
Abstract
Non-native species often acquire novel interspecific interactions, which are central to several hypotheses of invasion success, including biotic resistance and invasional meltdown. However, the outcome of these interactions is not often linked with the demographic evidence based on the full life cycle of the species. The Philippine Ground Orchid (Spathoglottis plicata) has invaded Puerto Rico and has acquired both negative and positive interspecific interactions involving the native weevil Stethobaris polita and the invasive red fire ant Solenopsis invicta, respectively. We studied a population in the Rio Abajo Forest, and asked how these interactions affect population demography by using a combination of field, experimental and modelling approaches. Stage-structured matrix population models based on four years of field observations showed that the population of S. plicata is growing at a rate (λ) of 1.05 under natural conditions. When we modified fecundity values based on experimental exclusion of weevils and ants, the control treatment showed a similar λ. Excluding weevils increased λ to 1.20, whereas the exclusion of ants decreased λ to 1.03. When we incorporate demographic and environmental stochasticity in our models, exclusion of invasive red fire ants significantly reduces the orchid abundance over time. Although weevils offer some biotic resistance to S. plicata, these effects do not prevent orchid population growth and expansion. On the other hand, invasive red fire ants have a positive effect on the invasive orchid’s λ, partially supporting the invasional meltdown hypothesis. This study presents a method that allows one to combine opposing mechanisms of species interactions within the same quantitative framework, and the results highlight the importance of considering acquired plant–animal interactions and stochastic processes when evaluating the population growth rates and dynamics of invasive plants.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Introduced non-native species often acquire novel interspecific interactions outside their native range, which affect the outcome of the invasion (Levine and D’Antonio 1999; Levine et al. 2004; Richardson et al. 2007). The effects of these biotic interactions on the population performance of non-native species may vary from negative, neutral, to positive, and serve as a basis for different hypotheses in invasion biology. For example, non-native species that are kept in check by predators or herbivores in their native range may experience a release from these antagonistic interactions in their introduced range, which positively influences the population performance (Elton 1977). On the other hand, receptive native communities may have local predators or herbivores that preferentially consume non-native species, and thus negatively affect their invasion success (Parker and Hay 2005). Moreover, biotic interactions between non-native species may have positive synergistic effects in a way that increases their invasion success, which in turn negatively affect the receptive communities (Simberloff and Von Holle 1999; Lach et al. 2010).
Understanding the demography of invasive species is essential to comprehend the processes that contribute to the establishment and spread of a particular species outside its native range, and to develop sound management strategies when needed. Many studies on species invasions measure specific life history traits to study the role of acquired interspecific interactions in the novel ecosystems (Levine et al. 2004; Mitchell et al. 2006). However, the outcomes of these interactions are not often linked with the demographic evidence which integrates an approximation of the full life cycle of the species, and measured at the population level.
Matrix population models provide the means to combine multiple vital rates based on life cycles, that allows one to evaluate how changes in these rates affect population dynamics (Caswell 2001; Sakai et al. 2001; Morris and Doak 2002). Although such models have been used to evaluate population viability, and to develop management strategies for the conservation or control of species populations (Hastings 1997; Caswell 2001; Morris and Doak 2002; Fieberg 2004), their use to understand the effects of acquired interspecific interactions on the population dynamics of invasive plants remains limited. Using these models could offer a more quantitative assessment to dissect the importance of processes regulated by biotic interactions in relation to other processes affecting the invasion success of a given species, and allows one to combine opposing mechanisms within the same quantitative framework.
In this study we assess how matrix population models can serve to quantify the effects of antagonistic and beneficial acquired interspecific interactions on the demography of invasive plants by coupling demographic data of the invasive Philippine Ground Orchid (Spathoglottis plicata Blume) with data from exclusion experiments in Puerto Rico. The Philippine Ground Orchid is one of several species of Orchidaceae that have escaped cultivation and established invasive populations in Puerto Rico, and was generally considered innocuous (Ackerman 2007; Ackerman et al. 2011; Recart et al. 2013; Ackerman et al. 2014). However, a recent study found that S. plicata is having negative ecological impacts on the native orchid Bletia patula Graham through apparent competition mediated by a native orchid-specialist weevil (Stethobaris polita Chevrolat) (Recart et al. 2013). Moreover, invasive red fire ants (Solenopsis invicta Buren) forage at the extrafloral nectaries present in the inflorescences of S. plicata, and may protect the inflorescences against weevil florivory and oviposition (Ackerman et al. 2014).
We first asked what life-cycle stages are more important for the invasive orchid’s success, and what is the population growth rate (λ) under natural field conditions? By evaluating the resulting projection matrices based on the demographic field study carried out in a population of S. plicata for four continuous years, we expected to identify important life-cycle stages and transitions for the orchid’s population performance. Moreover, and because S. plicata is an invasive species expanding its range in Puerto Rico, we expected our population to exhibit a positive λ. We then asked what are the effects of acquired interspecific interactions on the invasive orchid’s population growth rate? We assessed this question by incorporating into our matrix population models the expected fecundity based on reproductive success data derived from ex situ weevil and ant exclusion experiments. We anticipated to find agreement between the resulting population performance metrics of both our field (observed demography) and control (exclusion experiment treatment)-derived matrices as a validation to our approach (both include the three-way interactions). We expected to find that native weevils significantly reduce λ, but fail to control the invasive orchid, whereas invasive red fire ants increase λ by protecting the inflorescences. Finally, we sampled plants throughout their invasive range in Puerto Rico and Hawai’i, and compared their reproductive success in the presence or absence of antagonistic weevils. We expected to find significantly lower reproductive success in populations were weevils were present.
Methods
Study area
Our study site was located along Highway 10 in the Río Abajo State Forest (18°20.96N, 66°41.07W) in the municipality of Arecibo, which lies within the northern karst region of Puerto Rico, and is classified as Subtropical Moist Forest according to the Holdridge Life Zone system (Ewel and Whitmore 1973). The vegetation in the study area was characteristic of early secondary succession, with grasses and ferns dominating the area, followed by scattered shrubs (e.g., Piper aduncum L., Gesneria pedunculosa (DC.) Fritsch) and young trees (e.g., Tabebuia heterophylla (DC.) Britton). As a result, the area is open with high light availability, although local variation exists.
Study system
Spathoglottis plicata is a caespitose perennial herb that depends on seed production and wind dispersal for seedling establishment and population growth, with a mixture of autopollination (sensu Kearns and Inouye 1993) to outcrossing reproductive strategies within and between populations throughout its range (Ackerman et al. 2014). Spathoglottis has sympodial growth but because rhizomes are very short, the pseudobulbous shoots are caespitose. As pseudobulbs age they may lose their leaves, but new shoots form so that healthy plants are continuously photosynthetic (and lack a dormant stage at our site). Spathoglottis plicata can survive for over four years (this study), and rarely do plants become large enough to form independent ramets so clonal reproduction is unlikely to be a significant contributor to λ. Individual plants usually produce a single many-flowered inflorescence at a time, but will develop several over the course of a year. Flowers either develop into mature fruits approximately four weeks after pollination, or are either dropped (if not pollinated) or aborted, leaving a scar in the inflorescence. Fruits stay attached to the inflorescence long after dehiscence. Once seeds are dispersed, they may germinate in 1–2 weeks and flower within a year (Prakash and Leelee 1973; Thompson and Wright 1995; Ackerman et al. 2014).
The Philippine Ground Orchid is the most common and widespread species of the genus, with several forms cultivated and sold as ornamentals. The autopollinated forms of S. plicata have escaped cultivation and exotic populations have been reported in West Africa, Kenya, Mascarene Islands, Seychelles, Hawai’i, Florida, Cuba, Dominican Republic, Puerto Rico, US Virgin Islands, the Lesser Antilles, and Panama (Jolliffe 2010; Ackerman et al. 2014). In Puerto Rico, S. plicata mostly exploits naturally or anthropogenically disturbed (open) areas of moist and wet regions from near sea level to about 1000 m (Ackerman 1995; Ackerman et al. 2014). However, they can also be found inside mature forests, especially along riparian areas (WF personal observation). The first naturalized population was discovered in the mid 1970s within El Yunque National Forest in eastern Puerto Rico (R. Woodbury personal communication to JDA, 1981), and became noticeably common by the first decade of the new millennium. Based on local herbarium holdings (MAPR, UPR, and UPRRP), two specimens were collected in the 1980s, one in the 1990s, and 31 in the 2000s, and location records over time seem to indicate east to west spread. Collecting efforts were intense during these decades as floristic studies were in progress (Ackerman 1995; Axelrod 2011), so we may assume that the collection records reflect the lag-phase often observed in invading species. Populations of S. plicata have been established in the Río Abajo State Forest since at least 1995, when the first populations were reported (Axelrod 9190, UPRRP). Currently, white and pink flowered morphs are found on the island, and fruit set (FS; a measure of reproductive success given by the number of fruits divided by the sum of fruits and scars) can be over 50% (Recart et al. 2013). Seed set (proportion of seeds with visible embryos) in our study area can reach up to 90% (JDA personal observation). This is comparable to populations in Hawai’i, where fruit set is 66% (Kores 1980), and the mean seed set is 76% (±19; N = 12).
In Puerto Rico, S. plicata has acquired several interspecific interactions (see Ackerman et al. 2014 for a detailed description of the system). Stethobaris polita is a relatively small black weevil, which is an orchid specialist native to Puerto Rico that feeds on flowers and oviposits in fruits, thus reducing the reproductive output of both invasive and native orchids (Recart et al. 2013; Ackerman et al. 2014). The average seed set of S. plicata fruits with oviposition is 8% ± 0.17 (N = 20 fruits), but it is unknown whether these seeds would ultimately be viable as we often found rotten fruits with fungal and/or bacterial infections. When weevil damage is severe, an inflorescence will exhibit rotting fruits, buds and apices (Ackerman et al. 2014).
In Puerto Rico, red fire ants (Solenopsis) forage for extrafloral nectar produced on buds and floral bracts of S. plicata. Ants of the genus Solenopsis have a cosmopolitan distribution, occupy a wide range of niches, and several species are native to Puerto Rico (Smith 1936). In the Rio Abajo State Forest, the invasive S. invicta was the only red fire ant observed foraging for extrafloral nectar, they outnumbered other species of ants visiting the inflorescences 2 to 1, and they can attack or deter weevils from feeding on the orchids (Ackerman et al. 2014). Solenopsis invicta is a common invasive species throughout the world, and the polygynous forms are widespread in disturbed and open areas in Puerto Rico and other Caribbean islands (Global Invasive Species Database; Davis et al. 2001; Wetterer and Snelling 2006). While they may be passengers rather than drivers of ecosystem change (King and Tschinkel 2008), they are known to cause negative impacts on community structure, particularly arthropods (e.g. Porter and Savignano 1990).
Field methods
To study the population biology of S. plicata in our study site, we established a total of 8 (1 × 3 m) plots (8–10 m distant from each other), from which we gathered demographic data. In each of the plots, we located and identified all individuals of S. plicata using unique-numbered metal tags (attached to the base of each plant). Four censuses were conducted: the first in January 2009, and subsequently in November of 2009–2011. As a consequence, the first time interval is 11 months, and the other two were 12 months. After taking into account the differences in time intervals, we found that the effects of having an 11-month time interval for the first census did not affect our results and inference, so we used this time interval without correcting for time. The four different censuses conducted from early 2009 until late 2011 are referred to as t 0, t 1, t 2, and t 3, and the time transitions 2009–2009, 2009–2010, and 2010–2011 are transitions t 0–t 1, t 1–t 2, and t 2–t 3, respectively.
For every individual in each census, we measured the width and length of the longest leaf, as well as the reproductive status (i.e. presence of active inflorescences, number of opened flowers, the number of fruits, the proportion of damaged fruits, and the number of scars present). Beginning in the second census, we recorded the number of new recruits, as well as the number of dead individuals.
Model construction and parameterization
To project the population dynamics of S. plicata at this site, we used size-structured matrix population models (Lefkovitch 1965). The width of the longest leaf was used as a measure of size for the model. Previous studies have successfully used a variety of leaf measures as indicators of size for orchids (Montalvo and Ackerman 1987; Tremblay and Hutchings 2003), and preliminary analyses indicated that reproductive effort (RE; total number of flowers produced given by the sum of fruits and scars) was significantly correlated with this variable (Kendall τ = 0.28, p < 0.001; for the final dataset). We employed the Vandermeer–Moloney algorithm for determination of size-stage categories (Vandermeer 1978; Moloney 1986). This algorithm minimizes both the sampling error, and the error of distribution (Vandermeer 1978). The sampling error measures choosing a category that is too small, while the error of distribution measures choosing a category that is too big. This method is appropriate when a species has no traits that distinctly define life history stages (Rosenberg et al. 2005).
Precise measures of fitness in the Orchidaceae are not practical because of the minute size of the seeds, the large number of seeds produced in a single fruit, and the difficulty in measuring the contribution of adult plants to seedling establishment. As an alternative, Tremblay and Hutchings (2003) suggested that fecundity values for this family may be calculated using the number of fruits produced by flowering plants or the number of new recruits to the population (with the latter being more biologically relevant). Unfortunately, this method does not credit the production of recruits to specific parents (Tremblay and Hutchings 2003), which could be problematic when more than one size-stage contributes to the production of new recruits and that recruitment is not a linear function. However, the number of recruits to an orchid population is often assumed to be related to fruit-set (Tremblay and Hutchings 2003), which has been demonstrated in epiphytic orchids (Ackerman et al. 1996). Assuming that the number of new recruits added to a population is related to fruit-set, and that fruit production differs among plant size-stages, we calculated the recruitment rate as a ratio of new recruits added to the population to the total number of fruits produced in the previous year. We used this approach to determine the expected proportional contribution of a given size-stage as a function of the number of fruits produced at a given size-stage (see S1 for detailed methodology).
The Vandermeer–Moloney algorithm applied to the size distribution of the first transition (t 0–t 1) resulted in six size categories (Table 1). After establishing the size categories, we calculated the average life stage transition probabilities for each matrix (Caswell 2001) (see S2 for detailed methodology). The calculations of the demographic parameters and the asymptotic population growth rates (λ; maximum eigenvalue) from the projection matrices were done using the R package ‘popbio’ (Caswell 2001; Stubben and Milligan 2007).
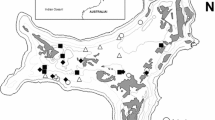
We calculated the fecundities as the number of recruits produced per individual at a given size stage under observed natural field conditions within our demography plots (from here on, “field conditions”; see S2 for details). This represents the matrix under natural conditions, which includes the influence of the interactions with weevils and ants. Thus, the resulting asymptotic population growth rates (λ), and the geometric mean population growth rates (λgeom; mean λ between time transitions) obtained from the matrices derived from our field data should reflect what is happening in our study population. Figure 1 shows the simplified life cycle of S. plicata based on the observed transitions in our field study, and the interspecific interactions with native weevils and invasive red fire ants, which occur only on reproductive individuals.

Weevil image by Tom Murray, and ant image by WP Amstrong (modified and used with authorization)
Simplified life cycle of Spathoglottis plicata based on the observed stage-size transitions and fecundity contributions in our field demography study, and the acquired interactions with native weevils and invasive ants in Puerto Rico. In the life cycle, “R” represents recruits, and numbers represent the size-stages. Native weevils (Stethobaris polita) feed on orchid flowers and oviposit in fruits, while invasive red fire ants (Solenopsis invicta) forage at extrafloral nectaries and often attack weevils. Orchid drawing from Lewis Roberts (modified): [CC BY-SA 3.0 (http://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia Commons.
We analyzed the contribution of each demographic parameter to the asymptotic population growth rate using the elasticity analyses (Caswell 2001). This analysis shows the relative contribution of the individual life-cycle transitions to the population growth rate (Caswell 2001). Elasticities can be used to determine which life-cycle transitions have the most influence on λ, and thus help delineate sound management strategies for the control (or conservation) of populations (Horvitz and Schemske 1995; Hastings 1997; Morris and Doak 2002; McCullough et al. 2006).
Effects of acquired interspecific interactions on λ
To investigate the effects of acquired plant–animal interactions on the orchid’s λ, we used the approach described previously for calculating the fecundities, in combination with data from this study and exclusion experiments from Ackerman et al. (2014), whose study sites were approximately 100 m distant from ours, and whose study duration overlapped with ours. They performed four treatments that included: (a) control-unmanipulated inflorescences accessible to weevils and ants (0.145 ± 0.045 fruit set), (b) weevil exclusion (0.563 ± 0.050), and (c) ant exclusion (0.097 ± 0.047). To calculate the expected fecundity values for any given treatment, we used the reproductive effort (total number of flowers produced; given by the sum of the number of flowers, fruits and scars) of each plant during each census and the estimated fruit set according to each experimental treatment (see S2 for details). To incorporate the observed variation in fruit set for each treatment, we generated state-fate transition data frames with estimated fruit sets by drawing random numbers from a normal distribution based on the mean and standard deviation of the fruit set for each treatment, and replicated this 100 times per treatment per time-transition.
Reproductive success throughout the invasive range
To put our results in context, we performed rapid surveys in different sites/populations of S. plicata outside our demography study area to collect information on reproductive success (fruit set) following the same methods indicated above (see S5 for locations). In Puerto Rico, we re-analyzed data of the 4 m2 plots sampled in adjacent sites to our demographic study area in the Río Abajo State Forest in 2011 (during t 3, 100–500 m apart; Ackerman et al. 2014), and sampled three additional populations during 2012 found around El Yunque National Forest (east of the island, where S. plicata populations were first reported). Furthermore, we sampled populations in the Hawaiian Archipelago in 2010–2011 (Big Island, Kauai, Maui, and O’ahu islands), where S. plicata is invasive and orchid-specialist weevils (Orchidophilus sp.) are rare on these plants.
Statistical analyses
All statistical analyses were run with R (v. 3.2.0, R Core Team 2015). Mean values ±SD are reported, unless otherwise stated. We selected and performed parametric and non-parametric tests based on the distribution of the data. To determine if there were population growth rate differences between time transitions and treatments, we calculated the bootstrap distribution of λ by randomly sampling with replacement from the stage-fate data of observed transitions, and observed and modified fecundities—based on the exclusion experiments (reported as λboot) (Caswell 2001; Morris and Doak 2002; Stubben and Milligan 2007). To project the long-term effects of the acquired interactions with weevils and ants, we projected the three transition matrices derived from this study (field conditions), and the matrices with the different fecundity values calculated based on the exclusion experiments (100 matrices per treatment per time transition). We used the number of individuals at each of the six size-stages recorded in the first census (t 0; Table 1) as the initial population vector, and projected the matrices for 60 time steps, with demographic and environmental stochasticity. Because these projections are based on a stochastic population growth rate (λsto) rather than the asymptotic population growth rate (λ), we calculated the log stochastic population growth rate by Tuljapukar’s approximation (λT approx) and by simulation (λSim), and generated 95% CI for comparison from simulations using 5000 iterations (Morris and Doak 2002). Finally, we tested for differences between the reproductive success data from our demographic study and the other sites/populations, and visualized the data using the R package ‘yarrr’ (ND Phillips 2016, https://github.com/ndphillips/yarrr), which allows to plot the raw data together with the frequency distribution, mean, and the 95% Bayesian Highest Density Interval (HDI).
Results
Demography of Spathoglottis plicata
We marked a total of 281 individuals of S. plicata, and found a tendency for plants to increase in size (width of the longest leaf) over time (Table 1). The mean growth per year was 4 mm ± 16, however, when we compare the change in plant size (growth) between years, we found no significant differences (N = 555 for all available pairs, X2 = 1.51, df = 2, p = 0.47). During our study period, and based on analysis of the projection matrices derived from the observed field conditions, S. plicata was not at stable-stage distribution in our site. Intriguingly, there was a reduction in the number of individuals recruited to size categories 1–3 compared to previous and subsequent year at t 2 (Table 1).
The demographic parameters of the orchids exhibited variation throughout the study period (Table 2). Flowering probabilities ranged from 0.12 to 0.42 across years, and reproductive individuals produced a mean of 19–33 flowers per year. We found a strong positive correlation between reproductive effort and weevils damage, given by the sum of fruits with weevil oviposition and scars (Kendallτ = 0.84, p < 0.001). Fruit set ranged from 21 to 46%, and weevil damage to fruits reduced the effective fruit set (EFS) by even more than half in the first census (i.e. the number of fruits undamaged by weevils). The total numbers of recruits ranged from 15 to 28 per census, and between 9 and 13 plants died each year. The mean plant density per year was 9.3/m2, and ranged between 2 and 16 plants per m2 (for all plots and censuses combined). We found no indication of density-dependent recruitment during the three time transitions (Kendallτ, p = 0.70) when we compared the association between the density of each plot during the census at time t, and the number of new recruits found at time t + 1. Moreover, in all censuses following t 0, some individuals were not found, but were recorded alive when observed in subsequent years (Table 2).
Differences in the size-stage projection matrices for the individual time transitions reflect the population’s demographic variation over the years. Survival probability increases as the size-stage increases, and there was considerable variation in size-stage 1 (see S4A). The most noticeable difference is the survival of individuals in size-stage class 1 in t 0–t 1, in comparison to the other time transitions. Moreover, as size-stage increased, so did the mean flowering probability, as well as the mean production of flowers and fruits (see S4B–D). The population projection matrices derived from our field study reflected these temporal differences (Table 3). Overall, the average projection matrix showed that plants tended to remain in stasis or increase in size—especially to size stages ≥3, and that plants from size-stages 5 to 6 contributed a disproportionately higher fecundity (Fig. 3a). This was supported by the elasticity analyses (Table 3) whereby the average elasticity matrix indicated that survival and stasis of plants in size stages 3 and 5–6, along with transitions between the two larger size classes contributed the most to λ (Fig. 3b). Moreover, the elasticity analysis (and average elasticity matrix) suggested that while the influence of survival and growth on λ was stronger than that of fecundity, the effects of the latter were also greatest in the two largest size stages (Fig. 3b).
Are there temporal differences in population growth rates of the invasive orchids? Does damage to flowers and fruits by the native weevil S. polita drive these differences? Under field conditions, the 95% CI of λ showed no differences between years (Table 4A). However, λ values within transitions do respond to weevil effects. When weevils were excluded, λ was higher than that produced under field conditions. Still, the 95% CI of the transition t 1–t 2 include values of λ <1 for both field and weevil exclusion conditions.
Effects of acquired interspecific interactions on λ
When the three-way interactions with weevils and ants occur, the invasive orchid population at our site exhibited a positive λ both under observed fecundities in the field as well as in those estimated based on the control treatment of the exclusion experiments (Table 4B; based on sampling with replacement from the stage-fate data). Here, the ant exclusion treatment is not significantly different from the controls, while the weevil exclusion treatment significantly increases λ. The stochastic population growth rates, on the other hand, showed significant differences between treatment types: under field and control conditions λsto = 1.05, without ant protection λsto = 1.03, and without weevil damage λsto = 1.21 (Table 5). Similarly, when we projected populations over time, the exclusion of ants and weevils showed significantly different effects from field conditions and controls (Fig. 2). The projected abundance without ants after 60 time steps was negative (42% decrease) in relation to field conditions, whereas under weevil-free conditions it was positive (367 times greater).

Visual representation of the average population projection- (a) and elasticity (b) matrices of Spathoglottis plicata based on four continuous years of demographic data in the Río Abajo Forest, where the invasive orchid acquired novel interactions with weevils and ants. The average matrices reflect the effects of the acquired interactions on the population dynamics of S. plicata under natural field conditions during the study period
Reproductive success throughout the invasive range
We found and sampled populations of S. plicata in different habitat types at different elevations, with presence or absence of antagonist weevils (Fig. 4; see S5 for locations and summary statistics). For reproductive plants, despite finding significant differences in terms of plant size (Kruskall-Wallis X 2 = 213.46, df = 8, p < 0.01), the mean reproductive effort was similar among all populations (Pairwise Wilcoxon with Bonferroni–Holm correction p > 0.06, for all pairs) (see Hochberg 1988 for details on Bonferroni-Holm correction). Moreover, reproductive success data from populations of the Philippine Ground Orchid throughout its invasive range shows a clear distinction with regards to the presence or absence of weevils (Wilcoxon rank sum test W = 13,680, p < 0.01). In Puerto Rico, when weevils are present, the mean fruit set was 0.31 ± 0.30, showing high variability. If fruits with weevil damage are excluded (effective fruit set), the mean reproductive success is reduced further to 0.19 ± 0.33. Moreover, damage to flowers and fruits by weevils in these populations seem to skew the reproductive success of invasive orchids towards the lower end of the distribution (Fig. 4). On the other hand, weevil-free populations in Puerto Rico and Hawai’i had a considerably higher mean reproductive success of 0.59 ± 0.30. These results were consistent even when we excluded the four years of data from our demography plots.
Discussion
The effects of novel interspecific interactions on introduced species may span from negative, neutral, to positive, and may involve both native and non-native species. Kelly and Dyer (2002) suggested that life cycle based models incorporating demographic and environmental stochasticity would provide a better understanding of the population dynamics. Nevertheless, studies looking at the effects of novel interspecific interactions on introduced species seldom use population performance metrics. Here, we combined field demographic and fecundity data, as well as the results of the effects of exclusion experiments on fecundity to construct life-cycle based population models to quantify and assess the role of acquired interspecific interactions on the population performance (λ) of an invasive plant.
Demography of Spathoglottis plicata
As expected, our analyses showed that the Philippine Ground Orchid exhibits λ > 1 under natural conditions, despite acquiring antagonistic interactions with a native herbivorous weevil. Nevertheless, we found values of λ ≤ 1 during one of the census periods (t 1–t 2), even when fecundities were modified to reflect the exclusion of weevils. Elasticity analyses of the projection matrices for all the time intervals, with fecundities under field conditions, showed that small changes in the survival and size-stage transitions, and fecundities of size-stages 5–6, would have a greater effect on λ (see Table 3; Fig. 2b). Furthermore, when compared to the other time intervals, the recruitment rate per plot was 2–7% lower in t 2, and the overall survival rate of this time interval was also lower, especially for size-stage 1. This dramatically reduced the number of individuals in size-stages 2 and 3 on t 2 by approximately 77 and 19% respectively, which probably affected the recruitment to larger size-stages in subsequent years. The fact that there was a 12% increase of individuals in size-stages 1–3 recorded the next year (t 3) suggests that the reduction in the survival rate of size-stage 1 (during t 1–t 2) was the main cause of population decline.
What caused the reduction in survival rate for size-stage 1 during the t 1–t 2 time interval? While other factors may have been involved, we suspect that unusual weather was a major cause. Climatological data from the nearby Arecibo Observatory (18.349°N, 66.752°W; National Climatic Data Center; www.ncdc.noaa.gov) showed that total precipitation was above normal during the timespan of our study period. However, the precipitation during t 1–t 2 was not only much higher than the other periods, but it was also the most variable (t 0–t 1: 2176 mm ± 72; t 1–t 2: 2510 mm ± 110; t 2–t 3: 2348 mm ± 89). In other words, plants experienced very high precipitation events over short, intermittent periods of time during t 1–t 2 when compared to the other time transitions.
Although higher precipitation can be associated with increased recruitment rates in orchids (Zotz and Schmidt 2006; Diez 2007), the fact that the Río Abajo State Forest is a Subtropical Moist Forest suggests that water is not often a limiting factor. Excessive soil water moisture is known to cause increased mortality in cultured orchids and other plants due to susceptibility to disease and changes in the soil nutrient concentration and pH (Leonhardt and Sewake 1999). Two other mechanisms that could explain this are competitive exclusion of small individuals due to increased growth in larger plants or the disruption of mycorrhizal interactions. It should be noted that we are just beginning to understand how mycorrhizal associations affect the recruitment and survival of wild orchids (e.g. Bayman et al. 2002).
While our focus has been on biotic interactions with insects, fluctuations in weather certainly can play a role in the demography of orchid populations (Pfeifer et al. 2006). Our analysis showed that environmental fluctuations may result in uncertain survival and fecundity rates in smaller size-stages, that there is a trade-off between growth and reproduction (i.e. energy allocated to growth reduces reproductive success), and that fecundities from size-stages 5 to 6, although variable, are important vital rates contributing to population growth. While one may argue that our results are perhaps artifacts of the matrix, they are actually expected outcomes for iteroparous species with indeterminate growth inhabiting stochastic environments (Katsukawa et al. 2002), including invasive plants (Ramula et al. 2008). These traits and conditions are just what S. plicata possesses and experiences.
Effects of acquired interspecific interactions on λ
Biotic interactions are important factors determining the invasion success of exotic species (Mitchell et al. 2006), and the effects of plant–animal interactions, even at small time frames, may have long-term effects on plant populations (Strauss and Irwin 2004). When we projected the population of S. plicata over time assuming field conditions (all interactions included) and stochasticity, the average abundances predicted by the model increase as expected for an invasive species expanding its range. The model including demographic and environmental stochasticity showed a lag phase of ~20–30 time-steps (equivalent to years in our case; Fig. 2). While we have found that the ants and weevils interact with S. plicata throughout most of its distribution in Puerto Rico, the actual population growth curve in a given location is likely to be somewhere between weevil exclusion treatment and field and control conditions because of (1) resource discovery time lag of weevils (supported by the low frequencies of weevils on isolated plants and plant patches), and (2) a presumed weevil population growth lag-period. These processes may be linked to the estimated invasion lag phase recorded in Puerto Rico for this orchid (assuming an overall equal herbarium collection effort per decade, which is likely the case; see “Study system” section).
Here we showed that weevil-mediated biotic resistance proposed for S. plicata invasion in Puerto Rico (Ackerman et al. 2014) does indeed have demographic consequences. When weevils are excluded from inflorescences of S. plicata, λ significantly increased to 1.20, which underscores how detrimental S. polita can be to the population growth rates of this invasive orchid. Propagule pressure, the number or sets of individuals that are dispersed to new area, is an important factor in determining invasion success and invasion speed (e.g. D’Antonio et al. 2001; Von Holle and Simberloff 2005; Richardson and Pysek 2006). Although S. plicata produces thousands of seeds in each fruit, weevils dramatically reduce the number of mature fruits, thereby reducing propagule pressure. Without weevils, the expected population size is not only (on average) 367 times higher than projected with the influence of weevils, but the model also indicates a significantly higher stochastic population growth rate (λsto). Larger individuals of S. plicata in our study were associated with a higher reproductive effort, and the proportion of weevil damage to flowers and fruits was in turn positively correlated with reproductive effort. This is congruent with the results of the elasticity analyses, which revealed that only changes in the fecundities of the two larger size-stage classes had a substantial effect on λ (Fig. 2b). According to a meta-analysis of biotic resistance to exotic plant invasions, there is little evidence that plant–native herbivore interactions repelled invasions (Levine et al. 2004). Instead, herbivores limited the abundance of invasive plants once they become established (Levine et al. 2004). Similarly, we found support for biotic resistance imposed by native weevils, but this resistance is incapable of deterring the S. plicata invasion since weevils alone do not reduce λ ≤ 1, suggesting that propagule pressure is an important mechanism at play.
The invasional meltdown hypothesis predicts that biotic interactions between invaders will have positive synergistic effects, thus increasing the chances of success for both interacting species (Simberloff and Von Holle 1999). Is the Spathoglottis plicata–Solenopsis invicta interaction an example of invasional meltdown? Similar to the results of the ant exclusion experiments where the mean fruit set was reduced by 33%, our bootstrapped values of λ showed that although invasive ants tend to reduce λ, this reduction was not significant, suggesting they do not effectively protect the invasive orchids. Yet this modelling approach does not consider the observed variation of fruit set, and the effects of stochasticity (which is substantial in our system). Although the experimental exclusion of ants in particular did not showed statistically significant protection to the invasive orchid, the overall evidence from previous studies indicated that red fire ants are an important factor affecting positively the reproductive success of S. plicata, with fruit set increasing with increasing number of ants visiting the inflorescences (Ackerman et al. 2014). Ants are known to respond to the quality and quantity of resources (Mailleux et al. 2000; Schilman 2011), and presence of invasive red fire ants and abundance on S. plicata was quite variable within days (mornings and afternoons), and between weeks in sites close to our demography study site (Ackerman et al. 2014). When we included demographic and environmental stochasticity in our analyses, the absence of ant protection decreased the expected population size over time almost by half (compared to field conditions; Fig. 3), and the λsto values were significantly lower than those expected under both the control treatment and field conditions.

The effects of acquired interspecific interactions on the projected population size of the invasive orchid (Spathoglottis plicata) over time. The population estimates were calculated by simulation (with 5000 iterations) based on the three projection matrices derived from the observed field conditions, and from the 100 matrices generated from each of the ex situ exclusion treatment (control, ant exclusion, and weevil exclusion; see methods). The initial population vector was based on the number of individuals at each size-stage observed at t 0 (N = 209)
It is possible that the lack of statistically significant ant protection to invasive orchids based on exclusion experiments was due to the sample size, and the highly variable reproductive success when both weevils and ants have the potential to interact with the orchid. We believe that considering the variation associated to each exclusion treatment (rather than static mean values) and including stochasticity enabled the model to better capture the dynamics in the system, yielding results that are consistent with the overall evidence on Solenopsis invicta protection against weevils, and with protection of other plants throughout its invasive range (reviewed in Ness and Bronstein 2004). Furthermore, our results are consistent with the findings of Rico-Gray and Thien (1989), who had showed that (1) different species of ants foraging on extra-floral nectaries of Schomburgkia tibicinis in Mexico deterred the main herbivore, Stethobaris sp., (2) that the prevalence of a given species varied with time, and (3) that the reproductive success of the orchid increased with increasing ant size (which was also related to species foraging dominance). Taken together, our results partially support the invasional meltdown hypothesis: invasive ants and invasive orchids are engaged in a mutually beneficial interaction that results in increased population growth for the orchids, and presumably fitness gains for ants (from energy derived from extrafloral nectar; Ness and Bronstein 2004, and references therein; Ackerman et al. 2014).
How prevalent are these interactions with ants across the invaded range? Plant–ant mutualisms, where ants protect the plants from herbivores and receive a reward from the plant, have been described or assumed for many plants (Rico-Gray and Thien 1989; Bronstein 1998; Chamberlain and Holland 2009). These interactions can be facultative and very generalized, not only for invasive species that have them, but for orchids as well (Fisher and Zimmerman 1988; Rico-Gray and Thien 1989; van der Cingel 2001). In Puerto Rico, there are at least seven native species of red fire ants (Solenopsis), and forest species may rely heavily on liquid foods (e.g. nectar, honeydew) (Torres 1984). Native S. geminata can be found in both disturbed, and relatively mature lowland dry to montane wet forests (50–600 m asl), and while it is considered invasive in Hawai’i (along with S. papuana), it can be displaced by invasive S. invicta when they co-occur (Porter et al. 1988). We have observed unidentified Solenopsis ants visiting the invasive orchid across the invaded range in Puerto Rico and Hawai’i, including relatively anthropogenically-undisturbed riparian areas inside mature forests (we only identified ants to the species level in our study site). Many other species of ants visit the inflorescences of the invasive orchid, and although there are several potentially aggressive species that specialize in liquid foods in Puerto Rico (Torres 1984), the only other species (very rarely) observed attacking and deterring weevils in our study area was the native Wasmannia auropunctata. This species can be found in mature forests from sea level up to 2400 m, is an aggressive invader elsewhere, and has been shown to increase the reproductive success of some plants (Ness and Bronstein 2004). Given the promiscuity of extrafloral nectaries-bearing plant and ant associations, and that many potential ant partners exist across Puerto Rico, we expect facultative mutualistic Spathoglottis-ant associations to be prevalent, but variable and governed by the orchid’s residence time, plant (inflorescence) densities, disturbance regimes, as well as by physiological and behavioral aspects of ant species.
There are some limitations to this study, which need to be considered. Firstly, when evaluating the effects of interspecific interactions based on experimental exclusions, the calculated fecundities represent the relative effect of different treatments on population growth and projected abundances, rather than precise estimates, which limits the characterization of populations elsewhere. Secondly, our models lack the seed stage in the life cycle of S. plicata. However, we consider highly unlikely that there is an active seed bank that contributes to the population growth rate, especially because this is a tropical species (e.g. Batty et al. 2000). Also, S. plicata produces inflorescences throughout the year, yet we only gathered reproductive data during the census periods (i.e. once a year). Thus, we are assuming that this reproductive “snapshot” is proportional to- and represents the annual contribution of a given size-stage to fecundity. Moreover, our models assume that new recruits are the progeny of plants within the study plots. Seed dispersal may affect this assumption and produce biases in the sense that (1) fecundity could be overestimated if new recruits are from plants outside the study plots and (2) the fecundity could be underestimated if the plants within the study plots produce new recruits outside the plots. However, we believe that their overall effects are minimal because studies on orchid seed dispersal and seedling establishment have shown that despite having the potential for long distance dispersal (which is inherently important for range expansion), a high proportion of seeds fall in the vicinity of parent plants, and sharply decreases with distance (e.g. Ackerman et al. 1996; Murren and Ellison 1998; Jersáková and Malinova 2007). In addition, demographic studies that have excluded the seed stages of orchids have successfully predicted future populations sizes (Tremblay and Hutchings 2003; Schödelbauerová et al. 2009).
Despite these limitations, the demographic consequences of interactions with native weevils and ants on the reproductive success of the invasive orchid observed in our demographic study seems to capture the variation observed throughout the invaded range in Puerto Rico (Fig. 4). Moreover, both the weevil-free population found in PR, as well as those in Hawai’i, have fruit sets close to that of plants in which weevils were experimentally excluded. Considering this, we expect the dynamics we revealed to be generally applicable across Puerto Rico because actively invading species usually have a λ > 1, and most S. plicata populations that we have surveyed in Puerto Rico have both ants and weevils. Because populations of S. plicata in our study area have been present for at least 15 years, the results derived from field conditions probably do not reflect what’s occurring at the invasion front. We expect reproductive success (and λs) in weevil-free and newly established populations to be similar to that of our weevil exclusion treatments, as observed in PDE (where weevils were absent). In agreement with the findings of Ramula et al. (2008) for invasive plant populations, and assuming populations of S. plicata across Puerto Rico exhibit similar dynamics to those in our study, simultaneous reductions in either survival or growth in combination with targeted reduction of fecundity for the two larger size-stages would be necessary to reduce λ.

Weevil image by Tom Murray (modified and used with authorization)
Reproductive success (fruit set) of Spathoglottis plicata throughout its invasive range in Puerto Rico and Hawai’i in the presence or absence of herbivorous weevils. Points represent raw data (individual reproductive plants sampled), the frequency distribution of observations is denoted by the shape of the beans, the solid line represents the mean, and the grey bar represents the 95% Bayesian Highest Density Interval. Data from the Río Abajo State Forest population include fruit set from the demographic censuses per year, ABO(t0–t3), and two sites located 80–500 m, ABO(out), sampled at t 3. Data collected in El Yunque National Forest (northeast of PR) include three populations: EYQ, EVE and PDE—with the latter being weevil-free. Hawaiian populations, where weevils are also absent, were sampled in Maui (MAU), Kauai (KAU), the Big Island (BIS), and O’ahu (OAU).
Studies aiming to assess whether biotic interactions can affect the invasion success and/or management of introduced species should include multiple (and replicated) independent populations representing the different ecological conditions which the species experiences throughout the invasive range where (1) demographic censuses are conducted for at least three years (and designed based on the life-cycle of the species), (2) exclusion experiments are conducted preferably within the demographic monitoring timeframe and area, and (3) additional rapid surveys are conducted in additional areas to maximize capturing the variation present in the invaded range of the species.
To finalize, the fate and invasion success of non-native introduced species is ultimately determined by processes occurring at the population level, and their populations do not occur in a vacuum. As a result, focusing on single species populations while ignoring the array of interactions that they encounter in their introduced range hinders our understanding of biological invasions. Here we showed how the combination of field, experimental and matrix population modelling approaches provide a quantitative framework to evaluate the effects of multiple biotic interactions on the performance of invasive plants at the population level. The implementation of approaches such as this not only will contribute to our understanding of biological invasions, but it can also serve to explicitly identify critical life-stages to target for management and assess the efficacy of different management strategies when interactions with opposing outcomes are at play (Sakai et al. 2001; D’Antonio and Meyerson 2002; Kelly and Dyer 2002; Allendorf and Lundquist 2003).
Change history
25 November 2016
An Erratum to this paper has been published: https://doi.org/10.1007/s10530-016-1337-5
References
Ackerman JD (1995) An orchid flora of Puerto Rico and the Virgin Islands. Mem N Y Bot Gard 73:1–201
Ackerman JD (2007) Invasive orchids: weeds we hate to love. Lankesteriana 7:19–21. doi:10.15517/lank.v7i1-2.18386
Ackerman JD, Sabat A, Zimmerman JK (1996) Seedling establishment in an epiphytic orchid: an experimental study of seed limitation. Oecologia 106:192–198. doi:10.1007/BF00328598
Ackerman JD, Falcón W, Recart W (2011) New records of naturalized orchids for the Hawaiian Islands. Bish Mus Occas Pap 110:1–4
Ackerman JD, Falcón W, Molinari J et al (2014) Biotic resistance and invasional meltdown: consequences of acquired interspecific interactions for an invasive orchid, Spathoglottis plicata in Puerto Rico. Biol Invasions 16:2435–2447. doi:10.1007/s10530-014-0676-3
Allendorf FW, Lundquist LL (2003) Introduction: population biology, evolution, and control of invasive species. Conserv Biol 17:24–30. doi:10.1046/j.1523-1739.2003.02365.x
Axelrod FS (2011) A systematic vademecum to the vascular plants of Puerto Rico. BRIT Press, Fort Worth
Batty AL, Dixon KW, Sivasithamparam K (2000) Soil seed-bank dynamics of terrestrial orchids. Lindleyana 15:227–236
Bayman P, Gonzalez EJ, Fumero JJ, Tremblay RL (2002) Are fungi necessary? How fungicides affect growth and survival of the orchid Lepanthes rupestris in the field. J Ecol 90:1002–1008
Bronstein JL (1998) The contribution of ant-plant protection studies to our understanding of mutualism. Biotropica 30:150–161. doi:10.1111/j.1744-7429.1998.tb00050.x
Caswell H (2001) Matrix population models. Sinauer Associates Incorporated, Sunderland
Chamberlain SA, Holland JN (2009) Quantitative synthesis of context dependency in ant–plant protection mutualisms. Ecology 90:2384–2392. doi:10.1890/08-1490.1
D’Antonio C, Meyerson LA (2002) Exotic plant species as problems and solutions in ecological restoration: a synthesis. Restor Ecol 10:703–713. doi:10.1046/j.1526-100X.2002.01051.x
D’Antonio CD, Levine J, Thomsen M (2001) Ecosystem resistance to invasion and the role of propagule supply: a California perspective. J Mediterr Ecol 2:233–245
Davis LR, Vander Meer RK, Porter SD (2001) Red imported fire ants expand their range across the West Indies. Fla Entomol 84:735–736. doi:10.2307/3496416
Diez JM (2007) Hierarchical patterns of symbiotic orchid germination linked to adult proximity and environmental gradients. J Ecol 95:159–170. doi:10.1111/j.1365-2745.2006.01194.x
Elton CC (1977) The invaders. The ecology of invasions by animals and plants. Springer, Dordrecht, pp 15–32
Ewel JJ, Whitmore JL (1973) The ecological life zones of Puerto Rico and the U.S. Virgin Islands. Forest Service Research Paper ITF-18. Institute of Tropical Forestry, Río Piedras, Puerto Rico
Fieberg J (2004) Population viability analysis. J Biogeogr 31:515–516. doi:10.1046/j.0305-0270.2003.01027.x
Fisher BL, Zimmerman JK (1988) Ant/orchid associations in the Barro Colorado national monument, Panama. Lindleyana 3:12–16
Hastings A (1997) Population biology: models and concepts. Springer, New York
Hochberg Y (1988) A sharper Bonferroni procedure for multiple tests of significance. Biometrika 75:800–802. doi:10.1093/biomet/75.4.800
Holle BV, Simberloff D (2005) Ecological resistance to biological invasion overwhelmed by propagule pressure. Ecology 86:3212–3218. doi:10.1890/05-0427
Horvitz CC, Schemske DW (1995) Spatiotemporal variation in demographic transitions of a tropical understory herb: projection matrix analysis. Ecol Monogr 65:155. doi:10.2307/2937136
Jersáková J, Malinova T (2007) Spatial aspects of seed dispersal and seedling recruitment in orchids. New Phytol 176:237–241. doi:10.1111/j.1469-8137.2007.02223.x
Jolliffe K (2010) Epiphytic orchids of the Seychelles. Kapisen 10:6–8
Katsukawa Y, Katsukawa T, Matsuda H (2002) Indeterminate growth is selected by a trade-off between high fecundity and risk avoidance in stochastic environments. Popul Ecol 44:265–272. doi:10.1007/s101440200030
Kearns CA, Inouye DW (1993) Techniques for pollination biologists. University Press of Colorado, Boulder
Kelly C, Dyer R (2002) Demographic consequences of inflorescence-feeding insects for Liatris cylindracea, an iteroparous perennial. Oecologia 132:350–360. doi:10.1007/s00442-002-0948-y
King JR, Tschinkel WR (2008) Experimental evidence that human impacts drive fire ant invasions and ecological change. Proc Natl Acad Sci USA 105:20339–20343. doi:10.1073/pnas.0809423105
Kores P (1980) Pollination mechanisms as a limiting factor in the development of the Orchidaceous Flora of Hawai’i. Department of Botany, University of Hawai’i at Manoa
Lach L, Tillberg CV, Suarez AV (2010) Contrasting effects of an invasive ant on a native and an invasive plant. Biol Invasions 12:3123–3133. doi:10.1007/s10530-010-9703-1
Lefkovitch LP (1965) The study of population growth in organisms grouped by stages. Biometrics 21:1–18. doi:10.2307/2528348
Leonhardt K, Sewake K (eds) (1999) Growing Dendrobium orchids in Hawaii. College of Tropical Agriculture and Human Resources, University of Hawaii, Honolulu
Levine JM, D’Antonio CM (1999) Elton revisited: a review of evidence linking diversity and invasibility. Oikos 87:15. doi:10.2307/3546992
Levine JM, Adler PB, Yelenik SG (2004) A meta-analysis of biotic resistance to exotic plant invasions. Ecol Lett 7:975–989. doi:10.1111/j.1461-0248.2004.00657.x
Mailleux A-C, Deneubourg J-L, Detrain C (2000) How do ants assess food volume? Anim Behav 59:1061–1069. doi:10.1006/anbe.2000.1396
McCullough DR, Work TT, Cavey JF et al (2006) Interception of nonindigenous plant pests at US ports of entry and border crossing over a 17-year period. Biol Invasions 8:611–630. doi:10.1007/s10530-005-1798-4
Mitchell CE, Agrawal AA, Bever JD et al (2006) Biotic interactions and plant invasions. Ecol Lett 9:726–740. doi:10.1111/j.1461-0248.2006.00908.x
Moloney KA (1986) A generalized algorithm for determining category size. Oecologia 69:176–180. doi:10.1007/BF00377618
Montalvo AM, Ackerman JD (1987) Limitations to fruit production in Ionopsis utricularioides (Orchidaceae). Biotropica 19:24–31
Morris WF, Doak DF (2002) Quantitative conservation biology. Sinauer Associates Inc, Sunderland
Murren CJ, Ellison AM (1998) Seed dispersal characteristics of Brassavola nodosa (Orchidaceae). Am J Bot 85:675–680
Ness JH, Bronstein JL (2004) The effects of invasive ants on prospective ant mutualists. Biol Invasions 6:445–461
Parker JD, Hay ME (2005) Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecol Lett 8:959–967. doi:10.1111/j.1461-0248.2005.00799.x
Pfeifer M, Wiegand K, Heinrich W, Jetschke G (2006) Long-term demographic fluctuations in an orchid species driven by weather: implications for conservation planning. J Appl Ecol 43:313–324. doi:10.1111/j.1365-2664.2006.01148.x
Porter SD, Savignano DA (1990) Invasion of polygyne fire ants decimates native ants and disrupts arthropod community. Ecology 71:2095. doi:10.2307/1938623
Porter SD, Van Eimeren B, Gilbert LE (1988) Invasion of red imported fire ants (Hymenoptera: Formicidae): microgeography of competitive replacement. Ann Entomol Soc Am 81:913–918
Prakash N, Leelee A (1973) Life-history of a common Malaysian orchid Spathoglottis plicata. Phytomorphology 23:9–17
R Core Team (2015) R: a language and environment for statistical computing
Ramula S, Knight TM, Burns JH, Buckley YM (2008) General guidelines for invasive plant management based on comparative demography of invasive and native plant populations. J Appl Ecol 45:1124–1133
Recart W, Ackerman JD, Cuevas AA (2013) There goes the neighborhood: apparent competition between invasive and native orchids mediated by a specialist florivorous weevil. Biol Invasions 15:283–293. doi:10.1007/s10530-012-0283-0
Richardson DM, Pysek P (2006) Plant invasions: merging the concepts of species invasiveness and community invasibility. Prog Phys Geogr 30:409–431. doi:10.1191/0309133306pp490pr
Richardson DM, Allsopp N, D’Antonio CM et al (2007) Plant invasions—the role of mutualisms. Biol Rev 75:65–93. doi:10.1111/j.1469-185X.1999.tb00041.x
Rico-Gray V, Thien LB (1989) Effect of different ant species on reproductive fitness of Schomburgkia tibicinis (Orchidaceae). Oecologia 81:487–489
Rosenberg K, Boland J, Tiver F, Watson I (2005) Application of the Vandermeer-Moloney Algorithm for determination of category size to Australian native plants. MODSIM 05 International Conference on Modelling and Simulation, December 2005. University of Melbourne, Modelling and Simulation Society of Australia and New Zealand
Sakai AK, Allendorf FW, Holt JS et al (2001) The population biology of invasive species. Annu Rev Ecol Syst 32:305–332. doi:10.1146/annurev.ecolsys.32.081501.114037
Schilman PE (2011) Trail-laying behaviour as a function of resource quality in the ant Camponotus rufipes. Psyche 2011:1–5. doi:10.1155/2011/139385
Schödelbauerová I, Tremblay RL, Kindlmann P (2009) Prediction vs. reality: can a PVA model predict population persistence 13 years later? Biodivers Conserv 19:637–650. doi:10.1007/s10531-009-9724-1
Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: invasional meltdown? Biol Invasions 1:21–32. doi:10.1023/A:1010086329619
Smith MR (1936) The ants of Puerto Rico. J Agric Univ P R 20:819–875
Strauss SY, Irwin RE (2004) Ecological and evolutionary consequences of multispecies plant-animal interactions. Annu Rev Ecol Evol Syst 35:435–466. doi:10.1146/annurev.ecolsys.35.112202.130215
Stubben C, Milligan B (2007) Estimating and analyzing demographic models using the popbio package in R. J Stat Softw 22:1–23. doi:10.18637/jss.v022.i11
Thompson SA, Wright FW (1995) Spathoglottis plicata (Orchidaceae): new to Dominica, another record from the Lesser Antilles. Caribb J Sci 31:2
Torres JA (1984) Niches and coexistence of ant communities in Puerto Rico: repeated patterns. Biotropica 16:284–295
Tremblay RL, Hutchings MJ (2003) Population dynamics in orchid conservation: a review of analytical methods based on the rare species Lepanthes eltoroensis. In: Dixon KW, Kell SP, Barret RL, Cribb PJ (eds) Orchid conservation. Natural History Publications, Kota Kinabalu
Van Der Cingel NA (2001) An atlas of orchid pollination: European orchids. CRC Press, Boca Raton
Vandermeer J (1978) Choosing category size in a stage projection matrix. Oecologia 32:79–84. doi:10.1007/BF00344691
Wetterer JK, Snelling RR (2006) The red imported fire ant, Solenopsis invicta, in the Virgin Islands (Hymenoptera: Formicidae). Fla Entomol 89:431–434
Zotz G, Schmidt G (2006) Population decline in the epiphytic orchid Aspasia principissa. Biol Conserv 129:82–90. doi:10.1016/j.biocon.2005.07.022
Acknowledgements
The PR-LSAMP Bridge to the Doctorate Fellowship (HRD-0601843), and the Center for Applied Tropical Ecology and Conservation (HRD-0734826) provided economical and logistical support to this project, for which we are grateful. We also want to thank Rafael D. Rodríguez, Joham Ríos, Taisa Alicea, Gilberto Colón, Rebecca Hernández, Wilnelia Recart and Jonathan Molinary for their help the during the many hours of fieldwork. Drs. Riccardo Papa and Ingi Agnarsson provided insightful comments and suggestion to do this project, for which we are grateful. We also thank Caner Aktas for his much appreciated help with part of the R code. Finally, we are very grateful to the anonymous reviewers, and the editors, whose comments and suggestions greatly improved this article.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Falcón, W., Ackerman, J.D. & Tremblay, R.L. Quantifying how acquired interactions with native and invasive insects influence population growth rates of a non-indigenous plant. Biol Invasions 19, 895–911 (2017). https://doi.org/10.1007/s10530-016-1318-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-016-1318-8