
Abstract
The ‘spectral variation hypothesis (SVH)’ assumes spectral variability as a result of variation in species richness. In the present study, we explore the potential of satellite datasets in identifying the patterns in species richness in part of three global biodiversity hotspots falling in India viz., Himalaya, Indo-Burma, and Western Ghats. We used generalized linear models to correlate remote sensing based vegetation indices (VIs) and physiographic indices (PIs) with plant richness calculated using 1264, 1114, and 1004 field plots across 21 different forest vegetation classes in Himalaya, Indo-Burma, and Western Ghats respectively. Three different vegetation indices ranked highest in explaining the variance in plant richness in the three hotspots. The variance in species richness explained by models based on only VIs was highest (69%, P < 0.01) for Bamboo vegetation in Indo Burma hotspot with Normalized Difference Vegetation Index, followed by that for dry deciduous forest in Western Ghats (57%, P < 0.001) with Normalized Difference Water Index, and for grasslands (54%, P < 0.05) in Himalaya by Modified Soil Adjusted Vegetation Index. The explained variance increased with combined models that are based on PIs and VIs to up to 85% (P < 0.05). Overall, we observed very high correlation between VIs and plant richness in open canopy vegetation classes with low species richness such as grasslands, scrubs, and dry deciduous forests, followed by vegetation classes with moderately dense canopy. Our study provides crucial insights on utility of satellite datasets as a proxy for estimating plant richness for better conservation of diverse ecosystems.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Various studies have used satellite data observations to explore patterns of plant richness (Gillespie 2005; Viedma et al. 2012). Vegetation indices have been utilized on a wide range of spectral and temporal scales to understand the ‘spectral variation hypothesis’, which assumes spectral variability as a result of variation in plant diversity (Rocchini et al. 2007; Gaitán et al. 2013). In tropical forests, normalized difference vegetation index (NDVI) has been shown to increase with diversity, explaining moderate range of variance i.e., 30–60%, nonetheless the mechanisms driving the relationship between vegetation indices and species richness are not well established. Integration of climatic and physiographic variables along with spectral indices in the model could better explain the variance in plant richness (Levin et al. 2007; Pau et al. 2012). Satellite-derived vegetation indices provide clues about diversity patterns as they are used for productivity estimation and quantification of spatial heterogeneity of vegetation (Oindo and Skidmore 2002). The spectral variation of remotely sensed data is associated with the heterogeneity in the environment which may, in turn, be correlated with species diversity (i.e., the Spectral Variation Hypothesis (SVH). Large sites are likely to be more heterogeneous than small sites (Öster et al. 2007), therefore the spectral variation measured over entire sites can be expected to be associated with the area of the sites. The variability in observations from satellites with broad spectral bands has been shown to function as a useful predictor of species diversity at local scales in a variety of vegetation types and geographic areas (Rocchini et al. 2007) and also for estimating levels of beta diversity. Although much progress has been made in estimating species diversity from satellite data (Gillespie et al. 2008), it has been concluded that more empirical studies are needed in order to establish the potential of remote sensing data in biodiversity studies (Nagendra et al. 2010).
There is an increased interest in measuring and modelling biodiversity using remote sensing datasets (Nagendra 2001). Biodiversity is a multifaceted variable; therefore it can be difficult to measure and express it simply (Duro et al. 2007). Biogeographers are particularly interested in measuring or quantifying patterns of species’ diversity, distribution, movements, and in modelling or providing probability maps of species distributions and patterns of diversity. Remote sensing datasets have considerable potential as a source of information on biodiversity at various spatial and temporal scales (Willis and Whittaker 2002). Remote sensing offers an inexpensive means of deriving complete spatial coverage of environmental information for large areas in a consistent manner that may be updated regularly (Duro et al. 2007). Direct remote sensing approaches use space borne sensors to identify either species or land cover types, and directly map the distribution of species assemblages, whereas indirect approaches use space borne sensors to model species distributions and the distributions of species diversity. Both these approaches have significant applications for species and ecosystem conservation that have still not been completely utilized to their full potential. Alternatively, a direct relationship between measures of species diversity and spectral variation has been sought. Normalized difference vegetation index (NDVI) from passive sensors has been the most famous index, because it is easy to calculate using the red (R) and near infrared (NIR) bands common to almost all passive space borne sensors (Gillespie 2005). NDVI has been associated with net primary productivity and has been hypothesized to quantify species diversity based on species energy theory (Currie 1991). Many studies have reported significant positive correlations between plant species diversity from plot or regions and NDVI in both temperate (Fairbanks and McGwire 2004; Levin et al. 2007) and tropical ecosystems (Cayuela et al. 2006). NDVI can explain between 30% and 87% of the variation in species richness or diversity within a vegetation type, landscape, or region (Pouteau et al. 2018). Heterogeneity in land-cover types, spectral indices, and spectral variability derived from satellite imagery has also been correlated with species diversity (Rocchini 2007). This is largely based on the hypothesis that heterogeneity in land cover, spectral indices, or spectral variability within an area or landscape acts as an indicator of habitat heterogeneity, which allows more species to coexist and hence greater species diversity (Simpson 1949; Carlson et al. 2007; Rocchini et al. 2007). Variation in spectral indices has been shown to be positively associated with species richness and diversity for a number of taxa in different regions. The ability to provide complete data coverage for large areas is often seen as a major advantage of remote sensing; some problems of working with large areas have not been addressed. It is generally assumed that relationships between the biodiversity variable of interest and the remotely sensed response are spatially stationary and hence transferable between sites within the region of study.
The ability to provide complete data coverage for large areas is often seen as a major advantage of remote sensing. It is generally assumed that relationships between the biodiversity variable of interest and the remotely sensed response are spatially stationary and hence transferable between sites within the region of study. There has been an increase in sophisticated statistical and spatial analyses to study plant SR. The prediction of richness has substantially relied on simple univariate regression or multiple regression models appropriately scaling sensor imagery to field data on vascular plants (Gould 2000). While these approaches provide a basic understanding of patterns and can be used to create predictive richness maps for a landscape, region, or continent, more sophisticated techniques are being examined and developed to model patterns of richness (Foody 2004). General linear models and general additive models have become increasingly important in the spatial prediction of biodiversity patterns; however, they have been less used for exploring the relationship between remote sensing data and ground based plant richness (Schwarz and Zimmermann 2005).
Our general hypothesis is that SR at a coarse scale is a function of the area and the environmental heterogeneity. Physiography and anthropogenic disturbance are local parameters, in order to take into account both meso- and micro-scale effects. Spatial heterogeneity may contribute to the control of local plant SR, thus are mainly related to gap dynamics (Pausas 1994). Increased heterogeneity implies diversified niches, thus more species coexistence. Often some species might be favored by relatively uniform or stable habitats, while others by heterogeneous habitats. As the size of the sites under study increases, within site topographic complexity becomes an important determinant of plant richness (O’brien et al. 2000). Studies with large data sets have shown the importance of the interaction between temperature and rainfall for explaining SR (Austin et al. 1996).
In this study, we test following two hypotheses in total 21 different vegetation types from three global biodiversity hotspots falling within India viz., Himalaya, Indo-Burma, and Western Ghats using open source satellite datasets and vegetation sampling data.
We undertake the present study based on following hypotheses:
-
(a)
Plant richness combined models based on vegetation indices and physiographic indices perform better than individual index based models in explaining the variation in plant richness.
-
(b)
Variation in plant richness explained by spectral indices varies across plant life forms with maximum variation explained for trees followed by shrubs and herbs.
Materials and methods
Study area
The study area comprises of three global biodiversity hotspots falling in India viz., Himalaya, Indo-Burma (only the mainland), and Western Ghats. Himalaya hotspot, hereafter called as “Himalaya” is located between 25°39′28″ and 35°49′48″N latitude; and between 73°08′04″ and 97°24′44″E longitude spanning over ca. 329, 109 km2 (Fig. 1). The elevational gradient in Himalaya ranges from 500 to 8800 m and distinct climatic gradients can be observed in this hotspot, where dry and cold climate is prevalent in western Himalaya with mean annual temperature of ca. 5 °C and total annual precipitation of 2500 mm/year; while wet and cold conditions are observed in eastern Himalaya with mean annual temperature of ca. 10 °C and total annual precipitation of 4000 mm/year. Owing to dry climatic conditions, scrubs and grasslands are prevalent in western Himalaya; while moist deciduous and sub-tropical broadleaved forests are dominant in eastern Himalaya. Indo-Burma hotspot, hereafter called “Indo-Burma” covers ca. 121,721 km2 and is located between 10°30′34″ and 26°55′50″N latitude; and 89°51′16″ and 95°22′48″E longitude (Fig. 1). The climate is wet and humid with mean annual temperature of ca. 25 °C and very high total annual precipitation of 6000 mm/year. The elevational gradient ranges from 750 to 2300 m above mean sea level and exhibits a wide variety of ecosystems ranging from mixed wet evergreen, dry evergreen, deciduous, and montane forests. Western Ghats hotspots is located between 8°04′45″ and 22°01′40″N latitude; 72°38′34″ and 78°28′18″E longitude (Fig. 1). In general, the mean temperature of the coldest month ranges from 25 °C at sea level to 11 °C at 2400 m. The elevation gradient ranges from 300 to 2700 m with increasing trend from north to south in the hotspot. The wide variation of rainfall patterns in the Western Ghats, coupled with the region’s complex geography, produces a great variety of vegetation types (Myers et al. 2000).

Three global biodiversity hotspots falling in India: a Himalaya, b Indo-Burma, c Western Ghats. (Color figure online)
Data used
Satellite data
We used multi-season multispectral satellite imagery of Landsat Thematic Mapper of year 2010 with spatial resolution of 30 m and a total of 3382 locations of field samples from all three hotspots to generate four most widely used spectral vegetation indices (VIs); Normalized Difference Vegetation Index (NDVI), Enhanced Vegetation Index (EVI), Modified Soil Adjusted Vegetation Index (MSAVI2, Qi et al. 1994), and Normalized Difference Water Index (NDWI, Gao 1996) (Supplementary Information) for a total of 21 vegetation classes. We also calculated and extracted the physiographic indices (PIs) such as altitude, slope, and aspect using Shuttle Radar Topographic Mission (SRTM) satellite data with spatial resolution of 30 m.
Plant richness data
The field samples used in our analysis were laid during ‘biodiversity characterization at landscape level project’ conducted for whole India during 1998 to 2008. As a part of the project stratified random sampling based 15, 565 nested quadrates of 400 m2 were laid in different natural vegetation classes in India (Roy et al. 2012, 2015), which also covered the forests in the three biodiversity hotspots covered in the present study. A random distribution of sample points was chosen in the vegetation type strata to determine the type-specific relative species composition. Minimum sampling intensity of 0.001–0.002% was selected on the basis of the remote sensing based vegetation type strata along with the physiography and climatic zones. This sampling intensity was selected so as to optimize the available resources and time, given the forest vegetation cover and other characteristics of the eco-regions in India. Species richness (Number of species) of different lifeforms (Tree, Shrub, Herb) was calculated based on 1264 plots from Himalaya, 1004 plots from Western Ghats and 1114 plots from Indo-Burma making to a total of 3382 field plots.
Testing spectral variation hypothesis
‘Spectral variation hypothesis’ (SVH) was tested using generalized linear models (GLMs) to understand the relationship between spectral indices and plant richness across different life forms viz., herbs, shrubs, trees, as well as of whole plot. We created generalized linear models in R software based on single variable as well as multiple variables (R Development Core Team 2014) using spectral vegetation indices (VIs) and physiographic indices (PIs) to test our hypotheses.
Results
Patterns of spectral reflectance in three biodiversity hotspots
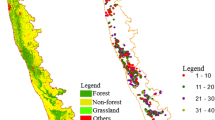
We observed varied patterns of spectral reflectance of the 21 vegetation types falling in the three hotspots. In Himalaya, the spectral reflectance ranged from 7 to 45%, where highest reflectance (~ 45%) observed in moist deciduous forests while lowest reflectance (~ 8%) was observed in grasslands. In Indo Burma, the spectral reflectance ranged from 9 to 45%, with highest reflectance (~ 45%) in evergreen forests and lowest reflectance (~ 9%) in scrubs and dry deciduous forests. In Western Ghats, the reflectance ranged from 9 to 33%, with highest reflectance (32%) in evergreen forests and lowest (9%) in forest plantations. Same vegetation types e.g., grasslands, scrublands, and dry deciduous forests that occurred in all three hotspots depicted different patterns of spectral reflectance, which could be attributed to local conditions that include climate, composition of these vegetation types, and anthropogenic disturbance (Fig. 2).

The spectral reflectance curve of 21 vegetation types from three biodiversity hotspots a Himalaya, b Indo-Burma, and c Western Ghats. (Color figure online)
Spectral variation and species richness relationship
The variance in species richness explained by models based on vegetation indices only (VIs) was highest (69%, P < 0.01) in Bamboo vegetation in Indo Burma hotspot by NDVI, followed by that for dry deciduous forest in Western Ghats (57%, P < 0.001) by NDWI, and grasslands (54%, P < 0.05) in Himalaya by MSAVI (Figs. 3, 4, 5). Additionally, integration of physiographic indices (PIs) with VIs increased the explained variance in SR to up to 85% (P < 0.05) by MSAVI + Aspect for grasslands in Himalaya, followed by 80% (P < 0.01) for scrubland in Western Ghats by NDVI + altitude + slope, and 78% (P < 0.01) for grasslands in Indo-Burma by NDVI + altitude (Equations). The variance explained by different models varied according to the predictor variables as well as the type of life form e.g., tree, shrub, herb etc. Variance explained in plant richness in trees was the highest in Pine forests in Himalaya (83%, P < 0.05, NDWI + altitude), followed by degraded forests in Indo- Burma (78%, P < 0.01, NDVI + altitude), and semi-evergreen forests in Western Ghats (76%, P < 0.01, NDVI + altitude + slope). The variance explained in shrub and herb species richness was low in most of the vegetation classes, because these life forms remain as understory in multi-canopy complex forests and are not directly visible to the satellite sensors. The variance explained only based on VIs was 54% in grasslands for shrub richness in Himalaya, which increased to 85% after we added physiography indices as a predictor variables.

The variance explained in life form wise species richness in seven vegetation classes explained in Himalaya by vegetation index based models and combined models comprising of vegetation index and physiographic index. (Color figure online)

The variance explained in life form wise species richness in seven vegetation classes explained in Indo Burma by vegetation index based models and combined models comprising of vegetation index and physiographic index. (Color figure online)

The variance explained in life form wise species richness in seven vegetation classes explained in Western Ghats by vegetation index based models and combined models comprising of vegetation index and physiographic index. (Color figure online)
Equations: The models that explained highest variance in plant species richness in the three biodiversity hotspots.
For Himalaya:
For Indo-Burma:
For Western Ghats:
In Himalaya, the variation in overall species richness (SR) was best explained in grasslands (54%, P < 0.05, MSAVI + aspect) and dry deciduous forests (53%, P < 0.01, NDWI + altitude), which could be attributed to open canopy, low moisture levels, and simpler canopy structure, as compared to other vegetation classes (Fig. 3). The variation in SR in moisture rich diverse ecosystems such as moist deciduous forests was poorly explained (41%, P < 0.05, NDWI + altitude) by the spectral vegetation indices, because of higher moisture levels, and complex canopy structure with multilayer canopy cover. The variance in SR in Pine, Pine mixed forests and scrubs was moderately explained (42%, P < 0.05; 51%, P < 0.01; and 45%, P < 0.01 respectively) by vegetation indices. However, the explained variance significantly increased after addition of physiographic variables such as altitude, slope and aspect to vegetation index based models. Highest increase in the variance explained was observed in grasslands and dry deciduous forests, where the variation in SR explained by combined model with vegetation index and physiography index (VI + P) was 85%, P < 0.01; and 84%, P < 0.05 respectively. In Indo-Burma, the variation in overall species richness (SR) was best explained for bamboo vegetation (69%, P < 0.01) followed by evergreen forests (63%, P < 0.05), which could be attributed to dominance of few plant species in the top canopy in Bamboo vegetation, as compared to other vegetation classes (Fig. 3). Similar to other hotspots, addition of physiographic variables to vegetation index based models increased the predictive capacity of the models by a large amount. Highest increase in the explained variance due to addition of physiography to spectral index based models was observed in grasslands and degraded forests, where the variation in SR explained increased from 47% to 78% and 42% to 75% respectively. In Western Ghats, the variation in overall species richness was best explained in dry deciduous forests (57%, P < 0.01) followed by semi-evergreen forests (56%, P < 0.05) and grasslands (52%, P < 0.05), which could be attributed relatively open canopy in these vegetation classes as compared to other dense vegetation types (Fig. 4). Highest variance in tree species richness was explained in scrub (61%, P < 0.01), which has sparse distribution of trees, followed by plantations (55%, P < 0.01), which has less species richness due to monotonous composition.
Discussion and conclusions
We observed highest spectral reflectance among all the vegetation types in moist deciduous forests in Himalaya and evergreen forests in Indo Burma hotspot, whereas lowest reflectance was observed in grasslands, scrublands, and dry deciduous forest. This could be attributed to canopy structure, species richness, and local conditions such as microclimate, species composition, and anthropogenic disturbance. In addition, we found highest range of spectral reflectance in Himalaya, among the three hotspots that could be pertinent to larger gradient of species richness due to large climatic variation in western and eastern Himalaya, which is line with the results of Tripathi et al. (2017). In Himalaya, highest reflectance was observed in moist deciduous forest, followed by temperate coniferous, and pine mixed forest, while among the lowest reflectance classes were grassland, scrubs, and dry deciduous forest. This could be clearly attributed to the canopy structure in terms of presence of trees in top canopy, canopy density etc. In Indo-Burma, we observed highest reflectance in evergreen forest followed by moist deciduous forest and grasslands. This could be due to dense canopy structure and high species richness in these vegetation types, which is peculiarly visible in pristine ecosystems in Indo-Burma hotspot. Lowest reflectance was observed in scrub, degraded forest, and bamboo, which could be attributed to less dense canopy, degraded patches of forests, and monotonous vegetation in case of bamboo vegetation. In Western Ghats, highest reflectance was observed in evergreen followed by semi-evergreen, and moist deciduous forest, which is pertinent to dense canopy and high species richness, whereas lowest reflectance was observed in dry deciduous forest, forest plantation, and scrub. The reason behind same vegetation types e.g., grasslands, scrublands, and dry deciduous forests depicting different patterns of spectral reflectance could be the differences in local conditions among these hotspots. For investigating further, we tested two hypotheses for exploring the potential of remote sensing datasets in ecology and biodiversity applications as proxy indicators of plant species richness. We observed a varied contribution of vegetation indices as well physiographic indices in explaining the patterns in overall species richness as well as life form wise species richness in the three hotspots. Vegetation indices were able to explain only up to 50% of the variance in plant richness, while addition of physiographic variables to models based on vegetation indices increased the predictive power of the models (Fig. 3, 4). This satisfies our first hypothesis, which states combined models have more predictive power than VI based models for plant richness. In past, various studies have reported significant positive correlations between plant SR from plot or regions and NDVI in both temperate (e.g., Fairbanks and McGwire 2004; Levin et al. 2007) and tropical ecosystems (Cayuela et al. 2006).
The difference in contribution by the variables could be attributed to the differences in the physiognomic conditions experienced by the three biodiversity hotspots. Spectral variation hypothesis (SVH) is generally followed in open canopy and low diversity ecosystems such as grasslands, scrubs and dry deciduous forests. However, in case of moisture rich ecosystems, multi-canopy structure affects the spectral response recorded by the satellite sensor, which could not address the variation in species richness in life forms such as shrubs and herbs occurring in the ground flora (Levin et al. 2007). The variation in overall species richness in dry deciduous forests, grasslands and scrubs was observed to best explained by vegetation indices, which is in line with the observations of Nagendra et al. (2010) in dry deciduous forests in India. This satisfies our second hypothesis, which could be attributed to the fact that these ecosystems exhibit low tree richness, less complex canopy structure and low moisture levels as compared to other vegetation classes. In case of grasslands such as savannah or dry grasslands in Western Ghats, trees are sparsely distributed; hence the variance in tree species richness was better explained by vegetation indices as compared to other vegetation classes.
The variance explained in similar vegetation classes such as grasslands and scrubs differed among the three hotspots, which could be attributed to the variations in the structural as well as functional attributes of the vegetation types. For example, in Himalaya grasslands occur above the tree line making it amenable to be addressed through the spectral reflectance of the satellite data, however in case of Indo Burma and W. Ghats grasslands are interspersed with trees/shrubs, which reduced the predictive power of the model based on the vegetation indices. Therefore, the variance explained in GL by vegetation indices was highest in Himalaya, followed by Indo-Burma and Western Ghats. The variance explained in the similar vegetation classes from the three hotspots obtained in the present study provides a range of variance, which could be replicable to same vegetation classes occurring in other parts of the country that experience similar climatic conditions. Addition of physiographic variables resulted in prominent increase in the variance explained in GL in Western Ghats and Himalaya (Fig. 3, 5), as compared to Indo-Burma that exhibits lesser physiographic variation. This signifies the role of complex physiography in Himalaya in controlling the distribution of plants, where greater environmental heterogeneity provides favorable conditions for speciation thus accommodating higher species richness.
Addition of relevant climatic and edaphic variables could enhance the variance explained in species richness. In Indo-Burma, the variance in species richness was relatively higher in low richness vegetation classes such as grasslands and degraded forests, as compared to moisture rich diverse vegetation classes such as moist deciduous and evergreen forests (Fig. 3). This indicates the role of climatic factors such as precipitation in controlling patterns of species richness in Indo-Burma. This hotspot experiences highest annual precipitation among the three hotspots, which is evident through distribution of bamboo vegetation, moist deciduous and evergreen forests in Indo Burma. The variance explained in tree richness was highest in grasslands and degraded forests, which could be due to open canopy in grasslands with interspersed trees, which helps in better explaining the patterns in tree species richness than in tree dominant forest vegetation classes. In Western Ghats, the variance explained by vegetation index based model was highest in grasslands and scrubs (Fig. 4), which was further increased after addition of physiography variables, which highlights the role of topography in controlling the patterns of species richness. Ability of models to explain high variance in species richness in plantations could be attributed to monotonous composition of tree plantations, which exhibits less spectral variation as compared to other natural ecosystems.
The stratification of overall plant richness into different life form wise richness provided better control over the statistical modelling of satellite derived spectral variation and SR. Generally, tree richness is observed to be highly correlated with the spectral vegetation indices, because of dominant occurrence in the top canopy; which is mostly visible to the satellite data (Rocchini et al. 2017). However, the present study reports better correlation of vegetation indices with approximately 55 to 60% among the SR in even in shrubs and herbs.
The present study provides explanation for largely unknown role of remote sensing observations in accounting for the variations in the plant richness in the biodiversity hotspots in India. Varied patterns of spectral reflectance among the same vegetation types viz., grasslands, scrublands, and dry deciduous forest falling in three hotspots hints at the role of local conditions in deriving the patterns of plant species richness. Highest range of spectral reflectance observed in Himalaya among the three hotspots, could be attributed to highest diversity in vegetation types in Himalaya due to variation in western and eastern Himalaya. Combined models created using physiographic indices and vegetation indices give better results in predicting life form wise SR. In the present study, we attempted to test spectral variation hypothesis, by integrating physiographic indices with vegetation indices to increase the predictive power of the models explaining variance in plant species richness. Integration of physiographic indices with vegetation indices created an overall scenario to represent the spectral heterogeneity in the study area, which certainly increased the model prediction accuracy. The investigation also provides novel insights on better correlation of vegetation indices with shrub and herb richness, which was sparsely attempted in the previous studies.
References
Austin GE, Thomas CJ, Houston DC, Thompson DB (1996) Predicting the spatial distribution of buzzard Buteo buteo nesting areas using a Geographical Information System and remote sensing. J Appl Ecol 33:1541–1550
Carlson KM, Asner GP, Hughes RF, Ostertag R, Martin RE (2007) Hyperspectral remote sensing of canopy biodiversity in Hawaiian lowland rainforests. Ecosystems 10(4):536–549
Cayuela L, Golicher DJ, Benayas JMR, González-Espinosa M, Ramírez N (2006) Fragmentation, disturbance and tree diversity conservation in tropical montane forests. J Appl Ecol 43(6):1172–1181
Currie DJ (1991) Energy and large-scale patterns of animal-and plant-species richness. Am Nat 137(1):27–49
Duro DC, Coops NC, Wulder MA, Han T (2007) Development of a large area biodiversity monitoring system driven by remote sensing. Prog Phys Geogr 31(3):235–260
Fairbanks DH, McGwire KC (2004) Patterns of floristic richness in vegetation communities of California: regional scale analysis with multi-temporal NDVI. Glob Ecol Biogeogr 13(3):221–235
Foody GM (2004) Sub-pixel methods in remote sensing Remote sensing image analysis: including the spatial domain. Springer, New York, pp 37–49
Gaitán JJ, Bran D, Oliva G, Ciari G, Nakamatsu V, Salomone J (2013) Evaluating the performance of multiple remote sensing indices to predict the spatial variability of ecosystem structure and functioning in Patagonian steppes. Ecol Ind 34:181–191
Gao B (1996) NDWI—a normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens Environ 58(3):257–266
Gillespie TW (2005) Predicting woody-plant species richness in tropical dry forests: a case study from south Florida, USA. Ecol Appl 15(1):27–37
Gillespie TW, Foody GM, Rocchini D, Giorgi AP, Saatchi S (2008) Measuring and modelling biodiversity from space. Prog Phys Geogr 32(2):203–221
Gould W (2000) Remote sensing of vegetation, plant species richness, and regional biodiversity hotspots. Ecol Appl 10(6):1861–1870
Levin N, Shmida A, Levanoni O, Tamari H, Kark S (2007) Predicting mountain plant richness and rarity from space using satellite-derived vegetation indices. Divers Distrib 13(6):692–703
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403(6772):853
Nagendra H (2001) Using remote sensing to assess biodiversity. Int J Remote Sens 22(12):2377–2400
Nagendra H, Rocchini D, Ghate R, Sharma B, Pareeth S (2010) Assessing plant diversity in a dry tropical forest: comparing the utility of Landsat and IKONOS satellite images. Remote Sens 2(2):478–496
O’brien EM, Field R, Whittaker RJ (2000) Climatic gradients in woody plant (tree and shrub) diversity: water-energy dynamics, residual variation, and topography. Oikos 89(3):588–600
Oindo BO, Skidmore AK (2002) Interannual variability of NDVI and species richness in Kenya. Int J Remote Sens 23(2):285–298
Öster M, Cousins SA, Eriksson O (2007) Size and heterogeneity rather than landscape context determine plant species richness in semi-natural grasslands. J Veg Sci 18(6):859–868
Pau S, Gillespie TW, Wolkovich EM (2012) Dissecting NDVI–species richness relationships in Hawaiian dry forests. J Biogeogr 39(9):1678–1686
Pausas JG (1994) Species richness patterns in the understorey of Pyrenean Pinus sylvestris forest. J Veg Sci 5(4):517–524
Pouteau R, Gillespie TW, Birnbaum P (2018) Predicting tropical tree species richness from Normalized Difference Vegetation Index time series: the devil is perhaps not in the detail. Remote Sens 10(5):698
Qi J, Chehbouni A, Huete A, Kerr Y, Sorooshian S (1994) A modified soil adjusted vegetation index. Remote Sens Environ 48(2):119–126
R Development Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org
Rocchini D (2007) Effects of spatial and spectral resolution in estimating ecosystem α-diversity by satellite imagery. Remote Sens Environ 111(4):423–434
Rocchini D, Ricotta C, Chiarucci A (2007) Using satellite imagery to assess plant species richness: the role of multispectral systems. Appl Veg Sci 10(3):325–331
Rocchini D, Petras V, Petrasova A, Horning N, Furtkevicova L, Neteler M (2017) Open data and open source for remote sensing training in ecology. Ecol Inform 40:57–61
Roy P, Karnatak H, Kushwaha S, Roy A, Saran S (2012) India’s plant diversity database at landscape level on geospatial platform: prospects and utility in today’s changing climate. Curr Sci (Bangalore) 102(8):1136–1142
Roy PS, Behera MD, Murthy M, Roy A, Singh S, Kushwaha S (2015) New vegetation type map of India prepared using satellite remote sensing: comparison with global vegetation maps and utilities. Int J Appl Earth Obs Geoinf 39:142–159
Schwarz M, Zimmermann NE (2005) A new GLM-based method for mapping tree cover continuous fields using regional MODIS reflectance data. Remote Sens Environ 95(4):428–443
Simpson EH (1949) Measurement of diversity. Nature 163:688
Tripathi P, Behera MD, Roy PS (2017) Optimized grid representation of plant species richness in India—Utility of an existing national database in integrated ecological analysis. PloS one 12(3):e0173774
Viedma O, Torres I, Pérez B, Moreno JM (2012) Modeling plant species richness using reflectance and texture data derived from QuickBird in a recently burned area of Central Spain. Remote Sens Environ 119:208–221
Willis KJ, Whittaker RJ (2002) Species diversity-scale matters. Science 295(5558):1245–1248
Acknowledgements
The authors are thankful to Dr. PS Roy, Project Director, Biodiversity Characterization project for providing the field sampling data for this study.
Disclaimer
The views and interpretations in this publication are those of the authors and are not necessarily attributable to ICIMOD.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M.D. Behera, S.K. Behera and S. Sharma.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Chitale, V.S., Behera, M.D. & Roy, P.S. Deciphering plant richness using satellite remote sensing: a study from three biodiversity hotspots. Biodivers Conserv 28, 2183–2196 (2019). https://doi.org/10.1007/s10531-019-01761-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-019-01761-4