
Abstract
One of the tree species with the greatest invasive potential worldwide is Pinus contorta Douglas ex Loundon, which is characterized by producing a great number of individuals in short periods of time, generating different impacts on the ecosystems it invades. The genus Chusquea is present in a diverse number of ecosystems in South America. In the temperate forests of Chile and Argentina, it forms dense thickets in the undergrowth as well as in open areas. Its rapid growth and vegetative reproduction, with large numbers of individuals, allow the species to dominate and restrict the establishment of other plant species. This study focused on analyzing the effect of the presence of individuals of Chusquea culeou E. Desv. on the establishment of the invasive species Pinus contorta, as well as on other native herbaceous and shrub species in Araucaria-Nothofagus forest. With this aim, we established 204 plots within the Malalcahuello National Reserve (in the Andes of south-central Chile) along a gradient of P. contorta invasion, where the presence or absence of these species was recorded through different percentages of Chusquea cover. The results showed that the probability of presence of P. contorta decreases significantly in areas where Chusquea cover is greater than 25%. Native species also decrease their presence, but to a lesser extent compared to P. contorta. Our results demonstrate the inhibitory effect of Chusquea for the establishment of other plant species, but more markedly for invasive conifers.
Resumen
Una de las especies arbóreas con mayor potencial de invasión a nivel mundial es Pinus contorta Douglas ex Loundon, la cual se caracteriza por producir una gran cantidad de individuos en cortos periodos de tiempo, lo que genera distintos impactos sobre los ecosistemas que invade. El género Chusquea, con varias especies de bambú, está presente en diversos ecosistemas de Sudamérica. En los bosques templados de Chile y Argentina forma densos matorrales en el sotobosque y zonas abiertas. Su rápido crecimiento y alta reproducción vegetativa, le permiten dominar y restringir el establecimiento de nuevas especies vegetales una vez establecido. El presente estudio se enfocó en analizar el efecto de la presencia de la especie Chusquea culeou E. Desv. sobre la abundancia y la probabilidad de establecimiento de la conífera invasora P. contorta y también de especies nativas de carácter herbáceas y arbustivas en Bosques de Araucaria -Nothofagus. Para ello se establecieron 204 parcelas en la Reserva Nacional Malalcahuello a lo largo de un gradiente de invasión de P. contorta, donde se registró la presencia/ausencia de estas especies a través de distintos porcentajes de cobertura de Ch. culeou. Los resultados mostraron que la abundancia y la probabilidad de establecimiento de P. contorta disminuye significativamente en lugares donde la cobertura de Chusquea es mayor al 25%. Las especies nativas también disminuyen su presencia, pero en menor magnitud en comparación al invasor P. contorta. Estos resultados demuestran el efecto inhibidor de Ch. culeou para el establecimiento de individuos de otras especies vegetales, pero más marcadamente para la conífera invasora.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The introduction of non-native tree species into different ecosystems is mediated by anthropogenic vectors, where two main sources can be identified (Brundu et al. 2020). The first corresponds to the introduction of non-native trees for ornamental purposes, which are concentrated in urban centers, where some of the species can escape from the original cultivation, mainly by human vectors such as vehicles and urban rivers (e.g., Acer platanoides, Acacia dealbata) (Säumel and Kowarik 2010). Secondly, the forest industry has historically functioned as a source of introduction of non-native tree species, being responsible for more than 65% of these tree species introductions. The main introduced species worldwide are of the genera Pinus, Eucalyptus, Acacia, Leucaena (Simberloff et al. 2010; Richardson et al. 2014). The success associated with the invasion of non-native trees is related to the fact that these species generally exhibit traits such as high propagule production, easy and early establishment, rapid growth, wide range of climatic tolerance and the ability to colonize disturbed ecosystems (Colautti and MacIsaac 2004; Peña et al. 2008; van Kleunen et al. 2010a, b; Van Kleunen et al. 2010a, b).
Some species of the genus Pinus are among the most invasive species worldwide (Lowe et al. 2000). In the southern hemisphere they are one of the most common genus of invasive tree species (Richardson 2006; Taylor et al. 2016). Within the genus Pinus, one of the most invasive species is Pinus contorta Douglas ex Loudon (Taylor et al. 2016), which invades Andean and Patagonian ecosystems in central and southern Chile (Langdon et al. 2010). The invasiveness of this species is mainly due to early seed production (5 to 10 years), small seeds that are easily transported by wind over long distances, and rapid seedling growth (Ledgard 2001; Peña et al. 2008; Langdon et al. 2010; Pauchard et al. 2016).
The invasion success of P. contorta also depends on the invasibility of ecosystems, where sites with high disturbance (e.g., fire-affected sites), exposed mineral soil, low temperatures and limited vegetation cover are favorable environments for its establishment and further invasion (Peña et al. 2008; Pauchard et al. 2016). Biotic resistance has historically been one of the factors that regulate the success of plant invasions, this regulation is primarily mediated by competition from resident plants (Levine et al. 2004; Beaury et al. 2020). However, other types of interactions can also regulate the success of plant invasions, such as predation, herbivory, and diseases caused by parasitic fungi, which can reduce both the establishment and recruitment of non-native plants (Nunez-Mir et al. 2017). In the case of Pinus species, it has been determined that the structure, composition, richness, and abundance of the resident community are key factors that reduce invasion (Richardson and Bond 1991). In this context, it has been observed that grassland communities, open soils, and abandoned fields are more susceptible to invasion by Pinus species (Richardson and Bond 1991). On the other hand, mixed shrublands, shrubs, and forests tend to generate higher biotic resistance against Pinus species (Richardson et al. 1994). For example, studies conducted in central Chile have indicated that the probability of establishment of Pinus radiata individuals decreases significantly with increasing native forest coverage (Gómez et al. 2019). Meanwhile, in the Argentinean pampas, it has been determined that the presence of native rodents through granivory has controlled the invasion of the invasive trees Robinia pseudoacacia and Gleditsia triacanthos (Muschetto et al. 2015).
The arrival of P. contorta in Chile and its establishment is due to its use for soil protection, erosion control and dune stabilization (Peña et al. 2008). During the 1970s, the Chilean Forestry Institute (INFOR) developed a program for the introduction of non-native species, aimed at the productive sector (Peña et al. 2008). Among these experimental plots, some that were located in the Malalcahuello National Reserve (MNR), in La Araucanía region stand out, from which an invasion process has been generated within the same reserve, mainly affecting the Andean ecosystem dominated by the long-lived, native conifer Araucaria araucana and species of the genus Nothofagus (hereafter termed as Araucaria-Nothofagus forest). The understory of these forests are dominated by Chusquea culeou and Chusquea quila (Peña et al. 2008; Langdon et al. 2010; Urrutia et al. 2013; Cóbar-Carranza et al. 2014). The main impacts of the invader Pinus contorta documented in the MNR are the reduction of native plant richness, which varies depending on the density of the invasion (Urrutia et al. 2013; Franzese et al. 2017), changes in soil biota, changes in fire regimes (Cóbar-Carranza et al. 2014; Taylor et al. 2016; Policelli et al. 2019), changes in the functional traits of local communities (Bravo-Monasterio et al. 2016) and changes in the microclimatic conditions of invaded ecosystems (García et al. 2023).
In the Nothofagus-Araucaria forests of the MRN, one of the main species that make up the understory are the species of the genus Chusquea spp. The species of the genus Chusquea (Fam: Poaceae, Subfam: Bambusoides) have a great effect on the distribution, establishment and growth of seedlings, mainly of the seed bank of native species (Holz and Veblen 2006). This species has a heterogeneous distribution within MRN forests, forming clusters of approximately 6 m in diameter that dominate the understory. Its occurrence is more frequent in forests that have been logged or previously affected by fires, whether natural or human induced (Marchesini Victoria et al. 2009).
The presence of populations of Chusquea spp. generates important changes in the microclimatic conditions of the ecosystem they inhabit (Caccia et al. 2015). For example, it has been found that dense stands of Chusquea spp. can reduce average light radiation levels, reduce soil and air temperature (~ 2 °C less than in sites without Chusquea spp.), and increase litter depth and soil moisture (Holz and Veblen 2006; Caccia et al. 2009). These conditions of low temperature, high humidity and low light availability generate direct impacts on vegetation, such as the generation of sub-optimal conditions for the establishment of native and non-native species (Caccia et al. 2015). In addition, there are other indirect impacts, such as the increase of pathogenic fungi and herbivory that increase the mortality rate of native and non-native species (Holz and Veblen 2006; Caccia et al. 2015).
It is also important to highlight the phenological processes of Chusquea spp., which are an important factor influencing the dynamics of regeneration and recruitment of native species in the forests of southern Chile (Sanguinetti and Kitzberger 2009). This is because species within the genus exhibit gregarious flowering, massive seed production, and rapid senescence, occurring at intervals ranging from 3 to 120 years (Marchesini Victoria et al. 2009). These phenological patterns result in significant changes within forests, such as alterations in microclimatic conditions (Holz and Veblen 2006) and reductions in granivory and herbivory (Sanguinetti and Kitzberger 2009). Additionally, this creates space and releases soil resources in the understory that can be utilized by new species (Caccia et al. 2015).
In this context, the presence of Chusquea sp. functions as a species that provides biotic resistance, regulating the establishment of both native plants, such as species of the Nothofagus genus, and non-native plants (Torres et al. 2018). The mechanisms associated with the biotic resistance exerted by this species encompass a wide range of interactions. On one hand, it increases competition for light, and its high seed production (50 million per hectare) enhances granivory and herbivory, reducing the recruitment capacity of native species. Additionally, it modifies microclimatic conditions, increasing humidity and decreasing temperature, thereby favoring the attack of pathogenic fungi that directly affect the recruitment of both native and non-native plants (Holz and Veblen 2006; Marchesini Victoria et al. 2009).Thus, the population dynamics of Chusquea spp. may be playing a fundamental role in the invasion dynamics of P. contorta in the Malalcahuello National Reserve, since on the one hand it may be functioning as a biotic filter that prevents the establishment of P. contorta individuals, but on the other hand, when a process of mass mortality occurs, P. contorta individuals may be benefited by finding the space, biotic and abiotic conditions necessary to establish themselves and expand their invasive range within this ecosystem. These patterns have already been observed in other ecosystems invaded by P. contorta. For example, in New Zealand the presence of native shrubs such as Fucosporta cliffortioides prevent the establishment of P. contorta individuals, functioning as a biotic filter to control the invasion of this species (Taylor et al. 2016).
In this context, it is important to evaluate how the presence of Ch. culeou could play a role on the invasion process of Pinus contorta, mainly by reducing its establishment and growth. For this we asked whether the increase in the cover of Ch. culeou decreases the abundance and the probability of establishment of individuals of P. contorta, as well as for other herbaceous and shrub species. We hypothesized that the increase in the cover of Ch. culeou within the Araucaria-Nothofagus forests of the Malalcahuello National Reserve will significantly decrease the abundance and probability of establishment of plant species, with this effect being more intense on the non-native invasive species P. contorta.
Methods
Study area
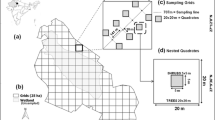
The study area corresponds to the Malalcahuello National Reserve located in La Araucanía Region, Chile (38 °S 25′; 71 °W 33′; 1420 m.a.s.l.) (Fig. 1). The area has a mean annual temperature ~ 9 °C and a mean annual precipitation of 3000 mm (Franzese et al. 2017). Topographically, it is an ecosystem that has been shaped by volcanic and glaciological activity that gives it unique characteristics in terms of the biodiversity (Gajardo 1994). This ecosystem is naturally dominated by open forests of Araucaria araucana ((Mol.) C. Koch.; Araucariaceae) and Festuca scabriuscula (Phil.; Poaceae) (Urrutia et al. 2013). There are also small and dense patches dominated by Nothofagus antarctica (G. Forst. Oerst.; Nothofagaceae) and Chusquea culeou (E. Desv.; Poaceae) (Franzese et al. 2017). The forest structure has undergone modification due to the invasion of P. contorta, leading to a significant increase in tree canopy cover attributable to the presence of this species (García et al. 2023). Ch. culeou is present in patches of varying sizes distributed heterogeneously throughout the landscape. The establishment of Ch. culeou predates the invasion and subsequent crown closure in areas with larger pine trees. Therefore, it is currently possible to find patches of this shrub in both invaded and non-invaded areas by P. contorta (Fig. S1).

a Location of the study site in the Malalcahuello National Reserve. b Distribution of sampling points along a fraction of the pine invasion gradient. At each point (yellow) between 4 and 6 plots of 1 m2 were established. c Un-invaded Araucaria-Nothofagus forest with presence of Ch. culeou patches. d Core area of P. contorta invasion
Study specie
Pinus contorta is a native species of North America that can grow up to 35 m in height and 60 cm in diameter at breast height (DBH). It is considered a pioneer species, intolerant to shade, and exhibits rapid growth. It has a broad ecological range and tolerance, growing from sea level up to 3600 m a.s.l. It can withstand temperatures ranging from − 57 to 38 °C and annual precipitation varying between 250 and 500 mm (Cóbar-Carranza et al. 2014). It was introduced in the 1970s (initial population < 0.5 ha) along with other species of the genus Pinus (e.g., P. sylvestris, P. ponderosa) in the Malalcahuello National Reserve. Currently, it is estimated that the invasion area of P. contorta exceeds 100 ha, reaching densities of < 6600 plants ha−1, specially in the sectors with greater invasion (Peña et al. 2008; Cóbar-Carranza et al. 2014; Franzese et al. 2017).
Sampling
To evaluate the abundance and probability of presence of species with herbaceous and shrub habit, as well as seedlings of P. contorta (height < 1 m), 204 plots of 1 m2 were randomly established in the Malalcahuello N.R., within an invasion gradient of P. contorta ranging from 12.5 ton/ha−1 to 150 on/ha−1. Within each plot, the abundance of all vascular plant species was quantified by analyzing the percentage of cover occupied by each species in the 1 m2 plot, which was subdivided into 25 quadrants. Chusquea quila cover was estimated by the projection of its “crown” over the 1 m2 plot. Tree canopy cover was quantified by means of a hemispherical photograph under low light conditions, which was placed in the center of the 1 m2 plot at 1 m above the ground. In each plot, the presence (1) and absence (0) of herbaceous and shrub (excluding Ch. culeou) species and individuals of P. contorta were recorded to evaluate the probability of establishment as a function of Ch. culeou cover and tree canopy cover. It is important to consider that a large percentage of the tree canopy cover in the study sites consists of P. contorta individuals, with a smaller percentage comprising tree species such as A. araucana and Nothofagus spp.
Data analysis
To determine the effect of the presence of Ch. culeou and its tree canopy cover on the abundance of the different growth forms, a generalized linear model (GLM) was performed, fitting a poisson distribution with a log link function. To determine the effect of the percentage cover of Ch. culeou and tree canopy cover on the probability of establishment of species of herbaceous and shrub growth form and the presence of P. contorta individuals, a logistic regression was modeled and fitted with a binomial distribution and a logit link function. The glm function was used in both models above-mentioned. To analyze the percentage of variance of the binomial model, the Pseudo R2 value was used. Finally, to determine the best model (Ch. culeou cover or tree canopy cover) to explain changes in abundance and establishment probability, the Akaike Information Criterion (AIC) was used, where the best model was chosen with the lowest AIC value (Akaike 1973). All data analyses were performed with R version 4.0.5 (Team 2015), and all the plots were performed using the ggplot2 R package (Wickham 2008).
Results
Species composition and abundance at the study site
A total of 41 plant species were recorded, of which 59% correspond to herbaceous species, 34% to shrubs and only 7% were trees. In relation to the biogeographic origin of the species, 93% of the species are native and only 7% correspond to non-native species (Table S1). The most frequent species were the non-native tree P. contorta that was present in 79.41% of the plots sampled. Followed the native conifer A. araucana (4.41%), and the shrubs Chusquea culeou (75.98%) and Gaultheria pumila (45.09%). Finally, the herbaceous layer was composed mainly by species Festuca acanthophylla (38.24%) and the exotic species Rumex acetosella (20.09%).
Chusquea culeou cover had a significant effect on the decrease of cumulative abundance of P. contorta individuals (p < 0.001) (Table 1; Fig. 2). This same effect was observed for species with herbaceous (p < 0.001) and shrub (p < 0.001) growth forms (Fig. 3a). In the case of tree canopy cover, the cumulative abundance of herbaceous (p = 0.003) and shrubs (p = 0.002) decreased, while a slight increase in the cumulative abundance of P. contorta (p < 0.001) was observed (Table 1, Fig. 3b). For both P. contorta species and the other growth forms, the interaction between Ch. culeou cover and tree canopy cover was significant (p < 0.001) (Table 1).

a Effect of Chusquea culeou and b tree canopy cover on the cumulative abundance of herbs (black), shrub (orange) and P. contorta (blue) individuals

Probability of establishment for a herbs, b shrubs and c P. contorta individuals, under the cover of Chusquea culeou. The curves indicate the values predicted by the binomial model for each of the individuals analyzed
Effect of Ch. Culeou cover and tree canopy cover on the probability of establishment of growth form
The probability of establishment of P. contorta individuals is significantly reduced as a function of increasing Ch. Culeou cover (Table 2; p < 0.001) (Fig. 4c), and increases as a function of tree canopy cover (p < 0.001) (Table 2; Fig. 4c). Regarding the percentage of variance explained, Ch. Culeou cover explains 19.76% of the data, while tree canopy cover explains 4.41% of the data. The Akaike information criterion (AIC) determined that the best model to explain the probability of establishment of P. contorta is the cover of Ch. Culeou (AIC = 240.63).

Probability of establishment for a herb, b shrub and c P. contorta individuals, under tree canopy cover. The curves indicate the values predicted by the binomial model for each of the individuals analyzed
The probability of establishment of other herbaceous and shrub growth forms also decreases significantly as a function of Ch. Culeou cover (Table 2; Fig. 4a and b) and tree canopy cover (Table 2; Fig. 4a and b). As for the percentage of variance explained, the variable Ch. Culeou cover explains 26.04% of the data for the probability of herbaceous establishment, while tree canopy cover explains only 17.39%. In the case of shrub growth form, the variable Ch. culeou cover explains only 11.88% of the data, while tree canopy cover explains 22.66% of the data. The AIC determined that the best model to explain the probability of herbaceous species establishment is the one containing the cover of Ch. culeou (AIC = 265.55), while for the probability of shrub species establishment the best model was for tree canopy cover (AIC = 309.08).
Discussion
Based on these results, we can assign an important role to the native bamboo Ch. culeou in reducing the abundance and probability of establishment of the invasive species P. contorta. This also occur to a lesser extent with other herbaceous and shrub species. In terms of tree cover, it tends to increase the abundance and establishment probability of P. contorta. On the other hand, tree canopy cover reduces the probability of occurrence of herbaceous and shrub species, with a maximum establishment probability of up to 45% under significant tree canopy cover.
The impact of Ch. culeou cover on the abundance and probability of establishment of herbaceous and shrubby growth form species, as well as P. contorta individuals, is mainly due to two key drivers. The first is the rapid growth of Chusquea sp species, which tends to quickly dominate the understory of Nothofagus-Araucaria forests (Veblen 1982). The second driver is the phenology of this species, which is characterized by a long period of time in vegetative state (3–20 years) and then a synchronized period of flowering and death, which varies within populations, but with an average periodicity of 15 years (Guerreiro 2014). The extent of this impact should be high, given that Ch. culeou dominates the understory in these temperate forests in the Andes (Holz and Veblen 2006).
This dominance of the understory in tropical and temperate forests of South America makes them an important component of the dynamics and structure of plant and animal communities (Budke et al. 2010). For example, the abundance of Ch. culeou may play an important role in the recruitment of different native species in the temperate forests of southern Chile (Holz and Veblen 2006; Guerreiro 2014). As for the wildlife, the understory of Chusquea sp. are the major source of food and shelter for most herbivores (Areta et al. 2009).
These characteristics cause species of the genus Chusquea to generate impacts on microclimatic conditions (Caccia et al. 2015), species diversity (Muñoz et al. 2012) and on the biotic interactions occurring under the understory (Caccia et al. 2009). In this context, it has been observed that the presence of Chusquea sp in forests of N. obliqua in the Chilean Patagonia, generates a decrease in light availability, higher soil moisture, and lower temperatures, which results in an increase in the mortality rate of juvenile individuals of N. obliqua (Caccia et al. 2009). Regarding the change in biotic interactions, it has been observed that the increase in the abundance of Chusquea sp. species generates an increase in the attack by pathogenic fungi (Abe et al. 2001), as well as an increase in herbivory and seed granivory of tree species (Sanguinetti and Kitzberger 2009).
This dominance of Ch. culeou in the understory and the impacts that its presence generates on the Nothofagus-Araucaria araucana forest may be a key factor that explains the dynamics of the invasion of P. contorta. The high dominance in the understory functions as a biotic filter that may be inhibiting germination and establishment of pines. In this context, our results are consistent with previous studies where it has been determined that the establishment success of P. contorta individuals within invaded ecosystems in southern Chile and New Zealand decreases as a function of the presence of native shrub and tree species (Taylor et al. 2016). This response is related to the fact that P. contorta is considered a pioneer species that in early stages of development needs a large amount of light for its establishment (Ledgard 2001; Pauchard et al. 2016). The results obtained regarding the higher abundance of P. contorta individuals in areas with greater tree canopy cover are mainly due to the fact that these areas also experience a higher invasion of P. contorta, resulting in increased recruitment of P. contorta seedlings in these zones (García et al. 2023). This invasion pattern has been observed in both forested and treeless ecosystems, where the propagule pressure is higher in the vicinity of mature trees (Pauchard et al. 2016; Langdon et al. 2019).
Sporadic flowering and gregarious death events could be a key factor explaining the mechanism by which P. contorta individuals can invade. In addition to this effect of flowering and gregarious death, Ch. culeou populations play a fundamental role in the fire regime, as it is a species prone to burn forming fire ladders that facilitate crowning. These disturbances may function as a driver to facilitate the dispersal and establishment of P. contorta individuals across the landscape (Langdon et al. 2019). It has also been shown that in areas invaded by P. contorta, fire can increase the quality of the seedbed and the rate of pine establishment, mainly if they are ecosystems with older invasion (Taylor et al. 2017).
Finally, it is important to consider these results from an invasion management point of view. Our results indicated that the presence of Ch. culeou significantly reduces the probability of establishment of P. contorta individuals and to a lesser extent herbaceous and tree species. Therefore, when carrying out management strategies in invaded areas, maintaining the cover of Ch. culeou could prevent recruitment of P. contorta individuals, allowing other native species (herbaceous or arboreal) to colonize these ecosystems. In addition, maintaining Ch. culeou could inhibit the effect of post-control or eradication invasion legacies, which will allow for better recruitment of native species in cases of restoring invaded ecosystems.
Conclusions
Our study determined that the cover of Ch. culeou significantly influences the abundance and probability of establishment of species with herbaceous and arboreal growth form, and especially on the non-native species P. contorta, even being more significant than the arboreal cover. These results are important for planning conservation strategies in the Malalcahuello National Reserve since the presence of a dense understory composed of species of the genus Chusquea could function as a biotic and even abiotic filter (changes in microclimatic conditions) that prevents the establishment and further invasion of P. contorta.
A deeper understanding of the phenological processes associated with historical and future Chusquea sp. species could be key to disentangle the invasion dynamics of P. contorta, as these stochastic processes of flowering and gregarious death, combined with natural and intentional disturbances, could favor the advance of the invasion in the Malalcahuello National Reserve.
It is important to be careful with these results for both management and control of P. contorta, because although the cover of Ch. culeou tends to reduce the probability of establishment of P. contorta, its spatial distribution within the Malalcahuello National Reserve tends to be heterogeneous within the landscape (clustered distribution in the landscape) unlike the tree canopy cover of P. contorta that tends to homogenize the ecosystem and be more constant in the landscape. Therefore, although the cover of Ch. culeou may increase biotic resistance, P. contorta individuals could establish in areas close to the propagule source that are free of Ch. culeou populations. The establishment of these pines could eventually suppress the Ch. culeou in the long term.
Data availability
Not applicable.
Code availability
Not applicable.
References
Abe M, Miguchi H, Nakashizuka T (2001) An interactive effect of simultaneous death of dwarf bamboo, canopy gap, and predatory rodents on beech regeneration. Oecologia 127:281–286. https://doi.org/10.1007/s004420000585
Akaike H (1973) Maximum likelihood identification of Gaussian autoregressive moving average models. Biometrika 60(2):255–265. https://doi.org/10.1093/biomet/60.2.255
Areta JI, Bodrati A, Cockle K (2009) Specialization on Guadua bamboo seeds by three bird species in the Atlantic forest of Argentina. Biotropica 41:66–73. https://doi.org/10.1111/j.1744-7429.2008.00458.x
Beaury EM, Finn JT, Corbin JD et al (2020) Biotic resistance to invasion is ubiquitous across ecosystems of the United States. Ecol Lett 23:476–482. https://doi.org/10.1111/ele.13446
Bravo-Monasterio P, Pauchard A, Fajardo A (2016) Pinus contorta invasion into treeless steppe reduces species richness and alters species traits of the local community. Biol Invasions 18:1883–1894. https://doi.org/10.1007/s10530-016-1131-4
Brundu G, Pauchard A, Pyšek P et al (2020) Global guidelines for the sustainable use of non-native trees to prevent tree invasions and mitigate their negative impacts. NeoBiota 61:65–116. https://doi.org/10.3897/neobiota.65.58380
Budke JC, Alberti MS, Zanardi C et al (2010) Bamboo dieback and tree regeneration responses in a subtropical forest of South America. For Ecol Manage 260:1345–1349. https://doi.org/10.1016/j.foreco.2010.07.028
Caccia FD, Chaneton EJ, Kitzberger T (2009) Direct and indirect effects of understorey bamboo shape tree regeneration niches in a mixed temperate forest. Oecologia 161:771–780. https://doi.org/10.1007/s00442-009-1412-z
Caccia FD, Kitzberger T, Chaneton EJ (2015) Episodic bamboo die-off, neighbourhood interactions and tree seedling performance in a Patagonian mixed forest. J Ecol 103:231–242. https://doi.org/10.1111/1365-2745.12349
Cóbar-Carranza AJ, García RA, Pauchard A, Peña E (2014) Effect of Pinus contorta invasion on forest fuel properties and its potential implications on the fire regime of Araucaria araucana and Nothofagus antarctica forests. Biol Invasions 16:2273–2291. https://doi.org/10.1007/s10530-014-0663-8
Colautti RI, MacIsaac HI (2004) A neutral terminology to define “invasive” species. Divers Distrib 10:135–141. https://doi.org/10.1111/j.1366-9516.2004.00061.x
Franzese J, Urrutia J, García RA et al (2017) Pine invasion impacts on plant diversity in Patagonia: invader size and invaded habitat matter. Biol Invasions 19:1015–1027. https://doi.org/10.1007/s10530-016-1344-6
Gajardo R (1994) La vegetaciόn natural de chile. clasificaciόn y Distribuciόn Geográfica. Editorial Universitaria, Chile
García RA, Fuentes-Lillo E, Cavieres L et al (2023) Pinus contorta alters microenvironmental conditions and reduces plant diversity in patagonian ecosystems. Diversity 15:320. https://doi.org/10.3390/d15030320
Gómez P, Murúa M, Martín JS et al (2019) Maintaining close canopy cover prevents the invasion of Pinus radiata: Basic ecology to manage native forest invasibility. PLoS ONE 14:1–13. https://doi.org/10.1371/journal.pone.0210849
Guerreiro C (2014) Flowering cycles of woody bamboos native to southern South America. J Plant Res 127(2):307–313. https://doi.org/10.1007/s10265-013-0593-z
Holz CA, Veblen TT (2006) Tree regeneration responses to Chusquea montana bamboo die-off in a subalpine Nothofagus forest in the southern Andes. J Veg Sci 17:19. https://doi.org/10.1658/1100-9233(2006)017[0019:trrtcm]2.0.co;2
Langdon B, Pauchard A, Aguayo M (2010) Pinus contorta invasion in the Chilean Patagonia: Local patterns in a global context. Biol Invasions 12:3961–3971. https://doi.org/10.1007/s10530-010-9817-5
Langdon B, Cavieres LA, Pauchard A (2019) At a microsite scale, native vegetation determines spatial patterns and survival of Pinus contorta invasion in Patagonia. Forests 10:1–15. https://doi.org/10.3390/f10080654
Ledgard N (2001) The spread of lodgepole pine (Pinus contorta, Dougl.) in New Zealand. For Ecol Manage 141:43–57. https://doi.org/10.1016/S0378-1127(00)00488-6
Levine JM, Adler PB, Yelenik SG (2004) A meta-analysis of biotic resistance to exotic plant invasions. Ecol Lett 7:975–989. https://doi.org/10.1111/j.1461-0248.2004.00657.x
Lowe S, Browne M, Boudjelas S, De Poorter M (2000) 100 of the world’s worst invasive alien species: a selection from the global invasive species database. Auckland: Invasive Species Specialist Group (ISSG), 12pp
Marchesini Victoria A, Sala Osvaldo E, Austin Amy T (2009) Ecological consequences of a massive flowering event of bamboo (Chusquea culeou) in a temperate forest of Patagonia, Argentina. J Veg Sci 20:424–432. https://doi.org/10.1111/j.1654-1103.2009.05768.x
Muñoz AA, González ME, Celedón C, Veblen TT (2012) Respuesta inicial de la regeneración arbórea luego de la floración y muerte de chusquea culeou (poaceae) en bosques andinos del centro-sur de Chile. Bosque 33:153–162. https://doi.org/10.4067/S0717-92002012000200005
Muschetto E, Mazia N, Cueto GR, Busch M (2015) Are rodents a source of biotic resistance to tree invasion in Pampean grasslands? Tree seed consumption under different conditions. Austral Ecol 40:255–266. https://doi.org/10.1111/aec.12208
Nunez-Mir GC, Liebhold AM, Guo Q et al (2017) Biotic resistance to exotic invasions: its role in forest ecosystems, confounding artifacts, and future directions. Biol Invasions 19:3287–3299. https://doi.org/10.1007/s10530-017-1413-5
Pauchard A, Escudero A, García RA et al (2016) Pine invasions in treeless environments: dispersal overruns microsite heterogeneity. Ecol Evol 6:447–459. https://doi.org/10.1002/ece3.1877
Peña E, Hidalgo M, Langdon B, Pauchard A (2008) Patterns of spread of Pinus contorta Dougl. ex Loud. invasion in a Natural Reserve in southern South America. For Ecol Manage 256:1049–1054. https://doi.org/10.1016/j.foreco.2008.06.020
Policelli N, Bruns TD, Vilgalys R, Nuñez MA (2019) Suilloid fungi as global drivers of pine invasions. New Phytol 222:714–725. https://doi.org/10.1111/nph.15660
Richardson DM (2006) Pinus: a model group for unlocking the secrets of alien plant invasions? Preslia 78:375–388
Richardson DM, Bond WJ (1991) Determinants of plant distribution: evidence from pine invasions. Am Nat 137:639–668. https://doi.org/10.1086/285186
Richardson DM, Williams PA, Hobbs RJ (1994) Pine invasions in the Southern hemisphere: determinants of spread and invadability. J Biogeogr 21:511. https://doi.org/10.2307/2845655
Richardson DM, Hui C, Nuñez MA, Pauchard A (2014) Tree invasions: patterns, processes, challenges and opportunities. Biol Invasions 16:473–481. https://doi.org/10.1007/s10530-013-0606-9
Sanguinetti J, Kitzberger T (2009) Efectos de la producción de semillas y la heterogeneidad vegetal sobre la supervivencia de semillas y el patrón espacio-temporal de establecimiento de plántulas en Araucaria araucana. Rev Chil Hist Nat 82:319–335. https://doi.org/10.4067/s0716-078x2009000300001
Säumel I, Kowarik I (2010) Urban rivers as dispersal corridors for primarily wind-dispersed invasive tree species. Landsc Urban Plan 94:244–249. https://doi.org/10.1016/j.landurbplan.2009.10.009
Simberloff D, Nuñez MA, Ledgard NJ et al (2010) Spread and impact of introduced conifers in South America: Lessons from other southern hemisphere regions. Austral Ecol 35:489–504. https://doi.org/10.1111/j.1442-9993.2009.02058.x
Taylor KT, Maxwell BD, Pauchard A et al (2016) Native versus non-native invasions: similarities and differences in the biodiversity impacts of Pinus contorta in introduced and native ranges. Divers Distrib 22:578–588. https://doi.org/10.1111/ddi.12419
Taylor KT, Maxwell BD, McWethy DB et al (2017) Pinus contorta invasions increase wildfire fuel loads and may create a positive feedback with fire. Ecology 98:678–687. https://doi.org/10.1002/ecy.1673
Team RC (2015) R Foundation for statistical computing. R Found Stat Comput Vienna, Austria, 2 , 2019
Torres A, Alarcón PAE, Rodríguez-Cabal MA, Nuñez MA (2018) Secondary invasions hinder the recovery of native communities after the removal of nonnative pines along a precipitation gradient in Patagonia. Forests 8:1–18. https://doi.org/10.3390/f9070394
Urrutia J, Pauchard A, García RA (2013) Diferencias en la composición vegetal de un bosque de Araucaria araucana (Molina) K.Koch y Nothofagus antarctica (G. Forst.) Oerst. asociadas a un gradiente de invasión de Pinus contorta Douglas ex Loudon. Gayana - Bot 70:92–100. https://doi.org/10.4067/S0717-66432013000100010
van Kleunen M, Dawson W, Schlaepfer DR et al (2010a) Are invaders different? A conceptual framework of comparative approaches for assessing determinants of invasiveness. Mark van Kleunen. 2010; Ecology Letters - Wiley InterScience. Ecol Lett 13:947–958
Van Kleunen M, Weber E, Fischer M (2010b) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol Lett 13:235–245. https://doi.org/10.1111/j.1461-0248.2009.01418.x
Veblen TT (1982) Growth patterns of chusquea bamboos in the understory of chilean nothofagus forests and their influences in forest dynamics. Bull Torrey Botanical Club 109(4):474–487
Wickham H (2008) ggplot2: Elganta graphics for data analysis
Acknowledgements
This work is part of the research carried out by the Laboratorio de Invasiones Biológicas (LIB), and much of the field work could not have been done without the support of the LIB team. Study funded by FONDECYT 1140485 and 1180205, CONAF FIBN N°014/2018. RGA, EFL, AP are funded by ANID/Basal FB210006. AFR thanks ANID/Basal FB210015 (CENAMAD) and is thankful for the support received from ANID SCIA-Anillo ACT210052 and grants DIUFRO DI20-0066 & DI22-2003.
Author information
Authors and Affiliations
Contributions
Research design by RG, AP; Fieldwork by RG, JU; Data analysis by EFL, JU; Manuscript writing and editing by JU, EFL, AFR, AP, RG.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ulloa, J., Fuentes-Lillo, E., Fuentes-Ramírez, A. et al. Native bamboo increases biotic resistance to Pinus contorta invasion in temperate forest ecosystems. Biol Invasions 25, 3905–3915 (2023). https://doi.org/10.1007/s10530-023-03147-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-023-03147-8