
Abstract
Pine plantations established on former heathland are common throughout Western Europe and North America. Such areas can continue to support high biodiversity values of the former heathlands in the more open areas, while simultaneously delivering ecosystem services such as wood production and recreation in the forested areas. Spatially optimizing wood harvest and recreation without threatening the biodiversity values, however, is challenging. Demand for woody biomass is increasing but other pressures on biodiversity including climate change, habitat fragmentation and air pollution are intensifying too. Strategies to spatially optimize different ecosystem services with biodiversity conservation are still underexplored in the research literature. Here we explore optimization scenarios for advancing ecosystem stewardship in a pine plantation in Belgium. Point observations of seven key indicator species were used to estimate habitat suitability using generalized linear models. Based on the habitat suitability and species’ characteristics, the spatially-explicit conservation value of different forested and open patches was determined with the help of a spatially-explicit conservation planning tool. Recreational pressure was quantified by interviewing forest managers and with automated trail counters. The impact of wood production and recreation on the conservation of the indicator species was evaluated. We found trade-offs between biodiversity conservation and both wood production and recreation, but were able to present a final scenario that combines biodiversity conservation with a restricted impact on both services. This case study illustrates that innovative forest management planning can achieve better integration of the delivery of different forest ecosystem services such as wood production and recreation with biodiversity conservation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Pine plantations on heathland
Since the nineteenth century heathland has been converted to pine plantations in order to increase wood production and economic profit of these areas in both Europe and North-America, (Foster et al. 2002; Bertoncelj and Dolman 2013a; Bieling et al. 2013; Moran-Ordonez et al. 2013). However this has often led to a loss of fauna and flora associated with grassland, heathland and sandy habitats (GHS species) (Andres and Ojeda 2002; Farren et al. 2010; Bertoncelj and Dolman 2013a). Heathland is now considered a rare and threatened habitat which is eligible for protection under e.g. the European Habitat Directive (Walker et al. 2004). The resulting landscape type, combining open and closed habitats, is widespread throughout Europe and North-America and holds important values for biodiversity conservation, wood production and recreation.
To restore biodiversity values in these pine-heathland systems many efforts have focused on re-converting plantations and restoring heathland (Eycott et al. 2006; De Valck et al. 2014). Nevertheless, while this can be a valuable and practical strategy in terms of biodiversity of the GHS species (Walker et al. 2004), recovery can be slow and results in a loss of other species that are linked to aggrading (pine) forests (Ozanne et al. 2000; Burton 2007). Moreover, re-conversion to heathland is not always possible and desirable for all stakeholders, because forest plantations also offer other key ecosystem services such as wood production, soil protection, water regulation and recreation (Zipper et al. 2011; Vihervaara et al. 2012; Jacobs et al. 2013; De Valck et al. 2014). The demand for woody biomass for example is high and rapidly increasing (Mantau et al. 2010) and forests in densely populated regions such as Flanders face a very high recreational demand (Hermy et al. 2008).
While trade-offs between pine plantations and GHS species conservation are obvious, it has often been overlooked that benefits could be non-exclusive (Bertoncelj and Dolman 2013a). Viable populations of some GHS species persist in the pine plantation matrix, thanks to the network of temporal (e.g. clear-cut areas) and permanent open patches (e.g. remnant heathland, forest rides) (Bertoncelj and Dolman 2013a; Pedley et al. 2013). In addition to these GHS species, also typical species of pine forests are hosted in these landscapes. These forest specialists, such as forest carabid beetles, are often negatively affected by increasing open areas (Barbaro et al. 2005, 2007). Hence, we argue that forest management in these systems, with a focus on wood harvest and recreation, definitely has certain trade-offs with biodiversity conservation. However, benefits of wood harvest and recreation could be non-exclusive, also leading to some synergies with biodiversity conservation. Additional quantitative data could help to further unravel the relation between the services of plantation forests and biodiversity conservation.
Recreation and biodiversity
There is a general consensus that recreation can have a direct negative impact on biodiversity (Steven et al. 2011), mainly by altering the ability of animals to exploit resources (Gill 2007). However effects of recreation vary across ecosystems, species, recreation forms and intensity levels (Liddle 1996; Ficetola et al. 2007). Some species groups are specifically vulnerable, such as ground-breeding birds (Mallord et al. 2007), ground-dwelling forest birds (Thompson 2015) and large mammals (George and Crooks 2006). Impact of recreation on ground dwelling arthropods is generally low (Zolotarev and Belskaya 2015), but butterflies were reported to be directly, negatively influenced by recreation (Bennett et al. 2013). There are also varying approaches to estimate the impact of recreation on species with divergent results (Gill 2007) and the relationship between the amount of recreational use and recreational impact is not always (curvi) linear (Monz et al. 2013). Mallord et al. (2007) found a clear negative effect of disturbance on the density of woodlarks in heathlands (Lullula arborea). George and Crooks (2006) found a lower density of large mammals along paths with more visitors in an urban nature reserve dominated by shrubs and open oak forests. Thompson (2015) underlines the need for trail-free refuge habitat for forest birds in deciduous forests. These examples show that there can be a strong impact of recreation on different species in different habitats. However, for the local context of our study area (pine plantations on former heathland), there is hardly any literature to be found. Only for the ‘flagship’ bird species, European nightjars (Caprimulgus europaeus), strong negative effects of visitors on nightjar populations were identified (Langston et al. 2007; Lowe et al. 2014).
To reduce the impact of recreation on biodiversity, a trail network can be designed to guide recreationists to spatiotemporally separate visitors from vulnerable species (Ferrarini et al. 2008). Standard trail design is already used to avoid vulnerable areas and to screen sensitive species from disturbance by recreation, but is sometimes too general for optimal results (Rodriguez-Prieto et al. 2014). A better way to design trails is based on empirical research (Fernandez-Juricic et al. 2007) and by the use of simulation models (Stillman and Goss-Custard 2010) to tailor the trail design to best fit the local context. However, Ficetola et al. (2007) and Rodriguez-Prieto et al. (2014) demonstrated that an appropriate design for one focal species is not necessarily appropriate for another species. Subsequently, adopting a multi-taxa approach might promote intelligent trail design to limit disturbance for a whole set of species. An example of such an intelligent steering of recreation pressure are the differing access rules for different users (walkers vs walkers with dogs vs boating activities) in the protection of bird colonies as proposed by Fernandez-Juricic et al. (2007).
Wood production and biodiversity
Wood harvest from clear-cuts can have a direct negative influence on forest species (Linden and Roloff 2013). Species dependent on shade, dead wood, old trees and cavities, such as shade-demanding woodland herbs, woodpeckers and saproxylic beetles are most vulnerable (Martin and Eadie 1999; Djupström et al. 2012). Clear-cuts also have a drastic influence on microclimatic environmental and biological conditions such as light, temperature and availability of food and shelter. However, species that are suited to more open conditions will use intensively managed forests and open, clear-cut areas as new valuable habitats (Bertoncelj and Dolman 2013b; Morris et al. 2013; Reidy et al. 2014). At landscape scale, the patchwork of open patches in a forest matrix can sustain viable metapopulations of GHS species. However, the success of these metapopulations will depend on the spatiotemporal lay-out of the clear-cuts and the dispersal capacity of the species (Johst et al. 2011).
Most programs to conserve forest biodiversity focus on setting aside protected areas and creating forest reserves (Lindenmayer et al. 2006). It has been stated that forest reserves alone are not enough because they generally only cover a limited area and are often isolated from each other (Daily et al. 2001; Lindenmayer et al. 2006; Mönkkönen et al. 2014). Another biodiversity conservation measure is retaining mature forest habitat elements on clear-cuts, such as green trees or snags, to reduce the negative impacts of wood harvest in clear-cuts. (Söderström 2009; Linden and Roloff 2013). Recent findings highlight that a green tree retention level of at least 10–15 % of all standing trees on large areas is needed to obtain a strong conservation effect on most forest bird species (Söderström 2009). This contrasts with current retention levels which are often around 2 % (Söderström 2009). Installing protected areas and retaining habitat elements could definitely be part of an effective forest biodiversity conservation strategy, but at the same time it is important to create structural diversity on different scales and to increase habitat connectivity for different species (Lindenmayer et al. 2006; Brockerhoff et al. 2008; Gustafsson and Perhans 2010).
For the protection of the vulnerable GHS species, conservation managers often create permanent open patches with grassland, heathland or sand dunes (Walker et al. 2004). Another classical conservation measure, also to increase habitat connectivity, is the broadening of forest roads (Bertoncelj and Dolman 2013a).
All of the above mentioned conservation strategies are valuable, but all have a clear trade-off with wood production. Management scenarios that optimize spatial design of temporal open patches to sustain metapopulations of both GHS and forest dwelling species are less conventional. However, these innovative methods could be highly effective (definitely when combined with classical conservation strategies), while more or less safeguarding the important wood and biomass production function of forests (Mönkkönen et al. 2011, 2014).
Management challenges
Forest managers have the challenging task to balance management between biodiversity conservation, wood production and recreation among other ecosystem services. Classic land sparing approaches, such as setting aside protected area are well known. Under land sparing, biodiversity conservation is spatially separated from production and other services, while under land sharing both goals are integrated on the same land (Phalan et al. 2011). Land sharing can be a very valuable conservation strategy if benefits of ecosystem services and biodiversity conservation are non-exclusive (Phalan et al. 2011). However, information and data on innovative land sharing approaches in forests, combining these three management goals and minimizing spatio-temporal trade-offs, are lacking. Moreover, land sparing and land sharing are often treated as alternative strategies (Phalan et al. 2011) but a combination of both approaches would likely be the most successful strategy since different actions benefit different species and ecosystem services (Rey Benayas and Bullock 2012). We thus set out to investigate the trade-offs between biodiversity conservation and both wood production and recreation in a pine plantation on former heathland and explored possible scenarios for improvement. We gathered empirical data for the different services and used spatially-explicit analyses to study the trade-offs between biodiversity and the two ecosystem services. Based on the analyses we formulate future management and recreation scenarios with their impact on biodiversity for our case study area. We complement contemporary management approaches to advance smart(er) ecosystem stewardship that can both benefit policy makers and practitioners beyond our study area.
Materials and methods
Study area
The study was performed in North-Eastern Belgium in Bosland (center of the study region: 51.17°N, 5.34°E). Bosland is a statutory partnership of four public owners and two non-profit organizations that used to work next to each other, but now closely collaborate to increase the impact and coherence of the management in their forest and nature areas (Vangansbeke et al. 2015a). Bosland covers a total surface area of 22,000 ha of which approximately 35 % is forest, 7 % heathland and 3 % grassland. The soils are characteristically dry, sandy and nutrient poor and were classified as Carbic Podzols (IUSS Working Group 2007). Until the middle of the nineteenth century, Bosland was mainly covered by an extensive heathland. Afterwards, gradual afforestation with conifers took place with Scots pine (Pinus sylvestris) and Corsican pine (Pinus nigra ssp. laricio var. Corsicana Loud.) as dominant tree species (Vangansbeke et al. 2015b). For this study we delimited a study area of 1347 ha in the heart of Bosland, commonly known as Pijnven and Slijkven. The study area is covered by a matrix of pine plantations and has traditionally been managed for wood production under a simple harvest regime including some thinnings (from 30 years stand age, ca each 6–9 years) and a final clear-cut after ca. 50–100 years. For biodiversity purposes certain areas have been set aside as forest reserves (26 ha) and as permanent open patches (77 ha). The forest matrix is interlaced with a network of forest rides that are both used for recreation (mostly walking, but also cycling and horseback riding) and for wood harvest and can also be a valuable habitat for the GHS species. The study area is also split up in two zones, one with a high recreational pressure (792 ha) and the other with a low recreational pressure (555 ha), without signposted tracks.
Data collection
Biodiversity
We chose to use an indicator species approach to monitor the biodiversity of the study area. We organized a brain-storm session with the local platform on fauna and flora, formally grouping people that work on management and research of species in the area with volunteers of local nature conservation organizations, often involved in monitoring (Vangansbeke et al. 2015a). We asked them to make a credible selection of indicator species that are locally relevant. Indicator species needed to be medium widespread within the study area and reasonably detectable. To increase the ecological relevance we asked for a large range in species’ habitat preference, mobility and home range. Ten indicator species were selected for the in-depth study, but only seven species were used in the analyses (Table 1), three other species (i.e., Coronella austriaca, Formica spec. and Genista pilosa), were removed from the analysis, because the total number of observations was below ten. The final indicator species pool consisted of two forest species (crested tit and coal tit; Lophophanes cristatus and Periparus ater), three GHS species (grayling, small heath and northern dune tiger beetle; Hipparchia semele, Coenonympha pamphilus and Cicindela hybrida) and two species that depend both on forest and open patches (nightjar and common lizard; Caprimulgus europaeus and Zootica vivipara). A literature review was performed to double check the habitat preference and the species mobility (Table 1).
An inventory of the butterflies and the tiger beetle was made three times along transects on the forest rides through the study area in June and August of 2013 and 2014 by bicycle or on foot (Fig. 1a). The insect inventory was done between 10 am and 4 pm and only on sunny days, binoculars were used for easier determination from a distance. The exact GPS location of each observation of an individual was registered. The crested tit and coal tit were also inventoried by walking the observation transects three times, between sunrise and 11 am on non-rainy days in April 2014. The observations were auditory (recognition of vocal sounds) and the exact location was not determined, but a stand was marked as occupied or not. We alternated the direction of the transects between days to compensate for a possible time effect (e.g. highest bird activity just after sunrise). The nightjar inventory was based on the sound of its churring song on one warm summer evening (July 10th 2014) with the help of no less than 60 volunteers spread over the entire study area. Each churring individual was marked on a map. The distribution of common lizards was assessed based on presence under black corrugated sheets that served as artificial refuges (Busby and Parmelee 1996). Eighty of these sheets were laid out across the entire study area, left for one year and checked for presence of lizards three times in August 2014. Finally the rough data for the biodiversity inventory were compiled in a map with 821 point observations of the seven species (Fig. 1a).

a Total number of observations of the different indicator species within the study area (n = 821). b number of visitors on the different roads in the forests as deducted from interviews with forest guards and counts from trail counters
Recreation
Bosland is a very important touristic destination with more than 1,000,000 yearly overnight stays. To determine the spatial distribution of the numerous visitors we compiled quantitative visitor data with questionnaires and automated trail counters. We started with interviewing the forest managers about the number of visitors on different road segments. We used a map with all roads and tracks and asked them to mark them with five different colors based on the relative recreational intensity. We then made up a relative recreational intensity map with an average score from the interviews. Then we installed six automated infrared trail counters (TRAFx research ltd, Canmore, Alberta, Canada) to quantify the exact number of visitors. The location of the trail counters was decided in consultation with the forest guards and with the goal to survey varying recreation intensities. We only had the counters available during a period of seven months between October 2014 and May 2015. To interpret these counts and the possibility to extrapolate the data we investigated data of four other counters in Bosland, just outside our study area. These counters were all located within 7 km of our study area in similar habitat and counts for three consecutive years were available. From this data we calculated a conversion factor as the ratio between the average daily number of visitors between October and May and the average daily number of visitors for a whole year. We found an average conversion factor of 1.05 (SD. 0.18) and used this to estimate an average daily number of visitors for a whole year for our own counter data.
Next we adjusted the relative recreational intensity map with the estimated average number of daily visitors for every road segment and obtained a map with an estimated recreational pressure for each road segment (Fig. 1b). We also calculated a recreation pressure score for each forest stand with the following formula (Fig. 4 in Appendix):
Wood production
We estimated the mean annual increment for each stand, based on the stand age, the dominant tree species and the site quality which was deducted from the soil map (after Broekx et al. (2013)). We then calculated the standing stock of every stand by multiplying the stand age with the stand area, the mean annual increment for each tree species and a harvest factor. The harvest factor was calculated as the ratio between the volume of the final harvest and the total volume of all thinnings and final harvest according to the growth table of Jansen et al. (1996).
The production of biomass from tree tops was calculated with the stem volume and a species-specific biomass expansion factor (after Vande Walle et al. (2005)) and with estimated harvest losses of 40 % (after Vangansbeke et al. (2015c)).
Habitat characteristics
We combined different data sources and layers to map the habitat in the study area. First of all, for the forest stands we used the map of the 2010 forest inventory. This map included all the forest stands and important habitat information such as dominant tree species and stand age. We added two important habitat features to this data layer, namely the recreational pressure of the stand (based on the pressure of the surrounding road segments) and the amount of neighboring open habitat.
We mapped the entire road network, based on aerial photographs and ground field data. The main habitat features for each road segment were the area, the orientation, the recreational pressure and the surface type (tracks on sand, grass, tree litter or paved with tarmac). We also mapped the non-forested patches; the area, the recreational pressure and the surface type (orchard, agriculture, sandy, heathland/grassland, clear-cut, plantation) were the main habitat features. We combined the road network layer with the non-forested patches layer in one layer for all open habitats.
An overview of the different habitat features of both forests and open habitats is given in Table 5 in Appendix.
Data analyses
Biodiversity
All spatial analyses were performed in QGIS 2.10.1 (QGIS Development Team 2015) and all statistical analyses were implemented in R 3.0.1 (R Core Team 2013), using the multi-model inference package (MuMIn). Every point observation was assigned to a forest stand (coal tit, crested tit, lizard and nightjar) or to an open habitat element (i.e. a road segment or a permanent open patch) (butterflies, beetles, lizard and nightjar). For nightjars we used a circular buffer with a radius of 20 m, because the exact location of a churring individual is hard to locate exactly. The presence of a certain species in a patch was modelled with a logistic regression with the different habitat features as predictors for the patches (either forest stands or open habitat patches) that were part of the inventory for this species. Patches were considered as part of the inventory when lying adjacent to an observation route (forest species and GHS species), containing a corrugated sheet (lizard) or lying within 400 m of an observation point (nightjar) (after Rebbeck et al. (2001)). Observation surface was included in the regression models as a covariate, to compensate for the fact that a higher observation surface automatically leads to a higher number of observations. We rescaled all numerical predictors by subtracting the mean value and dividing through the standard deviation to increase comparability. We ran generalized linear models (GLMs) using a binomial distribution for every possible combination of predictors (i.e. 256 models for forest patches, 16 for open patches). The models were ranked based on the AIC criterion, using the dredge function in the MuMIn package. Models with a delta AIC smaller than four were considered equivalent (Bolker 2008). These so-called top models were used to calculate an average model with the model averaging function in the MuMIn package (Symonds and Moussalli 2011). The R2 was calculated for the model containing all predictors that appeared in the top models. The importance value was used to evaluate the relevance of the different predictors for species distribution. Habitat features that did not appear in the top models were left out of the analysis. The final average coefficients were used to predict probabilities of presence of the different species in all patches. The probability of occurrence was considered as a measure for habitat suitability and was mapped with a value between 0 and 1 for every habitat patch.
These habitat suitability maps were imported in Zonation 4 (C-BIG, Helsinki), a framework and software tool for conservation prioritization and large-scale spatial conservation planning. It identifies areas that are important for retaining habitat quality and connectivity simultaneously for multiple species, thus providing a quantitative method for enhancing persistence of biodiversity in the long term (Moilanen et al. 2014). The software tool translated the habitat suitability maps to a raster with 5 × 5 m2 grid cells and ranked these cells according to their importance for the maintenance of a species. We used the basic core-area cell removal rule algorithm to decide which cells were least important for a species (Moilanen et al. 2014). To evaluate habitat quality and connectivity, this algorithm depends on two species-specific biological parameters: the dispersal capacity and the kernel width. The dispersion capacity was calculated as the inverse of half the dispersion distance in meters (Moilanen et al. 2014) (Table 1). The kernel width was based on the mobility of a species through the forest matrix. For the species that depended on forest we set the kernel width to 50 m, the species of open habitats were assigned a smaller kernel width of 35 m (small copper and grayling) and 20 m (Northern dune tiger beetle) depending on their dispersion distance. We grouped the forest species (coal and crested tit), the GHS species (two butterfly species and one beetle species) and the mixed species (nightjar and lizard). We thus obtained one rank of the different pixels in the study area for their suitability to sustain the current populations of the seven species under study.
Recreation and biodiversity
To evaluate trade-offs between recreation and biodiversity we selected the species for which recreation was an important variable in predicting the distribution (threshold set on an importance value larger than 0.5 (Lindtke et al. 2013)). These species were presumed to be vulnerable for recreation, as their distribution was negatively related to recreation intensity. The stands that had the highest average rank in Zonation for these vulnerable species were considered as the stands that were most vulnerable for recreation. The stands with the lowest average rank were considered as stands where recreation pressure has a lower impact on the distribution of the species involved. To test the trade-offs between recreation and biodiversity we calculated the impact of three recreation scenarios on the habitat suitability for the involved species. Scenario S1 doubles the amount of recreation everywhere (200 % visitor pressure); Scenario S2 doubles the amount of visitors in the least vulnerable areas (for the involved vulnerable species) and halves the amount of visitors in the most vulnerable areas, leading to an overall increase of 25 % in the number of visitors (125 % visitor pressure); Scenario S3 increases recreation with 25 % everywhere (125 % visitor pressure). We used these hypothetical recreation data to calculate the habitat suitability with the GLM for every species and compared the average habitat suitability score with the current reference.
Wood production and biodiversity
To evaluate the impact of harvesting on biodiversity we looked into the habitat preferences of the species. We considered a negative effect of clear-cuts on the habitat quality and connectivity for forest species and a positive effect on GHS species, that will profit from these new, temporal open patches. Nightjars depend on both forest stands and open patches and are very mobile, so spatial allocation of the harvested stands is probably less crucial to sustain populations. We next developed a harvesting plan for the next 20 years according to three different harvesting scenarios. First, in a wood production scenario, we followed the existing long-term vision on wood production (Moonen et al. 2011). Under this scenario, the oldest and most productive stands are harvested first. We ranked the stands by hand to a decreasing wood production score and harvested every year about 1 % of the total area (rotation period of 100 years). Second, in the biodiversity scenario, we first harvested the stands that have a low importance for forest species distribution and a high rank for the distribution of the GHS species. The third scenario is an integrated scenario that puts equal weights on the wood production rank and the biodiversity rank (as a low forest species rank and a high rank for GHS species). Finally, we calculated the output flow of harvested stem wood (and crown biomass) under these three scenarios.
Results
Habitat suitability
Our statistical models successfully explained the distribution of all but one of our study species (importance values and adjusted R squared in Table 2, average coefficients in Table 6). Only for the common lizard, we found that the best model was the intercept only model, without any environmental predictors. This is probably due to both the low number of patches in the inventory and the low number of observations. This species was left out of the analysis.
The probability of occurrence of the coal tit was strongly negatively related to a higher recreation pressure and to the amount of adjacent open patches. Coal tits seemed to prefer closed high forest without open patches, without too much recreation and from age class 81–100. Crested tits had a higher probability of occurrence in large high forest stands, with a low recreational intensity and a limited amount of border with open habitat.
The probability of occurrence for churring nightjars was higher in smaller stands with a high amount of adjacent open habitat. Also some stand age classes had a much higher probability of occurrence for nightjars, particularly stands from age class 81–100, 21–40 and uneven aged stands. To a much lesser extent the probability to find churring nightjars was also negatively related to the amount of recreational intensity. In the open patches, probability of presence of churring nightjars was mainly related to patch type (high probability in young plantations and low probability in agricultural and orchard patches) and size (again higher probability in smaller patches). In general, there was a higher number of churring nightjars in forest stands (104) than in open habitats (41).
The probability of occurrence of small heath was positively related to large open patches with grassland, heathland or sandy habitats and to a low number of visitors. Grayling had a higher probability of occurrence in clear-cuts and plantations and to a lesser extent in grassland, heathland and sandy habitats. Also for grayling we found a negative relation between the recreation intensity and the probability of occurrence. The probability of occurrence of tiger beetle was highest in large open patches with a sandy surface and in grassland or heathland.
The average coefficients of the top models were then applied to predict probability of occurrence for the indicator species in stands and open patches (Fig. 5 in Appendix). These habitat suitability maps were then imported in Zonation to evaluate the value of each grid cell for the conservation of a species groups (forest species, GHS species and species that depend both on stands and on forest), given the spatial distribution of the habitat patches and the mobility characteristics of the indicator species (Fig. 6 in Appendix).
Recreation and biodiversity
The coal tit, the small heath and the grayling were most vulnerable for recreational pressure. We ranked all landscape cells according to their conservation priority for these species and made up a map with the most and the least vulnerable areas for these species concerning recreation (Fig. 2).

Vulnerability of forest stands to recreational pressure for coal tit, small heath and grayling. This map can aid management and recreation planning
Next we investigated the effect of different hypothetical recreation scenarios on the habitat suitability for coal tit, small heath and grayling (Table 3). We found a negative effect of increased recreation on habitat suitability, but the impact was much smaller if recreation in the most vulnerable patches was limited. There is thus a trade-off between recreation and biodiversity conservation, but it can be minimized if both goals are spatially separated.
Wood harvest and biodiversity
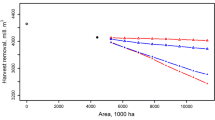
Harvesting wood and biomass from final cuttings transforms mature stands to clear-cuts. This could have both negative (forest species) and positive (GHS species) effects on conservation of species. Depending on the developed scenarios, the management plan for the next twenty years diverge substantially. The biodiversity and wood production scenario share hardly any stands spatially, while under the integration scenario most harvested stands are also harvested under the biodiversity or wood production scenario (Fig. 3). As expected the harvest of woody biomass is strongly determined by the chosen scenario (Table 4). In general, the more biodiversity conservation is included as a management target, the less wood is harvested, indicating a clear trade-off between these management goals.

Harvest schedule for the next twenty years under three different scenarios, a wood production scenario (WPS), a biodiversity scenario (BS) and an integrated scenario (IS). Some stands are only harvested under one scenario, some are harvested under two scenarios (brown and green dots) and some even under three scenarios (black dots). (Color figure online)
Discussion
Habitat suitability
Our results demonstrate that patch habitat features play an important role in the probability of occurrence of the indicator species. Only for the lizard, we found no significant relationship with any of the analysed habitat features. For the other species that use the forest matrix as a habitat, important features are the recreational pressure, the amount of forest border, the stand age or management type and the area. The contrast between coal tits and nightjars was interesting, with the former preferring large stands with limited borders and the latter preferring small stands with adjacent open space. This is in line with our expectations since coal tit was classified as a forest species (Brotons 2000) and nightjar as a mixed habitat species (Verstraeten et al. 2011). For the trail network and the open patches, we found a strong relation between the type of ground cover and the probability of occurrence of all indicator species. The butterfly species seemed to be more abundant when the number of visitors was lower. It is not surprising that the tiger beetle preferred large, sandy patches, however it is necessary to treat the results for this species with some caution, considering the limited number of observations. The probability of occurrence of small heath was bigger in larger open patches, while the opposite was true for nightjars. Nightjars thus occurred more in both smaller patches of forest and smaller open habitats, this links to its preference to a varied landscape. Nightjars were described to be vulnerable to recreational pressure (Langston et al. 2007; Lowe et al. 2014), however we did not detect a strong relation between recreational pressure. A possible explanation could be the mismatch between the location of a churring bird and the breeding location and the fact that we gathered data after sunset, when there is hardly any disturbance by recreation. Langston et al. (2007) mentioned that the main disturbance by recreationists on nightjars was related to a lower breeding success. Disturbance at the song posts after sunset will be much more limited.
Recreation and biodiversity
The stands that were important for the conservation of the populations of the coal tit and the butterflies were determined as the stands most vulnerable to recreational pressure. Most, but not all of the stands that were mapped as ‘vulnerable’ are already located in the actual zone with a low recreational pressure. This was expected, because the current distribution of the three species is the main parameter to determine the stand vulnerability and the distribution of these species is already influenced by the actual recreational pressure. The fact that we did not find a strong relationship between the number of visitors and the other indicator species does not necessarily mean that there is no such a negative effect of recreation on these species. However our data do not allow to assign certain stands for protecting these species. It is important to note that most of the stands that were mapped as tolerant to recreation were located at the edge of the study area. This is partly because of their habitat features that are less suited to sustain the indicator species. The effect is reinforced by the basic core-area cell removal rule algorithm implemented in Zonation, that promoted suitable habitat that is connected to other suitable habitat (Moilanen et al. 2014). When looking into the results of the hypothetical recreation scenarios we observe a decrease of the habitat suitability for the three species with an increasing number of visitors. Protection of the most vulnerable patches seems indeed crucial to sustain populations. After all, we found a very low decrease in habitat suitability in scenario S2 compared to S3, for the same total amount of visitors.
Our results thus seem to support the classical, land sparing approach to design the track network mostly in the border of a nature reserve, while safeguarding the core of the area from visitors for conservation purposes (Rodriguez-Prieto et al. 2014).
Wood production and biodiversity
Depending on the management focus, the temporal lay-out of the clear-cuts is almost entirely different, with the integrated scenario as an intermediate solution between both mono-functional scenarios. Stands harvested under the biodiversity scenario are mostly located closer to the edge of the study area where there is a low conservation value for the forest species and adjacent to existing open patches to increase habitat of GHS species. Adoption of the biodiversity scenario would reduce yearly stem harvest with ca. 22 %. When using an average resale price of 23 € m−3 stem wood and of 4 € m−3 crown wood (after Vangansbeke et al. (2015b)), subtracting a 33 % margin of profit for the harvesting company), the biodiversity scenario results in an income decrease of 29,000 euro per year compared to the wood production scenario over a planning period of 20 years. In the integrated scenario, annual harvest declines by ca. 13 % and total income by 17,000 € yr−1. It is important to be cautious in interpreting these economic values which are based on a rough estimation of growth and for instance neglect possible positive biodiversity effects on tree growth. With the given data, forest managers can easily develop their own scenarios with a different weight for biodiversity or harvesting. Although including biodiversity conservation as a management goal negatively affects wood production, the results show that a land-sharing approach is possible with limited impact on either wood production and biodiversity conservation.
Integration of services
In order to better support complex ecosystem dynamics, we will need to develop a new kind of (planetary) stewardship [e.g. Power and Chapin (2010); von Heland et al. (2014)] which combines a systems approach with transformative action. The current study can be seen as a first stepping stone in this regard since we combine first notions of systems thinking (linking biomass production, biodiversity and recreation; using multi-species analysis; scenario development) with a more transformational approach (involving volunteers, action research design, focus on practical applicability and close cooperation with policy makers) in a real-life setting. We believe that this exploratory study furthers our understanding of what ecosystem stewardship entails by adding new insights on the trade-offs of different management scenarios which may be of particular interest for policy makers or practitioners on the field.
Developing a management scenario that includes recreation pressure, wood harvest and reaches biodiversity conservation goals is not easy. Comparing different management scenarios can help forest managers to identify knowledge gaps that need to be addressed for better ecosystem management and can help policy makers to develop adaptive management approaches that are more appropriate to support a multitude of ecosystem services. The different scenarios show how management can be focused locally on increasing either biodiversity or biomass harvest. By bringing these two together in an integrated scenario, an approach can be developed where the trade-offs can be minimized. Installing the integrated harvest plan would increase the value of the landscape for biodiversity conservation, while safeguarding 87 % of the current wood harvest. In combination with an intelligent trail design and conventional conservation strategies this could be an important step towards bringing into practice better stewardship management arrangements.
Scenarios such as the ones developed here can be very useful for forest managers since they provide first indications on the estimate of the income loss (or suspended income) when incorporating biodiversity conservation as a management goal. They can better balance installation of this scenario with the costs of other biodiversity conservation measures. Mönkkönen et al. (2011) modeled the cost-effectiveness of different biodiversity conservation measures: installation of a few permanent large reserves, of many permanent/temporary small reserves (‘SLOSS dilemma’), and green tree retention. An important next step would be to investigate what additional costs might arise over a longer time period when choosing the wood production scenario.
When management is focused solely on biodiversity conservation, both recreation and harvest are restricted to the stands at the border of the study area. While not included in our study, there also exist trade-offs and synergies between recreation and wood harvest. On the one hand, recreationists value structural variation at the landscape scale. On the other hand, clear-cuts can evoke strong objections by visitors (Brunson and Reiter 1996). Forest management measures such as thinning can also affect recreation. There is little information available, but Heyman et al. (2011), for example, studied the effect of openness in the understory of plantation forests and found a preference of visitors for a more open understory, but a slightly negative effect on bird biodiversity in more open plots. In order to develop better stewardship practices, more research is thus needed to cover a wider spectrum of ecosystem services and a more encompassing set of species.
Methodological remarks
Of course, studying multi-species habitat preferences on a landscape scale is susceptible to uncertainties such as parametrization of the modeling. First of all, the selection of the indicator species is an important a priori choice that will have an important influence on the results. Ideally, all biodiversity components across all taxa are included but this is virtually impossible. Therefore, indicator species are chosen that are assumed to well represent the biodiversity values of a patch. It is important to choose different indicator species from a wide range of taxa, habitat preferences and mobility (Heink and Kowarik 2010; Rodriguez-Prieto et al. 2014; Pakkala et al. 2014). With help of the local volunteers we succeeded in fulfilling these requirements. However, due to limited observations we had to exclude some species from the analysis, causing a slightly unbalanced distribution of indicator species with only insects as GHS species and only birds as forest species.
Second, we considered only presence/absence of a species in a habitat patch as an indicator for a suitable or unsuitable territory. We believe this to be quite accurate for the insects, the lizard and the singing coal and crested tit. Nightjar home ranges, however, are much bigger and absence of a churring bird is probably not a solid indicator of unsuitable habitat (Sharps et al. 2015). However, given the complex life strategy of nightjars and the difficulty in mapping nightjar territories (Rebbeck et al. 2001), our methodology seems a good compromise with practical feasibility. We also chose for a high spatial resolution with a high number of observers, but as a drawback we only used data from one night, which could distort the results.
A third element that could distort the interpretation of the result is a possible mismatch between the scale of habitat mapping and preferences of the smaller indicator species. We worked at the landscape scale and performed analyses on the patch level (forest stands, forest road segments and open patches). Distribution of some indicator species will depend on micro-habitat features within patches, such as the presence of a host plant, microclimates or a smaller structure element [e.g. for grayling (Maes et al. 2006)], and bear little relationship with patch-level habitat features (Pakkala et al. 2014). However, only working at the patch level in such a large landscape was practically feasible.
A fourth element that influenced our final results was the delineation of our study area. Our study area can more or less be considered as an ecological unity with sharp borders, agricultural areas in the north-west and main roads in the south and east. We thus considered every cell outside our study area as unsuitable habitat for the studied populations. Well-connected habitats occurred logically more in the center of our study area than at the border and were thus awarded a higher conservation value. Setting the importance value threshold on 0.5 (after Lindtke et al. (2013)) to evaluate vulnerability to recreation can also be subject to debate. (Calcagno and de Mazancourt 2010) suggest a threshold of 0.8, which would give a different result in our analysis. A direct measurement of recreation pressure at the same time of species distribution mapping would maybe also have yielded better results. However we used the best available alternative, by estimating the year round recreation pressure with help of a conversion factor.
Finally there are also lots of related issues that were not looked into in this study, but where supplementary research could be highly valuable. A next step could be a more formal trade-off analysis between wood harvest, recreation and biodiversity conservation. This can be achieved through a multi-criteria analysis of a set of alternative scenarios that combine different harvest and recreation regimes. However, to quantify the impact of each scenario on biodiversity conservation, absolute values are required instead of relative biodiversity conservation scores as those provided by the program ‘Zonation’. Another interesting way to assess these trade-offs could be to execute the proposed management scenarios in the field and evaluate their impact on biodiversity. The future biodiversity surveys can then be used in future management plans, adopting a true adaptive management cycle (Lindenmayer et al. 2006). Other aspects that urgently require further research include the relationship between other harvesting techniques than clear-cuts and biodiversity (e.g. Fuller (2013)), the biological interactions between species (Pakkala et al. 2014) and the economic (Schou et al. 2012) and ecological (Brown et al. 2015) impact of the planned conversion to broadleaves.
Conclusion
To conclude, the combined valuation of biodiversity conservation and wood production led to an integrated harvest plan that increases the biodiversity conservation value of the landscape, while safeguarding 87 % of the current wood harvest. In addition, knowledge on the conservation value of stands for species vulnerable to recreation can help to improve the trail network design, guiding visitor streams and sheltering biodiversity hotspots. We showed that wood production and recreation have certain trade-offs with biodiversity conservation. However, with a better spatiotemporal design, important biodiversity conservation gains can be made without greatly reducing the delivery of other services. The current study will help policy makers and practitioners to develop future management schedules, for Bosland and beyond. Moreover it demonstrates that a combined land-sharing (for wood harvest) and land-sparing (for recreation) approach allow finding compromises between ecosystem services and biodiversity. There is an urgent need for additional research on the science-management interface, mainly on the interplay of different forest ecosystem services and the impacts for biodiversity.
References
Andres C, Ojeda F (2002) Effects of afforestation with pines on woody plant diversity of Mediterranean heathlands in Southern Spain. Biodivers Conserv 11:1511–1520
Barbaro L, Pontcharraud L, Vetillard F, Guyon D, Jactel H (2005) Comparative responses of bird, carabid, and spider assemblages to stand and landscape diversity in maritime pine plantation forests. Ecoscience 12:110–121
Barbaro L, Rossi JP, Vetillard F, Nezan J, Jactel H (2007) The spatial distribution of birds and carabid beetles in pine plantation forests: the role of landscape composition and structure. J Biogeogr 34:652–664
Bennett V, Quinn V, Zollner P (2013) Exploring the implications of recreational disturbance on an endangered butterfly using a novel modelling approach. Biodivers Conserv 22:1783–1798
Bertoncelj I, Dolman PM (2013a) Conservation potential for heathland carabid beetle fauna of linear trackways within a plantation forest. Insect Conserv Divers 6:300–308
Bertoncelj I, Dolman PM (2013b) The matrix affects trackway corridor suitability for an arenicolous specialist beetle. J Insect Conserv 17:503–510
Bieling C, Plieninger T, Schaich H (2013) Patterns and causes of land change: empirical results and conceptual considerations derived from a case study in the Swabian Alb, Germany. Land Use Policy 35:192–203
Bolker BM (2008) Ecological models and data in R. Princeton University Press, Princeton
Brockerhoff EG, Jactel H, Parrotta JA, Quine CP, Sayer J (2008) Plantation forests and biodiversity: oxymoron or opportunity? Biodivers Conserv 17:925–951
Broekx S, De Nocker L, Liekens I et al (2013) Estimation of the benefits by the natura 2000 network in Flanders (in Dutch). VITO, Mol
Brotons L (2000) Winter spacing and non-breeding social system of the coal tit (Parus ater) in a subalpine forest. Ibis 142:657–667
Brown ND, Curtis T, Adams EC (2015) Effects of clear-felling versus gradual removal of conifer trees on the survival of understorey plants during the restoration of ancient woodlands. For Ecol Manag 348:15–22
Brunson MW, Reiter DK (1996) Effects of ecological information on judgments about scenic impacts of timber harvest. J Environ Manag 46:31–41
Burton NHK (2007) Influences of restock age and habitat patchiness on tree pipits anthus trivialis breeding in Breckland pine plantations. Ibis 149:193–204
Busby WH, Parmelee JR (1996) Historical changes in a herpetofaunal assemblage in the flint hills of Kansas. Am Midl Nat 135:81–91
Calcagno V, de Mazancourt C (2010) Glmulti: an R package for easy automated model selection with (generalized) linear models. J Stat Softw 34(12):1–29
Clobert J, Massot M, Lecomte J et al (1994) Lizard ecology: historical and experimental perspectives. In: Vitt LJ, Pianka ER (eds) Determinants of dispersal behavior: the common lizard as a case study. Princeton University Press, Princeton, pp 183–206
Cormont A, Malinowska A, Kostenko O et al (2011) Effect of local weather on butterfly flight behaviour, movement, and colonization: significance for dispersal under climate change. Biodivers Conserv 20:483–503
Daily GC, Ehrlich PR, Sánchez-Azofeifa GA (2001) Countryside biogeography: use of human-dominated habitats by the avifauna of Southern Costa Rica. Ecol Appl 11:1–13
De Valck J, Vlaeminck P, Broekx S et al (2014) Benefits of clearing forest plantations to restore nature? Evidence from a discrete choice experiment in Flanders, Belgium. Landsc Urban Plan 125:65–75
De Vos K, Anselin A, Vermeersch G (2004) Atlas of the breeding birds in Flanders 2000–2002 (in Dutch). In: Vermeersch G, Anselin A, Devos K (eds) A new red list for breeding birds in Flanders (version 2004) (in Dutch). Instituut voor natuurbehoud, Brussels, pp 60–75
Desender K, Dekoninck W, Maes D (2008) An updated red List of the ground and tiger beetles (Coleoptera, Carabidae) in Flanders (Belgium). Bulletin van het Koninklijk Belgisch Instituut voor Natuurwetenschappen 78:113–131
Djupström LB, Weslien J, Jt Hoopen, Schroeder LM (2012) Restoration of habitats for a threatened saproxylic beetle species in a boreal landscape by retaining dead wood on clear-cuts. Biol Conserv 155:44–49
Eycott AE, Watkinson AR, Dolman PM (2006) The soil seedbank of a lowland conifer forest: the impacts of clear-fell management and implications for heathland restoration. For Ecol Manag 237:280–289
Farren A, Prodohl PA, Laming P, Reid N (2010) Distribution of the common lizard (Zootoca vivipara) and landscape favourability for the species in Northern Ireland. Amphib Reptil 31:387–394
Fernandez-Juricic E, Zollner PA, LeBlanc C, Westphal LM (2007) Responses of nestling black-crowned night herons (Nycticorax nycticorax) to aquatic and terrestrial recreational activities: a manipulative study. Waterbirds 30:554–565
Ferrarini A, Rossi G, Parolo G, Ferloni M (2008) Planning low-impact tourist paths within a site of community importance through the optimisation of biological and logistic criteria. Biol Conserv 141:1067–1077
Ficetola GF, Sacchi R, Scali S et al (2007) Vertebrates respond differently to human disturbance: implications for the use of a focal species approach. Acta Oecol 31:109–118
Foster DR, Hall B, Barry S, Clayden S, Parshall T (2002) Cultural, environmental and historical controls of vegetation patterns and the modern conservation setting on the island of Martha’s Vineyard, USA. J Biogeogr 29:1381–1400
Fuller RJ (2013) FORUM: searching for biodiversity gains through woodfuel and forest management. J App Ecol 50:1295–1300
George SL, Crooks KR (2006) Recreation and large mammal activity in an urban nature reserve. Biol Conserv 133:107–117
Gill JA (2007) Approaches to measuring the effects of human disturbance on birds. Ibis 149:9–14
Gustafsson L, Perhans K (2010) Biodiversity conservation in Swedish forests: ways forward for a 30-Year-old multi-scaled approach. Ambio 39:546–554
Heink U, Kowarik I (2010) What criteria should be used to select biodiversity indicators? Biodivers Conserv 19:3769–3797
Hermy M, Van Der Veken S, Van Calster H, Plue J (2008) Forest ecosystem assessment, changes in biodiversity and climate change in a densely populated region (Flanders, Belgium). Plant Biosys 142:623–629
Heyman E, Gunnarsson B, Stenseke M, Henningsson S, Tim G (2011) Openness as a key-variable for analysis of management trade-offs in urban woodlands. Urban For Urban Green 10:281–293
IUSS Working Group WRB (2007) World Reference Base for Soil Resources 2006, first update 2007. World Soil Resources Reports No. 103. FAO, Rome, Italy
Jacobs S, Stevens M, Van Daele T et al (2013) Ecosystem services delivery potential-Evaluation of a method based on land use and expert knowledge in Flanders. Instituut voor natuurbehoud, Brussels
Jansen JJ, Sevenster J, Faber PJ (1996) Yield tables for the most important tree species in the Netherlands (in dutch). Landbouwuniversiteit Wageningen, Wageningen
Johst K, Drechsler M, van Teeffelen AJA et al (2011) Biodiversity conservation in dynamic landscapes: trade-offs between number, connectivity and turnover of habitat patches. J Appl Ecol 48:1227–1235
Jooris R, Engelen P, Speybroeck J et al (2012) The IUCN red list for amphibians and reptiles in Flanders (in Dutch). Instituut voor natuurbehoud, Brussels
Langston RHW, Liley D, Murison G, Woodfield E, Clarke RT (2007) What effects do walkers and dogs have on the distribution and productivity of breeding European Nightjar Caprimulgus europaeus? Ibis 149:27–36
Lens L, Dhondt AA (1994) Effects of habitat fragmentation on the timing of crested tit (Parus cristatus) natal dispersal. Ibis 136:147–152
Liddle M (1996) Recreation ecology: the ecological impact of outdoor recreation and ecotourism. Chapman and Hall, London
Linden DW, Roloff GJ (2013) Retained structures and bird communities in clearcut forests of the Pacific Northwest, USA. For Ecol Manag 310:1045–1056
Lindenmayer DB, Franklin JF, Fischer J (2006) General management principles and a checklist of strategies to guide forest biodiversity conservation. Biol Conserv 131:433–445
Lindtke D, González-Martínez SC, Macaya-Sanz D, Lexer C (2013) Admixture mapping of quantitative traits in Populus hybrid zones: power and limitations. Heredity 111:474–485
Lowe A, Rogers AC, Durrant KL (2014) Effect of human disturbance on long-term habitat use and breeding success of the European Nightjar Caprimulgus europaeus. Av Conserv Ecol 9:6
Maes D, Bonte D (2006) Using distribution patterns of five threatened invertebrates in a highly fragmented dune landscape to develop a multispecies conservation approach. Biol Conserv 133:490–499
Maes D, Ghesquiere A, Logie M, Bonte D (2006) Habitat use and mobility of two threatened coastal dune insects: implications for conservation. J Insect Conserv 10:105–115
Maes D, Vanreusel W, Jacobs I, Berwaerts K, Van Dyck H (2011) A new red list for butterflies. APPLICATION of the IUCN criteria in Flanders. (in Dutch). Nat focus 10:62–71
Mallord JW, Dolman PM, Brown AF, Sutherland WJ (2007) Linking recreational disturbance to population size in a ground-nesting passerine. J Appl Ecol 44:185–195
Mantau U, Saal U, Prins K et al (2010) Real potential for changes in growth and use of EU forests. EU Wood, Hamburg
Martin K, Eadie JM (1999) Nest webs: a community-wide approach to the management and conservation of cavity-nesting forest birds. For Ecol Manag 115:243–257
Moilanen A, Pouzols FM, Meller L et al. (2014) Spatial conservation planning framework and software: Zonation. Version 4. User manual. (4.1), Pp.1–290. BCIG, Department of Biosciences, University of Helsinki, Helsinki, Finland
Mönkkönen M, Reunanen P, Kotiaho JS et al (2011) Cost-effective strategies to conserve boreal forest biodiversity and long-term landscape-level maintenance of habitats. Eur J For Res 130:717–727
Mönkkönen M, Juutinen A, Mazziotta A et al (2014) Spatially dynamic forest management to sustain biodiversity and economic returns. J Environ Manag 134:80–89
Monz CA, Pickering CM, Hadwen WL (2013) Recent advances in recreation ecology and the implications of different relationships between recreation use and ecological impacts. Front Ecol Environ 11:441–446
Moonen P, Kint V, Deckmyn G, Muys B (2011) Scientific support of a long-term planning for wood production in Bosland (in Dutch). K.U.Leuven, Leuven
Moran-Ordonez A, Bugter R, Suarez-Seoane S, de Luis E, Calvo L (2013) Temporal changes in socio-ecological systems and their impact on ecosystem services at different governance scales: a case study of heathlands. Ecosystems 16:765–782
Morris DL, Porneluzi A, Haslerig J, Clawson RL, Faaborg J (2013) Results of 20 years of experimental forest management on breeding birds in Ozark forests of Missouri, USA. For Ecol Manag 310:747–760
Ozanne CMP, Speight MR, Hambler C, Evans HF (2000) Isolated trees and forest patches: patterns in canopy arthropod abundance and diversity in Pinus sylvestris (Scots Pine). For Ecol Manag 137:53–63
Pakkala T, Linden A, Tiainen J, Tomppo E, Kouki J (2014) Indicators of forest biodiversity: which bird species predict high breeding bird assemblage diversity in boreal forests at multiple spatial scales? Ann Zool Fenn 51:457–476
Pedley S, Bertoncelj I, Dolman P (2013) The value of the trackway system within a lowland plantation forest for ground-active spiders. J Insect Conserv 17:127–137
Phalan B, Onial M, Balmford A, Green RE (2011) Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science 333:1289–1291
Power ME, Chapin FS (2010) Planetary stewardship, with an introduction from the editor-in-chief. Bull Ecol Soc Am 91:143–175
QGIS Development Team (2015). Qgis Geographic Information System. (2.10.1). Open Source Geospatial Foundation Project
Rebbeck M, Corrick R, Eaglestone B, Stainton C (2001) Recognition of individual European Nightjars Caprimulgus europaeus from their song. Ibis 143:468–475
Reidy JL, Thompson FR III, Kendrick SW (2014) Breeding bird response to habitat and landscape factors across a gradient of savanna, woodland, and forest in the Missouri Ozarks. For Ecol Manag 313:34–46
Benayas R, Bullock (2012) Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 6:883–899
R Core Team (2013) R 3.0.1. R Foundation for Statistical Computing, Vienna, Austria
Rodriguez-Prieto I, Bennett VJ, Zollner PA et al (2014) Simulating the responses of forest bird species to multi-use recreational trails. Landsc Urban Plan 127:164–172
Schou E, Jacobsen JB, Kristensen KL (2012) An economic evaluation of strategies for transforming even-aged into near-natural forestry in a conifer-dominated forest in Denmark. For Policy Econ 20:89–98
Sharps K, Henderson I, Conway G, Armour-Chelu N, Dolman PM (2015) Home-range size and habitat use of European Nightjars Caprimulgus europaeus nesting in a complex plantation-forest landscape. Ibis 157:260–272
Simon-Reising EM, Heidt E, Plachter H (1996) Life cycle and population structure of the tiger beetle cicindela hybrida L. (Coleoptera: cicindelidae). Dtsch Entomol Z 43:251–264
Söderström B (2009) Effects of different levels of green- and dead-tree retention on hemi-boreal forest bird communities in Sweden. For Ecol Manag 257:215–222
Steven R, Pickering C, Castley JG (2011) A review of the impacts of nature based recreation on birds. J Environ Manag 92:2287–2294
Stillman RA, Goss-Custard JD (2010) Individual-based ecology of coastal birds. Biol Rev 85:413–434
Symonds MR, Moussalli A (2011) A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav Ecol Sociobiol 65:13–21
Thompson B (2015) Recreational trails reduce the density of ground-dwelling birds in protected areas. Environ Manag 55:1181–1190
Vande Walle I, Van Camp N, Perrin D et al (2005) Growing stock-based assessment of the carbon stock in the Belgian forest biomass. Ann For Sci 62:853–864
Vangansbeke P, Gorissen L, Nevens F, Verheyen K (2015a) Towards co-ownership in forest management: analysis of a pioneering case Bosland (Flanders, Belgium) through transition lenses. For Policy Econ 50:98–109
Vangansbeke P, Osselaere J, Van Dael M et al (2015b) Logging operations in pine stands in Belgium with additional harvest of woody biomass: yield, economics and energy balance. Can J For Res 45:987–997
Vangansbeke P, De Schrijver A, De Frenne P et al (2015c) Strong negative impacts of whole tree harvesting in pine stands on poor, sandy soils: a long-term nutrient budget modelling approach. For Ecol Manag 356:101–111
Verstraeten G, Baeten L, Verheyen K (2011) Habitat preferences of European Nightjars Caprimulgus europaeus in forests on sandy soils. Bird Study 58:120–129
Vihervaara P, Marjokorpi A, Kumpula T, Walls M, Kamppinen M (2012) Ecosystem services of fast-growing tree plantations: a case study on integrating social valuations with land-use changes in Uruguay. For Policy Econ 14:58–68
von Heland F, Clifton J, Olsson P (2014) Improving stewardship of marine resources: linking strategy to opportunity. Sustainability 6:4470–4496
Walker KJ, Pywell RF, Warman EA et al (2004) The importance of former land use in determining successful re-creation of lowland heath in Southern England. Biol Conserv 116:289–303
Zipper CE, Burger JA, Skousen JG et al (2011) Restoring forests and associated ecosystem services on appalachian coal surface mines. Environ Manag 47:751–765
Zolotarev MP, Belskaya EA (2015) Ground-dwelling invertebrates in a large industrial city: differentiation of recreation and urbanization effects. Contemp Probl Ecol 8:83–90
Acknowledgments
We are very grateful to all people that have helped with the data collection: Lien Poelmans (butterflies); Ruben Evens, Eddy Ulenaers and volunteers (nightjars); Johan Agten, Eddy Ulenaers and Dries Gorissen (recreation). We also want to thank the biodiversity platform of Bosland for input on the indicator species and Lander Baeten, Leen Depauw, Renato Toledo, Sanne Van Den Berge and Laura Van Vooren on the data analyses. This paper was written while PDF held a postdoctoral fellowship from the Research Foundation–Flanders (FWO). PVG performed this research in the framework of a PhD, funded by the Flemish Institute for Technological Research (VITO). We are very grateful to E. Brockerhoff, H. Jactel and two anonymous reviewers for their comments and corrections on the previous version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Eckehard Brockerhoff, Hervé Jactel and Ian Thompson.
This is part of the special issue on ‘Forest biodiversity and ecosystem services’.
Appendix
Appendix
See Figs. 4, 5, 6 and Tables 5, 6.

Classification of the forest stands based on recreational pressure. Stands in reds have the highest recreation pressure based on the number of visitors on adjacent roads (see text for details on calculation), stands in blues have the lowest recreation pressure. (Color figure online)

Habitat suitability maps for the indicator species, based on the GLMs, blues stand for a high habitat suitability, reds for a low habitat suitability. a Coal tit, b Crested tit, c Nightjar, d Small heath, e Grayling, f Northern dune tiger beetle. (Color figure online)

Zonation rank of the landscape for the different species groups, blues stand for a high conservation value for a species group, red for a low conservation value. a the forest species, b the species that depend on both forest and open habitat, c the GHS species. (Color figure online)
Rights and permissions
About this article

Cite this article
Vangansbeke, P., Blondeel, H., Landuyt, D. et al. Spatially combining wood production and recreation with biodiversity conservation. Biodivers Conserv 26, 3213–3239 (2017). https://doi.org/10.1007/s10531-016-1135-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-016-1135-5