfall likely produced enough moisture to continue microbial decay of wood because net C mineralization occurred during and after drought conditions at the study toposequence (Fig. 3b). While microbial decay and mineralization of C are recognized as major drivers of wood decomposition (Harmon et al. 1986; Currie et al. 2002), substantial losses of wood mass can also occur via faunal activity and physical fragmentation.
Invertebrate activity and wood decay
Regional drought conditions within the study period (Fig. 2) lowered ground water tables, prevented substantial overbank flooding, and allowed for the establishment of RIFA colonies throughout our study site. The RIFA is a non-native invasive species that was introduced into the southern USA during the early 20th century (Allen et al. 2004). Since their introduction, RIFAs have had many documented effects on North American ecosystems. Native invertebrate and vertebrate species (ground nesting birds, reptiles, mammals) have all seen negative population impacts due to RIFAs (Morrison 2002; Allen et al. 2004). RIFA activity can also alter soil properties and biogeochemistry. Many researchers have reported significant increases in soil organic matter, clay, P, K, Ca, and Mg content within RIFA mounds (Lockaby and Adams 1985; Green et al. 1998; Lafleur et al. 2005). Inherent changes in soil properties can also increase heterotrophic respiration and greenhouse gas emissions (N2O, CH4, CO2) from RIFA mounds (Bender and Wood 2003). Although many changes to soil properties are highly localized (i.e., at a mound scale), entire landscapes can be impacted within a matter of decades because RIFA colonies tend to construct and abandon mound sites frequently in the southern USA (Green et al. 1999).
In contrast to the research of RIFA impacts on native fauna and soils, there have been relatively few studies of their effects on litter decomposition. Recent studies in the southern USA have indicated that the presence of ant colonies in woody debris can slow decomposition, because ants prey on wood-consuming termites and limit saprophytic fungi growth through anti-microbial metapleural secretions (Warren and Bradford 2012). These processes are not directly applicable in CONG because significant RIFA infestation occurred in response to a major drought event and there was no evidence that the study wood was used as permanent nesting habitat after the autumn of 2011. The data from CONG suggest that RIFA damage facilitated increased decomposition via microbial colonization of the wood impacted by burrowing. Similar invertebrate impacts on wood decomposition have been reported. For example, borings in fresh woody debris from ghost moths (Hepialus californicus Boisduval) have been shown to accelerate decay rates by increasing surface area and stimulating microbial decomposer colonization (Bastow et al. 2008).
RIFA presence in and utilization of wood as a food source is not unprecedented in the southern USA. RIFA nests have been observed in large downed wood and tree stumps following clear-cut operations in the region (Lockaby and Adams 1985). Outside of their native range RIFAs are omnivorous feeders and multiple reports of RIFA consuming tree sap, floral/extrafloral nectar, and bark have been made in North America (Adams 1986; Banks et al. 1991; Wilder et al. 2011). Furthermore, damage to wood from RIFA activity most frequently occurs during autumn and winter months, corresponding to periods of colony demand for carbohydrates (Adams 1986; Banks et al. 1991). Our wood samples were infested and damaged by RIFAs in September and October, similar to previously reported time periods of RIFA wood infestation in the region.
We hypothesize that RIFA infestation may have facilitated wood decomposition. Results from our study show that the surficial damage caused by invertebrate activity was significantly correlated with the wood decomposition process. Over time, we noted that the total percent surficial damage was increasing as wood decayed (see Fig. 6 x-axis). Mean wood surface damage immediately following RIFA infestation was only 11.4 ± 2.6 % in October 2011, but increased during subsequent collections to 44.6 ± 3.9 % after 700 days in the field. Wood damage was limited to the outer bark and sapwood when RIFAs were found in the samples, but after 459 days in the field (July 2012), extensive invertebrate activity had stripped much of the bark down to the sapwood and wood-borings were observed penetrating to the inner heartwood of many sticks. Following their initial discovery, there were no more instances of RIFA presence in the maple wood. Therefore, the significant increase in wood-borings, surface damage, and wood fragmentation observed in collections from 459 to 700 days are probably not a direct result of RIFAs, but rather foraging invertebrates, such as termites or beetles. These trends highlight the importance of RIFA as possible facilitators of advanced wood decomposition by opening the outermost bark and sapwood for further invertebrate and microbial utilization over time.
Downed woody debris nutrient dynamics
In CONG, N content displayed multiple periods of immobilization, especially after RIFA infestation (Fig. 4). Sink activity of N has been noted in the early stages of wood decay in floodplain wetlands (Chueng and Brown 1995; Rice et al. 1997) which has been attributed to overstory throughfall additions, senesced litter leachates, and microbial immobilization of soil N (Day 1982; Harmon et al. 1986). The largest N immobilization peaks were at 183 and 550 days corresponding to autumn 2011 and 2012, time periods where the potential availability of N from throughfall and senesced litter is greatest. Increased N content during these periods suggests that microbial decomposer communities within the wood are utilizing seasonal nutrient additions and hence may be N limited. Wood P dynamics at CONG differed from those observed for N and generally showed source activity (net mineralization) except for the 183 days collection immediately after RIFA infestation (Fig. 5). The large peaks in both N and P after RIFA infestation suggest both nutrients were increased as a result of initial ant activity. Invertebrates can increase N and P content in forest litter as a direct result of insect frass (fecal matter) deposition, which has high nutrient contents relative to woody material (Bocock 1964; Day 1982) and also through indirect inoculation of the substrate with microorganisms which can further immobilize nutrients (Harmon et al. 1986; Berg and McClaugherty 2008).
Wood N and P dynamics suggest more pronounced N limitation, because P generally mineralized after RIFA infestation and N showed more sink behavior indicating the microbial community was likely N deficient. In addition, the initial wood N:P ratios were narrow (<7.0, Table 2) further suggesting N limitation to vegetation growth within CONG (Bedford et al. 1999). These data fit the framework of terrestrial nutrient limitation, because CONG is a eutrophic floodplain forest associated with an alluvial red water river that receives mineral additions from watershed sources. Alluviation deposits mineral and organic P at the soil surface in red water floodplains which can exacerbate N limitation over time (Vitousek et al. 2010). Although these data support the idea that the ecosystem is N limited, further fertilization experiments are needed to determine the exact nature of nutrient deficiency within the floodplain forests of CONG.
Unlike N and P dynamics, C immobilization is rare in wood and net mineralization occurred at all stages of decomposition in CONG (Fig. 3b). A major effect of C mineralization is relative increases in wood N and lignin concentrations during advanced stages of decay (Rice et al. 1997; Berg and McClaugherty 2008). Wood C:N ratios narrowed and average lignin concentrations after 700 days ranged from 24.6 ± 1.8 to 26.9 ± 2.7 (Table 2), which are close to those reported for fresh red maple wood (20.6–24.0 %, Koch 1985; Harmon et al. 1986). The activity of invertebrates and high wood litter quality likely contributed to the observed fast turnover times and validates earlier assumptions that rapid decay is partially responsible for the relatively small downed woody debris pools within the old-growth floodplain forests of CONG (Wohl et al. 2011).
Conclusions
In this study, we quantified downed woody debris pools and turnover times within an old-growth floodplain forest. Our results suggest that the relatively low standing stocks of woody debris are a function of fast turnover times at CONG. We hypothesize that the rapid decomposition we observed was influenced by invertebrate activity leading to wood fragmentation and microbial inoculation during the initial stages of decay. These processes could facilitate additional mineralization of organic C because the warm, humid subtropical climate of the region rarely limits microbial decomposition. Additional studies of these processes are clearly necessary to better quantify the specific impacts of invasive RIFA on woody litter decay. In addition, the red maple wood utilized in our study was found to have high initial litter quality, characterized by low lignin concentration. The fast decay rates within the floodplain forests of CONG has obvious consequences for the long term C storage function of woody debris. Although increased annual losses of C via mineralization implies a net negative ecosystem function, associated rapid cycling of stored N and P may subsidize plant growth leading to increases in net primary productivity and a complete or partial offset of total ecosystem C losses associated with woody debris stocks. Our analysis indicates that woody debris pools are capable of cycling nutrients rapidly, especially in regions where climatic factors allow for nearly continuous year-round faunal disturbance and microbial mineralization. Thus, future floodplain nutrient budgets and ecosystem models should take into account woody debris as a changing and dynamic nutrient pool to better understand the impact of potentially rapid nutrient cycling on the functions and values associated with large riparian forests.
References
Adams CT (1986) Agricultural and medical impact of the imported fire ants. In: Lofgren CS, Vander Meer RK (eds) Fire ants and leaf-cutting ants, biology and management. Westview Press, Boulder, pp 48–57
Allen CR, Epperson DM, Garmestani AS (2004) Red imported fire ant impacts on wildlife: a decade of research. Am Midl Nat 152:88–103
Banks WA, Adams CT, Lofgren CS (1991) Damage to young citrus trees by red imported fire ants (Hymenoptera: Formicidae). J Econ Entomol 84:241–246
Bastow JL, Preisser EL, Strong DR (2008) Wood decomposition following a perennial lupine die-off: a 3-year litterbag study. Ecosystems 11:442–453
Bedford BL, Walbridge MR, Aldous A (1999) Patterns in nutrient availability and plant diversity of temperate North American wetlands. Ecology 80:2151–2169
Bender MR, Wood CW (2003) Influence of red imported fire ants on greenhouse gas emissions from a Piedmont Plateau pasture. Commun Soil Sci Plant Anal 34:1873–1889
Berg B, McClaugherty C (2008) Plant litter: decomposition, humus formation, carbon sequestration, 2nd edn. Springer-Verlag, Berlin
Bocock KL (1964) Changes in the amounts of dry matter, nitrogen, carbon and energy in decomposing woodland leaf litter in relation to the activities of soil fauna. J Ecol 52:273–284
Braccia A, Batzer DP (2008) Breakdown and invertebrate colonization of dead wood in wetland, upland, and river habitats. Can J For Res 38:2697–2704
Buxton RD (1981) Termites and the turnover of dead wood in an arid tropical environment. Oecologia 51:379–384
Chambers JQ, Higuchi N, Schimel JP, Ferreira LV, Melack JM (2000) Decomposition and carbon cycling of dead trees in tropical forests of the central Amazon. Oecologia 122:380–388
Chueng N, Brown S (1995) Decomposition of silver maple (Acer saccharinum L.) woody debris in a central Illinois bottomland forest. Wetlands 15:232–241
Clark DB, Clark DA, Brown S, Oberbauer SF, Veldkamp E (2002) Stocks and flows of coarse woody debris across a tropical rain forest nutrient and topography gradient. For Ecol Manag 164:237–248
Currie WS, Yanai RD, Piatek KB, Prescott CE, Goodale CL (2002) Processes affecting carbon storage in the forest floor and in downed woody debris. In: Kimble JM et al (eds) The potential of U.S. forest soils to sequester carbon and mitigate the greenhouse effect. CRC Press, Boca Raton, pp 135–157
Day FP (1982) Litter decomposition rates in the seasonally flooded Great Dismal Swamp. Ecology 63:670–678
Day FP (1983) Effects of flooding on leaf litter decomposition in microcosms. Oecologia 56:180–184
Delaney M, Brown S, Lugo AE, Torres-Lezama A, Quintero NB (1998) The quantity and turnover of dead wood in permanent forest plots in six life zones of Venezuela. Biotropica 30:2–11
Gee GW, Bauder JW (1986) Particle-size analysis. In: Klute A (ed) Methods of soil analysis Part 1, 2nd edn. ASA and SSSA, Madison, pp 383–411
Green WP, Pettry DE, Switzer RE (1998) Impact of imported fire ants on the texture and fertility of Mississippi soils. Commun Soil Sci Plant Anal 29:447–457
Green WP, Pettry DE, Switzer RE (1999) Structure and hydrology of mounds of the imported fire ants in the southeastern United States. Geoderma 93:1–17
Harmon ME, Hua C (1991) Coarse woody debris dynamics in two old-growth ecosystems. Bioscience 41:604–610
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD, Anderson NH, Cline SP, Aumen NG, Sedell JR, Lienkaemper GW, Cromack K Jr, Cummins KW (1986) Ecology of coarse woody debris in temperate ecosystems. Adv Ecol Res 15:133–302
Hättenschwiler S, Tiunov AV, Scheu S (2005) Biodiversity and litter decomposition in terrestrial ecosystems. Annu Rev Ecol Evol Syst 36:191–218
Jackson ML (1958) Soil chemical analysis. Prentice-Hall, Englewood Cliffs
Koch P (1985) Utilization of hardwoods growing on southern pine sites. Forest service agricultural handbook number 605. Government Printing Office, Washington DC
Lafleur B, Hooper-Bui LM, Mumma EP, Geaghan JP (2005) Soil fertility and plant growth in soils from forests and plantations: effect of invasive imported fire ants Solenopsis invicta (Buren). Pedobiologia 49:415–423
Laiho R, Prescott CE (2004) Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: a synthesis. Can J For Res 34:763–777
Lockaby BG, Adams JC (1985) Pedoturbation of a forest soil by fire ants. Soil Sci Soc Am J 49:220–223
Mattson KG, Swank WT, Waide JB (1987) Decomposition of woody debris in a regenerating, clear-cut forest in the southern Appalachians. Can J For Res 17:712–721
Melillo JM, Aber JD, Muratore JF (1982) Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 63:621–626
Morrison LW (2002) Long-term impacts of an arthropod-community invasion by imported fire ant, Solenopsis invicta. Ecology 83:2337–2345
Muller RN (2003) Landscape patterns of change in coarse woody debris accumulation in an old-growth deciduous forest on the Cumberland Plateau, southeastern Kentucky. Can J For Res 33:763–769
Olson JS (1963) Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44:322–331
Polit JI, Brown S (1996) Mass and nutrient content of dead wood in a central Illinois floodplain forest. Wetlands 16:488–494
Pregitzer KS, Euskirchen EN (2004) Carbon cycling and storage in world forests: biome patterns related to forest age. Glob Change Biol 10:1–26
Putz FE, Sharitz RR (1991) Hurricane damage to old-growth forest in Congaree Swamp National Monument, South Carolina, USA. Can J For Res 21:1765–1770
Rice MD, Lockaby BG, Stanturf JA, Keeland BJ (1997) Woody debris decomposition in the Atchafalaya River Basin of Louisiana following hurricane disturbance. Soil Sci Soc Am J 61:1264–1274
Schilling EB, Lockaby BG (2005) Microsite influence on productivity and nutrient circulation within two southeastern floodplain forests. Soil Sci Soc Am J 69:1185–1195
Soil Survey Laboratory Staff (2004) Soil survey laboratory methods manual. Soil survey investigation report 42 version 4.0. Government Printing Office, Washington DC
Soil Survey Staff (2013) Web soil survey: Soil data mart. NRCS, Washington DC. http://websoilsurvey.nrcs.usda.gov. Accessed 18 July 2013
Taylor BR, Parkinson D, Parsons WFJ (1989) Nitrogen and lignin content as predictors of litter decay rates: a microcosm test. Ecology 70:97–104
Tyrrell LE, Crow TR (1994) Dynamics of dead wood in old-growth hemlock-hardwood forests of northern Wisconsin and northern Michigan. Can J For Res 24:1672–1683
Van Soest PJ, Wine RH (1968) Determination of lignin and cellulose in acid detergent fiber with permanganate. J Assoc Off Anal Chem 51:780–785
Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010) Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecol Appl 20:5–15
Waddell KL (2002) Sampling coarse woody debris for multiple attributes in extensive resource inventories. Ecol Ind 1:139–153
Warren RJ II, Bradford MA (2012) Ant colonization and coarse woody debris decomposition in temperate forests. Insectes Sociaux 59:215–221
Wilder SM, Holway DA, Suarez AV, LeBrun EG, Eubanks MD (2011) Intercontinental differences in resource use reveal the importance of mutualisms in fire ant invasions. Proc Natl Acad Sci 108:20639–20644
Wohl E (2013) Floodplains and wood. Earth Sci Rev 123:194–212
Wohl E, Polvi LE, Cadol D (2011) Wood distribution along streams draining old-growth floodplain forests in Congaree National Park, South Carolina, USA. Geomorphology 126:108–120
Woodall C, Williams MS (2005) Sampling protocol, estimation, and analysis procedures for down woody materials indicator of the FIA program. General technical report NC-256, North Central Research Station. USDA-Forest Service, St. Paul
Zhang D, Hui D, Luo Y, Zhou G (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1:85–93
Acknowledgments
The authors are grateful for field and laboratory assistance provided by Robin Governo, Andrew Parsons, Robert Price, Lauren Behnke, Nate Click, Meg Bloodworth, Catherine Justice, Theresa Thom, Cliff Hupp, Ed Schenk, Frank Henning, and David Shelley. We also would like to thank Joey Shaw and Yucheng Feng who helped improve draft versions of this manuscript. Funding and support for this research was provided by the United States Geological Survey, the United States National Park Service, and the United States Department of Agriculture National Institute of Food and Agriculture under project number SC-1700424. Technical Contribution No. 6408 of the Clemson University Experiment Station.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: E. Veldkamp.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Ricker, M.C., Lockaby, B.G., Blosser, G.D. et al. Rapid wood decay and nutrient mineralization in an old-growth bottomland hardwood forest. Biogeochemistry 127, 323–338 (2016). https://doi.org/10.1007/s10533-016-0183-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-016-0183-y