
Abstract
We applied an intermediate disturbance-complexity approach to the land-use change of cultural landscapes in the island of Mallorca from c. 1850 to the present, which accounts for the joint behaviour of human appropriation of photosynthetic capacity used as a measure of disturbance, and a selection of land metrics at different spatial scales that account for ecological functionality as a proxy of biodiversity. We also delved deeper into local land-use changes in order to identify the main socioeconomic drivers and ruling agencies at stake. A second degree polynomial regression was obtained linking socio-metabolic disturbance and landscape ecological functioning (jointly assessing landscape patterns and processes). The results confirm our intermediate disturbance-complexity hypothesis by showing a hump-shaped relationship where the highest level of landscape complexity (heterogeneity connectivity) is attained when disturbance peaks at 50–60 %. The study proves the usefulness of transferring the concept of intermediate disturbance to Mediterranean cultural landscapes, and suggests that the conservation of heterogeneous and well connected land-use mosaics with a positive interplay between intermediate level of farming disturbances and land-cover complexity endowed with a rich bio-cultural heritage will preserve a wildlife-friendly agro-ecological matrix that is likely to house high biodiversity.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Biodiversity has been related to the existence of intermediate disturbances in ecosystems for a long time. Despite the intense debate raised by its detractors (Wilkinson 1999; Fox 2013; Sheil and Burslem 2013; Pierce 2014; Huston 2014), the intermediate disturbance hypothesis (IDH) is used in a growing number of scientific research (Svensson et al. 2012). Yet, since its introduction (Connell 1978) the IDH has hardly been applied to the socio-natural interplay or to study agricultural landscapes.
Assuming that agro-ecosystems are the result of energy flows and knowledge that farmers invest in a land matrix, the biodiversity associated to cultural landscapes (Altieri 1999) can be related on the one hand to their own complexity, and on the other hand to the degree of disturbance they exert upon natural systems. Traditional agro-ecological landscapes are endowed with an age-old bio-cultural heritage accumulated by rural communities that experienced a long-lasting joint adaptation with nature. Their maintenance are indissolubly tied to the practical knowledge handed down from one generation of farmers, shepherds and lumberjacks to the next, a complex set of ingenious techniques and local know-how that have contributed to historically compound this cultural and biological legacy. As a result, the complexity of cultural landscapes diminishes either when the farming intervention is intensified beyond a certain threshold in industrial monocultures, or abandoned (Fig. 1). Both may entail a process of landscape deterioration and biodiversity loss (Farina 2000; Antrop 2005; Agnoletti 2014).

Long-term bio-cultural heritage. Conceptual scheme of the intermediate disturbance hypothesis (IDH) in a Mediterranean cultural landscape context. Source our own
We have started to develop an intermediate disturbance-complexity (IDC) model of cultural landscapes (Marull et al. 2015a) using a multi-scalar experimental design in the island of Mallorca, at the core of the Mediterranean biodiversity hotspot (Myers et al. 2000), taking as a natural experiment the land-cover and land-use change (LCLUC) from c. 1850 to 2012. The main results of this LCLUC and their impact on landscape ecology are presented in this article. In this section we expose the aims and background of our research. Section two presents the case study and methods used. Section three discusses the results obtained and suggests a few hypotheses on the economic driving forces and socio-political agencies behind. Section four concludes.
Cultural landscapes in a globally changing world
Cultural landscapes are the historical outcome of interactions between socioeconomic and biophysical spatial patterns and metabolic flows (Wrbka et al. 2004; Liu et al. 2007; Rindfuss et al. 2008). Four decades ago pioneering work on the energy analysis of agro-ecosystems revealed a substantial decline in energy throughputs of contemporary farming, brought about by the consumption of fossil fuels and other external inputs (Odum 1984, 2007; Giampietro et al. 2011; Pelletier et al. 2011). More recently, several studies are reassessing the role traditional agrarian knowledge and practices have played to create complex-heterogeneous landscapes whose legacy is increasingly praised for its role in biological conservation (Tress et al. 2001; Kumaraswamy and Kunte 2013; Hong et al. 2014). Yet, the role of energy and material flows (Haberl 2001) as driving forces of contemporary LCLUC is still a pending research issue (Peterseil et al. 2004). We aim to contribute to the IDH research by exploring the relationships between socio metabolic impact as a proxy of human pressure, and landscape metrics that account for ecological functionality, applied to a multi-scalar analysis of LCLUC throughout socio-ecological transitions (Fischer-Kowalski and Haberl 2007; González de Molina and Toledo 2014).
LCLUC is a global factor of biodiversity loss that poses significant land-use policy questions (Schroter et al. 2005; Young et al. 2014), and challenges scientific research to develop better models and indicators (De Groot 2006; Turner et al. 2007; Haines-Young 2009). In turn, landscape ecology provides quantitative tools to characterize landscapes (Turner and Ruscher 1988; Li 2000) and land-use change (Reed et al. 1996) by linking ecological patterns and processes (Tischendorf 2001; Helming et al. 2007; Verburg et al. 2009). However a considerable disagreement still remains on whether the removal of human intervention in landscapes undergoing an abandonment process results in a positive impact on biodiversity conservation (as seen from a land sparing or a forest transition approach) or rather a negative one (as seen from a land sharing and a wildlife-friendly farming approach) (Green et al. 2005; Matson and Vitousek 2006; Bengston et al. 2003; Fischer et al. 2008; Perfecto and Vandermeer 2010; Tscharntke et al. 2012). According to Robson and Berkes (2011), land-use decline may result in a loss of agro-forest mosaics and to local biodiversity decrease. A meta-analysis made by Plieninger et al. (2014) founds some patterns linking biodiversity and land abandonment in the Mediterranean, but they seem too complex to draw definite conclusions.
Exploring this bio-cultural interface is an exciting and pressing scientific challenge (Phalan et al. 2011) that calls for a better understanding on how farm systems affect the relationship between farming land-uses, biological primary productivity and landscape functionality. A useful indicator is the human appropriation of net primary production (HANPP), a top-level indicator of environmental pressure (Vitousek et al. 1986; Haberl et al. 2007; Krausmann et al. 2013) that can assess the impact of farming on biodiversity (Firbank et al. 2008) according to the species-energy hypothesis (Hawkins et al. 2003). Although mathematical modelling suggests that the output of ecosystem services generally peaks at some intermediate level of LCLUC intensity (Braat and ten Brink 2008), this is rather complex interplay. Schwartz et al. (2000) found little support to establish a linear relationship between biodiversity and ecosystem functioning (i.e., biomass, nutrient cycling, etc.), while Balvanera et al. (2006) suggested the contrary from a meta-analysis on different biodiversity components that corroborate the basic scientific consensus and the remaining uncertainties on the subject (Hooper et al. 2005).
We consider that simple gradients of LCLUC are unable to explain the variations in biodiversity, unless the functional ecological complexity of landscapes is taken into account (Opdam et al. 2006; Pino and Marull 2012; Marull et al. 2014, 2015b). It is known that landscape heterogeneity arises in nature as one among many looping ways through which energy dissipation leads to the formation of self-organized structures, able to perform a historical succession ruled by adaptive selection (Morowitz 2002). When humans increase the dissipated energy up to a critical point, complexity is reduced and environmental degradation ensues (Ulanowicz 1997). In complex agro-ecosystems, instead, the storage of energy and information at some points reduces internal entropy thanks to the exploitation of other spaces of lower complexity but larger production within a joint encompassing structure (Margalef 2006). As in other living organisms, these heterogeneous space–time structures may allow keeping more mature organized spaces linked together with simpler productive ones within an interdependent set of patterns and flows able to provide resilience to the system (Ho and Ulanowicz 2005).
Disturbance ecology in cultural landscapes
The intermediate disturbance hypothesis (IDH) is a non-equilibrium explanation to understand the maintenance of biodiversity in ecosystems (Wilson 1990). Yet, there is considerable debate around which are the mechanisms that promote coexistence among species (Dial and Roughgarden 1998; Buckling et al. 2000; Sheil and Burslem 2003; Miller et al. 2012; Fox 2013; Huston 2014). There are different definitions of disturbance (van der Maarel 1993), but a common one is the destruction (or harvest) of biomass (Calow 1987) leading to the opening up of space and resources for recolonizing species—an approach that foregrounds the variation of its spatial extent in ecosystem communities (Wilson 1994). The earliest version by Hutchinson (1951) already considered disturbance intensity in a spatial context, that led to the idea of a humped-shaped trend later introduced by Horn (1975) and further amplified by Connell (1978). Coexistence would require spatially patchy disturbance that leads to a trade-off between species able to perform best at different stages of post-disturbance succession (Chesson and Huntly 1997). At intermediate disturbance frequencies both competitive and dispersal species may coexist (Roxburgh et al. 2004; Shea et al. 2004; Barnes et al. 2006). Wilson (1994) labelled it a between-patch mechanism (Collins and Glenn 1997), which has been renamed as a succession-mosaic hypothesis that views disturbances as events that alter niche opportunities (Shea and Chesson 2002).
Whereas IDH has been evaluated by mathematical modelling (Petraitis et al. 1989), and widely supported in studies of terrestrial (Molino and Sabatier 2001), freshwater (Padisak 1993) and marine communities (Johst et al. 2006), it has been seldom used in agro-ecosystem so far (Gliessman 1990; Fahrig and Jonsen 1998; Sasaki et al. 2009). Yet, if IDH holds true in natural ecosystems, it should play a similar role in the interplay of human activity with ecological processes (Farina 2000). Agro-forest mosaics offer habitats to different species, creating a greater amount of ecotones which in turn provide opportunities to edge species (Benton et al. 2003), as well as more permeable land-matrix allowing dispersion among local populations (Shreeve et al. 2004). Thanks to the edge effect and high connectivity, a complex land-cover pattern may host greater biodiversity than more uniform landscapes (Harper et al. 2005). Understanding and managing correctly these patchy agro-forest mosaics require an interdisciplinary approach to the bio-cultural diversity (Arts et al. 2012; Parrotta and Trosper 2012; Cocks and Wiersum 2014) embedded in agro-ecological landscapes (Antrop 2006; Matthews and Selman 2006; Blondel 2006; Verdasca et al. 2012).
In order to create and maintain agro-ecosystems, farmers have to continuously invest over the land matrix certain amounts of energy and information that shape the spatial patterns of an agro-ecological landscape embodied with a bio-cultural heritage (Marull et al. 2015c). The impact of this farming ecological disturbance (Margalef 2006) on biodiversity may be either positive or negative, depending on the intensity and shape of these socio-metabolic flows and the complexity of landscape mosaics (Altieri 1999; Swift et al. 2004; Cardinale et al. 2012).
Materials and methods
A multi-scalar experimental design of the study area
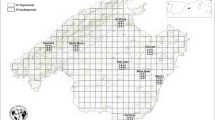
In the Mediterranean World, wilderness was early disturbed by human action. Since Ancient times, farmers and shepherds have long shaped the land with agroforest and grazing mosaics (Grove and Rackham 2003). The starting point of our case study is not from a pristine wilderness but a much transformed nature (Gil-Sánchez et al. 2002). The island of Mallorca, located in the Mediterranean Sea (Fig. 2), has an extension of 3603 km2 of calcareous origin. The coast combines sand beaches with cliffs raised by a mountain range that runs parallel to the North coast, the Serra de Tramuntana, and the eastward Serres de Llevant. Between them there is a great plain with a Mediterranean mild climate. Annual precipitation ranges from 300 mm in the South to 1800 mm in the North, largely concentrated in winter, while the average annual temperature is around 16 °C and peaks during the dry summers. The island vegetation, adapted to these agro-climatic features as well as to a long-lasting human intervention (Murray 2012), combines scrubland, pines and residual oak forests with a variety of annual crops (grains and vegetables) and arboriculture (olive groves, almonds, figs, carobs, vineyards).

Location of the Mallorca case study performed at three scales: SF-1 (1:50,000), SF-2 (1:5000), SF-3 (1:500). Source our own
There are six regions in Mallorca (Rullan 2002) with different traits (Fig. 2): (i) Tramuntana comprises all the northern mountains, with an abrupt morphology and a rainfall of 1400–1800 mm a year (the 3 × 3 km2 studied area is ‘Esporles’ scene); (ii) Raiguer is the piedmont between Tramuntana and the inland plane, whose soil, precipitation and edge character provide the best conditions for an intensive and diversified agriculture (the 3 × 3 km2 study area is ‘Santa Maria’ scene; next to Raiguer we find ‘Sa Pobla’ scene characterized by its drying works of wetlands and watering intensification); (iii) The Pla is a central plane where cereal crops have been most cultivated (we take the 3 × 3 km2 ‘Sant Joan’ scene); (iv) Llevant is located eastward and combines relative small elevations with valleys that contribute to its rich landscape diversity, representative of all Mallorca landscapes, including flat grain-growing zones, agro-forest mosaics in the hills and areas of shallow soil and arid vegetation (we set three 3 × 3 km2 scenes: ‘Albocàsser’, quite similar to ‘Sant Joan’; ‘Calicant’, similar to ‘Esporles’; and ‘Marina’ similar to the Migjorn region); (v) Migjorn, in the Southeast, is the driest region with barren land with shrubs that hinders agriculture (the 3 × 3 km2 scene is ‘Santanyí’).
This set of scenes allows us to gain in-depth insights that might be lost in the broader view of the whole island. In order to test the relationship between HANPP and ecological patterns and processes taking place in these cultural landscapes, we used the following multi-scalar experimental design: (1) regional scale (SF-1; 1:50,000) takes into account the entire island divided into 3 × 3 km2 cells (Fig. 2), and to avoid the sea edge effect the analysis area is limited to 331 inland cells studied in three time points (1956, 1973, 2000) using land-cover digital cartography (GIST 2009); (2) landscape scale (SF-2; 1:5000) takes into account eight 3 × 3 km2 analysis scenes distributed in five agro-ecological regions of Mallorca divided into nine 1 × 1 km2 cells (Fig. 2), so as to have a better approximation to the landscape transitions along three time points (1956, 1989 and 2010); and (3) local scale (SF-3; 1:500) takes into account three 3 × 3 km2 analysis scenes (Fig. 2) in the Llevant region, as a representative sample of Mallorca landscapes, dividing each scene into 36 cells of 0.5 × 0.5 km2 and extending backwards the time frame from the 1850s to 1956 and 2012 using land-cover cartography digitized from historical land-use maps.
This multi-scalar dataset will be used to test in Mallorca the hypothesis that landscape heterogeneity in a well-connected land matrix could potentially host greater biodiversity than in the more uniform land-covers we tend to have at present. This hypothesis has already been tried out for different species and ecosystems (Bengtsson et al. 2003; Tscharntke et al. 2012; Gabriel et al. 2013). The novelty is to apply this to cultural landscapes, by adopting a bio-cultural approach that relates the farming disturbance exerted through HANPP to the landscape ecology assessment of land-use patterns.
Assessing HANPP and land-cover change at three different scales
Based on the digital maps available for the whole island in 1956, 1973, 1989 and 2000 provided by GIST (2009), we have analysed the historical shifts in land-cover patterns of the study area (SF-1; Fig. 3) by using the metrics listed and explained in Table 1. Also relying on photointerpretation of the landscape scenes (SF-2; Fig. 6), we analysed in 1956, 1989 and 2011 the ecological landscape patterns listed and explained in Table 2. After digitising some of the cadastral land-use maps available at local scale (SF-3; Fig. 8) from historical archives (Rosselló-Verger 1982), we analysed the corresponding shifts in land-use patterns calculated per parcel and/or within 0.5 × 0.5 km2 sample cells for three study areas located in the Manacor municipality (‘Albocàsser’, ‘Callicant’ and ‘Marina’) c. 1850, in 1956 and 2012 by using the metrics listed and explained in Table 3.

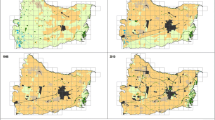
Land-cover changes at regional scale (SF-1; 1:50,000) in 1956, 1973, 2000. Source our own, from GIST (2009)
Our intermediate disturbance hypothesis (IDH) is based on variables that describe both spatial land pattern (Shannon–Wiener index, H′) and human disturbance (human appropriation of net primary production, HANPP). We work with squared cells from land-unit (LU) maps, so that:
where p i is the proportion of LU i in a specific cell, and k is the number of LU. We will refer to p as vector p = (p1,…pk).
In order to check the IDH with the historical LU maps available, we analysed the corresponding change in the spatial pattern of the study area by using H′ (Shannon 1948) that measures equi-diversity of LU in a cell:
where k is the total number of LU in the study area, and is the proportion of LU i in a specific cell.
HANPP is used as a measure of disturbance, where NPP is the net amount of biomass produced by autotrophic organisms (green plants) that constitutes the main nutritional basis for all food chains over a year. HANPP measures the extent to which humans modify the amount of NPP available for other species, either by changing the land-covers or removing a share of NPP (Haberl et al. 2007; Krausmann et al. 2013). Hence, HANPP is calculated using the following identities:
where NPP h is the NPP appropriation through harvest, and ΔNPP LU is the change of NPP through human-induced land conversions. ΔNPP LU is defined as the difference between the NPP of the potential (NPP 0 ), and actual (NPP act ) vegetation. HANPP is associated to each LU of the study area, so that HANPP is calculated multiplying a fixed coefficient (w i ) for some LU i by the surface occupied by this LU:
where denote the weight of LU i. Variations in HANPP not only depend on the variations of p, but on the variations of w as well. As a result we have spatially-explicit values of H′ and HANPP for each cell measured on the same LU database. Taking as reference the work done by Schwarzlmüller (2009) on Spain, these HANPP values have been estimated after assessing different NPP and harvested amounts (in tonnes of dry matter per LU and year).
In the work presented here bio-cultural diversity is represented in the land matrix and not in the species richness. Recent studies in Mediterranean cultural landscapes reveal that the conservation of heterogeneous and well-connected land matrix with a positive interplay between human disturbances and land-cover/land-use complexity are able to hold high species richness at regional scale (i.e. birds; Marull et al. 2015b), landscape scale (i.e. orchids; Marull et al. 2014) and local scale (i.e. butterflies; Marull et al. 2015a). In order to test our hypothesis at the regional scale, we analyse a set of landscape ecology metrics as a function of HANPP. To do this, we obtain a new variable L (‘Landscape Metrics’ as a proxy of biodiversity) using principal components analysis (PCA). Once we have L, we will perform a regression analysis with HANPP as the independent variable and L as the dependent.
Results and discussion
Land-cover dynamics at regional scale (SF-1)
Despite the seemingly low land-cover change seen from a regional view (Fig. 3), landscape metrics show a decrease from 1956 to 2000 as the joint result of urban sprawl, agricultural intensification and rural abandonment (Fig. 4). Urban areas (277 %) and golf courses (1796 %) increased the most. Agricultural covers decreased, mainly in dry crops (−8.8 %), dry groves (−4.3 %) and olive trees (−9.6 %). Shrubs (−3.6 %), woodland (−4.5 %) and wetlands (−5.2 %) experienced a lesser decrease, while irrigated cropland grew 14.6 % (Table 4).

Metrics applied at regional scale (SF-1): MLC main land cover, LCR land cover richness, H’ Shannon–Wiener index, MESH effective mesh size, LMI landscape metric index, ECI ecological connectivity index in 1956, 1973, 2000. a Land-cover change, b Land-cover structure, c Land-cover functionality, Source our own
Accordingly, the number of patch types per cell (LCR) tended to diminish. Land-cover richness (H′) measured by the number of different patch types and their proportional area distribution (richness and evenness), presented lower values as well—strongly correlated with MESH values as the inverse of fragmentation. LMI values confirm the progressive loss of landscape functional structure, thus lessening its capacity to support ecological processes and likely biodiversity. ECI values of landscape ecological connectivity also decreased (Figs. 4, 5) due to the impact of new transport facilities and low-density urban developments. Urban sprawl has isolated woodland, cropland and natural protected areas one another, while the retreat of farming decreased landscape diversity and ecotones. Taken together these metrics indicate a loss in landscape heterogeneity that would ultimately lead to lesser biodiversity. Some critical areas for the potential ecological connectivity between protected natural areas and the remaining agricultural mosaics can be detected in Fig. 5, which should be preserved from the barrier effect of linear infrastructures and urban developments in future.

Ecological connectivity index (ECI) at regional scale (SF-1) in 1956, 1973, 2000. Source our own
Transitions seen at landscape scale (SF-2)
The aerial photointerpretation highlights three main landscape changes from 1956 to 1989 and 2011 in the eight scenes (Fig. 6): abandonment of rain-fed arboriculture (almond groves change to cereals; olive groves change to woodland); spontaneous reforestation following the abandonment of forestry uses (charcoal making, wood pasture, etc.); and urban sprawl (mainly tourism in coastal areas and new inland urban developments in former farm dwellings). The traditional integrated polycultures tended to be replaced by disjoint patch units of grassland, woodland, cropland and urban covers, that in most cases have led to a higher number of possible land-uses in a cell—e.g. in the ‘Sant Joan’ scene. In others, the predominant trend has been towards more uniform land-covers—as the loss of land-use diversity driven by tourist urbanization in the ‘Marina’ scene. In all cases this polarization has tended to the vanishing of the former landscape mosaics.


Transitions at landscape scale (SF-2; 1:5000) from 1956 to 1989 and 2011. a Albocàsser, Calicant, Esporles and Marina landscape scenes b Sa Pobla, Sant Joan, Santa Maria and Santanyí landscape scenes Source our own
These contrasting trends of land-use intensification and abandonment have taken place along different scales and periods, as landscape metrics help to reveal (Fig. 7). Less than a quarter of the sample cells have experienced low degrees of land-cover change along the period 1956–2011. Yet during the first phase from 1956 to 1989, there were more land-use changes mainly driven by the green revolution in farm management and mass tourism in the coast. After 1989, the main drivers were rural abandonment ensuing Spanish entry to the EU (1986) and a new inward-oriented urban sprawl. These differences are shown in the rising values of land pressure (LP) and human-altered landscapes (LA) during the first phase, and the polarization trend towards either low and high levels of pressure (LP) or naturalness (LN) together with increasingly homogenised levels of human-altered landscapes (LA) in the second phase.

LD landscape dynamics, LP landscape pressure, LN landscape naturalness, LA landscape anthropogeneity assessed at landscape scale (SF-2) from 1956, to 1989 and 2011. a Landscape transitions, b Landscape naturalness Source our own
In ‘Santanyí’ and ‘Marina’ the loss of cultivated groves at the expense of urban developments was lower, and former rangelands were substituted by scrubland (in the southwest angle of ‘Santanyí’ an unchanged area appears which corresponds to a single big estate). In ‘Esporles’, in the Tramuntana mountains, the land-cover changed from olive groves to pine forest. In ‘Santa Maria’, in the Raiguer, dry groves predominated and are still found despite the proliferation of isolated houses and reforestation. Due to the lack of replacement of dead almond, carob and fig trees, arboriculture has been lost in ‘Calicant’, although an interesting landscape mosaic remains there except in the reforested hills. The plain areas of ‘Albocàsser’ and ‘Sant Joan’ have evolved from a polyculture of dry groves combined with rain-fed crops to a cereal monoculture devoid of tree cover, while some abandoned cropland and grazing areas have been conquered by woods. In ‘Sa Pobla’ irrigated land remained unchanged except by the growing number of dwellings and small wetlands. The maintenance of shelterbelts is also noticeable (Fig. 6).
Land-use patterns at local scale (SF3)
The closest approach allows us to capture finer relationships between land-use changes, ownership regimes and socioeconomic drivers of landscape change. We can observe in the three local scenes of Manacor municipality the expansion of dry polycultural groves from c. 1850 to 1956, at the expense of rain-fed arable land, woodland and scrubs (Fig. 8; Table 5). This happened as a result of the financial and political crisis of the old large estates (the so-called possessions) during the second half of the nineteenth century and the first two decades of the twentieth, which opened up a process of land parcelling allotted to small peasants offering them an option to make a living with a labour-intensive farming (Suau 1991; Manera 2001). The allotment process is more clearly shown in Albocàsser than in the mountainous area of Calicant, and even more than in Marina due to poor soils and aridity (Table 5), but everywhere crop diversity increased with the extent of landownership (Table 6). Not only leguminous carobs, but also almond and fig trees were grown in association with cereals and legumes, and even caper plants were grown in summer at the foot of the trees in the whole island (Bisson 1977). These multi-cropping groves of almonds and carobs grew from 6048 and 7789 ha in 1860 to 47,560 and 21,875 ha in 1930 respectively (Urech y Cifre 1869; Cela Conde 1979).

Land-use changes at local scale (SF-3; 1:500) in Albocàsser, Calicant and Marina scenes of the Manacor municipality in c. 1850, 1956 and 2012. Source our own
Thanks to smallholders’ work and inventiveness, that took advantage of the growing international demand for almonds, capers, potatoes, dried fruits (figs, apricots) and vegetables (Manera 2001), there was a shift towards complex agro-forest mosaics of higher diversity—as shown in the landscape metrics of these three scenes (Fig. 10). Values of land-use richness (LUR), edge density (ED) and polygon density (PD) increased while large patch index (LPI) decreased from c. 1850 to 1956, reflecting the greater land-cover diversity and ecotones of those multi-cropping mosaics interwoven with woods and pastures. Conversely, from 1956 to 2012 these scenes confirm the trend towards the disappearance of polycultural landscapes (Fig. 10; Table 5) already observed at larger scales.
This local scale also reveals that up to the present the withdrawal of farmer’s labour and knowledge has been only partial in Mallorca. The average or high values of LUR, land-cover diversity and ecotones (ED, PD) attained in 1956 are still found at present. This feature highlights the need to delve deeper into the socioeconomic drivers and ruling agencies behind this socio-ecological transition—a task which requires another forthcoming article whose main interpretive lines are outlined in the following “Driving forces and ruling agencies of socio-ecological change” section.
Human disturbance and landscape complexity in cultural landscapes
To conclude our intermediate disturbance analysis, we studied the statistical relationships between HANPP and all the landscape ecology metrics used as proxy for biodiversity, in the set of cells of our experimental design at regional scale. The high correlations (Table 7a) among land-cover metrics (H′, MESH, ECI, LMI and LCR) aim us to carry out a PCA. Hence, we performed a PCA of the variables involved (Table 7b) that shows that the major contributors for the first component (C1) are H’′ and MESH; and for the second component (C2) are ECI and LMI. LCR goes alone in all dimensions. These results have led us to consider a PCA taking only two variables, H′ and ECI, so that the two first dimensions are represented—which include patterns as landscape heterogeneity, and processes by means of ecological connectivity. Once we have reduced the dimensions of the land-cover metrics, we obtain a component resulting of the linear combination of H’ and ECI (component coefficient = 0.707; explained variance = 65 %). We call this new H’—ECI component ‘Landscape Metrics’ (L).
Figure 9 shows the results of a quadratic regression analysis, where HANPP is the independent variable that influences L as a proxy of landscape’s ecological patterns and processes (Table 7). In all time periods we obtain a second degree polynomial regression linking the two sets of data (socio-metabolic disturbance and landscape ecological functioning), that confirms our intermediate disturbance-complexity hypothesis (IDC) by showing a hump-shaped relationship where the highest level of landscape complexity (heterogeneity-connectivity as biodiversity proxy) is attained when HANPP peaks at 50–60 %. The time factor should not affect the relationship between variables, given that the IDC hypothesis represented in the non-lineal regression does not depend on time. By changing the perspective from regional to local scale, the results found in the three Manacor scenes (Figs. 10, 11) confirm that the historical trend that attained the highest land-cover diversity (H’) in 1956 was also linked to shifts in HANPP values. Yet the relationship seems to be more differentiated locally, which calls for a further geo-historical study of this complex interplay between biological and cultural factors.

Relationship between Landscape Metrics (L) and Human Appropriation of Net Primary Production (HANPP) at regional scale (SF-1) in 1956, 1973 and 2000. a 1956 b 1973 c 2000 Source our own. Results of the quadratic regression analysis, where HANPP is the independent variable that influences L as proxy of ecological patterns and processes (Table 7)

Landscape metrics applied at local scale (SF-3) in Albocàsser, Calicant and Marina scenes of the Manacor municipality in c. 1850, 1956 and 2012: LUR land use richness, LPI largest patch index, ED edge density, PD polygon density. Source our own

Shannon–Wiener Index of land-cover diversity (H’) and Human Appropriation of Net Primary Production (HANPP) applied at local scale (SF-3) in Albocàsser, Calicant and Marina scenes of the Manacor municipality in c. 1850, 1956 and 2012. Source our own
Driving forces and ruling agencies of socio-ecological change
From Middle Ages onwards (Jover and Soto 2002; Soto 2015) the agrarian change in the island was driven by the conflicting relationship between large estates (possessions) that hoarded most of the land, and peasant smallholders of tiny plots confined in the outskirts of the inner villages—who, in turn, supplied the wage labour hired to farm big estates. While the landowners practised extensive land usages and an export-oriented farm management (with olive oil trade as the main commercial driver), small peasants’ farming was highly intensive, diversified, and household or locally oriented (Bisson 1997, Manera 2001). In order to prevent a rise of agricultural wages as a result of a reduction of farmhands’ supply, big landowners tried to restrain the advance of those peasant land belts of intensive poly-culture, until they went bankrupt in the nineteenth century (Jover and Manera 2009). The parcelling of many large estates from the 1860s to the 1920s entailed a significant change in the cultural landscapes kept by this dual agrarian class structure (Cela Conde 1979; Rosselló-Verger 1982). Thus, and foremost, the wonderful ‘traditional’ landscapes which attracted elite visitors to Mallorca, from George Sand and Frederic Chopin (1838-39) to the Archduke Ludwig Salvator von Habsurbg-Lorena (1847-1915) who wrote a famous nine-volume treatise on the Balearic Islands, were to a large extent a relatively recent creation of small peasants who made advances in the age-old fight to have access to the land.
Tourism development of Mallorca from the elites of the Belle Époque up to the mass invasion of sun-and-sea holidaymakers has cast a Midas curse. Urban sprawl extended from coastal hotels to inland houses built in former rural dwellings, together with the highways linking them, which jointly entailed a growing environmental impact that tended to destroy the same landscape beauty that led Mallorca to become a tourist destination known worldwide (Pons et al. 2014). Developed land multiplied by 3.8 from 1956 to 2000, and doubled after 1973, as seen in Fig. 3 and Table 4 (Murray 2012). Yet the impact of tourism on the island’s agriculture has been twofold. On the one hand it has entailed a strong socioeconomic marginalisation of farming, leading to rural abandonment—with the usual ecological impacts such as wildfires (Gil-Sánchez et al. 2002) and disruption of complex dry stone hydraulic systems (Estrany et al. 2010). On the other hand, this effect started so early that, after the halt of Franco’s autarky (Naredo 2004), the intensification of farm and livestock management following the green revolution lines was tempered to some extent—with the usual outcomes of monocultures, soil degradation and water pollution (Roca 1992). Our SF-2 assessment shows that industrialization of agriculture left a clear imprint in the evolution of cultural landscapes mainly during the 1956 to 1989 period. But it was comparatively soft in regard to what happened in other parts of the Mediterranean basin, such as the province of Barcelona in Catalonia (Marull et al. 2010).
Three factors may explain the relatively high resilience (Marull et al. 2015b) of the cultural landscapes that peasants created in Mallorca before traditional organic farming ended. First, the commitment of local population that kept buying foodstuffs grown on the island (many years before the zero-km and slow food movements began) helped to maintain a precarious part-time agriculture that sought a compromise between traditional-organic and industrial farm managements. Second, following the Spanish EU membership in 1986 the main socioeconomic driver was rural abandonment that pushed towards relying on the increasing amount of imported food (Murray 2012). Small farms have been maintained mostly thanks to the hard work of non-professional peasants who have remained attached to the land for cultural and emotional reasons. The ageing of this group is one of the most important threats for bio-cultural preservation currently (Binimelis and Ordines 2008). In spite of this, the esteem of the local population for their food, tastes and landscapes was reinforced from then on by the growing environmental movement (Rayó 2004) led by the Grup d’Ornitologia Balear (GOB). Together with the EU environmental directives, this social pressure became a third factor that helped to preserve some natural sites and restrain urban sprawl to some extent—despite the ambiguous and shifting policies adopted by the autonomous and Spanish governments (Rullan 2010).
Not only the agricultural landscape and traditional peasant knowledge are currently threatened by low incomes and lack of farmers’ replacement, but also the rich diversity of local species varieties as well (Socies 2013). The entire bio-cultural heritage of the Mallorca Island is at stake. Last but not least, a local turning towards organic farming is on the way. Its promoters are younger and with a higher education than old peasants, and the shift towards high-quality foodstuffs can help to increase farming incomes—provided that consumers are willing to pay for them, and public policies are reoriented to foster local organic food instead of promoting tourism and urban developments at the expense of farming as it currently does. Despite the lack of political support, organic food is growing thanks to the efforts of small peasants and social movements. If there is a sustainable future for a cultural landscape able to hold a high biodiversity in Mallorca, this clearly belongs to the role of organic farming as heir of the rich bio-cultural heritage of this beautiful Mediterranean island (Alcover et al. 2003).
Conclusion
An intermediate-disturbance conceptual approach has been applied to the land-use changes of cultural landscapes underwent in the island of Mallorca from c. 1850 to the present. It accounts for the joint multi-scalar behaviour of human appropriation of photosynthetic capacity (HANPP) and landscape heterogeneity. We obtained a second-degree polynomial regression linking HANPP with landscape ecological functioning, jointly assessed by Shannon Index (H’) of land-cover patterns and ecological connectivity (ECI) of landscape processes, which confirms our intermediate disturbance-complexity hypothesis. As far as we know, few authors have studied the relationship between these variables, or other similar ones (Wrbka et al. 2004; Haberl et al. 2005; Vackar et al. 2012).
The results found show the usefulness of transferring the concept of intermediate disturbance to agro-ecological landscapes (Gliessman 1990; González de Molina and Toledo 2014), and suggest that rural development and land-use planning policies should consider the territory as a whole instead of applying a string of ad hoc decisions on minor parts of cultural landscapes as usual (Rullan, 2010; Agnoletti 2014). The historical landscape analysis performed and the driving forces described show that traditional farming played a crucial role in shaping and maintaining a complex set of land-use mosaics. Our results suggest that a great deal of the biodiversity currently existing in Mallorca may actually be associated to the remaining agricultural and forest mosaics still worked by the local peasantry. We deem that the keeping of this bio-cultural heritage may underlie the hump-shaped relationship we have found between HANPP and landscape ecological functionality jointly assessed with land-cover diversity and ecological connectivity -a result that fits with the intermediate disturbance hypothesis. Protecting natural spaces but at the same time allowing their isolation by the spread of anthropogenic barriers that decrease ecological connectivity will eventually lead to a biodiversity loss in the whole land matrix (Pino and Marull 2012). Conversely, the conservation of heterogeneous and well-connected landscapes with a positive interplay between intermediate level of farming disturbances and land-use complexity would preserve a wildlife-friendly agro-ecological matrix that is likely to hold a great biodiversity—perhaps with the exception of rare specialist species that require some specific habitats and other conservation policies (Loreau 2000; Tscharntke et al. 2012).
References
Agnoletti M (2014) Rural landscape, nature conservation and culture: some notes on research trends and management approaches from a (southern) European perspective. Landsc Urban Plan 126:66–73
Alcover JM, Pons GX, Palmer M (2003) Biodiversitat i Societat. In: Duarte C, Grasses F (eds) El papel social de la ciencia en Baleares: un homenaje a Javier Benedí. Llibre homenatge a Javier Benedí, Universitat de les Illes Balears, Balears, pp 111–130
Altieri M (1999) The ecological role of biodiversity in agroecosystems. Agric Ecosyst Environ 74:19–31
Antrop M (2005) Why landscapes of the past are important for the future. Landsc Urban Plan 70:21–34
Antrop M (2006) Sustainable landscapes: contradiction, fiction or utopia? Landsc Urban Plan 75:187–197
Arts B, van Bommer S, Ros-Tonen M, Verschoor G (2012) Forest-people interfaces: understanding community forestry and biocultural diversity. Wageningen Academic Publishers, Wageningen
Balvanera P, Pfisterer AB, Buchmann N, He J-S, Nakashizuka T, Raffaelli D, Schmid B (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol Lett 9:1146–1156
Barnes B, Sidhu HS, Roxburgh SH (2006) A model integrating patch dynamics, competing species and the intermediate disturbance hypothesis. Ecol Model 194:414–420
Bender DJ, Contreras TA, Fahrig L (1998) Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79:517–533
Bengtsson J, Angelstam P, Elmqvist T, Emanuelsson U, Folke C, Ihse M, Moberg F, Nyström M (2003) Reserves resilience and dynamic landscapes. Ambio 32(6):389–396
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Binimelis J, Ordines A (2008) La pagesia illenca als albirs del segle XXI. Agricultura i postproductivisme a les Illes Balears, El Gall
Bisson J (1977) L’homme et la terre aux îles Baléares. Edisud, Aix -en-Provence
Blondel J (2006) The ‘Design’ of mediterranean landscapes: a millennial story of humans and ecological systems during the historic period. Hum Ecol 34(5):713–729
Braat L, ten Brink P (2008) The cost of policy inaction. The case of not Meeting the 2010 biodiversity target. Study/Report for the European Commission, DG Environment
Buckling A, Kassen R, Bell G, Rainey PB (2000) Disturbance and diversity in experimental microcosms. Nature 408:961–964
Calow P (1987) Evolutionary physiological ecology. Cambridge University Press, Cambridge
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature 486:59–67
Cela Conde CJ (1979) Capitalismo y campesinado en la isla de Mallorca. Siglo XXI, Madrid
Chesson P, Huntly N (1997) The roles of disturbance, mortality, and stress in the dynamics of ecological communities. Am Nat 150:519–553
Cocks ML, Wiersum F (2014) Reappraising the concept of biocultural diversity: a perspective from South Africa. Hum Ecol 42(5):727–737
Collins SL, Glenn SM (1997) Intermediate disturbance and its relationship to within- and between-patch dynamics. N Z J Ecol 21:103–110
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310
De Groot R (2006) Function-analysis and valuation as a tool to assess land use conflicts in planning for sustainable, multi-functional landscapes. Landsc Urban Plan 75:175–186
Dial R, Roughgarden J (1998) Theory of marine communities: the intermediate disturbance hypothesis. Ecology 79:1412–1424
Estrany J, Garcia C, Batalla RJ (2010) Hydrological response of a small mediterranean agricultural catchment. J Hydrol 380(1–2):180–190
Fahrig L, Jonsen I (1998) Effect of habitat patch characteristics on abundance and diversity of insects in an agricultural landscape. Ecosystems 1(2):197–205
Farina A (2000) The cultural landscape as a model for the integration of ecology and economics. Bioscience 50:313–320
Firbank LG, Petit S, Smart S, Blain A, Fuller RJ (2008) Assessing the impacts of agricultural intensification on biodiversity: a British perspective. Philos Trans R Soc B 363:777–787
Fischer J, Lindenmayer DB (2007) Landscape modification and habitat fragmentation: a synthesis. Glob Ecol Biogeogr 216:265–280
Fischer J, Brosi B, Daily GC et al (2008) Should agricultural policies encourage land sparing or wildlife-friendly farming? Front Ecol Environ 6(7):380–385
Fischer-Kowalski M, Haberl H (eds) (2007) Socioecological transitions and global change. Trajectories of social metabolism and land use. Edward Elgar, Cheltenham
Forman RTT (1995) Land Mosaics. The ecology of landscapes and regions. Cambridge University Press, Cambridge
Forman RTT, Godron M (1986) Landscape ecology. Wiley, New York
Fox JW (2013) The intermediate disturbance hypothesis should be abandoned. Trends Ecol Evol 28(2):86–92
Gabriel D, Sait SM, Kunin WE, Benton TM (2013) Food production vs. biodiversity: comparing organic and conventional agriculture. J Appl Ecol 50:355–364
Giampietro M, Mayumi K, Sorman AH (2011) The metabolic pattern of societies: where economists fall short. Routledge, Oxon
Gilbert-Norton L, Wilson R, Stevens JR, Beard KH (2010) A meta-analytic review of corridor effectiveness. Conserv Biol 24:660–668
Gil-Sánchez L, Valdés CM, Díaz-Fernández P (2002) La transformación histórica del paisaje forestal en las islas Baleares. Ministerio de Medio Ambiente, Madrid
GIST (2009) Mapes de cobertes del sòl de les Illes Balears (1:25.000): 1956(1973), 1995, 2000. Universitat de les Illes Balears, Departament de Ciències de la Terra, Grup d’Investigació de Sostenibilitat i Territori, Palma de Mallorca
Gliessman SR (ed) (1990) Agroecology: researching the ecological basis for sustainable agriculture. Springer, New York
González de Molina M, Toledo V (2014) The social metabolism. A socio-ecological theory of historical change. Springer, New York
Green RE, Cornell SJ, Scharlemann JPW, Balmford A (2005) Farming and the fate of wild nature. Science 307:550–551
Grove AT, Rackham O (2003) The nature of Mediterranean Europe: an ecological history. Yale University Press, New Haven
Haberl H (2001) The energetic metabolism of societies. part I: accounting concepts. J Ind Ecol 5:107–136
Haberl H, Plutzar C, Erb KH, Gaube V, Pollheimer M, Niels B, Schulz NB (2005) Human appropriation of net primary production as determinant of avifauna diversity in Austria. Agric Ecosyst Environ 110:119–131
Haberl H, Erb KH, Krausmann F, Gaube V, Bondeau A, Plutzar C, Gingrich S, Lucht W, Fischer-Kowalski M (2007) Quantifying and mapping the human appropriation of net primary production in earth’s terrestrial ecosystems. Proc Natl Acad Sci 104(34):12942–12947
Haines-Young R (2009) Land use and biodiversity relationships. Land Use Policy 26:179–186
Harper KA, MacDonald SE, Burton PhJ, Chen J, Brosofsfe KD, Saunders SC, Euskirchen ES, Robert D, Jaiteh MS, Esseen PA (2005) Edge influence on forest structure and composition in fragmented landscapes. Conserv Biol 19:768–782
Hawkins BA, Field R, Cornell HV, Currie DJ, Guegan JF, Kaufman DM, Kerr JT, Mittelbach GG, Oberdorff T, O’Brien EM, Porter EE, Turner JRG (2003) Energy, water, and broad-scale geographic patterns of species richness. Ecology 84(12):3105–3117
Helming K, Perez-Soba M, Tabbush P (eds) (2007) Sustainability impact assessment of land use changes. Springer, New York
Ho MW, Ulanowicz R (2005) Sustainable systems as organisms? BioSystems 82(1):39–51
Hong SK, Bogaert J, Min Q (2014) Biocultural landscapes. Diversity, functions and values. Springer, New York
Hooper DU, Chapin FS III, Ewel JJ et al (2005) Effects of biodiversity of ecosystem function: a consensus of current knowledge. ESA report. Ecol Monogr 75(1):3–35
Horn HS (1975) Markovian properties of forest succession. In: Cody ML, Diamond JM (eds) Ecology and evolution of communities. Belknap Press, Cambridge
Huston M (2014) Disturbance, productivity, and species diversity: empiricism versus logic in ecological theory. Ecology 95:2382–2396
Hutchinson GE (1951) Copepodology for the ornithologist. Ecology 32:571–577
Jaeger J (2000) Landscape division, splitting index, and effective mesh size: new measures of landscape fragmentation. Landsc Ecol 15(2):115–130
Johst K, Gutt J, Wissel C, Grimm V (2006) Diversity and disturbances in the antarctic megabenthos: feasible versus theoretical disturbance ranges. Ecosystems 9:1145–1155
Jover G, Manera C (2009) Producción y productividad agrícola en la isla de Mallorca, 1590-1860. Revista de Historia Económica—Journal of Iberian and Latin American Economic History XXVII(3):463-498
Jover G, Soto R (2002) Colonización feudal y organización del territorio, Mallorca, 1230-1350. Revista de Historia Económica—Journal of Iberian and Latin American Economic History XX(3):437-475
Krausmann F, Erb KH, Gingrich S, Haberl H, Bondeau A, Gaube V, Lauk C, Plutzar C, Searchinger T (2013) Global human appropriation of net primary production doubled in the 20th century. Proc Natl Acad Sci 110:10324–10329
Kumaraswamy S, Kunte K (2013) Integrating biodiversity and conservation with modern agricultural landscapes. Biodivers Conserv 22(12):2735–2750
Li BL (2000) Why is the holistic approach becoming so important in landscape ecology? Landsc Urban Plan 50:27–41
Liu L, Dietz T, Carpenter SR, Alberti M, Folke C, Moran E, Pell AN, Deadman P, Kratz T, Lubchenco J, Ostrom E, Ouyang Z, Provencher W, Redman CL, Schneider SH, Taylor WW (2007) Complexity of coupled human and natural systems. Science 317(5844):1513–1516
Loreau M (2000) Are communities saturated? On the relationship between α, β and γ diversity. Ecol Lett 3:73–76
Manera C (2001) Història del creixement econòmic a Mallorca (1700–2000). Lleonard Muntaner Editor, Palma de Mallorca
Margalef R (2006) Ecological theory and prediction in the study of the interaction between man and the rest of biosphere. In: Siolo H (ed) Ökologie und Lebensschutz in Intrenationaler Sicht, Rombach Freiburg, 1973; reprinted in Catalan, Spanish and English in Medi Ambient. Tecnologia i Cultura 38:114–125
Marull J, Mallarach JM (2005) A GIS methodology for assessing ecological connectivity: application to the Barcelona Metropolitan area. Landsc Urban Plan 71:243–262
Marull J, Pino J, Mallarach JM, Cordobilla MJ (2007) A land suitability index for strategic environmental assessment in metropolitan areas. Lands Urban Plan 81:200–212
Marull J, Pino J, Tello E, Cordobilla MJ (2010) Social metabolism, landscape change and land-use planning in the Barcelona Metropolitan region. Land Use Policy 27:497–510
Marull J, Tello E, Wilcox PT, Coll F, Pons M, Warde P, Valldeperas N, Ollés A (2014) Recovering the land-use history behind a Mediterranean edge environment: the importance of cultural landscapes in biological conservation. Appl Geogr 54:1–17
Marull J, Otero I, Stefanescu C, Tello E, Coll F (2015a) Exploring the links between forest transition and landscape changes in the Mediterranean. Can forest recovery lead to lower landscape quality? Agroforest Syst 20:1–15 (in press)
Marull J, Font C, Tello E, Fullana N, Domene E (2015b) Towards an Energy-Landscape Integrated Analysis? Exploring the links between socio-metabolic disturbance and landscape ecology performance (Mallorca, Spain, 1956–2011). Landsc Ecol (accepted)
Marull J, Font C, Padró R, Tello E, Panazzolo A (2015c) Energy–Landscape Integrated Analysis of agro-ecosystems: how the complexity of energy flows shapes landscape patterns (Barcelona province, 1860–2000). Ecol Indic (submitted)
Matson PA, Vitousek PM (2006) Agricultural intensification: will land spared from farming be land spred for nature? Conserv Biol 20(3):709–710
Matthews R, Selman P (2006) Landscape as a focus for integrating human and environmental processes. J Agric Econ 57:199
McGarigal K, Marks B (1994) FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. General Technical Report PNW-GTR-351. Forest Service, Pacific Northwest Research Station, Portland
Miller A, Reilly D, Bauman S, Shea K (2012) Interactions between frequency and size of disturbance affect competitive outcomes. Ecol Res 27:783–791
Molino JF, Sabatier D (2001) Tree diversity in tropical rain forests: a validation of the intermediate disturbance hypothesis. Science 294:1702–1704
Morowitz HJ (2002) The Emergence of Everything: how the world became complex. Oxford University Press, Oxford
Murray I (2012) Geografies del capitalisme balear. Poder, metabolisme socio-econòmic i petjada ecològica d’una superpotència turística. Dissertation, University of the Balearic Islands
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GA, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Naredo JM (2004) La evolución de la agricultura en España (1940–2000). Universidad de Granada, Granada
O’Neill RV, Krummel JR, Gardner RH, Sugihara G, Jackson B, DeAngelis DL, Milne BT, Turner MG, Zygmunt B, Christensen SW, Dale VH, Graham RL (1988) Indices of landscape pattern. Landsc Ecol 1(3):153–162
Odum HT (1984) Energy analysis of the environmental role in agriculture. In: Stanhill E (ed) Energy and agriculture. Springer, Dordrecht, pp 24–51
Odum HT (2007) Environment, power, and society for the twenty-first century. The hierarchy of energy. Columbia University Press, New York
Opdam P, Steingrover E, van Rooij S (2006) Ecological networks: a spatial concept for multi-actor planning of sustainable landscapes. Landsc Urban Plan 75:322–332
Padisak J (1993) The influence of different disturbance frequencies on the species richness, diversity and equitability of phytoplankton in shallow lakes. Hydrobiologia 249:135–156
Parrotta JA, Trosper RL (2012) Traditional forest-related knowledge: sustaining communities, ecosystems and biocultural diversity. World Forests 12:1–621
Pelletier N, Audsley E, Brodt S, Garnett T, Henriksson P, Kendall A, Kramer K, Murphy D, Nemecek T, Troell M (2011) Energy intensity of agriculture and food systems. Annu Rev Environ Resour 36:223–246
Perfecto I, Vandermeer J (2010) The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proc Natl Acad Sci USA 107(13):5786–5791
Peterseil J, Wrbka T, Pultzar C, Schmitzberger I, Kiss A, Szerencsits E, Reiter K, Schneider W, Suppan F, Beissmann H (2004) Evaluating the ecological sustainability of Austrian agricultural landscapes—the SINUS approach. Land Use Policy 21:307–320
Petraitis PS, Latham RE, Niesenbaum RA (1989) The maintenance of species diversity by disturbance. Q Rev Biol 64:393–418
Phalan B, Onial M, Balmford A, Green RE (2011) Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science 333:1289–1291
Pierce S (2014) Implications for biodiversity conservation of the lack of consensus regarding the humped-back model of species richness and biomass production. Funct Ecol 28:253–257
Pino J, Marull J (2012) Ecological networks: are they enough for connectivity conservation? A case study in the Barcelona Metropolitan Region (NE Spain). Land Use Policy 29:684–690
Plieninger T, Hui C, Gaertner M, Huntsinger L (2014) The impact of land abandonment on species richness and abundance in the Mediterranean basin: a meta-analysis. PLoS One 9(5):e98355. doi:10.1371/journal.pone.0098355
Pons A, Rullan O, Murray I (2014) Tourism capitalism and island urbanization: tourist accommodation diffusion in the Balearics, 1936–2010. Isl Stud J 9(2):239–258
Rayó M (2004) L’ecologisme a les Balears. Edicions Documenta Balear, Palma de Mallorca
Reed RA, Johnson J, Baker WL (1996) Fragmentation of a forested rocky mountain landscape 1950–1993. Biol Conserv 75:267–277
Rindfuss RR, Walsh SJ, Turner BL, Fox J, Mishra V (2008) Developing a science of land change: challenges and methodological issues. Proc Natl Acad Sci 101(39):13976–13981
Robson JP, Berkes F (2011) Exploring some of the myths of land use change: can rural to urban migration drive declines in biodiversity? Global Environ Chang 21(3):844–854
Roca J (1992) Modernització agrícola i desenvolupament industrial. El cas de Mallorca (1850–1950). Estudis Baleàrics 43:109–118
Rosselló-Verger VM (1982) Canvis de propietat i parcel.lacions al camp mallorquí entre els segles XIX i XX. Randa. Història i Cultura de Mallorca 12:19–60
Roxburgh SH, Shea K, Wilson JB (2004) The intermediate disturbance hypothesis, patch dynamics and mechanisms of species coexistence. Ecology 85:359–371
Rullan O (2002) La construcció territorial de Mallorca. Editorial Moll, Palma de Mallorca
Rullan O (2010) Las políticas territoriales en las Islas Baleares. Cuadernos geográficos de la Universidad de Granada 47:403–428
Sasaki T, Okubo S, Okayasu T, Jamsram U, Ohkuro T, Takeuchi K (2009) Management applicability of the intermediate disturbance hypothesis across Mongolian rangeland ecosystems. Ecol Appl 19(2):423–432
Schroter D, Cramer W, Leemans R, Prentice IC, Araújo MB, Arnell NW, Bondeau A, Bugmann H, Carter TR, Gracia CA, de la Vega-Leinert AC, Erhard M, Ewert F, Glendining M, House JI, Kankaanpää S, Klein RJT, Lavorel S, Lindner M, Metzger MJ, Meyer J, Mitchell TD, Reginster I, Rounsevell M, Sabaté S, Sitch S, Smith B, Smith J, Smith P, Sykes MT, Thonicke K, Thuiller W, Tuck G, Zaehle S, Zier B (2005) Ecosystem service supply and vulnerability to global change in Europe. Science 310:1333–1337
Schwartz MW, Bringham CA, Hoeksema JD, Lyons KG, Mills MH, van Mantgem PJ (2000) Linking biodiversity to ecosystem function: implications for conservation ecology. Oecologia 122:297–305
Schwarzlmüller E (2009) Human appropriation of aboveground net primary production in Spain, 1955–2003: an empirical analysis of the industrialization of land use. Ecol Econ 69(2):282–291
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27(379–423):623–656
Shea K, Chesson P (2002) Community ecology theory as a framework for biological invasions. Trends Ecol Evol 17:170–176
Shea K, Roxburgh SH, Rauschert ESJ (2004) Moving from pattern to process: coexistence mechanisms under intermediate disturbance regimes. Ecol Lett 7:491–508
Sheil D, Burslem D (2003) Disturbing hypotheses in tropical forests. Trends Ecol Evol 18:18–26
Sheil D, Burslem D (2013) Defining and defending Connell’s intermediate disturbance hypothesis: a response to Fox. Trends Ecol Evol 28:571–572
Shreeve TG, Dennis RLH, Van Dick H (2004) Resources, habitats and metapopulations—whither reality? Oikos 106:404–408
Socies A (2013) Varietats locals de les Illes Balears. Documenta Balear, Palma de Mallorca
Soto R (2015) Feudal colonization and socio-ecological transition in the island of Mallorca in the thirteenth century. Continuity and Change (in print)
Suau J (1991) El món rural mallorquí, segles XVIII-XIX. Curial, Barcelona
Svensson JR, Lindegarth M, Jonsson PR, Pavia H (2012) Disturbance-diversity models: what do they really predict and how are they tested? Proc R Soc Lond B 279:2163–2170
Swift MJ, Izac AMN, van Noordwijk M (2004) Biodiversity and ecosystem services in agricultural landscapes—are we asking the right questions? Agric Ecosyst Environ 104(1):113–134
Tischendorf L (2001) Can landscape indices predict ecological processes consistently? Landsc Ecol 16:235–254
Tress B, Tress G, Décamps H, d’Hauteserre AM (2001) Bridging human and natural sciences in landscape research. Landsc Urban Plan 57:137–141
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I, Perfecto I, Vandermeer J, Whitbread A (2012) Global food security, biodiversity conservation and the future of agricultural intensification. Biol Conserv 151:53–59
Turner MG, Ruscher CL (1988) Changes in landscape pattern in Georgia, USA. Landsc Ecol 1:241–251
Turner BL, Lambin EF, Reenberg A (2007) The emergence of land change science for global environmental change and sustainability. Proc Natl Acad Sci USA 104(52):20666–20671
Ulanowicz RE (1997) Ecology, the ascendent perspective. Columbia University Press, New York
Urech y Cifre C (1869) Estudios sobre la riqueza territorial de las Islas Baleares. Felipe Guasp, Palma de Malloca
Vackar D, Chobot K, Orlitova E (2012) Spatial relationship between human population density, land use intensity and biodiversity in the Czech Republic. Landsc Ecol 27(9):1279–1290
Van der Maarel E (1993) Some remarks on disturbance and its relations to diversity and stability. J Veg Sci 4:733–736
Verburg PH, van de Steeg J, Veldkamp A, Willemen L (2009) From land cover change to land function dynamics: a major challenge to improve land characterization. J Environ Manag 90:1327–1335
Verdasca MJ, Leitao AS, Santana J, Porto M, Dias S, Beja P (2012) Forest fuel management as a conservation tool for early successional species under agricultural abandonment: the case of Mediterranean butterflies. Biol Conserv 146:14–23
Vitousek PM, Ehrlich PR, Ehrlich AH, Matson PA (1986) Human appropriation of the products of photosynthesis. Bioscience 36:363–373
Wilkinson DM (1999) The disturbing history of intermediate disturbance. Oikos 84(1):145–147
Wilson JB (1990) Mechanisms of species coexistence: twelve explanations for Hutchinson’s ‘paradox of the plankton’: evidence from New Zealand plant communities. N Z J Ecol 13:17–42
Wilson JB (1994) The ‘intermediate disturbance hypothesis’ of species coexistence is based in on patch dynamics. N Z J Ecol 18:176–181
Wrbka T, Erb K-H, Schulz NB, Peterseil J, Hahn C, Haberl H (2004) Linking pattern and process in cultural landscapes. An empirical study based on spatially explicit indicators. Land Use Policy 21:289–306
Young JC, Waylen KA, Sarkki S, Albon S, Bainbridge I, Balian E, Davidson D, Edwards D, Fairley R, Margerison C, McCracken D, Owen R, Quine CP, Stewart-Roper C, Thompson D, Tinch R, Van den Hove S, Watt A (2014) Improving the science-policy dialogue to meet the challenges of biodiversity conservation: having conversations rather than talking at one-another. Biodivers Conserv 23:387–404
Acknowledgments
This work has been supported by the research project HAR2012-38920-C02-02 funded by the Spanish Ministry of Economy and Competitiveness, and the international Partnership Grant SSHRC-895-2011-1020 ‘Sustainable farm systems: long-term socio-ecological metabolism in western agriculture’ funded by the Social Sciences and Humanities Research Council of Canada. We also thank the comments received in the oral presentation made in the European Conference on ‘Linking Biological and Cultural Diversity in Europe’ held in Florence (08-11/04/2014) for the Implementation of the UNESCO-SCBD Joint Programme on Biological and Cultural Diversity.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Mauro Agnoletti.
Rights and permissions
About this article

Cite this article
Marull, J., Tello, E., Fullana, N. et al. Long-term bio-cultural heritage: exploring the intermediate disturbance hypothesis in agro-ecological landscapes (Mallorca, c. 1850–2012). Biodivers Conserv 24, 3217–3251 (2015). https://doi.org/10.1007/s10531-015-0955-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-015-0955-z