
Abstract
Herbivore and plant invasions can modify the structure and functioning of ecosystems through positive interactions that facilitate their mutual establishment. An important consequence of the feeding behaviour of rabbits is that they can play a key role in seed dispersal by endozoochory. We examined the diet and potential for dispersing Rosa rubiginosa and R. canina (Rosa spp.) of European rabbits introduced in a semiarid protected area of Argentina. We found entire seeds in 100% of the samples of Rosa spp. analysed (n = 11, 187 pellets). Our results indicate that the passage of Rosa spp. seeds through the digestive tract of rabbits (64%) did not diminish their viability in comparison to the seeds taken directly from fruits (66%) (χ2 = 0.09, p > 0.05). This study has identified new positive interactions between an invasive mammal and two invasive plants, a finding that suggests that these invader complexes could have a reciprocal effect on one another, which would potentially aid their invasive process in a semiarid protected area of Argentina.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The presence of a new species in the system can cause various effects on a native community such as changes in interactions or appearance of new ones (Blackburn et al. 2014). Positive interactions occur frequently among non-native species, suggesting that mutualisms are important in several invasion scenarios (Braga et al. 2018). For example, introduced herbivorous mammals launch interactions with non-native plants (plant-disperser interactions), which promotes species integration in the invaded community. Thus, if the interacting species are both introduced and prove a beneficial outcome to each other, they could be contributing to a process by which non-native species facilitate one another’s invasion (invasional meltdown) (Traveset and Richardson 2006, Simberloff and Van Holle 1999). The fruit and seed consumption by herbivores may benefit the plant, not only, due to viable seeds eliminated in faeces at suitable sites for germination and survival, but also, because the plant escape of the high mortality rates close to the parent plant (dispersal by endozoochory; Howe and Smallwood 1982; Janzen 1980). Hence, invader complexes created among non-native plant–seed dispersers may contribute to success of their invasion and, therefore, to their expansion in the invaded range (Traveset and Richardson 2006). Particularly, lagomorphs may disperse viable seeds of grasses and forbs consumed incidentally while feeding on foliage and, of fleshy-fruited plants whose fruits are intentionally consumed (Schupp et al. 1997).
Aridlands are water-controlled ecosystems with infrequent discrete largely unpredictable water inputs, where most of the plants have low nutritional value and where herbivores are dependent on vegetation for food and water (Noy-Meir 1973). Three invasive species that occur in arid and semiarid areas of Argentina, coexist in our study area: the mosqueta roses, Rosa rubiginosa and R. canina (hereafter Rosa spp.) and the European rabbit, Oryctolagus cuniculus (Dalmasso et al. 2011, Cuevas et al. 2011). Rosa spp. have a set of attributes that make them a good invader. These shrubs possess a broad range of reproductive strategies to produce fruits and seeds (e.g. production of a large mass of seeds, longevity of seeds, more fruits availability in periods of food shortage) which enable them to colonize new areas through dispersal vectors, such as birds and mammals (Mazzolari et al. 2017). Another invasive component of this arid ecosystem is the European rabbit, native to the Iberian Peninsula and the south of France. The European rabbit is a generalist herbivore with high plasticity in selecting food (Gálvez-Bravo et al. 2008). Rabbits act as endozoochorous seed dispersers of different plant species in their area of origin, as well as in areas where they have been introduced (Castro et al. 2008; Salas-Pascual et al. 2009). As a result of their feeding behaviour and their role as seed dispersers, rabbits can affect the structure, composition, and functioning of plant communities in invaded environments.
Based on the invasional meltdown hypothesis (Simberloff and Van Holle 1999), European rabbit and Rosa spp. have a reciprocal positive effect on one another. In particular, we hypothesize that European rabbit plays a major role as potential endozoochoric seed dispersers of Rosa spp., so it could be expected that the rabbits will consume fruits of Rosa spp. and they will eliminate its viable seeds through faeces. Therefore, the aim of our study was to explore the trophic ecology of the European rabbit, particularly its food habits, and the role as an endozoochorous seed disperser of Rosa spp. in a semiarid protected area of Argentina.
Materials and methods
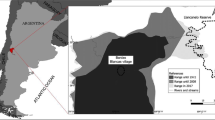
The study area was located on the semiarid Andean foothills in the Natural Reserve of Villavicencio, Mendoza province, Argentina (32° 31′ S–69° 1′ W, Fig. 1). The reserve occupies 62,000 ha and its altitudinal range varies from 900 to 3200 m a.s.l. Annual precipitation is between 120 and 300 mm. The reserve protects three important phytogeographic arid biomes: Puna, Cardonal and Monte (Dalmasso et al. 1999). Sampling was conducted during the wet season 2009–2010 (December to March). We established 24 strip transects (100 × 10 m) from the site where rabbits were intentionally released in 2006 (Cuevas et al. 2011) following the topography of the area (Fig. 1). Transects were placed randomly within two geomorphological units, hillsides (n = 12) and riverbeds (n = 12), of the three ravines covering the greater environmental heterogeneity: (1) Hornillos; (2) Darwin and (3) Toro (Fig. 1). These sites present a characteristic vegetation of the Cardonal Phytogeographic province dominated by shrubs of the genera Lycium, Baccharis, Artemisia and Adesmia, and by grasses of the genera Stipa, Poa and Aristida. Riverbeds are characterized by the presence of non-native plant species including Rosa spp. (Dalmasso et al. 1999).

modified from Cuevas et al. 2011
Left: Distribution of European rabbit (Oryctolagus cuniculus) in Mendoza province (dashed line) and study area (white circle). Right: Potential dispersal corridors: Hornillos Ravine (1), Darwin Ravine (2) and the third path towards the Monte desert foothills (3) in the Natural Reserve of Villavicencio. Map extracted and
We collected fresh rabbit faeces along the transects checked one time during wet season. All faeces collected in one transects constituted one sample for diet analysis. At this transects, plant cover was measure through point quadrat method (Passera et al. 1983) for food availability data. We also collected Rosa spp fruits and fruits from several plants to make a reference collection. To determine the rabbit’s diet, we used fresh faeces from each transect, which were evaluated through microhistological techniques and analysis that permit identification of leaf epidermis, stems, seed teguments and arthropod body parts (Dacar and Giannonni 2001). For each sample, we prepared microscope slides and systematically examined 50 fields under 40X microscope. Food items in the faecal samples were identified by comparison to reference material. Presence of a food item was recorded, and its relative frequency per slide was determined by dividing the number of microscope fields in which this item occurred by the total number of microscope fields observed × 100 (Holechek and Gross 1982).
Plant species were clustered into four categories according to life form: herbs, grasses, sub-shrubs (shrubs less than 1 m tall) and shrubs, and we estimated the proportion of each food category. Food availability was calculated from vegetation data obtained from transects. Selectivity of food items was estimated using Manly’s Selectivity Index (Manly et al. 2002): αi = (Pui/Pai)/Σ(Pui/Pai), where Pui is the observed proportion of item i in the rabbit’s diet and Pai is the available proportion of item i in the environment. If αi is greater than 1/k, being k the number of food items, indicate selection (greater than random). If αi is less than 1/k, indicate avoided, and, αi = 1/k indicate a consumption in accordance to availability (Manly et al. 2002). To test the reliability of Manly’s index, we resampled rabbit faeces 1000 times by bootstrapping. We then calculated the average values and the 95% confidence intervals of Manly’s index. All analyses were performed with the package Quantmod (Ryan et al. 2018) in environment R 3.5.1 software (R Development Core Team 2018).
To evaluate the viability of the entire sweet briar seeds found in rabbit faeces, we used a standard bioindicator (2, 3, 5 triphenyltetrazolium chloride (TTC)) that detects seed viability by staining the embryo tissue pink/red. Conditions for the tetrazolium test for the genus Rosa was taken from the International Rules for Seed Testing (2005). Viability of seeds was determined by their coloration, using microscope amplification in comparison to positive control seeds collected from the study site. A Chi square statistic test of homogeneity (Zar 2010) was used to analyse differences in the proportion of viable seeds obtained from faeces and the control treatment.
Results and discussion
We collected rabbit faeces from almost half the sampled transects (n = 11) where the 75% was recorded on riverbed transects and on only 2 hillside transects. The presence of rabbit signs in riverbed transects could be explained by the influence of factors like vegetation, soil, and places with streams or higher moisture (Cuevas et al. 2011). Hundred percent of the samples contained 135 entire seeds of Rosa spp. (0.7 ± 0.9 roses seeds. pellet−1), whereas 36.4% of the samples were found also damaged seeds (Table 1). Approximately, 27% of the 135 digested entire seeds were non-productive, that is, empty seeds, with a poorly developed embryo, infected by fungi or by wasp larvae (Table 1). The proportion between seed damaged/entire was low, suggesting that European rabbit plays a less important role as seed predator than as disperser to Rosa spp.
The rabbit’s diet was composed of different parts (leaves and seeds) of 30 plant species. Seed teguments represented 6% of the diet and all corresponded to Rosa spp. The European rabbit consumed mainly herbs (50%), followed by subshrubs (17.9%), shrubs (16.5%) and grasses (15.5%) (Fig. 2). The bulk of the diet (more than 70%) was composed of: Medicago lupulina (Fabaceae) (32.6%), Poa resinulosa (Poaceae) (11.7%), Lycium sp. (Solanaceae) (10.3%), Spartium junceum (Fabaceae) (6.5%), M. sativa (6.4%) and seeds teguments of Rosa spp. (Rosacea) (5.7%) (Table 2). Previous reports on the diet of rabbits in Argentina’s Patagonia showed that grasses represent the main category in the diet throughout the year whereas herbs had moderate participation in the spring and summer diets (Bonino and Borelli 2006). In its native Mediterranean ecosystem, the rabbit has been shown to accommodate its diet to the resource availability resulting from interactions with the other herbivores coexisting in the area (Soriguer 1983). Thus, when herbaceous plants are scarce by the presence of other herbivores, rabbits consume mainly grasses whereas, if grazing alone, they prefer herbs (Soriguer 1988). This could be happening inside the protected area, where the rabbit is one of the primary herbivores along with the non-native European hare (Lepus europaeus) and native guanaco (Lama guanicoe), and there is no great grazing pressure by livestock on herbaceous vegetation.

Frequency of occurrence (mean ± SE) of each food categories in European rabbit diet during wet seasons in Reserve Villavicencio, Argentina
Trophic selection for food categories was positive for herbaceous plants, while grasses, sub-shrubs and shrubs were avoided (Fig. 3a). Selection for food items was positive for four out of the 13 species of herbs consumed (Fig. 3b and Table 2). Rabbits consumed a higher proportion of the species they selected (about 50%), mainly the non-native Medicago lupulina. This species is considered a good forage resource due to its nutritional value (Passera et al. 1983). Consuming nourishing food could be important especially in arid and semi-arid ecosystems where the majority of plants have low nutritional value (Noy-Meir 1973). Even if the European rabbit did not select seeds of Rosa spp., showed a consumption greater than 5% in the diet during the wet season being the 6th item more consumed (Fig. 3b and Table 2). Previous studies have shown that the rabbit consumption of the woody species increases when the availability of herbaceous decreases by seasonality variation (Bonino and Borelli 2006). For example, comparison with dry season could showed a great consume fruits of Rosa spp. because constitute a nutritious, accessible and abundant food in this period. Therefore, the relative importance of rabbit–Rosa spp. interaction may largely depend on fruits and seeds availability in the invaded community in a given season.

Manly’s selectivity index (± 95% CI) for food categories (a) and to food items (b). Dotted line indicates 1/k = 0.25 and 1/k = 0.03 for a use proportional to the availability respectively. Black circle = Herbs; dark gray circle = Grasses; light gray circle = Subshrubs and white circle = Shrubs
Seventy-three percent of the entire seeds presented healthy embryos (Table 1). We found no significant viability differences (χ2 = 0.09, df = 1, p > 0.05) between seeds obtained from faeces (digested; 64% viable) and seeds obtained from fruits (control; 66% viable) (Table 1). Our results suggest that the passage of seeds of Rosa spp. through the digestive tract of rabbits does not affect their viability. The relatively large number of viable seeds after ingestion by rabbits could provide an advantage if seeds are deposited at suitable sites, far from the parent plant (Schupp et al. 2010). Further research regarding these new interactions should consider another aspect such as plant benefits resulting from seed deposition patterns, field germination success, and abundance, feeding strategies and movement of seed dispersers (Schupp et al. 2010).
Our study identifies new interactions between an introduced mammal and two invasive plants, and suggests that these invader complexes could have a reciprocal effect on one another, potentially aiding their invasive process (and subsequent impacts) in the semiarid foothills in the Andes of Argentina. Further studies should consider other components to understand the effectiveness of European rabbit as a seed disperser of Rosa spp. such as quality of treatment of seed by germination tests (legitimacy), quality of seeds deposition (efficiency) or quantitatively through the number of seeds dispersed per plant. We reinforce the importance of studies that may detect new positive interactions between non-native species, such as the other invasive herbivorous mammal Lepus europaeus inside of areas by high conservation value. These positive interactions would potentially cause an invasional meltdown and leading to an accelerating increase in number of introduced species and their impact on native community.
References
Blackburn TM, Essl E, Evans T, Hulme PE, Jeschke JM et al (2014) A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol. https://doi.org/10.1371/journal.pbio.1001850
Bonino NA, Borrelli L (2006) Variación estacional de la dieta del conejo silvestre europeo (Oryctolagus cuniculus) en la región andina de Neuquén, Argentina. Ecología Austral 16:7–13
Braga RR, Gómez-Aparicio L, Heger T, Vitule JRS, Jeschke JM (2018) Structuring evidence for invasional meltdown: broad support but with biases and gaps. Biol Invasions 20:923–936
Castro S, Bozinovic F, Jaksic FM (2008) Ecological efficiency and legitimacy in seed dispersal of an endemic shrub (Lithrea caustica) by the European rabbit (Oryctolagus cuniculus) in central Chile. J. Arid Environ 72:1164–1173
Cuevas MF, Chillo V, Marchetta A, Ojeda RA (2011) Mammalia, Lagomorpha, Leporidae, Oryctolagus cuniculus Linnaeus, 1758: new record and its potential dispersal corridors for northern Mendoza, Argentina. Check List 7:565–566
Dacar MA, Giannoni SM (2001) A simple method for preparing reference slides of seed. J Range Manag 54:191–193
Dalmasso DD, Martinez Carretero E, Videla F, Puig S, Candia R (1999) Reserva natural Villavicencio (Mendoza, Argentina). Plan de manejo. Multequina 8:11–50
Dalmasso AD, Almirón M, Arroyo N (2011) Cerramientos vivos con especies nativas y exóticas rústicas. Experimentia 1:3–56
Gálvez-Bravo L, Belliure J, Rebollo S (2008) European rabbits as ecosystem engineers: warrens increase lizard density and diversity. Biodivers Conserv 18:869–885
Holechek J, Gross B (1982) Evaluation of different calculation procedures for microhistological analysis. J Range Manag 35:721–723
Howe FH, Smallwood J (1982) Ecology of seed dispersal. Annu Rev Ecol Evol Syst 13:201–228
Janzen DH (1980) When is it coevolution? Evolution 34:611–612
Manly BFJ, McDonald LL, Thomas DL (2002) Resource selection by animals. Statistical design and analysis for field studies, 2nd edn. Chapman & Hall, London
Mazzolari AC, Marrero HJ, Vázquez DP (2017) Potential contribution to the invasion process of different reproductive strategies of two invasive roses. Biol Invasions 19:615–623
Noy-Meir I (1973) Desert ecosystems: environment and producers. Annu Rev Ecol Evol Syst 4:25–51
Passera CB, Dalmasso AD, Borsetto O (1983) Método de Point Quadrat Modificado. Taller sobre arbustos forrajeros. FAO, IADIZA, Mendoza
Ryan JA, Ulrich JM, Thielen W, Teetor P, Bronder S (2019) Quantmod: Quantitative Financial Modelling Framework. R package version 0.4-13. https://CRAN.R-project.org/package=quantmod. Accessed 17 June 2019
Salas-Pascual S, Lugo SF, Cigala AN (2009) Interaction between two exotic invading species: endozoochory of Acacia farnesiana seeds by the European rabbit (Oryctolagus cuniculus). Open For Sci J 2:86–90
Schupp EW, Heaton HJ, Gomez JM (1997) Lagomorphs and the dispersal of seeds into communities dominated by exotic annual weeds. Great Basin Nat 57:253–258
Schupp EW, Jordano P, Gomez JM (2010) Seed dispersal effectiveness revisited: a conceptual review. New Phytol 188:333–353
Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: invasional meltdown? Biol Invasions 1:21–32
Soriguer RC (1983) Consideraciones sobre el efecto de los conejos y los grandes herbívoros en los pastizales de la Vera de Doñana. Doñana Acta Vertebrata 10:155–163
Soriguer RC (1988) Alimentación del conejo (Oryctolagus cuniculus L. 1758) en Doñana. SO, España. Doñana Acta Vertebrata 15:141–150
Traveset A, Richardson DM (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol Evol 21:208–216
Zar JH (2010) Biostatistical analysis. Pearson Education, New Jersey
Acknowledgements
We thank Diego F. Vazquez for comments on this manuscript. Nelly Horak assisted in the English version. We are very grateful to the staff of Natural Reserve of Villavicencio and Recursos Naturales Renovables of Mendoza province. To the continuous support of our institution IADIZA-CONICET, CCT. This study was partially funded by ALARM (EU).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bobadilla, S.Y., Marchetta, A., Dacar, M.A. et al. Food habits of European rabbit and its role as seed dispersal of two Mosqueta roses: Facilitation among non-native species in a semiarid protected area of Argentina?. Biol Invasions 22, 1565–1571 (2020). https://doi.org/10.1007/s10530-020-02205-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-020-02205-9