
Abstract
Accurately characterizing the provenance and genetic diversity of non-native populations aids the management of biotic invasions because population-level genetic diversity is potentially correlated with invasive success. We sequenced the mitochondrial DNA of non-native Phelsuma grandis Gray 1870 (Cyt-b, 610 bp) and Gekko gecko (Linnaeus 1758) (ND2, 751 bp) specimens collected from southern Florida, and compared these sequences to those of georeferenced native-range conspecifics. Multiple mtDNA clades were identified in southern Florida populations of both species. In the case of G. gecko these were traceable to geographically distinct native-range locales, confirming that southern Florida’s population ultimately derives from multiple native-range regions. The majority of the P. grandis sequenced were closely allied with a specimen from the far north of Madagascar, while a minority clustered in a well-supported clade with P. grandis derived from northeastern Madagascar. Sympatry of individuals belonging to multiple mtDNA clades was confirmed for both species, and uncorrected pairwise distances as high as 11.41% were detected in sympatric G. gecko, highlighting the potential for interpopulation—and perhaps even interspecific—diversity to be transformed into intrapopulation diversity during invasion events. Our findings provide further evidence that introduced squamate populations are frequently highly heterogeneous and derived from multiple, distinct native-range lineages.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Determining the provenance, identity, and diversity of non-native organisms and populations is an important first step in characterizing any biotic invasion (Collins et al. 2002). The study of the provenance of non-native taxa is of great importance for its power to reveal whether non-native populations comprise individuals descended from multiple, genetically distinct lineages. Thus phenomenon serves to transform interpopulation diversity—and sometimes even overlooked or cryptic interspecific diversity (e.g., Hunter et al. 2018)—into intrapopulation diversity within the non-native range, resulting in elevated levels of genetic diversity and thereby potentially facilitating the adaptation, establishment, spread, and persistence of non-native organisms (Crawford and Whitney 2010; Gillis et al. 2009; Kolbe et al. 2004, 2007; Lavergne and Molofsky 2007; Smyser et al. 2020; Wagner et al. 2017). Furthermore, traits pertinent to invasive success often show spatial variation, including behaviors (Lycett et al. 2009), bioclimatic tolerances (Kolbe et al. 2013a), parasite assemblages (Kmentová et al. 2019), resistance to disease (Perrin et al. 2010), and degree of genetic admixture (Wielstra et al. 2017). Finally, provenance analysis can be used to pinpoint sources of introductions, as exemplified by the case of accidental introductions of organisms via cargo in ports accepting shipments from various locales (Júnior 2015; Kraus 2008; Krysko et al. 2016; Nania et al. 2020).
The state of Florida, USA is home to more non-native reptile and amphibian species than anywhere else on Earth (Krysko et al. 2016). Several studies have been conducted to establish the provenance, identity, and diversity of non-native squamate reptiles in the state. Kolbe et al. (2004) found evidence of at least eight separate introductions of Anolis sagrei to Florida, and Kolbe et al. (2007) conducted similar research on seven additional non-native Florida Anolis species, identifying mitochondrial (mtDNA) haplotypes derived from multiple native-range areas in all but one case. In a study of introduced anoles in Florida, the authors noted that the mean pairwise sequence divergences among haplotypes in introduced populations exceeded those of native populations in seven of Florida’s eight (at that time) non-native Anolis species, and speculated that highly heterogeneous populations derived from multiple native-range areas may be the norm in this group (Kolbe et al. 2007). More recently, Nuñez (2016) documented the occurrence of multiple introductions of both Ctenosaura similis and Agama picticauda to Florida. Hunter et al. (2018) revealed that Florida’s purported Python bivittatus population also comprises P. molurus as well as P. bivittatus × P. molurus hybrids, and thus clearly derives from multiple founding regions. Dowell et al. (2016) showed that the three extant Florida Varanus niloticus populations derive from three distinct regions of West Africa, but are apparently spatially and genetically isolated from one another. Similar studies did not yield evidence of multiple source populations for Florida’s Pituophis ruthveni (Krysko et al. 2014), Hemidactylus garnotii, or H. mabouia (Carranza and Arnold 2006) populations, but in each case only two specimens were assayed for a mitochondrial gene. Non-native squamate populations comprising multiple native-range sources have also been reported from other locales, such as Hemidactylus spp. in the Maldives (Agarwal et al. 2019), Podarcis muralis in England (Michaelides et al. 2013), and Anolis carolinensis on the Pacific Islands (Michaelides et al. 2018), confirming that this phenomenon is not unique to Florida. Over 85% of amphibian and reptile introductions to Florida are associated with the pet trade pathway (Krysko et al. 2016). Taxa introduced via this pathway might be especially likely to derive from multiple, distinct sources, given that novelty is often a driver in the pet trade (e.g., Nekaris and Bergin 2016; Nijman et al. 2019), and that this desire for novelty may drive the availability—and ultimately the escape or release—of multiple varieties or morphs of a given taxon (Kolbe et al. 2013b).
The Madagascar giant day gecko Phelsuma grandis Gray 1870 and the tokay gecko Gekko gecko (Linnaeus 1758) are two of the 16 non-native gecko species currently established in Florida (Krysko et al. 2019). Phelsuma grandis is native to northern Madagascar (Sanchez and Probst 2014) and was first recorded in Florida in the 1990s (Bartlett and Bartlett 1999). The species is currently confined to southern Florida, where it is established in Miami (Thawley and Stroud 2017) and Homestead (Fieldsend and Krysko 2019a) in Miami-Dade County, and on at least 14 of the Florida Keys (Fieldsend and Krysko 2019b), Monroe County. Gekko gecko has a wide native range spanning from Nepal to the Aru Islands, Indonesia (Rösler et al. 2011). This species was first reported in Florida in the 1960s (King and Krakauer 1966), and has since been recorded from Leon county in the Panhandle (Means 1996) southward to Key West, Monroe County (Meshaka et al. 2004). Both species were introduced to southern Florida via the pet trade (Krysko et al. 2016), and continue to be intentionally released in the State (Krysko et al. 2019). In the case of P. grandis, breeding populations are undoubtedly established with the intention of harvesting for the pet trade (Krysko et al. 2019); however, data are not available on the number of P. grandis and G. gecko in the pet trade that are imported versus captured domestically.
Both P. grandis and G. gecko pose a potential threat to Florida’s native biodiversity by virtue of being generalist predators (Dervin et al. 2013; Krysko et al. 2019; Meshaka Jr et al. 1997; Sanchez and Probst 2014). Both species consume gastropods (Meshaka Jr et al. 1997; Sanchez and Probst 2014), which is concerning given that Florida is home to native tree snails of the genera Liguus, Orthalicus, and Drymaeus (Kay 1995). Gekko gecko is particularly voracious, and has been documented feeding on a wide range of vertebrates including frogs, lizards, snakes, and birds (Krysko et al. 2019); it has also been observed preying on rodents in its native range (Bucol and Alcala 2013) and bats in its introduced range (Breuil et al. 2009), implying a threat to the threatened and endemic Florida bonneted bat Eumops floridanus, Key Largo woodrat Neotoma floridana smalli, Key Largo cotton mouse Peromyscus gossypinus allapaticola, and silver rice rat Oryzomys palustris natator, all of which apparently occur in sympatry with G. gecko (Beckmann 2011; Crouse 2007; de Torrez et al. 2018; Krysko and Daniels 2005; Krysko et al. 2019; Potts 2011).
The primary aim of this study was to establish the provenance and genetic diversity of southern Florida’s P. grandis and G. gecko populations. In particular, we were interested in determining whether these non-native squamate populations show evidence of genetic contributions from multiple, genetically distinct native-range lineages. This phenomenon has been reported for several other non-native squamates in Florida, thus potentially challenging the classical view that non-native populations are genetically homogenous and depauperate compared to native-range populations (Kolbe et al. 2007). Species in the family Gekkonidae have been shown to generally exhibit high levels of intraspecific mtDNA variation, making these geckos excellent candidates for studies in invasion genetics (Barrett et al. 2016; Harris 2002).
A secondary aim of this research was to confirm that these non-native gekkonids have been correctly identified. Establishing the identity of non-native taxa is not always straightforward, and non-native organisms are sometimes initially misidentified (e.g., Hunter et al. 2018; Nuñez et al. 2016), which can be problematic since the life histories of morphologically similar taxa can differ substantially (Collins et al. 2002; Hunter et al. 2018; Raxworthy et al. 2007; Zhang et al. 2014). Phelsuma grandis was elevated from its subspecific classification as P. madagascariensis grandis to specific rank by Raxworthy et al. (2007) along with P. kochi Mertens 1954 and P. madagascariensis Gray 1831, which were previously classified as P. m. kochi and P. m. madagascariensis, respectively. Two subspecies of the red-spotted tokay G. gecko are currently recognized: G. g. azhari, which is endemic to Bangladesh (Mahony and Reza 2008; Mertens 1955; Rösler et al. 2011), and the nominate taxon G. g. gecko, which occurs throughout the rest of the native range (Rösler et al. 2011) and is the only G. gecko ssp. identified in Florida thus far (Krysko et al. 2019). A second species of tokay gecko sensu lato, the “black-spotted tokay” G. reevesii (Gray 1831), was revalidated by Rösler et al. (2011), having long been treated as a distinctive morph of G. gecko (e.g., Peng et al. 2011). Importantly, neither P. grandis nor G. gecko was recognized by the scientific community as a distinct species from its morphologically similar congeners at the time of its original introduction to Florida. Consequently, it is entirely possible that P. kochi, P. madagascariensis, and G. reevesii were legitimately imported into the United States under the listing P. madagascariensis or G. gecko, and subsequently introduced to Florida. For example, “Chinese tokay geckos”—listed as G. gecko—were availalable for import from Hong Kong during the 1980s (Murphy and McCloud 2010), but G. reevesii is now known to occur in both Hong Kong (Chan et al. 2006) and mainland China (Rösler et al. 2011). Phelsuma grandis and G. gecko display very little overlap in bioclimatic niche with these congeners (Raxworthy et al. 2007; Rösler et al. 2011; Zhang et al. 2014), and so could differ markedly in their ability to colonize and spread through Florida, highlighting the importance of ensuring that these non-native gekkonids are accurately identified.
Methods
Thirty-four Gekko sp. and 27 Phelsuma sp. were captured via hand, noose pole, baited hook (Krysko 2000), or the fishing/glue trap technique (Fieldsend and Krysko 2020) from various sites in southern Florida (Fig. 1) between 24 May 2018 and 31 July 2020. Upon capture, lizards were euthanized via intracoelomic injection of MS222 (tricaine methanesulfonate) following Conroy et al. (2009), transported on dry ice, and stored at − 80 °C under Florida International University IACUC protocol # IACUC-17-019. Additionally, 12 Phelsuma sp. tissue samples were taken from specimens held in collections of the Florida Museum of Natural History, resulting in a total P. grandis sample size of 39. Full details of the specimens used in this study are given in Online Resource 1.

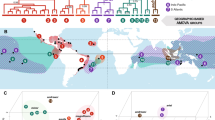
Locations and genetic affiliations of southern Florida Phelsuma grandis (a) and Gekko gecko (b) populations included in this study. Embedded mtDNA haplotype networks were generated using 610 bp of Cyt-b (P. grandis) and 751 bp of ND2 (G. gecko). Haplotype network circle size corresponds to haplotype frequency (n = 1–27). Black dots represent the number of mutations separating haplotypes, and numbers in parentheses denote the number of mutations separating major mtDNA clades/subclades from one another. Map circle sizes correspond to population sample sizes (n = 1–8), with circle segment coloration representing the proportion of individuals from the population found to either possess a given haplotype (P. grandis) or belong to a given haplogroup (G. gecko). Site abbreviations are as follows: BCK – Big Coppitt Key; BPK—Big Pine Key; CK—Cudjoe Key; FDK—Fat Deer Key; GK—Grassy Key; HS—Homestead; KL—Key Largo; KW—Key West; LTK—Little Torch Key; MI—Miami; NKL—North Key Largo; PC—Pinecrest; PK—Plantation Key; RK—Ramrod Key; SBK—Saddlebunch Keys; SK—Summerland Key; SM—South Miami; TV—Tavernier; UMK—Upper Matecumbe Key; VK—Vaca Key
DNA was extracted from tissue samples using cetyltrimethylammonium bromide (CTAB) as per the protocol of Saghai-Maroof et al. (1984). The cytochrome b (Cyt-b) and NADH dehydrogenase 2 (ND2) mitochondrial genes were chosen for analysis of Phelsuma and Gekko respectively, in order to maximize the number of native-range sequences available for comparison on GenBank. Double-stranded DNA amplification for Phelsuma samples was achieved via Polymerase Chain Reaction (PCR) using an initial hot-start step of 10 m at 57 °C, followed by 37 cycles of 30 s of 94 °C denaturation/45 s of 57 °C annealing/45 s of 72 °C extension, followed by 60 s at 49 °C, followed by a final 15 m extension period at 72 °C. The PCR parameters for Gekko samples were as follows: 10 m at 57 °C, followed by 37 or 39 cycles of 30 s of 94 °C denaturation/45 s of either 53 °C or 57 °C annealing/45 s of 72 °C extension, followed by 60 s at 49 °C, followed by a final 15 m extension period at 72 °C. PCRs were performed in 50 μl reactions containing 1-3 μl of total DNA extract of variable concentration (average ~ 31 ng/μl), 27.5–29.5 μl ddH2O, 10 μl of 5X Green GoTaq® Flexi Buffer (Promega, Madison, WI, USA), 3 μl of MgCl2 (25 mM), 2.5 μl each of 10 μM forward and reverse primer (Online Resource 2), 1 μl of dNTP solution (10 mM with respect to each dNTP), and 0.5 μl Taq polymerase (5U/μl). Double-stranded PCR products were then cleaned with ExoSap-IT (Thermo Fisher Scientific, Waltham, MA, USA), and cycle sequencing was performed using BigDye™ Terminator v3.1 Cycle Sequencing Kit as per the manufacturer’s instructions, but using 1/2 the reaction volume and 1/8 the amount of BigDye Terminator v3.1 (Thermo Fisher Scientific, Waltham, MA, USA). Sequences were read on both strands using an Applied Biosystems 3130XL Genetic Analyzer.
Sequence alignments comprised a combination of sequences generated for this study and sequences retrieved from GenBank (Online Resources 3 and 4). When multiple identical sequences were available on GenBank for a native-range locale, we included a single representative sequence in our alignment in order to reduce redundancy. Sequence alignments were conducted with MAFFT (Katoh and Standley 2013) in Mesquite (Maddison and Maddison 2019) and produced alignments with no internal gaps. Alignments were translated to amino acids with the vertebrate mitochondrial genetic code to verify in-frame reading through the sequences with no stop codons.
Phylogenetic analyses were undertaken using Bayesian inference as employed in MrBayes (v3.2.6, Ronquist et al. 2012) using the GTR + I + G model on each of three partitions by codon position, allowing parameters to vary independently for each partition, which differ in patterns of base compositional bias, selective constraints, and proportion of invariant sites. Default priors were used in all analyses. Posterior probabilities of phylogenetic trees were based on running 10 million generations of Metropolis coupled Markov Chain Monte Carlo (MC3), which included two simultaneous runs of four chains, starting with a random tree and sampling every 100 generations. Preliminary runs were carried out to ensure stationarity of the dataset, with the potential scale reduction factor approaching 1 for all parameters, and the standard deviation of split frequencies less than 0.01. Twenty-five percent of trees were discarded as burnin; the remaining trees were used to determine the 50% majority-rule consensus tree and estimate Bayesian posterior probabilities.
Haplotype networks (Fig. 1) were generated in R version 3.5.3 (R Core Team 2019) using the haploNet function in the ‘pegas’ package (Paradis 2010).
Results
The primers used for Phelsuma grandis amplified a mean product of 651 base pairs (bp) of the Cyt-b mitochondrial gene, 610 bp of which were used for analysis. The equivalent values for the Gekko gecko ND2 mitochondrial gene were 1262 bp and 751 bp respectively, the latter value corresponding to the length of the majority of GenBank sequences available for comparison (Saijuntha et al. 2019). All sequences were deposited into GenBank (Online Resource 1).
All 39 Phelsuma sp. mtDNA haplotypes from southern Florida were reliably assigned to P. grandis, rather than its morphologically similar congeners P. kochi and P. madagascariensis (Fig. 2). Five distinct mitochondrial haplotypes were identified in the southern Florida P. grandis populations (Figs. 1, 2). Haplotype I was present in 27 of the 39 sequenced specimens (69.2%), and was the only haplotype identified in the Key Largo, Fat Deer Key, Vaca Key, Ramrod Key, Big Coppitt Key, and Key West populations. This haplotype was also detected on Grassy Key (1 of 4 specimens), Big Pine Key (1 of 2 specimens), and Little Torch Key (7 of 8 specimens). Haplotype II was confined to Plantation Key, where it was the sole haplotype detected in the four sequenced specimens. Haplotype III was present on Tavernier (1 of 1 specimens) and Little Torch Key (1 of 8 specimens), whilst Haplotype IV was detected in Miami (1 of 1 specimens) and Homestead (1 of 1 specimens), and on Grassy Key (1 of 4 specimens). Haplotype V was found on Grassy Key (2 of 4 specimens) and Big Pine Key (1 of 2 specimens). Thirty-six of the 39 southern Florida P. grandis did not cluster with any native-range sequences in our phylogenetic analysis, but were found to be very closely allied (606–609 out of 610 bp identical, 99.34%–99.84% similarity) with a specimen from Antsiranana, Madagascar, which is located at the northern extreme of the species’ native range. The remaining three sequences clustered in a well-supported clade containing native-range specimens from Analalava Forest, Andilana, Cap Est, and Salafaina. All four of these sites are located on Madagascar’s northeastern coast, with Cap Est located on the Masoala peninsula at the southern edge of the species’ native distribution, some 351 km south-east of Antsiranana (Raxworthy et al. 2007, Supplementary data).

Phelsuma 50% majority-rule consensus tree (Cyt-b, 610 bp). The year, if given, denotes the year in which the specimen was captured. Asterisks (*) denote nodes with Bayesian posterior probabilities ≥ 0.95. Haplotype (I-V) classifications follow Fig. 1. GenBank Accession Numbers are given for sequences that were not generated as part of this study. Countries of origin are identified by their ISO 3166-1 alpha-3 codes: Madagascar—MDG; United States of America—USA. AY221380 and AY221381 are captive specimens from Mauritius and the United Kingdom respectively
No Gekko sp. ND2 haplotypes from southern Florida could reliably be assigned to G. reevesii. Indeed, our analysis found G. reevesii to be both polyphyletic and deeply nested within G. g. gecko, a finding corroborated by previous mtDNA studies (Saijuntha et al. 2019; Wang et al. 2013). The subspecies G. g. azhari was not included in this analysis; however, the high support for the placement of the southern Florida Gekko specimens within well-characterized native-range G. g. gecko clades apparently spatially disjunct from G. g. azhari’s known range in Bangladesh (Mahony and Reza 2008; Mertens 1955; Rösler et al. 2011; Saijuntha et al. 2019) makes it likely that no G. g. azhari haplotypes were detected. Saijuntha et al. (2019) demonstrated the existence of five, distinct native-range G. gecko mtDNA clades (Clades A-E), which between them contain 10 discrete “haplogroups” (G1-G10) (Fig. 3). A total of 12 mitochondrial haplotypes were detected in the 34 assayed southern Florida G. gecko specimens (Figs. 1, 3), all of which clustered within two of the five native-range clades, with three of the 10 haplogroups represented. Eight of the southern Florida G. gecko (23.5%)—including specimens from Homestead, Key Largo, and Plantation Key—clustered within Clade A, Haplogroup G3 from Cambodia, South China, peninsular Malaysia, Sumatra (Indonesia), northeastern Thailand, Timor-Leste, and northern Vietnam. The remaining 26 southern Florida G. gecko clustered within two well-supported subclades of native-range Clade B. Nine (26.5%)—including individuals from South Miami, Key Largo, Plantation Key, Summerland Key, and Cudjoe Key—placed within the Clade B, Haplogroup G6 subclade, which contains native-range specimens from peninsular Thailand. The remaining 17 specimens (50%) from Pinecrest, North Key Largo, Key Largo, Plantation Key, Upper Matecumbe Key, Big Pine Key, Summerland Key, and Cudjoe Key fell within Clade B, Haplogroup G1, which includes native-range specimens from Cambodia, Laos, and northeastern Thailand, as well as two specimens identified by Wang et al. (2013) as G. reevesii.

Gekko 50% majority-rule consensus tree (ND2, 751 bp). Asterisks (*) denote nodes with Bayesian posterior probabilities ≥ 0.95. Clade, subclade, and haplogroup (G1–G10) classifications follow Saijuntha et al. (2019) with some modification. Clades C, D, and E (Saijuntha et al. 2019) have been collapsed to better visualize the clades containing sequences from southern Florida G. gecko. GenBank Accession Numbers are given for sequences which were not generated as part of this study. Countries of origin are identified by their ISO 3166-1 alpha-3 codes: Cambodia—KHM; China—CHN; Indonesia—IDN; Laos—LAO; Malaysia—MYS; Myanmar—MMR; Thailand—THA; Timor-Leste—TLS; United States of America—USA; Vietnam—VNM
Mean uncorrected pairwise distance for all southern Florida P. grandis (n = 39) was 0.60%, while the equivalent value for G. gecko (n = 34) was 5.39%. Maximum uncorrected pairwise distances were 2.00% for P. grandis and 11.77% for G. gecko (Online Resources 3 and 4).
Discussion
This study identifies the presence of multiple, distinct mitochondrial clades in southern Florida’s non-native Phelsuma grandis and Gekko gecko populations. In doing so, it provides further evidence that genetically heterogeneous populations (Hunter et al. 2018; Kolbe et al. 2004, 2007; Nuñez 2016) may be the rule rather than the exception in the non-native squamate fauna of Florida. Similar research conducted outside of Florida (e.g., Agarwal et al. 2019; Michaelides et al. 2013, 2018) indicates that this may be a general phenomenon in non-native squamates, in contrast to the general pattern of reduced genetic diversity exhibited in introduced populations of most taxa (Dlugosch and Parker, 2008). While some of Florida’s highly diverse non-native squamate populations clearly derive from accidental importation via cargo (e.g., Anolis sagrei, Kolbe et al. 2004; Krysko et al. 2016), > 85% of introductions of reptiles and amphibians to the state can ultimately be traced to the pet trade (Krysko et al. 2016). The relationship between invasion pathway and genetic diversity should thus be considered a pressing issue within the field of invasion biology, given that high-or-heightened population-level genetic diversity has now been implicated as a driver of invasive success in several case studies (Crawford and Whitney 2010; Lavergne and Molofsky 2007; Smyser et al. 2020; Wagner et al. 2017).
The uncorrected mean pairwise distance value for southern Florida G. gecko (5.39%) was found to be much higher than the equivalent value (0.60%) for P. grandis. An important consideration is that different mitochondrial genes were assayed for each species: analysis of P. grandis was conducted using the Cyt-b gene, while ND2 was used for analysis of G. gecko. These pairwise distance values thus relate to different regions of the mitochondrial genome, across which substitution rates can differ substantially (Pesole et al. 1999), and care must therefore be taken when drawing inferences. To check that this discrepancy was not merely an artifact of differing substitution rates between these two mitochondrial genes, we first downloaded 12 Gekko spp. complete mitochondrial genomes from GenBank (Hao et al. 2016a, 2016b; Kim et al. 2016; Kumazawa 2007; Li et al. 2013; Zhou et al. 2006). We then calculated the mean uncorrected pairwise distance at both the 751 bp ND2 region used in our analysis of G. gecko, and the 610 bp Cyt-b region equivalent to that used for analysis of P. grandis. We found that the ND2 region was only slightly more variable than the Cyt-b region (mean uncorrected pairwise distances 28.94% vs 25.71%, Online Resource 5). We were unable to perform an equivalent analysis with Phelsuma spp. due to a lack of available GenBank data. Nevertheless, given that the mean uncorrected pairwise distance for southern Florida’s G. gecko population was found to be nearly nine times greater than the equivalent for P. grandis, analysis of different mitochondrial regions for the two species seems unlikely to be the major cause of this difference. It should however be noted that the two species are rather distantly related to one another within the Gekkonidae (Gamble et al. 2012), and differ substantially in various aspects of their natural history (e.g., size and behavior, Krysko et al. 2019).
Phylogenetic analysis (Fig. 2) revealed the existence of multiple P. grandis mtDNA clades in southern Florida, with northeastern Madagascar identified as the probable source of Haplotype V. Haplotypes I-IV proved to be more similar to that of a specimen from the city of Antsiranana in Madagascar’s far north, which would be consistent with its position at the center of the P. grandis trade (CJ Raxworthy, pers. comm.); nevertheless, the paucity of georeferenced native-range sequences precludes confident inference of the provenance of these four haplotypes. Interestingly however, two individuals from Big Pine Key—captured on the same day from adjacent buildings (Online Resource 1)—were found to possess Haplotypes I and V respectively (Figs. 1, 2), implying the possibility of admixture between these distinct clades. Given the small sample sizes in our study, deeper genetic sampling is required in order to address population-level questions. This is particularly true of the Big Pine Key, Little Torch Key, Grassy Key, and Plantation Key populations, as these represent the oldest extant P. grandis populations in southern Florida (Krysko et al. 2003). As such, knowledge of their genetic composition would likely shed light on the colonization history of P. grandis in southern Florida since its initial introduction in the 1990s (Bartlett and Bartlett 1999). Deeper sampling would also help to confirm whether Haplotype II is a private haplotype of the Plantation Key Population, and whether the low prevalence of Haplotype I in the Upper Keys and mainland (2 of 9 specimens) versus the Lower Keys (25 of 30 specimens) (χ2 (1, N = 39) = 12.14, p ≤ 0.0005) is indicative of genuine population structure, or merely an artifact of small sample sizes and/or sampling bias.
Our study revealed the presence of at least three, deeply divergent mtDNA lineages in southern Florida’s G. gecko population (Figs. 1, 3). Kongbuntad et al. (2016) postulate that G. g. gecko may actually represent a species complex. The maximum ND2 uncorrected pairwise distance of 18.44% detected between a G. g. gecko specimen from Ayeyarwady, Myanmar (GenBank Accession No. JN019052) and two G. g. gecko from northern Thailand (GenBank Accession Nos. MK117114 and MK117200) (Online Resource 3; Rösler et al. 2011; Saijuntha et al. 2019) provides some support for this this assertion, given that it exceeds the greatest ND2 uncorrected pairwise distance (18.2%) reported by Agarwal et al. (2019) in their study of the unresolved “Hemidactylus frenatus” gekkonid species complex. Nevertheless, this maximum uncorrected pairwise distance still falls short of the analogous Gekko interspecific uncorrected pairwise distances detailed in Online Resource 5, which range from 23.06% (G. japonicus and G. swinhonis) to 41.21% (G. gecko and G. vittatus). It must however be noted that the analysis is by no means exhaustive, and includes only six of the ~ 45 known Gekko species (Rösler et al. 2011). Interestingly, all three pairwise mtDNA haplogroup combinations (i.e., G1/G3, G1/G6, and G3/G6) were found in sympatry in southern Florida (Fig. 1; Online Resource 1). Of particular note, two specimens collected from the same private residence in Key Largo were found to belong to the G3 and G6 haplogroups respectively, and exhibited an uncorrected pairwise distance of 11.41% (Online Resource 3). In keeping with contemporary taxonomic nomenclature, southern Florida’s tokay gecko population is currently classified as one subspecies, i.e., G. g. gecko (Krysko et al. 2019). Nevertheless, the depth of genetic divergence between the three mtDNA lineages—particularly between Haplogroup G3 (Clade A) and the other two haplogroups (Clade B)—suggests at least the possibility that multiple, reproductively isolated tokay gecko lineages are present in southern Florida. Intriguingly, Clade A and Clade B show close phylogeographical correspondence with the distribution of the morphologically distinct “nominal” and “central mainland” forms of tokay gecko described by Rösler (2005) respectively, providing further evidence of their distinctiveness. If admixture does indeed occur between these lineages, it appears likely that it is serving to transform interpopulation—and perhaps even interspecific—native-range diversity into intrapopulation diversity in southern Florida, potentially resulting in a highly genetically diverse population. Such admixture could conceivably increase invasive potential (Crawford and Whitney 2010; Lavergne and Molofsky 2007; Smyser et al. 2020; Wagner et al. 2017), although it should be noted that it could also have the opposite effect (e.g., outbreeding depression, Barker et al. 2019; Pantoja et al. 2018). Conversely, reproductive isolation between these lineages in southern Florida would provide very strong support for the argument that G. g. gecko is actually a species complex. This study system may thus present an unusual opportunity to use an introduction event to help resolve a species complex (e.g., Wegener et al. 2019). Since mitochondrial DNA is generally inherited uniparentally (Avise et al. 1979)—and is thus a poor indicator of hybridization and admixture—our future research will combine mtDNA data with nuclear markers such as microsatellites or nuclear genes (Hunter et al. 2018; Kurita et al. 2018; Pinto et al. 2019; Vuillaume et al. 2015), in order to check for cytonuclear discordance indicative of these phenomena (Toews and Brelsford 2012).
The work of Rösler et al. (2011), Wang et al. (2013), and Saijuntha et al. (2019) has made available for comparison ~180 georeferenced G. gecko ND2 sequences. Sequencing effort has thus far focused predominantly on Thailand, China, Vietnam, and neighboring countries, resulting in relatively high sampling density for China (excluding Hong Kong, Chan et al. 2006) and most of mainland Southeast Asia. Nevertheless, the wide native range of this species (Rösler et al. 2011) means that our phylogeographic knowledge of G. gecko remains incomplete, thereby lowering the confidence with which native-range provenance can be inferred. Gekko gecko ND2 sequence data are unavailable for Brunei (UMMZ 201955-201957), Philippines (Bucol and Alcala 2013), Singapore (Tan et al. 1978), and the Subcontinental nations of Bangladesh, Bhutan, India, and Nepal (Rösler et al. 2011), meaning that genetic sampling of specimens from these countries should be a top priority. Myanmar is another country for which sampling should be prioritized. Several gekkonid species exhibit deep intraspecific genetic divergence in Myanmar (Carranza and Arnold 2006), and the two Myanmar G. gecko specimens sequenced thus far have an ND2 uncorrected pairwise difference of 17.93% (Online Resource 3) despite having been collected from locations ca. 400 km apart (Rösler et al. 2011), suggesting that the nation could harbor considerable G. gecko diversity. Deeper sampling of Indonesia and Malaysia would also be welcomed, given their prominence in the international G. gecko trade (Caillabet 2013; Nijman and Shepherd 2015) and the low number of ND2 sequences from these countries presently available for comparative analysis. Another priority for future research is the taxonomic status of the black-spotted tokay, G. reevesii, as our analysis found specimens identified as G. reevesii to be both polyphyletic and clustered deep within G. g. gecko (Fig. 3), and thus does not support its current taxonomic status as a separate species.
Our study adds to the growing body of evidence that non-native squamate populations in Florida and elsewhere often ultimately derive from multiple, distinct genetic lineages, and are frequently highly heterogeneous as a result. Such heterogeneity can enhance the fitness (Smyser et al. 2020), adaptive potential (Lavergne and Molofsky 2007), and invasive success (Crawford and Whitney 2010) of non-native organisms, illustrating both the importance of this genetic approach to invasion biology, and the need to understand how widespread this phenomenon is in other taxonomic groups.
Availability of data and materials (data transparency)
The datasets generated and analyzed during the current study are available in the GenBank repository, under the Accession Numbers MT554153-MT554166 and MW244758-MW244816. Specimen voucher numbers are listed in Online Resource 1.
Code availability (software application or custom code)
N/A.
References
Agarwal I, Jablonski D, Bauer AM (2019) The identity and probable origin of the Hemidactylus geckos of the Maldives. Herpetol J 29:230–236
Avise JC, Giblin-Davidson C, Laerm J, Patton JC, Lansman RA (1979) Mitochondrial DNA clones and matriarchal phylogeny within and among geographic populations of the pocket gopher, Geomys pinetis. Proc Natl Acad Sci 76:6694–6698
Barker BS, Cocio JE, Anderson SR et al (2019) Potential limits to the benefits of admixture during biological invasion. Mol Ecol 28:100–113
Barrett SC, Colautti RI, Dlugosch KM, Rieseberg LH (2016) Invasion genetics: the Baker and Stebbins legacy. Wiley, New York
Bartlett RD, Bartlett PP (1999) field guide to Florida reptiles and amphibians. Gulf Pub. Co., Houston
Beckmann SM (2011) The effects of climatic and geographic events on the cotton mouse (Peromyscus gossypinus). University of Miami, Coral Gables
Breuil M, Guiougou F, Questel K, Ibéné B (2009) Modifications du peuplement les Antilles françaises, 2 ème partie Le Courrier de la Nature:36–43
Bucol A, Alcala A (2013) Tokay gecko, Gekko gecko (Sauria: Gekkonidae) predation on juvenile house rats. Herpetol Notes 6:307–308
Caillabet O (2013) The trade in tokay geckos in South-East Asia: with a case study on novel medicinal claims in Peninsular Malaysia A TRAFFIC Southeast Asia Report Selangor, Malaysia
Carranza S, Arnold E (2006) Systematics, biogeography, and evolution of Hemidactylus geckos (Reptilia: Gekkonidae) elucidated using mitochondrial DNA sequences. Mol Phylogenet Evol 38:531–545
Chan SK, Cheung K-S, Ho C-Y, Lam F-N, Tang W-S (2006) The Geckos of Hong Kong. Hong Kong Biodivers 13:1–9
Collins TM, Trexler JC, Nico LG, Rawlings TA (2002) Genetic diversity in a morphologically conservative invasive taxon: multiple introductions of swamp eels to the southeastern United States. Conserv Biol 16:1024–1035
Conroy C, Papenfuss T, Parker J, Hahn N (2009) Use of tricaine methanesulfonate (MS222) for euthanasia of reptiles. J Am Assoc Lab Anim Sci 48:28–32
Crawford K, Whitney K (2010) Population genetic diversity influences colonization success. Mol Ecol 19:1253–1263
Crouse AL (2007) Genetic analysis of the endangered silver rice rat (Oryzomys palustris natator) and Lower Keys marsh rabbit (Sylvilagus palustris hefneri). Texas A&M University, College Station
de Torrez ECB, Ober HK, McCleery RA (2018) Restoring historical fire regimes increases activity of endangered bats. Fire Ecol 14:9
Dervin S, Baret S, Penin L, Sanchez M (2013) Régime alimentaire du grand gecko vert de Madagascar, Phelsuma grandis Gray, 1870 sur l’île de La Réunion (Squamata: Gekkonidae). Cahiers scientifiques de l’océan Indien occidental 4:29–38
Dowell SA, Wood JP, Campbell TS, Kolokotronis S-O, Hekkala ER (2016) Combining genetic and distributional approaches to sourcing introduced species: a case study on the Nile monitor (Varanus niloticus) in Florida. R Soc Open Sci 3:150619
Dlugosch KM, Parker IM (2008) Founding events in species invasions: genetic variation, adaptive evolution, and the role of multiple introductions. Mol Ecol 17:431–449
Fieldsend TW, Krysko KL (2019a) Madagascar giant day gecko (Phelsuma grandis) established in Homestead Miami-Dade County, Florida, USA. IRCF Reptiles Amphib 26:159–160
Fieldsend TW, Krysko KL (2019b) PHELSUMA GRANDIS (Madagascar Giant Day Gecko) GEOGRAPHIC DISTRIBUTION. Herpetol Rev 50:527
Fieldsend TW, Krysko KL (2020) Collecting non-native Madagascar Giant Day Geckos, Phelsuma grandis (Gray 1870), in Florida: a novel approach combining the fishing and glue-trap techniques. IRCF Reptiles Amphib 27:114–115
Gamble T, Greenbaum E, Jackman TR, Russell AP, Bauer AM (2012) Repeated origin and loss of adhesive toepads in geckos. PLoS ONE 7:e39429
Gillis NK, Walters LJ, Fernandes FC, Hoffman EA (2009) Higher genetic diversity in introduced than in native populations of the mussel Mytella charruana: evidence of population admixture at introduction sites. Divers Distrib 15:784–795
Hao S, Ping J, Zhang Y (2016a) Complete mitochondrial genome of Gekko chinensis (Squamata, Gekkonidae). Mitochondrial DNA Part A 27:4226–4227
Hao S, Yu D, Ping J, Zhou H, Zhang Y (2016b) Complete mitochondrial genomes of two gecko species, Gekko hokouensis and Gekko japonicus (Squamata, Gekkonidae). Mitochondrial DNA Part B 1:346–347
Harris D (2002) Reassessment of comparative genetic distance in reptiles from the mitochondrial cytochrome b gene. Herpetol J 12:85–87
Hunter ME, Johnson NA, Smith BJ, Davis MC, Butterfield JSS, Snow RW, Hart KM (2018) Cytonuclear discordance in the Florida Everglades invasive Burmese python (Python bivittatus) population reveals possible hybridization with the Indian python (P. molurus). Ecol Evol 8:9034–9047. https://doi.org/10.1002/ece3.4423
Júnior JCR (2015) Occurrence of the Tokay Gecko, Gekko gecko Linnaeus 1758 (Squamata, Gekkonidae), an exotic species in southern Brazil. Herpetol Notes 8:8–10
Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 30:772–780
Kay EA (1995) The conservation biology of molluscs. In: Proceedings of a symposium held at the 9th international malacological congress, Edinburgh, Scotland, 1986. vol 9. IUCN
Kim I-H, Park J, Cheon K-S, Lee H-J, Kim J-K, Park D (2016) Complete mitochondrial genome of Schlegel’s Japanese gecko Gekko japonicus (Squamata: Gekkonidae). Mitochondrial DNA Part A 27:3684–3686
King W, Krakauer T (1966) The exotic herpetofauna of southeast Florida. Q J Florida Acad Sci 29:144–154
Kmentová N et al (2019) Co-introduction success of monogeneans infecting the fisheries target Limnothrissa miodon differs between two non-native areas: the potential of parasites as a tag for introduction pathway. Biol Invasions 21:757–773
Kolbe JJ, Ehrenberger JC, Moniz HA, Angilletta MJ Jr (2013a) Physiological variation among invasive populations of the brown anole (Anolis sagrei). Physiol Biochem Zool 87:92–104
Kolbe JJ, Glor RE, Schettino LR, Lara AC, Larson A, Losos JB (2004) Genetic variation increases during biological invasion by a Cuban lizard. Nature 431:177
Kolbe JJ, Glor RE, Schettino LR, Lara AC, Larson A, Losos JB (2007) Multiple sources, admixture, and genetic variation in introduced anolis lizard populations. Conserv Biol 21:1612–1625. https://doi.org/10.1111/j.1523-1739.2007.00826.x
Kolbe JJ, Lavin BR, Burke RL, Rugiero L, Capula M, Luiselli L (2013b) The desire for variety: Italian wall lizard (Podarcis siculus) populations introduced to the United States via the pet trade are derived from multiple native-range sources. Biol Invasions 15:775–783
Kongbuntad W et al (2016) Genetic diversity of the red-spotted tokay gecko (Gekko gecko Linnaeus, 1758) (Squamata: Gekkonidae) in Southeast Asia determined with multilocus enzyme electrophoresis. J Asia-Pacific Biodivers 9:63–68
Kraus F (2008) Alien reptiles and amphibians: a scientific compendium and analysis, vol 4. Springer, Berlin
Krysko K (2000) A fishing technique for collecting the introduced knight Anole (Anolis equestris) in Southern Florida. Caribb J Sci 36:162–162
Krysko K, Smith D, Rochford M, Kieckhefer G III, Nuñez L (2014) Confirmation of introduced Louisiana pinesnakes Pituophis ruthveni, in Florida based on molecular analyses. Amphib Reptile Conserv 8:36–43
Krysko K et al (2016) New verified nonindigenous amphibians and reptiles in Florida through 2015, with a summary of over 152 years of introductions. Reptiles Amphib 23:110–143
Krysko KL, Daniels KJ (2005) A key to the geckos (Sauria: Gekkonidae) of Florida. Caribb J Sci 41:28–36
Krysko KL, Enge KM, Moler PE (2019) Amphibians and reptiles of Florida. University Press of Florida, Gainesville
Krysko KL, Hooper AN, Sheehy CM III (2003) The Madagascar giant day gecko, Phelsuma madagascariensis grandis Gray 1870 (Sauria: Gekkonidae): a new established species in Florida. Florida Sci 66:222–225
Kumazawa Y (2007) Mitochondrial genomes from major lizard families suggest their phylogenetic relationships and ancient radiations. Gene 388:19–26
Kurita T, Honda M, Toda M (2018) Species delimitation and biogeography of the Ryukyu ground geckos, Goniurosaurus kuroiwae ssp. (Squamata: Eublepharidae), by use of mitochondrial and nuclear DNA analyses. J Zool Syst Evolut Res 56:209–222. https://doi.org/10.1111/jzs.12198
Lavergne S, Molofsky J (2007) Increased genetic variation and evolutionary potential drive the success of an invasive grass. Proc Natl Acad Sci 104:3883–3888
Li H-M, Zeng D-L, Guan Q-X, Qin P-S, Qin X-M (2013) Complete mitochondrial genome of Gekko swinhonis (Squamata, Gekkonidae). Mitochondrial DNA 24:86–88
Lycett SJ, Collard M, McGrew WC (2009) Cladistic analyses of behavioural variation in wild Pan troglodytes: exploring the chimpanzee culture hypothesis. J Hum Evol 57:337–349
Maddison W, Maddison D (2019) Mesquite: a modular system for evolutionary analysis. Version 3.51. 2018.
Mahony S, Reza A (2008) A herpetofaunal collection from the Chittagong Hill Tracts, Bangladesh, with two new species records for the country. Hamadryad 32:34–35
Means D (1996) Gekko gecko (tokay gecko). Herpetol Rev 27:152
Mertens R (1955) Über eine eigenartige Rasse des Tokehs (Gekko gecko) aus Ost-Pakistan. Senckenbergiana Biologica 36:21–24
Meshaka W Jr, Clouse R, McMahon L (1997) Diet of the tokay gecko (Gekko gecko) in southern Florida. Florida Field Nat 25:105–107
Meshaka WE, Butterfield BP, Hauge JB (2004) Exotic amphibians and reptiles of Florida. Krieger Pub. Co., Malabar
Michaelides S, While GM, Bell C, Uller T (2013) Human introductions create opportunities for intra-specific hybridization in an alien lizard. Biol Invasions 15:1101–1112
Michaelides SN, Goodman RM, Crombie RI, Kolbe JJ, Cowie R (2018) Independent introductions and sequential founder events shape genetic differentiation and diversity of the invasive green anole (Anolis carolinensis) on Pacific Islands. Divers Distrib 24:666–679. https://doi.org/10.1111/ddi.12704
Murphy JB, McCloud K (2010) Reptile dealers and their price lists. Herpetol Rev 41:266–281
Nania D, Flecks M, Rödder D (2020) Continuous expansion of the geographic range linked to realized niche expansion in the invasive Mourning gecko Lepidodactylus lugubris (Duméril & Bibron, 1836). PLoS ONE 15:e0235060
Nekaris K, Bergin D (2016) Primate trade (Asia). International Encyclopedia of Primatology, pp 1–8
Nijman V et al (2019) Dynamics of illegal wildlife trade in Indonesian markets over two decades, illustrated by trade in Sunda Leopard Cats. Biodiversity 20:27–40
Nijman V, Shepherd CR (2015) Adding up the numbers. TRAFFIC
Nuñez LP (2016) Molecular analyses of three non-indigenous squamate species in Florida: testing various hypotheses regarding species introductions. University of Florida
Nuñez LP, Krysko KL, Avery ML (2016) Confirmation of introduced Agama picticauda in Florida based on molecular analyses. Bull Florida Museum Nat Hist 54:138–146
Pantoja PO, Paine CT, Vallejo-Marín M (2018) Natural selection and outbreeding depression suggest adaptive differentiation in the invasive range of a clonal plant. Proc R Soc B Biol Sci 285:20181091
Paradis E (2010) pegas: an R package for population genetics with an integrated–modular approach. Bioinformatics 26:419–420
Peng Q, Wang G, Tan S, Wang T, Li L, Zou F (2011) Isolation and characterization of thirteen polymorphic tetranucleotide microsatellite markers in the Tokay gecko (Gekko Gecko). Conserv Genet Resour 3:617–619
Perrin P, Herbreteau V, Hugot J-P, Morand S (2010) Biogeography, humans, and their parasites. In: Morand S, Krasnov BR (eds) The biogeography of host–parasite interactions. Oxford University Press, Oxford, p 41
Pesole G, Gissi C, De Chirico A, Saccone C (1999) Nucleotide substitution rate of mammalian mitochondrial genomes. J Mol Evol 48(4):427–434
Pinto BJ, Titus-McQuillan J, Daza JD, Gamble T (2019) Persistence of a geographically-stable hybrid zone in Puerto Rican dwarf geckos. J Hered 110:523–534
Potts JM (2011) Estimating abundance of rare, small mammals: a case study of the Key Largo woodrat (Neotoma floridana smalli). University of St Andrews
Raxworthy CJ, Ingram CM, Rabibisoa N, Pearson RG (2007) Applications of ecological niche modeling for species delimitation: a review and empirical evaluation using day geckos (Phelsuma) from Madagascar. Syst Biol 56:907–923. https://doi.org/10.1080/10635150701775111
Rösler H (2005) Studien am Tokeh: 2. Intraspezifische Variation der südostasiatischen Populationen von Gekko gecko (Linnaeus, 1758). Gekkota, pp. 65–149
Rösler H, Bauer AM, Heinicke MP, Greenbaum E, Jackman T, Nguyen TQ, Ziegler T (2011) Phylogeny, taxonomy, and zoogeography of the genus Gekko Laurenti, 1768 with the revalidation of G. reevesii Gray, 1831 (Sauria: Gekkonidae). Zootaxa 2989:1–50
Ronquist F et al (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61:539–542
Saghai-Maroof MA, Soliman KM, Jorgensen RA, Allard R (1984) Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc Natl Acad Sci 81:8014–8018
Saijuntha W et al (2019) Genetic Structure of the Red-spotted Tokay Gecko, Gekko gecko (Linnaeus, 1758) (Squamata: Gekkonidae) from Mainland Southeast Asia Asian. Herpetol Res 10:69–78
Sanchez M, Probst J-M (2014) Distribution and habitat of the invasive giant day gecko Phelsuma grandis Gray 1870 (Sauria: Gekkonidae) in Reunion Island, and conservation implication. Phelsuma 22:13–28
Smyser TJ, Tabak MA, Robeson SB, Blackburn H (2020) Mixed ancestry from wild and domestic lineages contributes to the rapid expansion of invasive feral swine. Mol Ecol 29:1103–1119
Tan R, Lim E-W, Ishak B (1978) Intestinal bacterial flora of the household lizard, Gecko gecko. Res Vet Sci 24:262–263
Team RC (2019) R version 3.5.3: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Thawley CJ, Stroud JT (2017) Phelsuma grandis (Madagascan day gecko). Herpetol Rev 48:812–812
Toews DP, Brelsford A (2012) The biogeography of mitochondrial and nuclear discordance in animals. Mol Ecol 21:3907–3930
Vuillaume B, Valette V, Lepais O, Grandjean F, Breuil M (2015) Genetic evidence of hybridization between the endangered native species Iguana delicatissima and the invasive Iguana iguana (Reptilia, Iguanidae) in the Lesser Antilles: management implications. PLoS ONE 10:e0127575
Wagner NK, Ochocki BM, Crawford KM, Compagnoni A, Miller TE (2017) Genetic mixture of multiple source populations accelerates invasive range expansion. J Anim Ecol 86:21–34
Wang G et al (2013) Genetic variability of the tokay gecko based on mitochondrial and nuclear DNA. Mitochondrial DNA 24:518–527
Wegener JE, Pita-Aquino JN, Atutubo J et al (2019) Hybridization and rapid differentiation after secondary contact between the native green anole (Anolis carolinensis) and the introduced green anole (Anolis porcatus). Ecol Evolut 9:4138–4148
Wielstra B et al (2017) A genomic footprint of hybrid zone movement in crested newts. Evolut Lett 1:93–101. https://doi.org/10.1002/evl3.9
Zhang Y, Chen C, Li L, Zhao C, Chen W, Huang Y (2014) Insights from ecological niche modeling on the taxonomic distinction and niche differentiation between the black-spotted and red-spotted tokay geckoes (Gekko gecko). Ecol Evol 4:3383–3394. https://doi.org/10.1002/ece3.1183
Zhou K, Li H, Han D, Bauer AM, Feng J (2006) The complete mitochondrial genome of Gekko gecko (Reptilia: Gekkonidae) and support for the monophyly of Sauria including Amphisbaenia. Mol Phylogenet Evolut 40:887–892
Acknowledgements
The authors wish to thank the Miami-Dade County Environmentally Endangered Lands Program and Miami-Dade County Parks, Recreation, and Open Spaces Department, Natural Areas Management Division; Castellow Hammock Preserve & Nature Center; Camp Owaissa Bauer; Florida Park Service; Florida Department of Environmental Protection Division of Recreation and Parks; United States Department of the Interior; United States Fish and Wildlife Service; FIU Tropics; the Susan S. Levine Trust; the FIU Institute of Environment; Coleman M. Sheehy III and Terry A. Lott of the Florida Museum of Natural History; Shahrzad Forouzanfar, Madison Harman, Enrico Vezzani, Jim Duquesnel, the Jankowski family, and the public who generously assisted in various ways with the collection of specimens for this study. All work was performed under Florida International University IACUC protocol # IACUC-17-019.
Funding
This research was funded by Student Grants awarded to Thomas William Fieldsend by FIU Tropics, the FIU Institute of Environment, and the Susan S. Levine Trust.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Fieldsend, T.W., Krysko, K.L., Sharp, P. et al. Provenance and genetic diversity of the non-native geckos Phelsuma grandis Gray 1870 and Gekko gecko (Linnaeus 1758) in southern Florida, USA. Biol Invasions 23, 1649–1662 (2021). https://doi.org/10.1007/s10530-021-02463-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-021-02463-1