
Abstract
Fire is responsible for the structure and composition of the biological communities in grasslands, despite this, few studies evaluate the effects of fire on bird communities, especially in the southern cone of South America. We compared the structure and composition of vegetation, bird communities, density of nests and predation rates between burnt and unburnt areas in grassland habitats in the Argentine Pampas during two reproductive seasons following a fire. The abundance, richness and diversity of birds were lower in the burnt areas and followed a pattern of interannual recovery associated with the increase in vegetation cover. The density of nests and the number of nesting species was higher in the controls only during the first season. The risk of predation of artificial nests in burnt areas was greater 1 year after the fire in respect to the controls, this difference decreased over time in inverse relationship with the percentage of plant cover surrounding the nests. Our results highlight the importance of a good vegetation structure and cover to support abundant and species-rich bird communities, as well as high values of nest density and reproductive success, and the ability of vegetation and bird communities to recover after the fire. However, this successional and temporary loss of habitats of high nesting value should be analyzed on a larger scale in order to consider the effect of the total reduction, and the fragmentation of the grasslands in a good state of conservation, on the population dynamics of grassland birds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fire is a central factor in the dynamics of different biomes around the world, to the point that several of them owe their distribution and ecological properties to the fire regime (Bond et al. 2004). Fire plays a crucial role in the dynamics of savannas, Mediterranean shrublands and boreal forests ecosystems (Meinrat and Goldammer 1992; Bowman and Murphy 2010; Pennington and Hughes 2014), being one of the factors responsible for the structure and composition of biological communities, particularly in grasslands (Blair et al. 2014).
Grasslands cover ca. one-third of the Earths vegetation cover and provide an important variety of ecosystem goods and services (Costanza et al. 1997; Zavaleta et al. 2010). Fire is a factor associated in a particularly strong way with grasslands, with roughly 80% of global fire taking place in these systems each year (Leys et al. 2018). Changes in fire regimes, active fire suppression policies and grazing modify the vegetation cover of grasslands and shrubs around the world (Eldridge et al. 2011; Ratajczak et al. 2014). A decrease in the frequency and the intensity of the fires with respect to their historical pattern is associated with increases in the cover of woody species and with the transformation of pastures in scrub and forest ecosystems (Ratajczak et al. 2014). Invasive plant species in particular often alter fire regimes, increasing their frequency, extent and intensity. Their rapid post-fire growth also results in positive feedback mechanisms that further increase the frequency of fires (D´ Antonio and Vitousek 1992; Williams and Zdravko 2000; Bowman and Murphy 2010). Over time this cycle can convert diverse ecosystems into environments dominated by a few exotic species and can result in key structural changes. While the effects of fire on the dynamics of vegetation have been widely studied, however there is less knowledge about the relationship between the fires and the wildlife (Fuhlendorf et al. 2006; Bendell 2012; Pausas and Parr 2018).
Diverse studies describe the reorganization of wildlife communities after a fire, which depends on the particular response of each species (Lyon et al. 2000; Vickery et al. 2005; Zozaya et al. 2010; Clavero et al. 2011). The effects of fire have been studied in invertebrates (Kiss and Magnin 2006; Sackmann and Farji-Brener 2006; Langlands et al. 2006), amphibians and reptiles (Homan 2014), birds (Barlow et al. 2006; Ukmar et al. 2007), and mammals (Recher et al. 2009; Homan 2012), including the direct consequences of fire on the behaviour of the animals and the indirect effects due to the impact on resources, such as food and shelter. In particular, the response of bird communities to a fire is determined by the characteristics of the fire (Lyon et al. 2000), the post-fire environment and the bird species in the community (Milesi et al. 2002; Bonier et al. 2007; Pons and Clavero 2010; Clavero et al. 2011). The generally small and fragmentary nature of grassland remnants in a good state of conservation implies that fire can have decisive consequences for the maintenance of the natural vegetation communities and the wild bird populations that depend on them, in particular the obligate grassland birds (Coppedge et al. 2008). Grassland birds are defined as those that spend a predominant part or their entire life cycle in grasslands, nesting on the ground or in the center or under clumps of grasses (Vickery et al. 1999). These include obligate grassland birds that are those strictly restricted to grasslands and do not use other habitats (Azpiroz et al. 2012).
Grassland birds are experiencing reductions in their abundance and distribution area globally, attributed mainly to the destruction, degradation and fragmentation of their habitats (Vickery et al. 1999; Askins et al. 2007). In particular, grasslands in southeastern South America (central and northeastern Argentina, Uruguay, southern Paraguay and Brazil) have undergone important changes due to the development of the livestock industry, agriculture and afforestation (Azpiroz et al. 2012), resulting in the replacement of natural grasslands. This has substantially changed the composition, structure and functioning of the original vegetation, and with this the associated bird communities (Comparatore et al. 1996; Krapovickas and Di Giacomo 1998; Cueto and Lopez de Casenave 1999; Codesido et al. 2008; Azpiroz et al. 2012). The borders of streams seem to be habitats of particular importance as they provide habitat for more grassland birds species than other vegetation associations in western North America prairies (Stanley and Knopf 2002) and the same pattern has been detected for grasslands in Argentina (Zalba and Cozzani 2004; Aguilar et al. 2008; Cozzani and Zalba 2009; Pretelli et al. 2013), where breeding of birds is favored by a high availability of resources, but, fundamentally, by a lower exposure to predators (Comparatore et al. 1996; Isacch and Martínez 2001; Aguilar et al. 2008).
In this study, we evaluate the response of bird communities in riparian grasslands in the south of the Pampas grassland in Argentina, and also the intensity of egg predation, at sites affected by fire compared to unburnt areas. We expect: (1) that bird communities in habitats affected by fire will exhibit lower values of total abundance, species richness and diversity compared to those associated with streams that were unaffected by fire, (2) that the density of nests and the richness of nesting species will be lower on burnt streams, where the vegetation is lower and open, (3) that the nest predation rate will be higher in the habitats affected by fire, and (4) that these differences will decrease as time passes after the fire and the cover of the upper strata of vegetation and the abundance of tall grasses increase.
Materials and methods
Study area
This study was carried out in grasslands in Sierra de la Ventana, Argentina (38°00′–38°07′S and 61°52′–62°03′W). The climate of the region is temperate, with an average annual temperature of 14 °C and an average annual rainfall of 800 mm (Burgos 1968). The vegetation is represented by a grass steppe (Cabrera 1976; Frangi and Bottino 1995). The limitations for agricultural activity imposed by rocky outcrops and slopes make it a relic of high value for biodiversity conservation (Kristensen and Frangi 1995). It is listed as an Important Bird and Biodiversity Area (IBA) of Argentina due to its ornithological value (Di Giacomo 2005). Tall grasslands associated with the stream banks in these mountains are recognized as preferential sites for the reproduction of grassland birds (Cozzani and Zalba 2009), where the density of vegetation reduces the visibility of nests in the face to potential predators (Zalba and Cozzani 2004; Cozzani and Zalba 2012).
The mountain area includes the Ernesto Tornquist Provincial Park (ETPP), one of the main conservation units of Pampas grassland in Argentina. The reserve faces several conservation challenges including the invasion of exotic woody species, such as Pinus halepensis, Spartium junceum, Genista monspessulana and Prunus mahaleb (Amodeo and Zalba 2013; Cuevas and Zalba 2013; Sanhueza and Zalba 2014), and grazing, mainly by feral horses (Scorolli and López Cazorla 2010). The presence of these large herbivores modifies the plant structure and composition, increasing the shrub density and the cover of bare soil and favouring the dispersal of exotic species (Loydi and Zalba 2009; De Villalobos and Zalba 2010; Loydi et al. 2010). Previous studies in the area have also recorded changes in the composition of the bird community according to the intensity of grazing, the destruction of nests by trampling and an increase in the rate of nest predation due to the reduction of safe nesting sites in overgrazed areas (Zalba and Cozzani 2004; Cozzani and Zalba 2012; Cozzani et al. in preparation). Grazing by feral horses results in a reduction in the percentage of coverage of the highest strata of vegetation which, in turn, is related to decreases in the density, richness and diversity of birds and with changes in the composition of the communities, including a drastic fall in the abundance of species such as the Grassland Yellow-Finch (Sicalis luteola), the Brown-and-Yellow Marshbird (Pseudoleistes virescens), several species of pipits (Anthus spp.) and the Great Pampa-Finch (Embernagra platensis) in areas under intensive grazing (Zalba and Cozzani 2004).
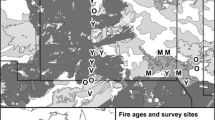
The study area is affected periodically by fires that are particularly frequent during the summer (Michalijos and Uboldi 2010). Between December 29, 2013 and February 2, 2014, the largest fire in the records of the history of the Southern Sierras of Buenos Aires occurred covering 73 thousand hectares (Fig. 1) and comprising almost the entire ETPP, as well as other areas outside the reserve (Marini 2014). This situation was exploited to test the predictions considered in this study. Three streams affected by the fire and three unburnt streams were selected. All of them rise in the mountains and run through riverbeds up to 4 m wide, surrounded by rocks in blocks and soil accumulations that allow the growth of tall grasses, including Paspalum quadrifarium and Cortaderia selloana, which in some places are very dense and can exceed 2 m in height. A belt transect of 400 × 20 m centered on the water courses was established at each stream, so that 10 m wide strips were surveyed at each side of the water course. Starting points were placed at an homogeneous height ranging from 550 to 600 m asl, and the transects were traveled downstream from that point. Vegetation sampling, bird surveys, nest searches and predation trials were conducted in three burned streams and in two stretches of an unburnt stream during the 2014–2015 reproductive season (November to March). The unburnt controls had to be relocated in the 2015–2016 breeding season by others located in two streams of similar characteristics in the following season due to changes in the livestock management of the surrounding fields that altered the structure of the vegetation (Fig. 2).

Study area including Ernesto Tornquist Provincial Park in Buenos Aires province, Argentina. The dark range corresponds to the area burnt in December 2013–February 2014 (73,000 ha)

Streams affected by fire inside the Ernesto Tornquist Provincial Park (1, 2 and 3) in December 2013–February 2014 and control unburnt streams in Estancia “Las Vertientes” (1/2, 3 and 4). a Burnt stream # 2 during the first sampling season (2014–2015). b Burnt stream # 2 during the second sampling season (2015–2016) c Stream not affected by fire used as control during the second sampling season (2015–2016)
Characterization of the vegetation
Four plots of 2 × 2 m were located every 100 m on each sampling strip. Vegetation samples were taken in November, at the beginning of the sampling, and in January and March at the end of the surveys in each season, completing three samples for each stream and season. The vegetation cover percentage was estimated in four height strata (0–15 cm, 15–30 cm, 30–50 cm, and > 50 cm) and the cover of the dominant species: C. selloana, P. quadrifarium, Eryngium spp., Silybum marianum and Echium plantagineum. Mean percentages of vegetation cover between seasons and between burned streams and controls were compared by Student´s t tests. Paired tests were used in the case of interannual comparisons between the same streams.
Bird communities
The transects associated with the stream margins were surveyed once a fortnight during both reproductive seasons, (November to March), completing nine censuses for each transect and season. We recorded the number of individuals of every bird species seen or heard in the strips. Birds that flew over the transects at a height of no more than 2 m, in obvious association with the underlying environment, were also recorded. The values of total density of birds per stream (individuals/ha) were calculated, as well as the richness and diversity of species (Simpson). A generalized linear mixed model (GLMM) was performed to look for possible differences in total density of birds, in species richness and in diversity of species between burned and unburned streams and between seasons. For density of birds we applied aLinear Mixed Model with R package LME4 fit by REML t-testusing Satterthwaite approximations to estimate the degrees of freedom [lmerMod], with Normal distribution, applying the square root of the sample size. The sampling seasons (2014–2015 and 2015–2016) and the treatments (burned streams and controls streams) were considered as fixed effects factors, and sample time(the date on which each sample was taken) and stream (the transect where the sample was taken)as random effects factors. For species richness we performed GLMM with Poisson distribution and log link function, considering the sampling seasons (2014–2015 and 2015–2016) and treatment (burnt or unburnt streams) as fixed effects factors, and sample date and stream as random effects factors. We modeled the diversity through a GLMM with R package LME4 fit by REML t-test use Satterhwaite approximations to estimate the degrees of freedom [lmerMod], with Normal distribution. The response variable, species diversity, was raised to the third power in order to improve homocedasticity and normality. The two sampling seasons (2014–2015 and 2015–2016) and the treatments (burned and controls streams) were treated as fixed effects factors, while sample date and stream were treated as random effects factors. Two samples in which diversity was equal to zero were excluded from the analysis so that the models fulfilled the assumptions. Analyzes were performed using R version 3.4.0 (R Development Core Team 2014).
Nests and associated vegetation
Nests were searched for in the sampling strips along the streams during both breeding seasons. We calculated the mean density of nests (nests/ha) at the burnt and unburnt streams in each season, only considering the bird species that nested in plants or on the ground, and discarding a few cases of birds nesting in scattered exotic trees growing alongside the streams. We compared the density of nests between burnt and unburnt streams, and between seasons, using Mann–Whitney tests. We made estimates of the percentage of vegetation cover by strata of height (0–15 cm, 15–30 cm, 30–50 cm, > 50 cm), and the percentage of bare soil, in plots of 0.5 m2 and 16 m2 centered on each nest (Cozzani and Zalba 2009). Using Student’s t tests, we compared these values at both sampling scales, between burnt streams and controls and between seasons. Paired tests were used in the case of interannual comparisons between the same streams.
Nest predation
To assess the intensity of nest predation, eight artificial nests were used per sampling strip, both on burnt streams and in controls, arranged every 50 m on plants of C. selloana or P. quadrifarium, simulating nests of grassland birds according to De la Peña (2013). The nests were constructed with leaves of grasses, and each one contained a common quail egg (Coturnix coturnix) and another molded in plasticine (Cozzani and Zalba 2012).
The nests were placed in the field during the second week of December in both sampling seasons and were then visited daily for 15 days. They were considered as predated when one or both eggs were broken or taken. We calculated the percentages of total nest predation and the average daily rate of predation by stream and season, calculated as the mean number of days elapsed before each predation event. We performed Student’s t tests to test the differences in the percentages of predation and predation rates between burnt streams and controls, and between seasons. As in the case of natural nests, the percentage of vegetation cover per height stratum (0–15 cm, 15–30 cm, 30–50 cm, > 50 cm) was estimated in plots of 0.5 m2 and 16 m2, centered on the nests, and the plant species present were identified. Linear regression analyzes were used to evaluate the effect of the percentage cover of the two upper layers of vegetation on the fate of the nests. Generalized linear models (GLM) with binomial distribution, backward criterion and logit link function were used to explore the relationship between the survival of artificial nests and the whole set of independent variables. Analyzes were performed using R version 3.4.0 (R Development Core Team 2014).
Results
Characterization of the vegetation
The total vegetation cover at the burnt streams was significantly lower in the first season compared to the second (t = − 4.37, p = 0.001, gl = 8) and also lower than that of the control streams during the first sampling season (t = − 3.19, p = 0.01, gl = 13), but not during the second season (t = 0.70, p = 0.24, gl = 13, Table 1).
Analysis of the changes in the cover of the different height strata showed that, a year after the fire, the burnt streams had high percentages of cover of bare soil and vegetation below 50 cm, except in the case of the #1 burnt stream. On the other hand, the control streams were associated with high percentages of plant cover in the highest stratum (> 50 cm). After 2 years of sampling, the differences between the burnt streams and controls were reduced (Fig. 3).

Mean percentage of bare soil and vegetation cover by height strata on stream banks burnt in December 2013–February 2014 and in unburnt controls in southern Argentinian Pampas, sampled in spring–summer 2014–2015 and 2015–2016. Control streams varied between years
Analysis of the most abundant plant species at the edges of the streams affected by fire showed a reduction in the representation of E. plantagineum and S. marianum, two invasive alien species, which covered up to 40% of the samples in the year of the fire and were no longer present after 2 years of fire, and an increase in the cover of P. quadrifarium (65.2 ± 5.46 vs. 22.6 ± 11) and C. selloana (27.73 ± 2.03 vs. 17 ± 0.43). None of these species showed any variation in abundance between seasons in the case of the streams unaffected by fire.
Bird communities
We detected a total of 44 species of birds during this study associated with the edges of the burnt streams and the controls unaffected by the fire (Table 2), of which 43% were found in both habitats. Four bird species, Great Pampa-Finch (Embernagra platensis), Grassland Yellow-Finch (Sicalis luteola), Tropical Kingbird (Tyrannus melancholicus) and Chalk-browed Mockingbird (Mimus saturninus), were only recorded in the controls and at the burnt streams during the second season, but they were missing from the sites affected by fire in the first season, when the effect of fire was more noticeable.
The GLMM for density of birds did not detect a significant interaction between season and treatment (p = 0.82). The mean density of birds at the edges of burnt streams was lower than that of the controls both in 2014–2015 (20.10 ± 5.22 individuals/ha vs. 42.5 ± 6.35 individuals/ha) and in 2015-2016 (37.77 ± 3.60 individuals/ha vs. 50.06 ± 5.42 individuals/ha).This was in spite of a mean increase of 8.67 ± 4.43 individuals/ha between seasons in burnt streams. Pairwise comparisons showed that, although the difference in the mean density of birds between burned streams and controls is maximum in the first season (2014–2015), it is not statistically significant (t4.3 = − 1.41, p = 0.03), while the density of birds in burned streams is significantly higher in the second breeding season with respect to that immediately following the fire (t37 = 2.86, p = 0.00678).
The GLMM for species richness did not detect a significant interaction between season and treatment (p = 0.27). The random effect associated with the stream was six times greater than that associated with the sample date. Species richness was significantly lower in control with respect to burnt streams (6.0 vs. 10.5, p = 0.036) during the 2014–2015 breeding season, with no significant difference between both treatments the following season (9.1 vs. 11.4, p = 0.39). Species richness increased significantly between seasons in burnt streams (3.56 ± 0.77 species, p < 0.001), with no significant change in the case of the unburnt controls (p = 0.78) (Fig. 4).

Changes in mean richness of bird species on stream banks burnt in December 2013–February 2014, and in the unburnt controls in southern Argentinian Pampas, sampled in spring–summer 2014–2015 and 2015–2016. Unburnt control streams varied between years
We observed increases in the diversity between seasons for the burnt streams (0.65 ± 0.049, in the first season, vs. 0.83 ± 0.013, in the second). Moreover, a year after the fire, the burnt streams presented values of diversity lower than the controls (0.65 ± 0.049 vs. 0.84 ± 0.025), but 2 years after the fire the burnt streams presented values of diversity approximately similar to those of the controls (0.83 ± 0.013 vs. 0.84 ± 0.018). At the same time, notoriously, the #1 burnt stream showed higher values of density, species richness and diversity than the other two streams affected by the fire since the beginning of the samplings, approaching the values recorded for the controls. The GLMM model did not detect a significant interaction between the treatments and the sampling seasons (t4.6 = 1.494, p = 0.1994), while pairwise comparisons showed that species diversity difference in the burned streams burned increased significantly from 2014–2015 to 2015–2016 (t20.48 = 3.51, p = 0.0002).
Nests and associated vegetation
During both sampling seasons, we detected a total of 52 nests corresponding to eleven bird species, of which only six were directly associated with the grassland nesting habitat, building their nests on the ground or on grasses: the Spectacled Tyrant (Hymenops perspicillatus) (25 nests), the Grassland Yellow-Finch (7 nests), the Brown-and-Yellow Marshbird (4 nests), the Great Pampa-Finch, the Rufous-collares Sparrow (Zonotrichia capensis) and the House Wren (Troglodytes aedon) (1 nest each).
The mean density of nests in the burnt streams was lower than in the controls during the first season (3.34 ± 1.81 nests/ha vs. 6.25 ± 1.25 nests/ha), although this difference was not statistically significant (U = 5, p = 0.33, p > 0.05). In the second season, the mean density of nests in burnt and unburnt areas were more similar (5.84 ± 1.50 nests/ha vs. 5.62 ± 1.875 nest/ha, U = 3.5, p = 1, p > 0.05). During this season, the higher density of nests in the burnt streams with respect to the previous year (3.34 ± 1.81 nests/ha vs. 5.84 ± 1.50 nests/ha, U = 2.5, p = 0.5, p > 0.05) was due to a nearly doubling of the nesting density of Spectacled Tyrant (11.25 vs. 6.25 nest/ha), and to the presence of nesting species, such as Grassland Yellow-Finch (3.75 nests/ha) and Brown-and-Yellow Marshbird (1.25 nests/ha), absent in the previous season. In control streams the most abundant nests in both seasons were those of Spectacled Tyrant, Grassland Yellow-Finch, Brown-and-Yellow Marshbird and Great Pampa-Finch.
Nest predation
In the first season the total percentage of nest predation in the burnt streams was 2.07 times higher than in the controls (79% vs. 38%), which was statistically significant (t = 10, p = 0.004, gl = 2) although it stopped being so in the second season (t = 1.96, p = 0.07, gl = 3) when this difference was reduced to 1.6 times (50% vs. 31%). Interannual differences for the burnt streams were also statistically significant (79% in 2014 vs. 50% in 2015, t = 3.3, p = 0.03, gl = 2).
During the first season the nests found in the burnt streams showed survival rates lower than those of the controls (t = − 4.36, p = 0.011, gl = 3) and these differences ceased to be significant 1 year later (t = − 2.80, p = 0.10, gl = 1). No significant interannual differences were detected between the burnt streams, nor between the controls (t = − 1.54, p = 0.13, gl = 2, and t = 0.24, p = 0.41, gl = 2, respectively).
We observed a weak, although statistically significant increasing trend in the probability of success of the artificial nests as the percentage of plant cover corresponding to the strata above 30 cm in height increased at both the smaller scale (R2 = 0.09, p = 0.03) and at the larger scale (R2 = 0.11, p = 0.001) (Fig. 5).

Probability of nest survival in response to the percentage of vegetation cover in surrounding 0.5 m2 (a) and 16 m2 (b) plots on banks of streams burnt in December 2013–February 2014 and in unburnt controls in southern Argentinian Pampas, sampled in spring–summer 2014–2015 and 2015–2016. One represents nest success and 0 nest failure. The total number of successful (Ns) and failed (Nf) nests are indicated for each axis. The grey scale of the diamonds represents the accumulated number of nests
The GLM also suggested an effect of the percentage cover of the upper layers of vegetation on the fate of the artificial nests, with the probability of nest survival showing a significant and positive correlation with the percentage cover of the > 50 cm high vegetation stratum [log (survival prob/prob. of dying) = − 0.721883 + 0.0144% cover of the stratum, p = 0.01).
Discussion
The impact of fires on wild animal communities has been studied very little in South America (Comparatore et al. 1996; Isacch and Martínez 2001; Di Giacomo et al. 2011; Cardoni et al. 2015) despite the predominant role of fire in most of the biomes in the region, especially in shrublands and savannas, and the trends in the alteration of their frequency, extension and intensity. This study represents an original contribution to the knowledge of the effect of fire on grassland bird communities in the south of the Pampas biome, in particular the first data are provided on the variation in species abundance, breeding activity and reproductive success as time elapses after a fire.
Our results revealed a sequence of changes in the structure and composition of the vegetation at stream banks, that constitutes a critical habitat for the nesting of grassland birds (Cozzani and Zalba 2009). One year after the fire there was a predominance of low-lying strata (0–15 and 15–30 cm) dominated by invasive alien species, such as E. plantagineum and S. marianum. This situation was particularly evident in two of the streams affected by the fire, but not in the case of stream #1, which showed an earlier recovery of vegetation, with a higher percentage of cover associated with taller native grasses, such as P. quadrifarium and C. selloana. This difference could respond to topographic variations between the streams: burnt stream #1 has steeper banks that could result in less accessibility for feral horses. Several authors describe the negative effects of large herbivores on the post-fire recovery of vegetation (Bowman and Murphy 2010; Blair et al. 2014; Foster et al. 2016). Studies in Pampean grasslands also emphasize the combined impact of fire and livestock on bird communities, detecting diverse effects depending on the intensity of grazing, the degree of modification of the vegetation structure and the spatial distribution of heterogeneous patches providing key habitats resources for birds (Isacch and Martínez 2001; Cardoni et al. 2015).The presence of large herbivores in the study area could delay the recovery of the vegetation structure and composition by trampling and consumption of native grass sprouts, favouring species of low palatability, including the exotic species better adapted to the conditions of overgrazing (Loydi and Zalba 2009; De Villalobos and Zalba 2010).
Confirming the predictions we propose in the introduction, the abundance, richness and diversity of birds were lower 1 year after the fire for the burnt streams, but increased significantly 2 years after the fire in association with a more complex vegetation structure and an overall interannual increase in the height of the species present. The initial impact of fire on grassland bird communities and their evolution until they reach values of diversity close to or higher than those of unburnt areas has been documented by several authors who agree that the main variable that determines this response of the grassland bird communities is the increase in the structural complexity and diversity of the vegetation on which they depend (Vickery et al. 2005; Zozaya et al. 2010; Watson et al. 2012). Similar associations between vegetation structure and bird communities are reported in other studies carried out in Pampean habitats (Comparatore et al. 1996; Cueto and Lopez de Casenave 1999; Isacch and Martínez 2001). The remarkable recovery of the vegetation after fire, particularly of tall grasses, could provide a greater abundance of food items, availability of refuge and nesting sites, generating a more favourable habitat for the establishment and reproduction of grassland birds.
However, even though the general pattern detected was an increase in the abundance, richness and diversity of birds, differences were observed in the particular response of different species that might be due to their habitat requirements or preferences. These individual differences may be of interest when identifying species that are particularly vulnerable to changes in the frequency or extent of fires. Such are the cases of the Great Pampa Finch, the Brown-and-Yellow Marshbird and the Grassland Yellow-Finch, three species of birds that are strongly associated with grasslands (Codesido et al. 2011), which were absent or only present in very low numbers in the early stages of succession (1 year after fire). Two of these species, the Great Pampa Finch and the Grassland Yellow-Finch, have diets that are based primarily on the consumption of seeds (Salvador and Salvador 1986; Beltzer 1990; Montalti et al. 2005), while the Brown-and-yellow Marshbird feeds primarily on insects (Darrieu et al. 1996). The three, on the other hand, concentrate their breeding activity in the area on patches of tall grasses (Cozzani and Zalba 2009). Subsequent studies could help establish whether their delay in responding to successional changes following a fire is related to the availability of food resources or an appropriate habitat for nesting.
We documented an increase in the nesting density in the burnt streams as time passed after the fire, in association with an increase in the cover of the highest strata of vegetation, confirming one of our predictions. Our results also reveal changes in the nesting bird species, revealing a delay in the response of strict grassland birds which prefer tall plants of P. quadrifarium and C. selloana (Comparatore et al. 1996; Isacch and Martínez 2001; Cozzani 2010) for nesting. Grassland birds would be particularly sensitive to changes produced by the fires, avoiding nesting in the burnt habitats during the initial stages of recovery. This situation may have consequences on the dynamics of their populations, which requires more detailed studies including aspects related to the eventual philopatry and/or displacements of the species, as well as regional patterns of loss and fragmentation of habitats. From the point of view of conservation, and until the possible consequences of this situation in the population dynamics of grassland bird species are better understood, it seems advisable to establish preventive actions, including the creation of firebreaks, in order to protect remnants of nesting habitats in the case of fires over large geographic areas, considering the relictual nature of the natural grasslands under study. These refuges should be burnt in an alternate manner to allow the continuity of the grass fire cycle.
The trials with artificial nests confirmed the expected negative effect of the habitat changes associated with fire on the breeding success of birds. Several authors propose that the differences in the intensity of predation respond to an increase in the detectability of nests as a consequence of changes in the vegetation (Willson et al. 2001; Renfrew et al. 2005; Aguilar et al. 2008; Cozzani and Zalba 2012). Although the visibility of the nests in this study was not measured directly, previous studies carried out in the area showed the relationship between this variable, the density of the highest vegetation strata and the risk of predation (Zalba and Cozzani 2004).Densely vegetated stream margins and low areas appear to be good refuges that could provide a visual and olfactory barrier that reduces the efficiency of predators and they have been shown to be preferential sites for the reproduction of grassland birds (Cozzani and Zalba 2009). Fire would temporarily neutralize this effect of concealment, as indicated by the increase in the predation percentage in the burnt streams, and the inverse relationship between the cover of high plant strata and failure of nests, which could be a selection pressure that explains the behaviour of avoiding these habitats on the part of grassland birds. In any case it is important to highlight that the use of artificial nests as an indicator of real predation rates has been criticized by numerous authors (Zanette and Jenkins 2000; Pärt and Wretenberg 2002; Berry and Lill 2003; Moore and Robinson 2004), who point out that natural and artificial nests may be subject to different predation rates due to variations in the preferences of the eggs by predators or the choice of the place where the nest is placed by the researcher, among other factors. The monitoring of nests by researchers has also been discussed because it could leave odors, mark roads or otherwise alert predators, although most agree that this effect this effect does not usually influence the results (Lloyd et al. 2000; Pärt and Wretenberg 2002). Despite criticism, this technique is a practical and economical tool, and its reliability is supported by numerous studies (Dion et al. 2000; Vander Haegen et al. 2002; Githiru et al. 2005; França and Marini 2009) even in this area of work (Cozzani 2010).
The results obtained in this study highlight the importance of vegetation density, particularly of tall grasses, for sustaining abundant, species-rich, bird communities in habitats on the edges of streams, and for maintaining high values of nest density and reproductive success. It also highlights the ability of the grassland to recover quickly after a fire and the response of the birds that, in this study, took between one and two reproductive seasons, depending on the species. Preliminary evidence of possible interference from grazing by exotic herbivores on the process of recovery of the vegetation and associated bird communities highlights the importance of considering both fire and herbivory when assessing the effect of fires on the biological diversity of the Pampas grasslands. The temporary exclusion of large herbivores, including cattle and feral horses, is a management measure that could accelerate the recovery of vegetation affected by fire and could be applied in stream sections in order to generate refuges for reproduction of the most vulnerable birds, without interfering with access to water by wildlife in the rest of the watercourse. The effect of fire should also be analyzed on a larger scale in order to evaluate the impact of the total reduction of the area of grassland that is in good condition, and other factors, such as the fragmentation of habitats that could enhance the effect of temporary successional loss of habitats of high value for nesting on the population dynamics of birds, especially of the obligate grassland species.
Despite the importance of fire as a structuring factor of natural communities in grasslands, management policies in protected natural areas of southern South America, in most cases, are limited to preventing and extinguishing fires. Important advances in the understanding of the effects of fire on natural communities have been made, however an important level of uncertainty still persists, especially in dynamic scenarios of loss and fragmentation of habitats, advance of invasive alien species and climate change (D´ Antonio and Vitousek 1992; Mouillot and Field 2005; Williams and Zdravko 2000). An approach based on the adaptive management of prescribed and natural fires seems the best alternative to preserve the structure and composition of plant communities and grassland birds.
References
Aguilar TM, Dias RI, Oliveira AC, Macedo RH (2008) Nest-site selection by Blue-black Grassquits in a Neotropical savanna: do choices influence nest success? J Field Ornithol 79:24–31
Amodeo MR, Zalba SM (2013) Wild cherries invading natural grasslands: unraveling colonization history from population structure and spatial patterns. Plant Ecol 214:1299–1307
Askins RA, Chávez-Ramírez F, Dale BC, Haas CA, Herkert JR, Knopf FL, Vickery PD (2007) Conservation of grassland birds in North America: understanding ecological processes in different regions. Ornithol Monogr 64:1–46
Azpiroz AB, Isacch JP, Dias RA, Di Giacomo SA, Suertegaray Fontana C, Morales Palarea C (2012) Ecology and conservation of grassland birds in southeastern South America: a review. J Field Ornithol 83:217–246
Barlow J, Peres CA, Henriques LMP, Stouffer PC, Wunderle JM (2006) The responses of understorey birds to forest fragmentation, logging and wildfires: an Amazonian synthesis. Biol Conserv 128:182–192
Barnett JM, Pearman M (2001) Listacomentada de las avesargentinas: annotated checklist of the birds of Argentina. Lynx, Barcelona
Beltzer A (1990) Biología alimentaria del verdon común Embernagraplatensisplatensis (Aves: Emberizidae) en el valle aluvial del rio Parana medio, Argentina. Ornitol Neotrop 1:25–30
Bendell JF (2012) Effects of fire on birds and mammals. Cap. 4 en. In: Kozlowski T, Ahlgren C (eds) Fire and ecosystems. Academic Press, New York
Berry L, Lill A (2003) Do predation rates on artificial nests accurately predict predation rates on natural nests? The effects of nest type, egg type and nest-site characteristics. Emu 103:207–214
Blair JM, Nippert J, Briggs J (2014) Grassland ecology. In: Monson R (ed) The plant sciences—ecology and the environment. Springer, New York, pp 389–423
Bond WJ, Woodward FI, Midgley GF (2004) The global distribution of ecosystems in a world without fire. New Phytol 165:525–538
Bonier F, Martin PR, Wingfield JC (2007) Urban birds have broader environmental tolerance. Biol Lett 3(6):670–673
Bowman DMJS, Murphy PB (2010) Fire and biodiversity. In: Sodhi NS, Ehrlich PR (eds) Conservation biology for all. Oxford University Press, Oxford, pp 163–180
Burgos J (1968) El clima de la provincia de Buenos Aires en relación con la vegetación natural y el suelo. In: Cabrera AL (ed) Flora de la provincia de Buenos Aires, vol 1. Colección científica INTA, Buenos Aires, pp 33–100
Cabrera AL (1976) Regiones Fitogeográficas Argentinas. In: Parodi LR (ed) Enciclopedia Argentina de Agricultura y Ganadería, Tomo 2, Fascículo 2. ACME, Buenos Aires, p 85
Cardoni DA, Isacch JP, Iribarne O (2015) Avian responses to varying intensity of cattle production in Spartinadensiflora saltmarshes of south-eastern South America. Emu 115:12–19
Clavero M, Brotons L, Herrando S (2011) Bird community specialization, bird conservation and disturbance: the role of wildfires. J Anim Ecol 80:128–136
Codesido M, González Fischer C, Bilenca D (2008) Asociaciones entre diferentes patrones de uso de la tierra y ensambles de aves en agroecosistemas de la Región Pampeana, Argentina. Ornitol Neotrop 19:575–585
Codesido M, González-Fischer C, Bilenca D (2011) Cambios en la distribución de especies de aves terrestres en agroecosistemas de la Argentina Central. El cóndor 113:266–273
Comparatore VM, Martinez MM, Vasallo AI, Barg M, Isacch JP (1996) Abundancia y relaciones con el hábitat de aves y mamíferos en pastizales de Paspalumquadrifarium(paja colorada) manejados con fuego (Provincia de Buenos Aires, Argentina). Interciencia 21:228–237
Coppedge BR, Fuhlendorf SD, Harrell WC, Engle DM (2008) Avian community response to vegetation and structural features in grasslands managed with fire and grazing. Biol Conserv 141(5):1196–1203
Costanza R, D’Arge R, De Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O’Neill RV, Paruelo J, Raskin RG, Sutton P, Van den Belt M (1997) The value of the world’s ecosystem services and natural capital. Nature 387:253–260
Cozzani NC (2010) Efectos del pastoreo sobre el éxito de cría de aves de pastizal pampeano. Tesis doctoral, Universidad Nacional del Sur. 156
Cozzani N, Zalba SM (2009) Estructura de la vegetación y selección de hábitats reproductivos en aves del pastizal pampeano. Ecol Austral 19:35–44
Cozzani N, Zalba SM (2012) Depredadores de nidos en pastizales del Parque Provincial Ernesto Tornquist (Provincia de Buenos Aires, Argentina), su importancia relativa bajo distintas intensidades de pastoreo. El Hornero 27(2):137–148
Cueto VR, Lopez de Casenave J (1999) Determinants of bird species richness: role of climate and vegetation structure at a regional scale. J Biogeogr 26:487–492
Cuevas YA, Zalba SM (2013) Efecto del tipo de corte y de tratamientos en el mantillo para la restauración de pastizales naturales invadidos por Pinus halepensis. Boletín de la Sociedad Argentina de Botánica. https://doi.org/10.31055/1851.2372.v48.n2.6265
D´ Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu Rev Ecol Syst 23:63–87
Darrieu CA, Camperi AR, Cicchino AC (1996) Ecologíaalimentaria de Pseudoleistesvirescens (Vieillot 1819) (Aves, Icteridae), enambientesribereñosdelnordeste de la provincia de Buenos Aires, Argentina.Serie Informe No 51. Comisión de Investigaciones Científicas de la provincia de Buenos Aires. La Plata, Argentina, pp 14
De la Peña M (2013) Nidos y reproducción de las aves argentinas. Serie Naturaleza, Conservación y Sociedad. de La Vega SG, Carrizo G, Chaippe A. 2003. Patagonia Las Leyes de La Estepa, Contacto Silvestre
De Villalobos AE, Zalba SM (2010) Continuous feral horse grazing and grazing exclusion in mountain pampean grasslands in Argentina. Acta Oecol 36(5):514–519
Di Giacomo AS (2005) Áreas Importantes para la Conservación de las Aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad. Temas de naturaleza y conservación 5:1–514. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires: Edición del autor
Di Giacomo AG, Di Giacomo AS, Reboreda JC (2011) Effects of grassland burning on reproductive success of globally threatened Strange-tailed Tyrants Alectrurus risora. Bird Conserv Int 21(4):411–422
Dion N, Hobson KA, Larivière S (2000) Interactive effects of vegetation and predators on the success of natural and simulated nests of grassland songbirds. Condor 102:629–634
Eldridge DJ, Bowker MA, Maestre FT, Roger E, Reynolds JF, Whitford WG (2011) Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol Lett 14:709–722
Foster CN, Barton PS, Sato CF, Wood JT, MacGregor CI, Lindenmayer DB (2016) Herbivory and fire interact to affect forest understory habitat, but not its use by small vertebrates. Anim Conserv 19:15–25
França LC, Marini MÂ (2009) Teste do efeito de borda na predação de ninhos naturais e artificiais no Cerrado. Zoología 26(2):241–250
Frangi JL, Bottino OJ (1995) Comunidades vegetales de la Sierra de la Ventana, Provincia de Buenos Aires, Argentina. Rev Fac Agron La Plata 71:93–133
Fuhlendorf SD, Harrell WC, Engle DM, Hamilton RG, Davis CA, Leslie DM Jr (2006) Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 16:1706–1716
Githiru M, Lens L, Cresswell W (2005) Nest predation in a fragmented Afrotropical forest: evidence from natural and artificial nests. Biol Conserv 123:189–196
Homan P (2012) Survival and recolonisationfollowing wildfire at Moyston West, Western Victoria: 1. Mammals. Vic Nat 129:192–202
Homan P (2014) Survival and recolonisation following wildfire at Moyston West, Western Victoria. 2. Herpetofauna. Vic Nat 131:4–14
Isacch JP, Martínez MM (2001) Estacionalidad y relaciones con la estructura del habitat de la comunidad de aves de pastizales de paja colorada (Paspalumquadrifarium) manejados con fuego en la provincia de Buenos Aires, Argentina. Ornitol Neotrop 12:345–354
Kiss L, Magnin F (2006) High resilience of Mediterranean land snail communities to wildfires. Biodivers Conserv 15:2925–2944
Krapovickas S, Di Giacomo AS (1998) Conservation of pampas and campos grasslands in Argentina. Parks 8:47–53
Kristensen MJ, Frangi JL (1995) La Sierra de la Ventana: una Isla de Biodiversidad. Ciencia Hoy 5:25–34
Langlands PR, Brennan KEC, Pearson DJ (2006) Spiders, spinifex, rainfall and fire: long-term changes in an arid spider assemblage. J Arid Environ 67(1):36–59
Leys B, Marlon JR, Umbanhowar C, Vannière B (2018) Global fire history of grassland biomes. Ecol Evol 8(17):8831–8852. https://doi.org/10.1002/ece3.4394
Lloyd P, Little RM, Crowe TM (2000) Investigator effects on the nesting success of arid-zone birds. J Field Ornithol 71(2):227–235
Loydi A, Zalba SM (2009) Feral horses dung piles as invasion windows in natural grasslands. Plant Ecol 201(2):471–480
Loydi A, Distel RA, Zalba SM (2010) Large herbivore grazing and non-native plant invasions in montane grasslands of central Argentina. Nat Areas J 30(2):148–155
Lyon LJ, Huff MH, Telfer ES, Schreiner DS, Smith L, Kapler J (2000) Fire effects on animal populations. In: Smith JK (ed) Wildland fire in ecosystems. Effects of fire on Fauna. Rocky Mountain Research Station, U.S. Department of Agriculture, Forest Service, Missoula
Marini F (2014) Informe complementario acerca del incendio en la comarca serrana del sudoeste de la Provincia de Buenos Aires utilizando imágenes satelitales. Teledetección y SIG - SSD. INTA
Meinrat A, Goldammer JG (1992) Tropical wildland fires and other biomass burning: Environmental impacts and implications for land use and fire management. Conservation of West and Central African Rainforests, pp 71–110
Michalijos MP, Uboldi J (2010)Caracterización del régimen de fuego en el parque provincial Ernesto Tornquist, provincia de Buenos Aires. Tecnologías de la Información Geográfica del sur argentino
Milesi FA, Marone L, Lopez de Casenave J, Cueto VR, Mezquida ET (2002) Gremios de manejo como indicadores de las condiciones del ambiente: un estudio de caso con aves y perturbaciones del hábitat en el Monte central, Argentina. Ecol Austral 12(2):149–161
Montalti D, Ferman LM, Camperi AR, Soave GE, Arambarrim AM, Darrieu CA (2005) Winter diet of great pampa-finches Embernagra platensis in Guaminí Lagoon, Argentina. Acta Ornithol 40:79–82
Moore RP, Robinson WD (2004) Artificial bird nests, external validity, and bias in ecological field studies. Ecology 85(6):1562–1567
Mouillot F, Field CB (2005) Fire history and the global carbon budget: A 1ox 1o fire history reconstruction for the 20th century. Glob Change Biol 11:398–420
Pärt T, Wretenberg J (2002) Do artificial nests reveal relative nest predation risk on real nests? J Avian Biol 33(1):39–46
Pausas JG, Parr CL (2018) Towards an undertanding of the evolutionary role of fire in animals. Evol Ecol 32:113–125
Pennington RT, Hughes CE (2014) The remarkable congruence of New and Old World savanna origins. New Phytol 204:4–6
Pons P, Clavero M (2010) Bird responses to fire severity and time since fire in managed mountain rangelands. Anim Conserv 13:294–305
Pretelli MG, Isacch JP, Cardoni DA (2013) Year-round abundance, richness and nesting of the bird assemblage of tall grasslands in the south-east pampas region, Argentina. Ardeola 60:327–343
R Development Core Team (2014) R: a language and environment for statistical computing. R foundation for Statistical Computing, Vienna, Austria
Ratajczak Z, Nippert JB, Briggs JM, Blair JM (2014) Fire dynamics distinguish grasslands, shrublands and woodlands as alternative attractors in the Central Great Plains of North American. J Ecol 102:1374–1385
Recher HF, Lunney D, Matthews A (2009) Small mammal populations in a eucalypt forest affected by fire and drought. I. Long-term patterns in an era of climate change. Wildl Res 36:143–158
Renfrew RB, Ribic CA, Nack JL (2005) Edge avoidance by nesting grassland birds: a futile strategy in a fragmented landscape. Auk 122(2):618–636
Sackmann P, Farji-Brener AA (2006) Effect of fire on ground beetles and ant assemblages along an environmental gradient in NW Patagonia: does habitat type matter? Ecoscience 13:360–371
Salvador SA, Salvador LA (1986) Nota sobre la reproducción del Misto (Sicalisluteola) en Córdoba, Argentina. Hornero 12:274–280
Sanhueza CC, Zalba SM (2014) Banco de semillas, germinación y longevidad de semillas de retama (Spartiumjunceum, Fabaceae): implicancias para su control. Boletín de la Sociedad Argentina de Botánica 49(1):67–76
Scorolli AL, López Cazorla AC (2010) Demography of feral horses (Equuscaballus): a long-term study in Tornquist Park, Argentina. Wildl Res 37:207–214
Stanley TR, Knopf FL (2002) Avian responses to late-season grazing in a shrub-willow floodplain. Conserv Biol 16:225–231
Ukmar E, Battisti C, Luiselli L, Bologna MA (2007) The effects of fire on communities, guilds and species of breeding birds in burnt and control pinewoods in central Italy. Biodivers Conserv 16:3287–3300
Vander Haegen WM, Schroeder MA, DeGraaf RM (2002) Predation on real and artificial nests in shrubsteppe landscapes fragmented by agriculture. Condor 104:496–506
Vickery PD, Tubaro PL, Cardoso Da Silva JM, Peterjohn BG, Herkert JR, Cavalcanti RB (1999) Conservation of grassland birds in the Western Hemisphere. Stud Avian Biol 19:2–26
Vickery PD, Zuckerberg B, Jones AL, Shriver WG, Weik AP (2005) Influence of fire and other anthropogenic practices on grassland and shrubland birds in New England. USDA Forest Service Gen. Tech. Rep. PSW-GTR-191
Watson SJ, Taylor RS, Nimmo DG, Kelly LT, Clarke MF, Bennett AF (2012) The influence of unburnt patches and distance from refuges on post-fire bird communities. Anim Conserv 15:499–507
Williams DG, Zdravko B (2000) African grass invasion in the Americas: ecosystem consequences and the role of ecophysiology. Biol Invasions 2:123–140
Willson M, Morrison JL, Sieving KE, De Santo TL, Santisteban L, Díaz I (2001) Patterns of predation risk and survival of birds nests in a chilean agricultural landscape. Conserv Biol 15(2):447–456
Zalba SM, Cozzani N (2004) The impact of feral horses on grassland bird communities. Anim Conserv 7(1):35–44
Zanette L, Jenkins B (2000) Nesting success and nest predators in forest fragments: a study using real and artifical nests. Auk 117(2):445–454
Zavaleta ES, Pasari JR, Hulvey KB, Tilman GD (2010) Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. PNAS 107(4):1443–1446
Zozaya EL, Brotons L, Herrando S, Pons P, Rost J, Clavero M (2010) Monitoring spatial and temporal dynamics of bird communities in Mediterranean landscapes affected by largewildfires. Ardeola 57:33–50
Acknowledgements
This study was supported by the Universidad Nacional del Sur, CONICET and CIN (Consejo Interuniversitario Nacional). The authors wish to thank Parque Provincial Ernesto Tornquist and Estancia Las Vertientes for their help during fieldwork and Valdemar Delhey for his valuable collaboration in statistical analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Ailsa J. McKenzie.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bahía, R., Zalba, S. Changes in grassland bird communities and breeding success after a fire in the Argentinian Pampas. Biodivers Conserv 28, 3767–3786 (2019). https://doi.org/10.1007/s10531-019-01850-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-019-01850-4