
Abstract
The brown widow spider, Latrodectus geometricus, is a hazardous and highly invasive species globally. One of the suggested mechanisms enhancing this spider’s invasiveness is lower susceptibility to natural enemies compared to other widow species. The parasitoid wasp, Philolema latrodecti, is known to attack egg sacs of L. geometricus, as well as those of other species of Latrodectus. Despite its potential importance as a natural enemy of L. geometricus, little is known about the development of this wasp and factors mediating its parasitism success. We investigated the development of this parasitoid in two widow spider host species, one native to Israel, the white widow, L. pallidus, and the invasive brown widow, L. geometricus. We compared wasp developmental success in egg sacs exposed to increasing wasp densities. We found increased wasp emergence and brood size, and smaller emerging wasp body size with increasing wasp density. Across all wasp densities, more and larger parasitoids developed in the egg sacs of the native host species, L. pallidus, compared to the invasive host species, L. geometricus. Parasitoid sex ratio was highly female-biased and the proportion of males increased with wasp density, suggesting local mate competition. Overall, our results suggest that L. pallidus is a better host for the egg sac parasitoid P. latrodecti, which could give a competitive advantage to the less-affected invasive brown widow spider.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Globalization and international trade increase transport and establishment of alien species to new environments (Holden et al. 2022; Hulme 2009; Parepa et al. 2013). Ecological and evolutionary hypotheses focused on invasive species traits have been proposed to explain invasion success, including high fecundity, high dispersal rate and high competitiveness of the invasive species (Abhilasha and Joshi 2009; Callaway and Ridenour 2004; Moravcová et al. 2015; Rehage and Sih 2004). In addition, interactions with natural enemies such as predators, parasites, and parasitoids may play a crucial role in the determining ability of a species to invade (Elton 1958; Heimpel and Mills 2017; Newsome and Noble 1986; Ricklefs and Cox 1978; White and Perkins 2012). Invasive species may have traits that lower their susceptibility to parasitism throughout their distribution. For example, the invasive harlequin ladybird beetle has chemical defenses that make the species unpalatable to predators and defend against infection (Ceryngier et al. 2018). Regardless of the mechanisms involved, lower susceptibility to parasites may give an invasive species a relative advantage over native species. Hence, comparing parasitism success on invasive versus native species can contribute to understanding the mechanisms of successful invasive establishment.
Studies of invasive species are biased towards relatively few taxonomic groups, which can affect our understanding of invasion mechanisms and success. Over 60% of studies focus on insects and plants (Pysek et al. 2008), while other groups are understudied. Studies related to invasive spiders have only recently started to attract scientific attention (Campbell et al. 2020; Mowery et al. 2021; Narimanov et al. 2021; Nentwig 2015), despite high prevalence both in agricultural (Hogg and Daane 2010) and natural habitats (Pétillon et al. 2020) and their essential role as generalist predators (Birkhofer et al. 2017; Michalko et al. 2019). Parasitoids are common and important predators of spiders at all life stages, including the egg sacs (Fei et al. 2023). Egg sac parasitoids may find hosts using species-specific spider silk cues, such as in a parasitoid of linyphiid spider egg sacs (van Baarlen et al. 1996). Spider egg sacs may differ in their quality for the parasitoid. For example, larger spider host egg sac mass was found to increase resulting parasitoid offspring body size (Morse 1994). Yet, we lack direct comparisons of predation and parasitism in congeneric invasive and native spider hosts (Snyder and Evans 2006).
The brown widow spider, L. geometricus, is an invasive species worldwide, in some cases outcompeting native species and threatening human health, as its bite can cause a serious reaction requiring hospitalization (Goddard et al. 2008; Marie and Vetter 2015; Muller 1993). One of the main natural enemies of widow spiders, including the brown widow, is the gregarious parasitoid wasp P. latrodecti (Eurytomidae, Hymenoptera). A single wasp will lay a clutch of eggs inside the spider’s egg sac, and multiple wasps emerge after consuming all the spider eggs, thereby completing their development inside the egg sac. It has been suggested that the brown widow spider is less susceptible to egg sac parasitism due to the occurrence of silk spike-like structures produced by the female spider on the surface of its egg sac (Vetter et al. 2012). Consistent with this idea, previous studies found lower parasitism rates on egg sacs of the invasive brown widow compared to native widow spider species in both field and lab settings (Mowery et al. 2022; Triana et al. 2012; Vetter et al. 2012). However, despite its potential ecological importance, very little is known about the development of this parasitoid inside the spider egg sac and on factors mediating its success.
The suitability and quality of hosts to parasitoids are often tested under unrealistic conditions, where a single host is exposed to a single parasitoid. In nature, when parasitoid density is high, several females may compete for a host, with more than one female parasitizing the same host individual (termed super-parasitism), thus reducing resource availability for the developing offspring (Godfray 1994; Van Alphen and Visser 1990). The outcome of super-parasitism may depend on the host species and quality, as well as on the relative timing of multiple ovipositions and the rate of depletion of resources inside of the host egg sac. Measuring successful development and rates of developmental failure may provide insight into the effects of superparasitism on parasitoid development (Keasar et al. 2006). However, to our knowledge, no studies have considered the effects of intraspecific competition and super-parasitism in invasive versus native hosts.
In this study, we first describe the development of P. latrodecti in widow spider egg sacs. This is one of few descriptions of the development of an egg sac parasitoid in general. We compared parasitism success in egg sacs of two widow spider host species, one native to Israel, the white widow, L. pallidus (Levy and Amitai 1983), and the invasive brown widow, L. geometricus, following exposure to an increasing number of parasitoid wasps (from one to four female wasps) under controlled conditions. We also measured parasitoid fecundity before and after feeding on honey to assess the potential for egg depletion, which could affect the number of eggs laid in the host. We predicted that parasitism success and brood size would increase with the number of parasitizing wasps up to a threshold, after which it could level off or even decrease. This may occur if female wasps interfere with one another preventing oviposition, or if the larvae compete for resources inside the host, preventing some of the parasitoids from completing their development (Hassell and Waage 1984; Van Alphen and Visser 1990). In addition, we predicted lower development success of the parasitoid on the egg sacs of the invasive L. geometricus. This prediction was based on the presumably better egg sac defences of L. geometricus, as well as on the smaller size of their egg sacs, which contain fewer spider eggs on average than L. pallidus egg sacs (Danielsen et al. 2014; Levy and Amitai 1983; Mowery et al. 2022). Female parasitoids adjust their offspring sex ratios based on host quality (Charnov 1982; Godfray 1994; King 1987) and conditions of superparasitism (Shuker et al. 2005). Evidence of varying sex ratios may also shed light on the wasp mating system (Werren 1983). We have compared the sex ratio of wasps emerging out of egg sacs of the two host species and at increasing parasitoid densities, and predicted proportionately more males in lower-quality L. geometricus and superparasitized hosts,
Methods
Spider and wasp rearing
We kept field-collected adult female L. geometricus and L. pallidus in laboratory conditions (25 ± 1 °C, 14:10 L:D). Females were fed weekly with one grasshopper nymph (Schistocerca gregaria). Spider egg sacs used in all experiments were less than seven days old. Wasps for the experiments emerged from L. pallidus spider egg sacs collected in the field (Hatzerim and Beer Sheva area, Israel). We fed wasps with honey immediately after emergence.
Development of P. latrodecti in L. geometricus egg sacs
We exposed egg sacs (n = 12) of L. geometricus each to a single P. latrodecti parasitoid female in a plastic vial. We then incubated the egg sacs under controlled conditions (25 ± 1 °C, 14:10 L:D). On days one, three, five, 10, 15, and 20 post-parasitism, we dissected two parasitized egg sacs each day and photographed and recorded parasitoid development using a Nikon stereomicroscope (SMZ-18) with a Nikon digital camera (DS-FI2).
Wasp density
We observed the effect of the presence of multiple parasitoid wasps on the outcome of parasitism in L. geometricus and L. pallidus egg sacs by exposing the egg sacs to one (L.g.: N = 69, L.p.: N = 26), two (L.g.: N = 33, L.p.: N = 30), three (L.g.: N = 37, Lp: N = 17), or four (L.g.: N = 35, Lp: N = 17) parasitoid females simultaneously. The sample size for each treatment was determined by the availability of fresh egg sacs (less than one week old) and wasps. Female wasps normally survived for few days under these conditions. After two months, we recorded the contents of each egg sac as: adult wasps that emerged; adult wasps that developed but did not successfully emerge; underdeveloped wasps; unhatched spider eggs; or emerged spiderlings. In egg sacs from which wasps emerged, we counted the total number of emerging wasps. Using a Nikon stereomicroscope and digital camera (see above), we photographed and measured the body length of a sample of 10 females and up to 10 males (depending on availability) per egg sac. We measured wasp body length from the anterior of the prosoma to the posterior of the metasoma using NIS-Elements D software. We recorded the sex ratio of wasps emerging from each egg sac, differentiating between males and females according to their metasomal size and shape, their funicular pubescence and presence or absence of an ovipositor (Burks 1971). Sex ratio was calculated as the number of adult males divided by the total number of adult offspring in the egg sac.
P. latrodecti fecundity
We dissected newly emerged, unfed adult female P. latrodecti wasps (n = 25) under a Nikon stereomicroscope on the day of their emergence and counted the number of eggs in their ovaries. We noted the size distribution of eggs of each female to assess the occurrence of immature eggs. To determine whether females mature more eggs following emergence, we fed wasps with honey solution and after three days, dissected them (n = 20) and counted their eggs.
Statistical methods
All analyses were conducted in R (version 4.0.2, R Core Team 2023). To compare measures of parasitism development, we first compared rates of failed parasitoid development in the two species’ egg sacs using Fisher’s exact test. We then compared parasitism emergence success from L. geometricus and L. pallidus egg sacs exposed to 1–4 wasps using a Generalized Linear Model (GLM) with successful parasitoid emergence as the binary response variable (yes or no), and species and wasp density as well as the interaction between them as predictors, with a binomial distribution and a logit link function in R (lme4; R Studio, Inc, Bates et al. 2015). We compared the total number of wasps emerging from the egg sacs of each spider species and at each wasp density using a generalized linear model with the number of wasps emerging as the response variable, modelled with a quasi-Poisson error distribution due to overdispersion, and wasp density, spider species, and the interaction between wasp density and spider species as predictors. In cases where data were found to be overdispersed based on a dispersion parameter greater than one, quasi-models were used.
To compare body size of emerging wasps we constructed a generalized linear mixed model (GLMM) with wasp density treatment and spider species as fixed factors, the body size of female wasps (up to ten per egg sac) as response variables, egg sac ID as a random factor, and a Gamma error distribution with a log link function. For comparing sex ratio (males/total offspring) among wasp density treatments and host species, we used a GLMM with sex ratio as a binomial outcome, with egg sac ID as a random factor. To assess the effect of brood size on emerging female wasp body length, we used a GLMM with wasp body size as the response variable with a Gamma distribution and log link function, species and brood size as the predictors, with egg sac ID as a random factor. Parameters and significance were calculated using Wald F tests for GLMs and Chi-squared testes for GLMMs (car package, Fox and Weisberg 2019), and multiple comparisons were conducted using Tukey HSD post-hoc tests using the multcomp package (Hothorn et al. 2022) in R. To compare female egg load before and after feeding, we used a two-tailed t-test.
Results
Development of P. latrodecti in L. geometricus
The different developmental stages of P. latrodecti are presented in Fig. 1. After one day, P. latrodecti eggs were found next to the spider eggs, close to the inner surface of the egg sac. P. latrodecti eggs hatched after two to three days inside of the egg sac. The larvae fed on the content of L. geometricus spider eggs, leaving empty spider egg shells, pupated by 15 days after oviposition, and emerged after 20 days of development inside of the spider egg sac after depleting all spider eggs.

Development of Philolema latrodecti in egg sacs of Latrodectus geometricus observed on days one (a, egg), three (b, larva), five (c, larva), 10 (d, larva), 15 (e, pupa), and 20 (f, adult wasp) after P. latrodecti female oviposition
Wasp density and host species
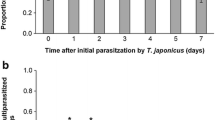
Increasing wasp density resulted in an increasing proportion of egg sacs in which parasitoids successfully emerged (Fig. 2a, GLM, F1,324 = 13.138, P < 0.001), with no significant effects of host species egg sac (F1,324 = 0.0005, P = 0.982) or the interaction between wasp density and host species (F1,324 = 1.181, P = 0.278). In particular, wasps were more likely to emerge from egg sacs exposed to three and four wasps compared to those exposed to a single wasp (Tukey HSD, three wasps: z = 2.874, P = 0.024; four wasps: z = 2.985, P = 0.017). In most cases where wasps did not emerge, spiderlings emerged out of the egg sac, suggesting that no parasitism occurred. However, in a few cases, we found evidence that the wasps failed to complete their development or to emerge out of the egg sac. This occurred in 7 out of 107 L. geometricus egg sacs and in one out of 65 L. pallidus egg sacs (Fisher’s exact test P = 0.261) and only occurred in cases where the egg sac was exposed to more than one wasp. However, the number of cases in which parasitoids were undeveloped may have been too low to detect differences among species. In a few cases, we observed behavioral interactions between wasps such as jumping off the egg sac following contact with another wasp that may have resulted in interference in oviposition.

a Parasitoid wasp (P. latrodecti) emergence success from L. geometricus egg sacs (brown bars) and L. pallidus (white bars) exposed to one, two, three, and four parasitoid wasps. Error bars are standard errors calculated for the proportions. b Number of parasitoid wasps emerged after exposure of L. pallidus egg sacs (white) and L. geometricus egg sacs (brown) to one, two, three, and four P. latrodecti females. Samples sizes indicate total number of egg sacs in each treatment. Boxplots show median and 25th and 75th percentiles. The whiskers on the plots indicate values within 1.5 times the interquartile range, and the outliers are indicated with black points
More wasps emerged with increasing wasp density (Fig. 2b, GLM, F1,133 = 20.825, P < 0.001) and from L. pallidus egg sacs compared to L. geometricus egg sacs (F1,133 = 11.023, P = 0.001), with no significant interaction between wasp density and host species (F1,133 = 0.280, P = 0.597). More wasps emerged from egg sacs exposed to more than one wasp than from the the single wasp exposure (Tukey HSD, one-two wasps: z = 3.970, P < 0.001; one-three wasps, z = 2.568, P = 0.059; one-four wasps: z = 5.189, P < 0.001). Larger female wasps emerged from L. pallidus egg sacs (Fig. 3a, GLMM, X21 = 19.520, P < 0.001) and from egg sacs exposed to lower wasp densities (X21 = 13.654, P < 0.001), with no interaction between host species and wasp density (X21 = 0.009, P = 0.925). We found a negative relationship between emerging wasp brood size and body size (Fig. 3b). Mean female wasp body size decreased with increasing brood size in both host species (GLMM, X21 = 69.101, P < 0.001), and wasps were consistently larger when developing in L. pallidus egg sacs (X21 = 41.423, P < 0.001), with no significant interaction between brood size and host species (X21 = 2.526, P = 0.112).

a Female parasitoid wasp body size in L. geometricus (brown) and L. pallidus egg sacs (white) after parasitism by one to four P. latrodecti females. Boxplots show median and 25th and 75th percentiles. The whiskers on the plots indicate values within 1.5 times the interquartile range, and the outliers are indicated with black points. b Parasitoid P. latrodecti brood size and female parasitoid body size in parasitoids that developed in L. geometricus (brown) and L. pallidus (white) egg sacs
Wasp density also affected the resulting sex ratio of the wasps emerging. Sex ratio was generally female biased. The proportion of males increased with increasing wasp density (Fig. 4, quasi-binomial GLM, F1,126 = 76.728, P < 0.001), but was not affected by the host species (F1,126 = 0.911, P = 0.342), nor was there an interaction between wasp density and host species (F1,126 = 0.070, P = 0.791).

Parasitoid wasp sex ratio (proportion of males out of all emerging offspring) in L. geometricus (brown) and L. pallidus host egg sacs (white) after parasitism by one to four P. latrodecti adult female wasps. Boxplots show median and 25th and 75th percentiles. The whiskers on the plots indicate values within 1.5 times the interquartile range, and the outliers are indicated with black points
P. latrodecti fecundity
All eggs from dissected P. latrodecti females were similarly shaped and sized, suggesting that they were already mature upon wasp emergence. The mean number of P. latrodecti eggs before feeding was 39 ± 1.6 eggs (mean ± SE; n = 25), and was not significantly different from the number of eggs in females dissected three days after feeding (40 ± 2.2, mean ± SE; n = 20 fed females, t = 0.518, P = 0.608).
Discussion
We compared measures of parasitism success of the egg sac parasitoid wasp P. latrodecti at increasing parasitoid densities between two host species: the globally invasive brown widow spider L. geometricus, and the white widow spider L. pallidus, native to Israel. We found evidence for increased parasitism success (wasp emergence) with increasing wasp density, and some evidence that the invasive species is an inferior host, as more and larger wasps emerged from native L. pallidus compared to L. geometricus egg sacs. These, combined with previous results (Mowery et al. 2022), suggest the lower susceptibility and suitability of L. geometricus to parasitism, potentially contributing to the invasion success of this species.
Greater parasitism success with greater wasp density was observed in previous studies (Gu and Dorn 2003; Khafagi and Hegazi 2008; Silva-Torres et al. 2009). In our study, however, succesful parasitism rate did not increase proportionally with wasp density, and differed mainly between the single versus multiple wasp treatments. A possible reason could be interference competition between wasps at higher densities that prevented females from laying their eggs. Direct interference between ovipositing females was observed in many parasitoid species (including our observations of P. latrodecti, current study) that could reduce parasitism success (Boivin and Martel 2012; Grenier et al. 2001; Huang et al. 2017). Failed parasitism could also result from intense resource competition of larvae within the host, causing parasitoid brood death. Indeed, in some cases we found dead immature wasps, or wasps that failed to emerge, inside the egg sacs. These cases of failed emergence occurred only when the egg sac was exposed to multiple wasps, and mostly in the smaller L. geometricus egg sacs, and could be related to extreme resource shortage within the host (Harvey et al. 2013; Wajnberg et al. 1990).
We found an increasing number of offspring emerging from egg sacs that were exposed to several wasps, compared to a single wasp, which suggests the occurrence of multiple parasitism events inside a single host (i.e., super-parasitism, (Dorn and Beckage 2007; Roitberg et al. 1992; Van Alphen and Visser 1990). However, similarly to parasitism success, brood size did not increase proportionally with wasp density. In addition, parasitoid brood size in L. geometricus egg sacs was smaller than in L. pallidus egg sacs. These observed patterns were probably at least partially due to fewer eggs laid per female wasp at higher wasp densities (Harvey 2000; Le Masurier 1991; West et al. 2001; Zaviezo and Mills 2000), and on the inferior host (Barrett and Schmidt 1991), but could also be due to lower survival of the developing wasps within the host due to resource shortage (Godfray 1994; Harvey 2005; Mackauer et al. 1997; Vinson and Iwantsch 1980). In the current experiment, we could not distinguish between these two options, since it is impossible to count wasp eggs laid inside the host without interfering with their development.
Results on wasp body size further supported the occurrence of resource shortage within the host. The body size of emerging females was smaller when developing in egg sacs exposed to multiple wasps, and in those developing on the inferior host L. geometricus compared to L. pallidus. This is in accordance with many studies suggesting that host quality affects parasitoids’ adult size (Farahani and Goldansaz 2013; Godfray 1994). Moreover, wasp body size was negatively affected by brood size in both species, suggesting a trade-off between the number and size of offspring, as observed in other parasitoids (Gu and Dorn 2003; Hardy 1992; Takagi 1987).
Despite the lack of alternatives hosts in our experiment, females did not seem to lay all of their eggs in a single host, even in the absence of competition. This is inferred by the smaller number of wasps emerging from host exposed to a single female (20–30), compared to the mean egg load of females (~ 40). In addition, in another experiment, we dissected females soon after they oviposited in the host, and found that usually eggs were still available in their reproductive tract (A. D. Johnson, unpublished data). The wasps may be adapted to spread their eggs among several hosts as a bet-hedging strategy (Cronin and Strong 1993; Hopper 1999), or to optimize their lifetime reproductive success (Bezemer and Mills 2003; Sevenster et al. 1998). Future studies should characterize clutch size decisions of these parasitoids when foraging freely in nature and while searching for native or invasive hosts.
Parasitoid sex ratio was highly female-biased in both host species. Although little is known about the mating behavior of these wasps, a skewed sex ratio suggests that mating in this species occurs locally inducing local mate competition between related males (Hamilton 1967; Werren 1980). Personal observations of P. latrodecti matings soon after emergence are consistent with this interpretation, although it should be considered that in our experimental set-up, dispersing away from the natal host was not possible. The observed increase in sex ratio with the increasing number of competing females is also consistent with the predictions of local mate competition theory, as well as with empirical evidence for increased sex ratio when the number of female founders in a patch increases (Hardy 1994; Herre 1987). However, even at a high wasp density sex ratio remained highly female-biased (~ 20% males), suggesting the potential occurrence of additional mechanisms that might favor a biased sex ratio, such as high relatedness between ovipositing females (Herre 1985).
We provide one of the first descriptions of wasp development inside a spider egg sac. Other studies reported the occurrence of egg sac parasitism in different spider species (Austin 1985; Bowden and Buddle 2012; Cobb and Cobb 2004; Finch 2005) including L. geometricus (Marie and Vetter 2015; Moura et al. 2021; Vetter et al. 2012), but none observed the developmental sequence from egg, throughout the different larval stages, to the pupal and adult stage. Our observations suggest that the eggs are laid close to the egg sac surface, and that the larvae hatch soon after egg laying and grow while consuming the contents of the spider eggs, almost always depleting the eggs (in very few cases we observed the emergence of both spiderlings and wasps). As the number of wasps emerging from an egg sac (3–76 wasps in our study) is lower than the number of spider eggs in a egg sacs of either species (100–300 eggs, Levy and Amitai 1983), each wasp must feed on multiple eggs during its development. Egg sac parasitoids are sometimes considered predators rather than parasitoids as they feed on several prey items during their lifetime (Eggleton and Belshaw 1992; Eggleton and Gaston 1990; Strand and Obrycki 1996). However, given the phylogenetic background, and the life cycle of this wasp, we think it is more reasonable to consider it a parasitoid (Cobb and Cobb 2004; Fei et al. 2023), with the whole egg sac, rather than a single egg, as a host.
To our knowledge, rarely have parasitism and predation been compared in native versus invasive spiders. One example involves a North American dwarf spider, Mermessus trilobatus, that has undergone a range expansion in Europe, spreading by more than 1000 km in less than 50 years (Hirna 2017; Schmidt et al. 2008). This invasion success was once thought to be attributed to reduced susceptibility to native predators of M. trilobatus in Europe. However, when Narimanov et al. (2021) compared the susceptibility of the invasive M. trilobatus and a native Erigone dentipalpis (Araneae: Linyphiidae) to a shared native predator, they found that the invasive species was consumed almost three times as often as the native species. In contrast, in stink bugs, invasive hosts were accepted equally by parasitoids compared to a native species, but there was no successful development in the invasive host (Abram et al. 2014). Parasitism can result in significant declines in native host populations (Abram et al. 2020), and thus could increase successful establishment of an invasive species that is not highly parasitized. Our study supports the notion of lower suitability and susceptibility of the invasive L. geometricus to parasitism under varying environmental conditions, either due to their smaller size, better defenses, or other aspects that were not measured here, which may potentially promote this species’ invasion success. Such knowledge enhances our understanding of biological invasions and may help better manage species invasions.
Data availability
All data associated with this manuscript are available on the Dryad data repository at https://doi.org/10.5061/dryad.tb2rbp053
References
Abhilasha D, Joshi J (2009) Enhanced fitness due to higher fecundity, increased defence against a specialist and tolerance towards a generalist herbivore in an invasive annual plant. J Plant Ecol 2(2):77–86. https://doi.org/10.1093/jpe/rtp008
Abram PK, Gariepy TD, Boivin G, Brodeur J (2014) An invasive stink bug as an evolutionary trap for an indigenous egg parasitoid. Biol Invasions 16(7):1387–1395. https://doi.org/10.1007/s10530-013-0576-y
Abram PK, Mills NJ, Beers EH (2020) Review: Classical biological control of invasive stink bugs with egg parasitoids—what does success look like? Pest Manag Sci 76(6):1980–1992. https://doi.org/10.1002/ps.5813
Austin AD (1985) The function of spider egg sacs in relation to parasitoids and predators, with special reference to the Australian fauna. J Nat Hist 19(2):359–376. https://doi.org/10.1080/00222938500770261
Barrett M, Schmidt JM (1991) A comparison between the amino acid composition of an egg parasitoid wasp and some of its hosts. Entomol Exp Appl 59(1):29–41. https://doi.org/10.1111/j.1570-7458.1991.tb01483.x
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1. https://doi.org/10.18637/jss.v067.i01
Bezemer TM, Mills NJ (2003) Clutch size decisions of a gregarious parasitoid under laboratory and field conditions. Anim Behav 66(6):1119–1128. https://doi.org/10.1006/anbe.2003.2296
Birkhofer K, Gossner MM, Diekötter T, Drees C, Ferlian O, Maraun M, Scheu S, Weisser WW, Wolters V, Wurst S, Zaitsev AS, Smith HG (2017) Land-use type and intensity differentially filter traits in above- and below-ground arthropod communities. J Anim Ecol 86(3):511–520. https://doi.org/10.1111/1365-2656.12641
Boivin G, Martel V (2012) Size-induced reproductive constraints in an egg parasitoid. J Insect Physiol 58(12):1694–1700. https://doi.org/10.1016/j.jinsphys.2012.10.014
Bowden JJ, Buddle CM (2012) Egg sac parasitism of Arctic wolf spiders (Araneae: Lycosidae) from northwestern North America. J Arachnol 40(3):348–350. https://doi.org/10.1636/P11-50.1
Burks BD (1971) A Synopsis of the genera of the family Eurytomidae (Hymenoptera: Chalcidoidea). Trans Am Entomolog Soc (1890) 97(1):1–89
Callaway RM, Ridenour WM (2004) Novel weapons: Invasive success and the evolution of increased competitive ability. Front Ecol Environ 2(8):436–443. https://doi.org/10.1890/1540-9295(2004)002[0436:NWISAT]2.0.CO;2
Campbell LD, Salazar AR, Rypstra AL (2020) Contests between web spiders: patience as a strategy for successful invasion. Biol Invasions 22(5):1737–1752. https://doi.org/10.1007/s10530-020-02216-6
Ceryngier P, Nedvěd O, Grez AA, Riddick EW, Roy HE, San Martin G, Steenberg T, Veselý P, Zaviezo T, Zúñiga-Reinoso Á, Haelewaters D (2018) Predators and parasitoids of the harlequin ladybird, Harmonia axyridis, in its native range and invaded areas. Biol Invasions 20(4):1009–1031. https://doi.org/10.1007/s10530-017-1608-9
Charnov EL (1982) The theory of sex allocation. Monogr Popul Biol 18:1–355
Cobb LM, Cobb VA (2004) Occurrence of Parasitoid Wasps, Baeus sp., and Gelis sp., in the egg sacs of the wolf spiders Pardosa moesta and Pardosa sternalis (Araneae, Lycosidae) in Southeastern Idaho. Can Field Nat 118(1):1. https://doi.org/10.22621/cfn.v118i1.894
Cronin JT, Strong DR (1993) Superparasitism and mutual interference in the egg parasitoid Anagrus delicatus (Hymenoptera: Mymaridae). Ecol Entomol 18(4):293–302
Danielsen DWR, Clarke DE, Valle SJ, Ansalmo AA, Vincent LS, Vetter RS (2014) Natural egg sac clutch size of the brown widow spider, Latrodectus geometricus (Araneae: Theridiidae) in southern California. Bull South Calif Acad Sci 113(2):100–102. https://doi.org/10.3160/0038-3872-113.2.100
Dorn S, Beckage NE (2007) Superparasitism in gregarious hymenopteran parasitoids: ecological, behavioural and physiological perspectives. Physiol Entomol 32(3):199–211. https://doi.org/10.1111/j.1365-3032.2007.00576.x
Eggleton P, Belshaw R (1992) Insect parasitoids: an evolutionary overview. Philos Trans R Soc Lond Series B Biol Sci 337(1279):1–20. https://doi.org/10.1098/rstb.1992.0079
Eggleton P, Gaston KJ (1990) “Parasitoid” species and assemblages: Convenient definitions or misleading compromises? Oikos 59(3):417–421. https://doi.org/10.2307/3545155
Elton CS (1958) The reasons for conservation. In: Elton CS (ed) The ecology of invasions by animals and plants. Springer, US, pp 143–153. https://doi.org/10.1007/978-1-4899-7214-9_8
Farahani HK, Goldansaz SH (2013) Is host age an important factor in the bionomics of Apanteles myeloenta (Hymenoptera: Braconidae)? Eur J Entomol 110(2):277–283. https://doi.org/10.14411/eje.2013.041
Fei M, Gols R, Harvey JA (2023) The biology and ecology of parasitoid wasps of predatory arthropods. Ann Rev Entomol. https://doi.org/10.1146/annurev-ento-120120-111607
Finch O (2005) The parasitoid complex and parasitoid-induced mortality of spiders (Araneae) in a Central European woodland. J Nat Hist 39(25):2339–2354. https://doi.org/10.1080/00222930502005720
Fox J, Weisberg S (2019) An R companion to applied regression (Third edition). Sage. https://socialsciences.mcmaster.ca/jfox/Books/Companion/
Goddard J, Upshaw S, Held D, Johnnson K (2008) Severe reaction from envenomation by the brown widow spider, Latrodectus geometricus (Araneae: Theridiidae). South Med J 101(12):1269–1270. https://doi.org/10.1097/smj.0b013e31817f4d89
Godfray HCJ (1994) Parasitoids: behavioral and evolutionary ecology, vol 67. Princeton University Press, New York. https://doi.org/10.2307/j.ctvs32rmp
Grenier S, Basso C, Pintureau B (2001) Effects of the host species and the number of parasitoids per host on the size of some Trichogramma species (Hymenoptera: Trichogrammatidae). Biocontrol Sci Tech 11(1):21–26. https://doi.org/10.1080/09583150020029709
Gu H, Dorn S (2003) Mating system and sex allocation in the gregarious parasitoid Cotesia glomerata. Anim Behav 66(2):259–264. https://doi.org/10.1006/anbe.2003.2185
Hamilton WD (1967) Extraordinary sex ratios. Science 156(3774):477–488. https://doi.org/10.1126/science.156.3774.477
Hardy ICW (1992) Non-binomial sex allocation and brood sex ratio variances in the parasitoid hymenoptera. Oikos 65(1):143–158. https://doi.org/10.2307/3544897
Hardy ICW (1994) Sex ratio and mating structure in the parasitoid Hymenoptera. Oikos 69(1):3–20. https://doi.org/10.2307/3545278
Harvey JA (2000) Dynamic effects of parasitism by an endoparasitoid wasp on the development of two host species: implications for host quality and parasitoid fitness. Ecol Entomol 25(3):267–278. https://doi.org/10.1046/j.1365-2311.2000.00265.x
Harvey JA (2005) Factors affecting the evolution of development strategies in parasitoid wasps: the importance of functional constraints and incorporating complexity. Entomol Exp Appl 117(1):1–13. https://doi.org/10.1111/j.1570-7458.2005.00348.x
Harvey JA, Poelman EH, Tanaka T (2013) Intrinsic inter- and intraspecific competition in parasitoid wasps. Annu Rev Entomol 58(1):333–351. https://doi.org/10.1146/annurev-ento-120811-153622
Hassell MP, Waage JK (1984) Host-parasitoid population interactions. Annu Rev Entomol 29(1):89–114. https://doi.org/10.1146/annurev.en.29.010184.000513
Heimpel GE, Mills NJ (2017) Biological control. Cambridge University Press, Cambridge
Herre EA (1985) Sex ratio adjustment in fig wasps. Science 228(4701):896–898. https://doi.org/10.1126/science.228.4701.896
Herre EA (1987) Optimality, plasticity and selective regime in fig wasp sex ratios. Nature 329(6140):6140. https://doi.org/10.1038/329627a0
Hirna A (2017) First record of the alien spider species Mermessus trilobatus (Araneae: Linyphiidae) in Ukraine. Arachnologische Mitteilungen 54:41–43. https://doi.org/10.5431/aramit5409
Hogg BN, Daane KM (2010) The role of dispersal from natural habitat in determining spider abundance and diversity in California vineyards. Agr Ecosyst Environ 135(4):260–267. https://doi.org/10.1016/j.agee.2009.10.004
Holden CA, Bailey JP, Taylor JE, Martin F, Beckett P, McAinsh M (2022) Know your enemy: Application of ATR-FTIR spectroscopy to invasive species control. PLoS ONE 17(1):e0261742. https://doi.org/10.1371/journal.pone.0261742
Hopper K (1999) Risk-spreading and bet-hedging in insect population biology. Annu Rev Entomol 44:535–560. https://doi.org/10.1146/annurev.ento.44.1.535
Hothorn T, Bretz F, Westfall P (2008) “Simultaneous Inference in General Parametric Models.” Biom J 50(3):346–363
Huang J, Hua H-Q, Wang L-Y, Zhang F, Li Y-X (2017) Number of attacks by Trichogramma dendrolimi (Hymenoptera: Trichogrammatidae) affects the successful parasitism of Ostrinia furnacalis (Lepidoptera: Crambidae) eggs. Bull Entomol Res 107(6):812–819. https://doi.org/10.1017/S0007485317000335
Hulme PE (2009) Trade, transport and trouble: managing invasive species pathways in an era of globalization. J Appl Ecol 46(1):10–18. https://doi.org/10.1111/j.1365-2664.2008.01600.x
Keasar T, Segoli M, Barak R, Steinberg S, Giron D, Strand MR, Bouskila A, Harari AR (2006) Costs and consequences of superparasitism in the polyembryonic parasitoid Copidosoma koehleri (Hymenoptera: Encyrtidae). Ecological Entomology 31(3):277–283. https://doi.org/10.1111/j.1365-2311.2006.00788.x
Khafagi WE, Hegazi EM (2008) Does superparasitism improve host suitability for parasitoid development? A case study in the Microplitis rufiventris-Spodoptera littoralissystem. Biocontrol 53(3):427–438. https://doi.org/10.1007/s10526-007-9078-0
King BH (1987) Offspring sex ratios in parasitoid wasps. Q Rev Biol 62(4):367–396. https://doi.org/10.1086/415618
Le Masurier AD (1991) Effect of host size on clutch size in Cotesia glomerata. J Anim Ecol 60(1):107–118. https://doi.org/10.2307/5448
Levy G, Amitai P (1983) Revision of the widow-spider genus Latrodectus (Araneae: Theridiidae) in Israel. Zool J Linn Soc 77(1):39–63. https://doi.org/10.1111/j.1096-3642.1983.tb01720.x
Mackauer M, Sequeira R, Otto M (1997) Growth and development in parasitoid wasps: adaptation to variable host resources. In: Dettner K, Bauer G, Völkl W (eds) Vertical food web interactions: evolutionary patterns and driving forces. Springer, London, pp 191–203. https://doi.org/10.1007/978-3-642-60725-7_11
Marie J, Vetter RS (2015) Establishment of the brown widow spider (Araneae: Theridiidae) and infestation of its egg sacs by a parasitoid, Philolema latrodecti (Hymenoptera: Eurytomidae), in French Polynesia and the Cook Islands. J Med Entomol 52(6):1291–1298. https://doi.org/10.1093/jme/tjv127
Michalko R, Pekár S, Dul’a M, Entling MH (2019) Global patterns in the biocontrol efficacy of spiders: a meta-analysis. Glob Ecol Biogeogr 28(9):1366–1378. https://doi.org/10.1111/geb.12927
Moravcová L, Pyšek P, Jarošík V, Pergl J (2015) Getting the right traits: reproductive and dispersal characteristics predict the invasiveness of herbaceous plant species. PLoS ONE 10(4):e0123634. https://doi.org/10.1371/journal.pone.0123634
Morse DH (1994) The effect of host size on sex ratio in the ichneumonid wasp, Trychosis cyperia, a spider parasitoid. Am Midl Nat 131(2):281–287. https://doi.org/10.2307/2426254
Moura RR, Tozzo Neto A, Gonzaga MO (2021) Don’t put all your eggs in small baskets: Ineffective guardians, incidence of parasitoids and clutch size of Latrodectus geometricus (Araneae, Theridiidae) along an urban gradient. Zool Anz 295:120–125. https://doi.org/10.1016/j.jcz.2021.10.005
Mowery MA, Vink C, Mason AC, Andrade MCB (2021) Behavioural, morphological, and life history shifts during invasive spread. Biol Invasions. https://doi.org/10.1007/s10530-021-02593-6
Mowery MA, Arabesky V, Lubin Y, Segoli M (2022) Differential parasitism of native and invasive widow spider egg sacs. Behav Ecol 33(3):565–572. https://doi.org/10.1093/beheco/arac017
Muller GJ (1993) Black and brown widow spider bites in South Africa. A series of 45 cases. S Afr Med J 83(6):399–405
Narimanov N, Hatamli K, Entling MH (2021) Prey naïveté rather than enemy release dominates the relation of an invasive spider toward a native predator. Ecol Evol 11(16):11200–11206. https://doi.org/10.1002/ece3.7905
Nentwig W (2015) Introduction, establishment rate, pathways and impact of spiders alien to Europe. Biol Invasions 17(9):2757–2778. https://doi.org/10.1007/s10530-015-0912-5
Newsome AE, Noble IR (1986) Ecological and physiological characters of invading species. In: R.H. Groves & J.J. Burdon (eds.), Ecology of biological invasions: an Australian perspective. Australian Academy of Science, Canberra, pp 1–20
Parepa M, Fischer M, Bossdorf O (2013) Environmental variability promotes plant invasion. Nat Commun 4(1):1. https://doi.org/10.1038/ncomms2632
Pétillon J, Privet K, Roderick GK, Gullespie RG, Price DK (2020) Non-native spiders change assemblages of Hawaiian forest fragment kipuka over space and time. NeoBiota 55:1
Pysek P, Richardson DM, Pergl J, Jarosík V, Sixtová Z, Weber E (2008) Geographical and taxonomic biases in invasion ecology. Trends Ecol Evol 23(5):237–244. https://doi.org/10.1016/j.tree.2008.02.002
R Core Team (2023) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Rehage JS, Sih A (2004) Dispersal behavior, boldness, and the link to invasiveness: a comparison of four Gambusia species. Biol Invasions 6(3):379–391. https://doi.org/10.1023/B:BINV.0000034618.93140.a5
Ricklefs RE, Cox GW (1978) Stage of taxon cycle, habitat distribution, and population density in the avifauna of the West Indies. Am Nat 112(987):875–895. https://doi.org/10.1086/283329
Roitberg BD, Mangel M, Lalonde RG, Roitberg CA, van Alphen JJM, Vet L (1992) Seasonal dynamic shifts in patch exploitation by parasitic wasps. Behav Ecol 3(2):156–165. https://doi.org/10.1093/beheco/3.2.156
Schmidt MH, Rocker S, Hanafi J, Gigon A (2008) Rotational fallows as overwintering habitat for grassland arthropods: the case of spiders in fen meadows. Biodivers Conserv 17(12):3003–3012. https://doi.org/10.1007/s10531-008-9412-6
Sevenster JG, Ellers J, Driessen G (1998) An evolutionary argument for time limitation. Evolution 52(4):1241–1244. https://doi.org/10.2307/2411256
Shuker DM, Pen I, Duncan AB, Reece SE, West SA (2005) Sex ratios under asymmetrical local mate competition: theory and a test with parasitoid wasps. Am Nat 166(3):301–316. https://doi.org/10.1086/432562
Silva-Torres CSA, Ramos Filho IT, Torres JB, Barros R (2009) Superparasitism and host size effects in Oomyzus sokolowskii, a parasitoid of diamondback moth. Entomol Exp Appl 133(1):65–73. https://doi.org/10.1111/j.1570-7458.2009.00903.x
Snyder WE, Evans EW (2006) Ecological effects of invasive arthropod generalist predators. Annu Rev Ecol Evol Syst 37(1):95–122. https://doi.org/10.1146/annurev.ecolsys.37.091305.110107
Strand MR, Obrycki JJ (1996) Host specificity of insect parasitoids and predators. Bioscience 46(6):422–429. https://doi.org/10.2307/1312876
Takagi M (1987) The reproductive strategy of the gregarious parasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae). Oecologia 71(3):321–324. https://doi.org/10.1007/BF00378702
Triana E, Barrantes G, Hanson P (2012) Incidence of parasitoids and predators on eggs of seven species of Therididae (Araneae). Arachnology 15(9):293–298. https://doi.org/10.13156/arac.2012.15.9.293
Van Alphen JJM, Visser ME (1990) Superparasitism as an adaptive strategy for insect parasitoids. Annu Rev Entomol 35(1):59–79. https://doi.org/10.1146/annurev.en.35.010190.000423
van Baarlen P, Topping CJ, Sunderland KD (1996) Host location by Gelis festinans, an eggsac parasitoid of the linyphiid spider Erigone atra. Entomol Exp Appl 81(2):155–163. https://doi.org/10.1111/j.1570-7458.1996.tb02027.x
Vetter RS, Vincent LS, Danielsen DWR, Reinker KI, Clarke DE, Itnyre AA, Kabashima JN, Rust MK (2012) The prevalence of brown widow and black widow spiders (Araneae: Theridiidae) in urban southern California. J Med Entomol 49(4):947–951. https://doi.org/10.1603/me11285
Vinson SB, Iwantsch GF (1980) Host suitability for insect parasitoids. Annu Rev Entomol 25(1):397–419. https://doi.org/10.1146/annurev.en.25.010180.002145
Wajnberg E, Boulétreau M, Prévost G, Fouillet P (1990) Developmental relationships between Drosophila larvae and their endoparasitoid Leptopilina (Hymenoptera: Cynipidae) as affected by crowding. Arch Insect Biochem Physiol 13(3–4):239–245. https://doi.org/10.1002/arch.940130310
Werren JH (1980) Sex ratio adaptations to local mate competition in a parasitic wasp. Science 208(4448):1157–1159. https://doi.org/10.1126/science.208.4448.1157
Werren JH (1983) Sex ratio evolution under local mate competition in a parasitic wasp. Evolution 37(1):116–124. https://doi.org/10.2307/2408180
West SA, Flanagan KE, Godfray HCJ (2001) Variable host quality, life-history invariants, and the reproductive strategy of a parasitoid wasp that produces single sex clutches. Behav Ecol 12(5):577–583. https://doi.org/10.1093/beheco/12.5.577
White TA, Perkins SE (2012) The ecoimmunology of invasive species. Funct Ecol 26(6):1313–1323. https://doi.org/10.1111/1365-2435.12012
Zaviezo T, Mills N (2000) Factors influencing the evolution of clutch size in a gregarious insect parasitoid. J Anim Ecol 69(6):1047–1057. https://doi.org/10.1111/j.1365-2656.2000.00460.x
Acknowledgements
We thank Ishai Hoffmann and Ofir Altstein for their assistance collecting wasps and spiders from the field. Thanks to Maayan Segoli and Denis Vostrikov for their help counting and measuring parasitoids.
Funding
This work was supported by a Zuckerman STEM Leadership Postdoctoral Fellowship to MAM. MS acknowledges support from the Israel Institute for Advanced Studies.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Daniel, J.A., Arabesky, V., Rozenberg, T. et al. Parasitoid development and superparasitism in invasive versus native widow spider host egg sacs. Biol Invasions 25, 2519–2530 (2023). https://doi.org/10.1007/s10530-023-03052-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-023-03052-0