
Abstract
Non-native plant invasions are often catalyzed by wildfires and may alter the structure, function, and composition of ecosystems. Of particular concern are non-native annual grasses that promote increased fire frequency and compete with native vegetation for early season moisture. While these effects are well studied in non-forested ecosystems, relatively few studies document invasions of non-native annual grasses in forested ecosystems. We document non-native plant invasion following repeated high-severity wildfire in mixed-conifer/hardwood forests dominated by Douglas-fir (Pseudotsuga menziesii) in the Klamath Mountains of northern California. Despite great concern regarding the loss of conifer dominance and transitions to non-forested states related to recent and projected increases in wildfire activity, there is little known about invasions of non-native plant species following fire in this region. Non-native plant species capable of long-distance dispersal had an average total cover of 26% and were ubiquitous across a systematic, gridded sample of twenty 800 m2 plots. Non-native forbs included multiple species of common post-disturbance invaders (e.g. Cirsium vulgare, Senecio sylvaticus, Lactuca serriola). Non-native annual grasses comprised more of the vegetative cover than non-native forbs (17% vs. 9%), and were dominated by two species, Aira caryophyllea and Vulpia myuros, which reached as high as 80% cover. Our findings indicate that non-native annual grasses are a novel stressor that may accelerate projected loss of conifer dominance in the Klamath Mountains by facilitating fire spread and increasing competition for early season soil moisture. Additional monitoring will be crucial to understanding the impacts of non-native plants as fire activity in this region continues to increase.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Larger and more frequent wildfires are altering landscapes throughout forested regions of western North America (Westerling et al. 2006). Although fire is part of the natural disturbance regime across this region, there is great concern about uncharacteristically large patches of high-severity fire (Miller et al. 2012; Reilly et al. 2017) associated with increased stand density and fuel loading from a century of fire exclusion, coupled with longer fire seasons, more extreme fire weather days, and drier fuels from decreased summer precipitation (Jolly et al. 2015; Abatzoglou and Williams 2016; Holden et al. 2018). Plant communities with species that are adapted to a particular fire regime may experience a shift in species composition with alterations to the frequency and severity of fire. Although wildfire may offer opportunities for restoring some aspects of native plant communities, these disturbances also offer opportunities for invasion of non-native plant species (Keeley 2006) that threaten ecosystem resilience and resistance to future fires (Chambers et al. 2019).
Invasive plants have the potential to alter the structure, function, and composition of native ecosystems (D’Antonio and Vitousek 1992; D’Antonio et al. 2000; Brooks et al. 2004). Many non-native plant species pose a competitive threat to native vegetation, but one of the greatest threats that non-native species pose is through alteration of natural disturbance regimes (Mack and D’Antonio 1998). In particular, non-native annual grasses can increase the frequency of fire through rapid accumulation of fine fuels (D’Antonio and Vitousek 1992). Invasions of non-native annual grasses [e.g., cheatgrass (Bromus tectorum)] are well documented in non-forested ecosystems of the western United States (Mack 1981; Mack and Pyke 1983; D’Antonio and Vitousek 1992), but the few documented invasions of non-native annual grasses in tree dominated systems are limited to warm, dry ponderosa pine (Pinus ponderosa) woodlands (Pierson and Mack 1990; Crawford et al. 2001; Keeley and McGinnis 2007; Fornwalt et al. 2010; Kerns and Day 2017). More productive forested ecosystems with closed canopies where fire was excluded throughout the 20th century are considered more resistant to invasion (Keeley 2006; Klinger et al. 2008; Kerns et al. In Press).
The Klamath Mountains of northern California are renowned for their ecological diversity (Whittaker 1960). This region is primarily associated with a mixed-severity fire regime historically with pre-European settlement fire return intervals for Douglas-fir (Pseudotsuga menziesii) dominated mixed-conifer/hardwood forests that ranged from 12 to 19 years (Taylor and Skinner 2003). Fire exclusion effectively eliminated fire across much of the region during the 20th century, but fire activity has increased since the early 1980′s (Miller et al. 2012) and there is great concern about the conversion of old-growth forests to non-forested states following high-severity wildfire (Tepley et al. 2017; McCord et al. 2020). Although non-native plants are common post-fire invaders in other regions of California (Keeley 2006), these forests are thought to be relatively resistant to invasion following wildfire in the Klamath Mountains (Klinger et al. 2008). We document invasion of non-native plant species following short-interval (11 years) repeated high-severity wildfire in old-growth, low elevation mixed-conifer/hardwood forests dominated by Douglas-fir and discuss the implications of our findings on projected losses of conifer dominance in the region.
Methods
Study site
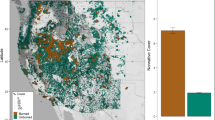
The Klamath Mountains of northern California and southwestern Oregon, USA (Fig. 1) are a hotspot biological diversity characterized by environmental and topographic complexity (Whittaker 1960). The region has a Mediterranean climate with warm, dry summers and cool, wet winters. Frequent (12–19 years) mixed severity fire drove the composition and structure of forests in the Klamath Mountains and maintaned landscape-scale patterns of low density, multi-aged stands during the pre-European settlement period (Taylor and Skinner 2003).

Plot locations with patterns of remotely-sensed burn severity from the relativized difference in the normalized burn ratio (RdNBR) in the Sims (2004) and Saddle Fire (2015). Inset in the top left shows the study location (green star) near Hyampom, California and perimeters of reburned areas from 1984 to 2017 (red) in the Klamath Mountains (dark gray)
Our study was located in the southcentral portion of the Klamath Mountains, northwest of Hyampom, California (Fig. 1). The area is composed of low elevation (500–1000 m) mixed-conifer and hardwood forests. Douglas-fir is the most common and dominant conifer, but other conifer species including ponderosa pine (Pinus ponderosa), Jeffrey pine (P. jeffreyi), sugar pine (P. lambertiana), and incense-cedar (Calocedrus decurrens) also occur occasionally. Evergreen and deciduous hardwoods are common in our study forest, especially tanoak (Notholithocarpus densiflorus), Pacific madrone (Arbutus menziesii), canyon live oak (Quercus chrysolepis), California black oak (Q. kelloggii), and Oregon white oak (Q. garryana). Common shrub species include wild lilac (Ceanothus spp.), manzanita (Arctostaphylos spp.), and poison oak (Toxicodendron diversilobum).
The Sims Fire was ignited by a fallen power line in July 2004 on an extreme fire weather day and burned 1630 ha during moderate drought conditions according to the Palmer Drought Severity Index (PDSI) (McCord et al. 2020). This landscape was highly departed from the historical fire regimes with no evidence of fire in the last 100 years. The fire included an approximately 700 ha patch of high- and very high-severity fire as measured by the relativized difference in the normalized burn ratio (RdNBR; Miller and Thode 2007) (RdNBR > 750) (Fig. 1). The Saddle Fire was ignited by lightning in June 2015 and burned 624 ha. Most of the Saddle Fire burned within the footprint of the previous Sims Fire, and a large portion of the reburned area occurred in an approximately 300 ha patch of high- and very-high burn severity.
Field methods
In 2017, two years following the Saddle fire, twenty 800 m2 (0.08 ha) circular plots were established on a 200 m grid system (Fig. 1). The plots were sampled according to an established regeneration plot protocol from the US Forest Service Common Stand Exam (USDA Forest Service 2007). We searched each plot and identified all plants to species. Plots were divided into quadrats and searched systematically at increasing distances from plot center. Once the entire plot was searched, the three person field crew came to a consensus and recorded ocular estimates of cover of each species across the entire plot. All species were characterized as native or non-native, then all non-native species were characterized by life form (annual/biennial/perennial) and dispersal capacity based on known species characteristics. Sources included the Manual of California Flora (Jepson Flora Project 2020), CalFlora (www.calflora.com), and the USDA plants database (https://plants.sc.egov.usda.gov).
Data analysis
In order to assess the effectiveness of our sample in capturing the potential pool of invasive species in the burned area, we used the specaccum function from the vegan package (Oksanen et al. 2009) in R Core Development Team (2018) to create species accumulation plots. Sites were added in random order with 50 permutations. Finally, we used the specpool function to provide an estimate of the total number of species in our study area. We then calculated the frequency of all species as the percent of all plots in which each species was found, as well as the percent of all vegetative cover composed of invasive plants. We also compared descriptive statistics of the abundance of each species, as well as taxonomic groups, functional groups, and dispersal mechanisms.
Results
Total cover of non-native plant species varied among plots but averaged 26.6% with a standard deviation (SD) of ± 25.9%, and ranged from 2% to 98.5%. Non-native plant species cover was > 40% in more than a quarter of the plots (Fig. 2). Across the study area, annual grasses made up more of the total non-native cover than forbs with a mean cover of 16.9% (SD = ±21.8%) and a range from 0.5% to 80%. Cover of annual forbs averaged 9.7% (SD = ±6.8%) and ranged from 0.5% to 23%.

Total cover of all non-native plants, non-native annual grasses, and non-native forbs in a systematic, gridded sample of twenty 800 m2 plots following repeated, short-interval high-severity fire. Boxplots represent the minimum and maximum values, the first and third quartiles, and the median value. Outliers are represented by open circles and means are represented by stars
Non-native plant species made up an average 17.7% (SD = ±10.4%) of the total vegetation cover and ranged from 3% to 34% (Fig. 3). Annual grasses made up an average of 10.2% (SD = ±10.2%) of the total vegetation cover and were < 5% of the total cover in almost half of the plots, but > 17% in a quarter of the plots. Annual forbs comprised an average of 7.5% (SD = ±4.9%) of the total vegetation and ranged from 0.3% to 15%.
We found a total of sixteen non-native plant species in our plots (Table 1). Non-native plants were found in all plots. The first order jackknife estimate of the total number of non-native species in our area was 19.8 with a standard error of 1.9. The species accumulation curve increased rapidly until five plots and began flattening out around ten plots (Fig. 4), indicating that our sample provides an adequate representation of the pool of non-native species present in the part of landscape we sampled.

Percent of total vegetation cover comprised of all non-native plants, non-native annual grasses, and non-native forbs in a systematic, gridded sample of twenty 800 m2 plots following repeated, short-interval high-severity fire. Boxplots represent the minimum and maximum values, the first and third quartiles, and the median value. Outliers are represented by open circles and means are represented by stars
We found a total of ten species of non-native forbs from four different families (Table 1). Eight species were annuals and all were capable of long-distance dispersal via either animals or wind. Seven species had very low average cover (< 1%), but half of the species were frequent and found in more than 65% of the plots. Woodland ragwort (Senecio sylvaticus) was found in 95% of all plots and had the highest average cover of any forb species (5.8%). Other species from the family Asteraceae were also present at high frequencies including bull thistle (Cirsium vulgare) and prickly lettuce (Lactuca serriola).
We found a total of six species of non-native grasses. All species of non-native grasses were annuals capable of long-distance dispersal via either animals or wind. Annual silver hair grass (Aira caryophyllea) and rattail fescue (Vulpia myuros) were the most frequent species occurring in 95% and 100% of the plots, respectively. Aira caryophyllea had the highest average cover (10.6%) of all non-native grasses followed by V. myuros (5.6%). Both of these species had a maximum cover of 40%. B. tectorum was present in 40% of the plots but generally occurred at very low cover (< 1%).
Discussion
Non-native plant species, including several species of annual grasses, were ubiquitous and abundant across the study landscape dominated by a large patch of early seral vegetation created by repeated high-severity wildfire. Although non-native annual grass invasions are well documented in non-forest ecosystems (Keeley 2006), documentation of annual grass invasions following wildfire in forested ecosystems are primarily limited to cheatgrass in warmer, drier ponderosa pine (Pierson and Mack 1990; Crawford et al. 2001; Keeley and McGinnis 2007; Kerns and Day 2017; Kerns et al. In Press. Given the high level of agreement among studies projecting increases in future wildfire activity in the Klamath Mountains (Reilly et al. 2018) and the capability of the species we found for long-distance dispersal via wind or adhesion, our findings indicate a growing threat to forest resilience in the Klamath Mountains.

Species accumulation curve for non-native plants in a systematic, gridded sample of twenty 800 m2 plots following repeated, short-interval high-severity fire. Boxplots represent the minimum and maximum values, the first and third quartiles, and the median value. Outliers are represented by open circles and means are represented by stars
Forests are generally considered to be more resistant to annual grasses than warmer, drier and less productive ecosystems dominated by shrubs (Keeley and McGinnis 2007; Chambers et al. 2019). However, we found that annual grasses composed the majority of species and made up approximately 17% percent of the total vegetative cover, and cover was > 40% in almost a quarter of the plots and as high as 80%. Invasion and establishment were most likely facilitated by the second fire which altered early trajectories of vegetation change away from canopy closure and conifer establishment towards more open canopy conditions with less cover of shrubs and conifers (McCord et al. 2020). The mild, wet winters typical of lower elevations in the Klamath Mountains and the occurrence of extreme drought conditions from 2012 to 2015 (McCord et al. 2020) likely also provided suitable conditions that facilitated the invasion of some species of non-native annual grasses (Abatzoglou and Kolden 2011). While the long-term effects of fire exclusion on understory plant communities are largely unknown in this region, lack of fire for over a century may have reduced the abundance of native forbs and shrubs that could help promote resistance to invasion following fire.
The two most common and abundant annual grass species, A. caryophyllea and V. myuros, are documented within Trinity County, California where the fires occurred and are common in other parts of California (Merriam et al. 2006), but have not been documented invading post-fire communities in the Klamath Mountains. Both species were likely present in the landscape along roads prior to the first fire and may have established immediately after, then subsequently increased in abundance and expanded following the second fire. V. myruos was noted as a major fuel component capable of carrying fire in the other parts of California (Florence 1986), and was the most abundant common annual grass ten years following soil disturbance in a meadow system in northern California (Kotanen 2004). There is little know about the post-fire response of A. caryophyllea or its potential as a fine fuel to carry fire. Annual grasses are a threat to forest persistence due to their potential to alter fire regimes by providing flammable and continuous fine fuels that facilitate the movement of fire across the landscape (D’Antonio and Vitousek 1992, Mack and D’Antonio 1998). Annual grasses may also compete for moisture during the early growing season with regenerating conifers.
Non-native forbs comprised more of the total non-native species and represented less cover than grasses, but several species were relatively widespread and present in more than 60% of the plots. Two of the most frequent species, S. sylvestris and C. vulgare, are also associated with invasion following logging in this region (McDonald et al. 1996). One of the primary threats from non-native forbs includes competition with native species for resources such as water and pollinators. Non-native forbs may negatively affect conifer regeneration (e.g., Douglas-fir) by reducing early growing season moisture (Harrington and Schoennholtz 2010). Lactuca serriola is known to increase competitive interactions with native annuals in southern California due to its earlier phenology (Alexander and Levine 2019), but little is known about how such interactions affect native plant diversity and abundance in mixed-conifer/hardwood forests of this region.
While some environmental settings (e.g., serpentine soils) may be more resistant to invasions of non-native plants (Jules et al. 2011), low elevation landscapes with legacies of past management such as clearcuts and extensive road networks may be particularly susceptible to invasion as these may facilitate the spread of non-native plants (Parendes and Jones 2000). Other management related activities aimed at increasing forest resilience and resistance to fire and drought and restoration treatments such as thinning and prescribed fire (Keeley 2006; Kerns et al. In Press) and creation of fuel breaks (Merriam et al. 2006) may also promote invasion of non-native plant species. Given that many of the species we documented in this landscape may be dispersed via adhesive appendages, humans may also facilitate dispersal and spread of non-native species throughout the landscape during post-fire mitigation practices such as replanting. Such trade-offs present a major management challenge given the widespread need to restore native ecosystems and mitigate future fire severity across much of the mixed-conifer/hardwood forests in the Klamath Mountains.
Forests of this region are considered a model system for conceptual development of ecological state changes following repeated high-severity fire (e.g., Odion et al. 2010). However, the focus of previous research has been on native shrubs and hardwoods as the primary fuel capable of carrying fire during post-fire recovery. Given the potential for annual grasses to carry fire relatively soon after burning, these models are likely to underestimate the potential for state-changes due to repeated fire. In addition to fire, increased competition from non-native plants may contribute to decreased moisture availability for post-fire conifer regeneration which is already expected to decline due to increased water deficits in the future (Tepley et al. 2017). Such dynamics could accelerate observed and projected changes of conifer loss and increased dominance of hardwoods throughout the region (McCord et al. 2020).
Projected increases in temperature, fire activity, and warmer, wetter winters are anticipated to increase suitability for establishment and spread of non-native annual grasses in many parts of the western United States (Abatzoglou and Kolden 2011). We propose that non-native plant invasions following fire, and specifically repeated wildfire, are a novel stressor and under-recognized mechanism that could accelerate the loss of conifer dominance in the Klamath Mountains. Given the high capacity for long distance dispersal in all the non-native plant species we found, areas subject to short interval, high-severity fire may serve as source populations for further spread as the frequency of fire increases in the future. Additional monitoring and research will be crucial as fire activity in this region continues to increase in the future.
References
Abatzoglou JT, Kolden CA (2011) Climate change in Western US Deserts: potential for increased wildfire and invasive annual grasses. Rangel Ecol Manag 64:471–478
Abatzoglou JT, Williams AP (2016) Impact of anthropogenic climate change on wildfire across western US forests. Proc Natl Acad Sci 113:11770–11775
Alexander JM, Levine JM (2019) Earlier phenology of a nonnative plant increases impacts on native competitors. Proc Natl Acad Sci 116:6199–6204
Brooks ML, D’Antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. Bioscience 54:677–688
Chambers JC, Brooks ML, Germino MJ, Maestas JD, Board DI, Jones MO, Allred BW (2019) Operationalizing resilience and resistance concepts to address invasive grass cycles. Front Ecol Evol. https://doi.org/10.3389/fevo.2019.00185
Crawford JA, Wahren CHA, Kyle S, Mohr WH (2001) Responses of exotic plant species to fires in Pinus ponderosa forests in northern Arizona. J Veg Sci 12:261–268
D’Antonio CM, Tunison JT, Loh RK (2000) Variation in the impact of exotic grasses on native plant composition in relation to fire across an elevation gradient in Hawaii. Austral Ecol 25:507–522
D’Antonio C, Vitousek P (1992) Biological invasions by exotic grasses, the grass-fire cycle and global change. Annu Rev Ecol Syst 23:63–88
Florence M (1986) Plant succession on prescribed burn sites at Pinnacles National Monument. Fremontia 14:31–33
Fornwalt PJ, Kaufmann MR, Stohlgren TJ (2010) Impacts of mixed severity wildfire on exotic plants in a Colorado ponderosa pine-Douglas-fir forest. Biol Invasions 12:2683–2695
Harrington TB, Schoenholtz SH (2010) Effects of logging debris treatment on five-year development of competing vegetation and planted Douglas-fir. Can J For Res 40:500–510
Jepson Flora Project (eds.) (2020) Jepson eFlora, http://ucjeps.berkeley.edu/eflora/. Accessed 12 Jan 2020
Jolly WM, Cochrane MA, Freeborn PH, Holden ZA, Brown TJ, Williamson GJ, Bowman DM (2015) Climate-induced variations in global wildfire danger from 1979 to 2013. Nat Commun 6:7537
Jules ES, Ellison AM, Gotelli NJ, Lillie S, Meindl GA, Sanders NJ, Young AN (2011) Influence of fire on a rare serpentine plant assemblage: a 5-year study of Darlingtonia fens. Am J Bot 98:801–811
Keeley JE (2006) Fire management impacts on invasive plants in the Western United States. Conserv Biol 20:375–384
Keeley JE, McGinnis TW (2007) Impact of prescribed fire and other factors on cheatgrass persistence in a Sierra Nevada ponderosa pine forest. Int J Wildland Fire 16:96–107
Kerns BK, Day MA (2017) The importance of disturbance by fire and other abiotic and biotic factors in driving cheatgrass invasion varies based on invasion stage. Biol Invasions 19:1853–1862
Kerns BK, Tortorelli C, Day MA, Nietupski T, Barrows AMG, Kim JB, Krawchuk MA (In Press) Invasive grasses: a new perfect storm for forested ecosystems. For Ecol Manag
Klinger R, Wills R, Brooks ML (2008) Fire and nonnative invasive plants in the southwest Coastal Bioregion. In: Zouhar K, Smith JK, Sutherland S, Brooks ML (eds) Wildland fire in ecosystems: fire and nonnative invasive plants. Gen Technol Rep RMRS-GTR-42, vol 6. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, p 355
Kotanen PM (2004) Revegetation following soil disturbance and invasion in a Californian meadow: a 10-year history of recovery. Biol Invasions 6:245–254
Mack RN (1981) Invasion of Bromus tectorum L. into western North America: an ecological chronicle. Agro Ecosyst 7:145–165
Mack MC, D’Antonio CM (1998) Impacts of biological invasions on disturbance regimes. Trends Ecol Evol 13:195–198
Mack RN, Pyke DA (1983) The demography of Bromus tectorum: variation in time and space. Journal of Ecology 71:69–93
McCord M, Reilly MJ, Butz RJ, Jules ES (2020) Early seral pathways of vegetation of vegetation change following short interval, repeated wildfire in low elevation mixed-conifer-hardwood forest landscape of the Klamath Mountains, Northern California. Can J For Res 50:13–23
McDonald PM, Fiddler GO (1996) Development of a mixed shrub-tanoak-Douglas-fir community in a treated and untreated condition. Res. Pap. PSW-RP-225. Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station, p 16
Merriam KE, Keeley JE, Beyers JL (2006) Fuel breaks affect nonnative species abundance in Californian plant communities. Ecol Appl 16:515–527
Miller JD, Thode AE (2007) Quantifying burn severity in a heterogenous landscape with a relative version of the delta Normalized Burn Ration (RdNBR). Remote Sens Environ 109:66–80
Miller JD, Skinner CN, Safford HD, Knapp EE, Ramirez CM (2012) Trends and causes of severity, size, and number of fires in northwestern California. Ecol Appl 22:184–203
Odion DC, Mortitz MA, DellaSala DA (2010) Alternative community states maintained by fire in the Klamath Mountains, USA. J Ecol 98:96–105
Oksanen J, Kindt R, Legendre P, O’Hara B, Stevens MHH, Oksanen MJ (2009) Vegan community ecology package. R package version 1.13-2. http://vegan.r-forge.r-project.org
Parendes LA, Jones JA (2000) Role of light availability and dispersal in exotic plant invasion along roads and streams in the H.J. Andrews Experimental Forest, Oregon. Conserv Biol 14:64–75
Pierson EA, Mack RN (1990) The population biology of Bromus tectorum in forests: distinguishing the opportunity for dispersal from environmental restriction. Oecologia 84:519–525
Reilly MJ, Dunn C, Meigs G, Spies TA, Kennedy R, Bailey J, Briggs K (2017) Contemporary patterns of fire extent and severity in forests of the Pacific Northwest. Ecosphere 8(3):e01695. https://doi.org/10.1002/ecs2.1695
Reilly MJ, Spies TA, Littell J, Butz R, Kim J (2018) Vulnerability of vegetation to climate change and disturbance in the Northwest Forest Plan area. In: Synthesis of science to inform land management within the Northwest Forest Plan Area. USFS PNW GTR-966, vol 1
Taylor AH, Skinner CN (2003) Spatial patterns and controls on historical fire regimes and forest structure in the Klamath Mountains. Ecol Appl 13:704–719
R Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Tepley AJ, Thompson JR, Epstein HE, Anderson-Teixeira KJ (2017) Vulnerability to forest loss through altered postfire recovery dynamics in a warming climate in the Klamath Mountains. Glob Change Biol 10:4117–4132
USDA Forest Service (2007) Common stand exam field guide. Natural Resource Information System, Washington, DC. http://www.fs.fed.us/nrm/fsveg/
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western U.S. forest wildfire activity. Science 313:940–943
Whittaker RH (1960) Vegetation of the Siskiyou Mountains, Oregon and California. Ecol Monogr 30:279–338
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Reilly, M.J., McCord, M.G., Brandt, S.M. et al. Repeated, high-severity wildfire catalyzes invasion of non-native plant species in forests of the Klamath Mountains, northern California, USA. Biol Invasions 22, 1821–1828 (2020). https://doi.org/10.1007/s10530-020-02227-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-020-02227-3