
Abstract
Endophytic fungi in grasses are often considered to be mutualistic because they can increase host resistance to herbivory and drought. However, not all endophytes are beneficial to their hosts, but may instead be specialist enemies. Brachypodium sylvaticum is an invasive grass in the USA. In its European native range, it is nearly always infected by the host-specific endophyte Epichloë sylvatica. While this fungus decreases herbivory, it also decreases the growth rate and size of infected plants, making them less competitive. After showing that B. sylvaticum has lost its endophyte in the invaded range, we use greenhouse assays to deconfound the effects of endophyte infection and range origin to test assumption of the evolution of increased competitive abilities (EICA) hypothesis. Brachypodium in its invaded range appears to have lost tolerance mechanisms present in the native range, allowing Epichloë to greatly increase seedling mortality and reduce growth rates. Additionally, there is some evidence for increased competitive abilities in the form of increased seedling growth rates in the invasive range. Together, these results provide strong support of the EICA hypothesis.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The enemy release hypothesis (ERH) postulates that one major factor facilitating invasion is the relative lack of specialized enemies in the invaded range, allowing for faster growth and spread (Keane and Crawley 2002). An alternative, the evolution of increased competitive ability hypothesis (EICA), assumes that the success of invasive species is evolutionary in nature, driven by the change in selective pressures of the new environment. Under the EICA framework, enemy release provides selective pressure to reallocate resources from defense to growth and reproduction (Blossey and Notzold 1995), though there are many complicating factors (Colautti et al. 2004). One complication is the distinct difference in defensive strategies used to protect against generalist versus specialist enemies (van der Meijden 1996; Müller-Schärer et al. 2004). Specialist enemies are theorized to be more important to plant invasions (Keane and Crawley 2002) because of the disproportionate effect they have on controlling populations in their native ranges, and the relative metabolic costliness of specific defenses. Thus, evolved increased competitive abilities may be due to reallocation of resources from specialist defenses to generalist defenses (Joshi and Vrieling 2005).
There has been much debate and intensive research surrounding the EICA hypothesis (reviewed in: Atwood and Meyerson 2011; Felker-Quinn et al. 2013), much of it inconclusive or contradictory (e.g., Willis et al. 2000; van Kleunen and Schmid 2003; Bossdorf et al. 2004; but see also Vilà et al. 2003; Meyer et al. 2005). Founder effects, novel hybridization opportunities, and strong abiotic environmental selective pressures can drive rapid evolutionary change in invasion. This complicates EICA research, and many studies have not tested competitive abilities and defense in the same organisms (see Bossdorf et al. 2005; Atwood and Meyerson 2011 and citations therein). It is impossible to make inferences about energetic tradeoff without a measure of both competitive abilities (usually growth) and defense. Here, we test the EICA hypothesis using Brachypodium sylvaticum (Huds.) P. Beauv., an aggressive invasive species in the northwest of the USA (Roy 2010), with the ERH (phenotypic plasticity in the face of specialist enemy loss) as an explicit alternative hypothesis. Previous work indicates that pathogens and herbivores of B. sylvaticum show some, but not all, of the characteristics predicted by the ERH (Roy et al. 2011). Insecticide and fungicide sprays were used to remove herbivorous insects and pathogenic fungi from the plants in multiple populations in both the native and invaded ranges. In accordance with the ERH, population growth rates were higher in the native range in the sprayed plots, where enemies were fewer than in the control plots. There was no statistically significant effect of enemy removal in the invaded range. Contrary to the ERH, all the common enemies were generalists and there was more herbivory in the invaded range relative to the native range (Roy et al. 2011; Halbritter et al. 2012).
Increased herbivory in the invaded range and reduced seed germination in the native range suggested that there might be differences in endophyte infection, since endophytes would not have been killed by the non-systemic fungicides used in previous studies (Roy et al. 2011; Halbritter et al. 2012). Endophytes are fungi that live between the cell walls of plants and cause no visible disease symptoms on the surface of the plant; they are common in grasses (Clay 1990; Rudgers et al. 2009). While it is not obvious that a plant is infected when endophytes are present, they may nonetheless have a range of consequences for their hosts, from true mutualism that increases insect or drought resistance, through commensalism, to antagonist pathogenicity that decreases survival and reproduction (Carroll 1988; Faeth and Sullivan 2003; Saikkonen et al. 2006). The same species of endophyte can either be a mutualist or pathogen depending upon its lifecycle stage, genotype, or environmental conditions. All symbioses exist on a continuum from pathogen to mutualist: if the benefit to the host (e.g., from reduced herbivory) is greater than the cost (e.g., reduced growth and seed-set) the fungus is a mutualist. The environment within which a host is embedded will impact the position of a symbiont along this continuum because it will alter the balance between costs and benefits.
Brachypodium sylvaticum in its native Eurasian range appears to be almost ubiquitously infected with a host-specific fungal endophyte, Epichloë sylvatica Leuchtm and Schardl (Eckblad and Torkelsen 1989; Raynal 1994; Väre and Itämies 1995; Bucheli and Leuchtmann 1996; Enomoto et al. 1998; Zabalgogeazcoa et al. 2000; Roy et al. 2011; Leuchtmann, pers. com.), which may act as a pathogen rather than a mutualist, despite common assumptions about Epichloë endophytes of grasses (Schardl 1996). Small-scale studies done with infected and uninfected plants in Switzerland, by Brem and Leuchtmann (2002), indicate that while plants infected with an asexual strain of E. sylvatica have less herbivory, they also have decreased growth rates and competitive abilities. Recent research in our lab (Roy et al. 2011; Halbritter et al. 2012) suggests that Epichloë infection may decrease germination rates in B. sylvaticum by seed infection, as the fungus is spread vertically from mother plant to daughter (Brem and Leuchtmann 1999). Thus, E. sylvatica appears to be a specialist enemy of B. sylvaticum.
Here, we document the near total absence of E. sylvatica infection in the invaded range. If E. sylvatica is generally pathogenic, the near lack of fungal endophyte infection in B. sylvaticum within the invaded range may constitute strong support for some form of the ERH or the EICA hypothesis.
In addition to documenting the virtual absence of the endophyte in the invaded range, we compare germination and growth rates of seedlings, a common proxy for fitness (Poorter and Garnier 1999; Matzek 2012), from the native and invaded ranges to explicitly test the EICA hypothesis. We utilized a greenhouse experiment using seeds collected during the same season in both ranges, clearing the seeds of Epichloë infection and then selectively re-inoculating half of each group. This bifactorial design permits us to effectively compare the effect of E. sylvatica and plant origin independent of each other, allowing us to distinguish between ERH, EICA, and potential founder effects (Fig. 1). If release of B. sylvaticum from control by E. sylvatica is sufficient to explain observed increases in fitness in the invasive range (Holmes et al. 2010; Roy et al. 2011; Halbritter et al. 2012), we expect that removing it from native range plants should increase their performance to be on par with those from the invasive range. Additionally, we would expect invasive range plants to be affected similarly to those from the native range. In short, under the hypothesis that E. sylvatica is directly impacting fitness in B. sylvaticum, such that it is a controlling specialist enemy in the native range, we expect the fungus to impact native and invasive range plants similarly (Fig. 1a).

Comparison of predictions of the ERH (a), the EICA hypothesis (b), and founder effects or other evolutionary forces unrelated to enemy release (c). Relative sizes of the cartoon grasses and the green bars indicate relative differences in fitness for the hosts. Presence (E+) or absence (E−) of E. sylvatica is shown on the X-axis, and range of origin on the Y-axis of each panel. ERH (a): If the observed increase in fitness in B. sylvaticum is due only to escape from E. sylvatica, re-introduction of the fungus to invasive range plants should recapitulate native range fitness levels, while removal of the fungus from native range grasses should release them from control, increasing fitness to levels to those observed in invasive range plants. EICA (b): If populations of B. sylvaticum have evolved in response to enemy release in the invasive range, we expect invasive range plants to experience a disproportionate fitness loss when infected by E. sylvatica compared to their native range conspecifics, as well as having increased fitness in the absence of the fungus. Founder effects (c): If founder effects, or evolution not related to enemy release, is responsible for the increase in fitness observed in the invasive range, we expect invasive range plants to be more fit than native range plants regardless of infection with E. sylvatica, though infection will likely still negatively affect the host, likely in a manner proportionate to the effect on native range plants
Alternatively, if release from the specialist enemy E. sylvatica has provided selective pressure for B. sylvaticum to have evolved some reallocation of resources from defense to growth and reproduction, we expect to be able to observe the reduction in defensive capabilities by invasive range plants in the form of strongly reduced fitness when infected as compared to infected native range plants. In other words, we expect the invasive range plants to be disproportionately negatively affected by infection with E. sylvatica. Additionally, if such evolution has occurred, we would also predict that even in the absence of the controlling enemy, invasive range plants will out-perform native range plants (Fig. 1b).
Factors independent of enemy release may be driving the evolution of invasive range populations of B. sylvaticum, such as founder effects and drift, or selection unrelated to enemy release. If the invasive range plants show increased fitness relative to their native range counterparts in both infected and uninfected states, such other evolutionary forces may be the best explanation for the observed increases in fitness in the invasive range (Holmes et al. 2010; Roy et al. 2011; Halbritter et al. 2012), though more work will be needed to determine the extent to which founder effects, genetic drift, or selective pressures not related to enemy release are responsible for such evolution (Fig. 1c).
This experiment also allowed us to test explicitly for effects of infection by E. sylvatica on germination rates. All germination rates observed to date of uninfected seeds of European origin are from naturally infected seeds that were treated to kill the endophyte (Roy et al. 2011; Halbritter et al. 2012). It is necessary to compare germination rates of seeds from the same population produced with and without the endophyte to accurately determine the effect of Epichloë on germination rate, because infection of seeds at any time may negatively impact germination, including prior to heat treatment.
Materials and methods
Focal species
We are working with the grass B. sylvaticum (Huds.) P. Beauv., an aggressive invasive species in the USA (Roy 2010), introduced in the early 1900s by the US Department of Agriculture (USDA) for agronomic research. Records from the Office of Foreign Plant Introduction dating back to 1912 indicate that B. sylvaticum was being imported from India, Sweden, Russia, and probably other localities (Rosenthal et al. 2008). The grass was first collected in the wild in Oregon in 1939 (Chambers 1966), and has become increasingly common during the last 15 years (Rosenthal et al. 2008). This grass is of particular concern because it is shade-tolerant (Holmes et al. 2010) and forms vast, virtually monospecific carpets in the forest, which crowd out other vegetation (Kaye and Blakeley-Smith 2006) and, similar to other grasses, may reduce conifer seed germination (Powell et al. 2006). It is found commonly and in high densities in the central Willamette valley, particularly from Eugene to Corvallis, and appears to be in the midst of rapid range expansion (Rosenthal et al. 2008; Roy 2010).
Study sites
We sourced seed and tested for endophyte infection in Switzerland (center of the native range) and Oregon (USA, epicenter of the invaded range) at 21 field sites (Supplementary Table S1; Fig. 2), a subset of which were used for germination, growth rate, and mortality assays. Climate in the two areas are similar: Zürich (Switzerland) has an annual precipitation of 1,086 mm and Eugene (Oregon, USA) has 1,254 mm. Mean temperatures for Zürich and Eugene are 8.5 and 11.9 °C, respectively (climate information from www.meteoschweiz.admin.ch and the Western Regional Climate Center www.wrcc.dri.edu). There are, however, seasonal differences between the two sites: summer is much drier in Oregon (mean precipitation for July and August in Zürich is 124.5 mm, but it is only 20 mm for Eugene).

Wild endophyte screen results. We only found evidence of E. sylvatica in one wild population in the invaded range, the Fisherman site near Mill City, Oregon. All individuals from all populations tested from the European native range were infected, however. *Native range data from Bucheli and Leuchtmann (1996)
Infection rates
To determine whether the fungus was present in the invaded range we used the Agrinostics Field Tiller immunoblot kit (Agrinostics Ltd. Co., Watkinsville, GA, USA). Because the kit was developed for a different species, we verified its efficacy by isolating E. sylvatica from B. sylvaticum seeds collected in Switzerland where infection rates are at or near 100 % (Fig. 2; Bucheli and Leuchtmann 1996; Leuchtmann and Schardl 1998). For additional positive controls we used leaf tissue from plants grown from Swiss seeds and Dactylis glomerata showing choke symptoms caused by Epichloë typhina (Pers.:Fr.) Tul. Finally, we verified a subset of immunoblot results with an E. sylvatica-specific PCR screen; these indicated that the immunoblot results were valid (see Supplemental Methods for details).
Genetic data and historical records suggest that B. sylvaticum was likely initially introduced from two Bureau of Plant Introduction experimental plots, one near Eugene, Oregon, and one near Corvallis, Oregon (Rosenthal et al. 2008). We therefore screened three populations near Eugene (Mount Pisgah, Jasper, and Jasper State Park) and two near Corvallis (Bald Hill and Sweet Home; see Table S1). The vegetation and other site characteristics have been described elsewhere (Roy et al. 2011).
We tested for E. sylvatica in the invaded range at three times: peak growing season (20 June 2010), 7 weeks later (4 August 2010), and at the end of the summer (26 August 2010). All tillers were collected at ground level within 24 h of analysis, wrapped in a paper towel and placed on ice. Thirty tillers per population were randomly sampled by taking the nearest tiller to a meter mark along two parallel transects each 15 m long and approximately 10 m apart.
Initial germination and growth rate assays
For the initial assay, seed material of B. sylvaticum was collected in late August 2007 from three wild populations in the native range and six in the invaded range (Table S1). Seeds were stored at 4 °C until needed. Prior to germination, seeds were deglumed and winnowed to remove aborted seeds. To reduce fungal attack, seeds were surface sterilized in 5 % bleach solution for 30 s and then rinsed twice with tap water. On May 22nd 2008 seeds were placed between four sheets of filter paper in a sterile Petri dish and dampened with a solution of gibberelic acid (50 mg GA3/500 ml tap water). Petri dishes were kept at room temperature and checked daily to ensure correct moisture level. As they germinated the seedlings from each of the US populations were transplanted into 200 cm3 Containers (D-40 cells, Steuwe and Sons, Corvallis, Oregon) filled with Rexius, Patio Potting Soil™ (one seedling per tube). The Swiss seeds had extremely low germination rates, and after day 11 we transferred the remaining seeds to trays filled with potting soil and transplanted them into containers upon germination.
We measured the aboveground height of seedlings 9 days after transplanting (with a few exceptions of 8–11 days). Seedling growth rates, a proxy for fitness (Poorter and Garnier 1999; Matzek 2012), were calculated by dividing the height at the time of measurement by the number of days since emergence. To test differences between ranges we used a mixed-model analysis of variance (ANOVA) with restricted maximum likelihood (REML) estimation of variance components. Range was designated as a fixed effect and population as a random effect.
EICA greenhouse experiment
A large number of seeds were collected from two native (Flaach:654, Rafz:395) and two invaded (Pisgah:293, Jasper:291; see Table S1) range populations of B. sylvaticum at the end of summer 2011. These were deglumed by hand, and then treated to remove the endophyte, following Nott and Latch (1993). The seeds were surface sterilized by immersion in 95 % EtOH for 1 min, full-strength bleach (6.15 % NaHClO) for 3 min, 30 s in 95 % EtOH, then triple rinsed in autoclaved deionized water. The seeds were allowed to dry on sterile filter paper, then were placed in sterile petri dishes and placed in 100 % humidity at 37 °C for 3 weeks in a sealed incubator. The seeds were then germinated on sterile water agar, with any seeds showing fungal infection being discarded.
Half of the germinants from each population were inoculated with E. sylvatica, for a total of 65 plants per population. Inoculation of B. sylvaticum with the endophyte was accomplished following Leuchtmann and Clay (1988). Working under a dissecting microscope, a 27-gauge sterile hypodermic needle was used to make a small incision just above the apical meristem of the seedlings at the two- or three-leaf stage of development (typically 5 days post germination). The needle was then used to collect a small sample of cultured fungal hyphae (isolated from Swiss seeds as described above), which was then inserted into the incision. Control plants were injected with a small drop of sterile deionized water (Leuchtmann and Clay 1988). Plants were grown for 5 days on agar before transplantation into soil (Black Gold, Sun Gro Horticulture, Agawam, MA, USA) in 10 cm pots.
The plants were randomly distributed in racks in the greenhouse, and were re-randomized every week. Height (length from longest leaf tip to soil surface) was measured every other day, from initial transplant into soil (19 December 2011) to harvest (29 February 2012). Daily growth rates were calculated as the difference in height between two subsequent measurements, divided by two. The data were analyzed using a repeated measures, mixed model analysis of variance (ANOVA), including site, nested in range, and inoculation as fixed effects, population included as a random effect, and daily growth rate as the response variable. Number of tillers at harvest and oven-dried biomass were analyzed by mixed model analysis of variance (ANOVA) as well, though without repeated measures. Tukey’s HSD was used to compare means. Mortality was analyzed using the log-rank Mantel–Haenszel test (Harrington and Fleming 1982).
A subset of both treatment groups for all populations (ten plants per treatment per population) were repotted into gallon pots and allowed to set seed, which was collected for second-generation germination assays. A subset of 100 seeds from each parent plant were deglumed by hand, surface sterilized as above, plated onto water agar to germinate, and observed for germination for 30 days. Ten seeds from each plant were also tested using the Agrinostics Seed Immunoblot kit (Agrinostics Ltd. Co., Watkinsville, GA, USA) to confirm infection status of the parent. Because seed infection by Epichloë was not entirely all-or-nothing, germination rates were analyzed by linear regression to examine trends in germination in response to rate of seed infection. Additionally, a mixed model ANOVA and Tukey’s test were used to examine treatment and range differences. Student’s t tests were used to examine pairwise differences.
All analysis was performed in R (version 2.15.1), using the packages vegan (Oksanen et al. 2013) and survival (Therneau 2013).
Results
Endophyte screening
Endophyte infection in Oregon, epicenter of the invasion, appears to be limited to a single population of the eight we sampled. Using the immunoblot test we found E. sylvatica in 41 of the 455 wild collected tillers from the invaded range (Fig. 2; Supplementary Table S1). The only infected plants from the invaded range were collected from Fisherman, near the northern limit of the invaded range. There was no effect of time sampled; within a population, all samples were either infected, or not infected. Our positive controls were consistently positive (see “Methods”). Immunoblot results were validated by screening a subset of samples (24 negative, 3 positive) with an Epichloë-specific PCR assay, which gave identical results to the immunoblot.
Germination rates
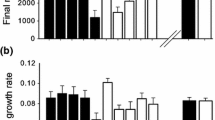
In our initial assay, germination was significantly higher in the invaded range (F = 13.101,7.33, P = 0.0079), and all populations in the invaded range had higher germination than the native range (Fig. 3a).

Results from our initial germination rate (a) and growth rate assays (b), May–June 2008. There is a significant difference in germination rates for the two ranges (t = 3.975.588, P = 0.0085), which we initially attributed to the fact that all native range populations are ubiquitously infected with E. sylvatica, and all invasive range populations tested lack the endophyte. There is also a significant difference in growth rates for the two ranges (t = 16.48159.539, P < 0.0001), in addition to differences by individual populations (letters represent differences at P < 0.05 by pairwise t test)
In our second germination assay with seeds originating from the common greenhouse study, endophyte infection and seed origin were decoupled (Fig. 4). We still found significantly higher germination in invasive populations (t = 5.0820.788, P < 0.0001), but there was also a significant interaction between range and infection status (F = 8.231,22, P = 0.0087), such that there is no difference in germination rates for infected seeds by range (t = 1.5911.001, P = 0.1397), while differences in germination rates of uninfected seed drove the entire trend (t = 9.318.736, P < 0.0001). In invasive populations there was a marginally significant trend towards reduced germination with endophyte infection (F = 3.791,10, P = 0.0801, r2 = 0.20), but in native range seeds germination rates significantly increased with increasing rates of endophyte infection (F = 5.111,12, P = 0.0432, r2 = 0.24). These results are in line with Brem and Leuchtmann (2002): they cite unpublished germination data showing higher germination rates in endophyte infected seeds in the native range.

Relationship between seed infection by E. sylvatica and germination. Each point represents a single maternal genotype originating in the native range (red) or invaded range (blue), with seed produced being open-pollenated in a common greenhouse. Two native and two invasive range populations were included, and there were no statistically significant differences between populations within a given range (invasive: t = 1.289.966, P = 0.2311; native: t = −0.4811.251, P = 0.6402). For linear regressions, the American range has r2 = 0.20 with P = 0.080; the European range has r2 = 0.24 with P = 0.043. Percentages were transformed using the standard arcsine square root transformation
Growth rates
In our initial seedling growth rate assay, we found that growth rates were significantly higher for plants from the invaded range (F = 20.921,7.22, P = 0.0024; Fig. 3b).
Our second growth rate assay was designed to de-couple endophyte infection from range of origin. We found that growth rates were not significantly reduced in the inoculated treatment for those seedlings from the native range (Fig. 5a; F = 0.0071,19, P = 0.933), but were significantly reduced for seedlings from the invaded range (Fig. 5b; F = 26.041,19, P < 0.0001). Interestingly, there was no significant effect of range origin in this greenhouse experiment once the negatively affected invasive range inoculated plants were removed from the analysis (F = 0.0661,19, P = 0.797). There was no significant effect of inoculation on final tiller number of surviving plants for either range (native: t = 1.894, P = 0.2326; invasive: t = 1.895, P = 0.2321), nor was there a significant effect of inoculation on the biomass of surviving plants for either range (native: t = 1.834, P = 0.2586; invasive: t = 0.103, P = 0.9996).

Daily growth rate (ratio of change/day) of B. sylvaticum seedlings in the greenhouse. In seedlings originating in the plant’s native range (Europe; a), there is no significant difference (F = 0.0071,19, P = 0.933) between seedlings inoculated with E. sylvatica (red) and those receiving the control treatment (blue). In seedlings originating in the invaded range (United States; b), however, there is significant effect of inoculation (F = 26.041,19, P < 0.0001)
Seedling mortality
Our second greenhouse experiment tracked mortality through time, in addition to growth rates. We observed a significant treatment by range interaction here, with inoculation not significantly changing mortality for Brachypodium originating from the native range (Fig. 6a; χ2 = 1.1, P = 0.299), but significantly increasing mortality for those seedlings originating from the invaded range (Fig. 6b; χ2 = 34, P < 0.0001).

Survival function estimates for B. sylvaticum seedlings. In seedlings originating in the plant’s native range (Europe; a), there is no significant difference (χ2 = 1.1, P = 0.299) between seedlings inoculated with E. sylvatica (red) and those receiving the control treatment (blue). In seedlings originating in the invaded range (United States; b), however, there is a significant effect of inoculation, leading to significantly reduced survivorship (χ2 = 34, P < 0.0001) within the first 2 weeks, and continuing to drop through time
Discussion
Pathogen or mutualist?
Harboring this endophyte has fitness costs for B. sylvaticum, but whether or not an endophyte is a pathogen or a mutualist depends on the specific context of host, symbiont, and environment (Carroll 1988; Scholthof 2007). Theoretically, if herbivores are present that significantly decrease fitness, then infected plants will have an advantage, provided the herbivores are deterred by the fungal alkaloids produced (Richardson et al. 2000; Brem and Leuchtmann 2001). Our results indicate that the endophyte E. sylvatica has low incidence in the invaded range (9 % overall; Fig. 2), while literature indicates that it is nearly ubiquitous in the native range (Fig. 2). In addition to the Swiss infections reported in Fig. 2 (Bucheli and Leuchtmann 1996; Roy et al. 2011; Leuchtmann, pers. com.), there are also reports of infection in Scandinavia, Finland, France, Spain, and Japan (Eckblad and Torkelsen 1989; Raynal 1994; Enomoto et al. 1998; Zabalgogeazcoa et al. 2000; Väre and Itämies 1995). Additionally, Adrian Leuchtmann reports having seen 100 % infection levels in populations of B. sylvaticum from Holland, Sweden, England, and Italy. He does, however, note that in one population from Sardinia, only two out of three plants were infected (A. Leuchtmann, pers. com.).
Data from the native range indicates that the endophyte increases resistance to insect herbivory (Brem and Leuchtmann 2001), but also decreases competitiveness (Brem and Leuchtmann 2002). We found that in the invaded range it can be detrimental to growth rates (Fig. 5) and seed germination (Fig. 4), but increases seed germination in the native range (Fig. 4). While the loss of anti-herbivore properties conferred by the endophyte could have made it more susceptible to being eaten in the invaded range, data show that vegetative insect herbivory has little effect on fitness in either range (Brem and Leuchtmann 2001; Halbritter et al. 2012). However, the observed elevation in herbivory in the invaded range may be evidence that loss of the endophyte does mean a loss of protection (Halbritter et al. 2012).
Brachypodium sylvaticum may be controlled by the host-specific endophytic fungus E. sylvatica in its native range, given the effects of the fungus on its host’s competitive abilities (Brem and Leuchtmann 2002), and the differences in performance between ranges (Halbritter et al. 2012). This may contribute to the grass’s success as an invader in the Pacific Northwest, where Epichloë is largely absent. Given preliminary germination data in both ranges (Fig. 3a), we expected to see control by E. sylvatica acting through reduced germination; however, when infection status and range are deconfounded, it appears that lower germination in the native range is not caused by E. sylvatica (Fig. 4). In the absence of the endophyte, the European seeds germinate at significantly lower rates than the American seeds, indicating that differences in germination may be determined by genetic factors. The increase in germination with Epichloë infection in the native range, and the trend towards decrease in germination rates with infection in the invaded range (Fig. 4), lends support to our hypothesis that B. sylvaticum in the USA has lost defense and/or tolerance mechanisms (the ability to survive and reproduce despite being infected; see Roy and Kirchner 2000) through evolution. Demonstrating that the loss of tolerance is in direct exchange for increased fitness will be interesting future work.
We show a near total absence of E. sylvatica infection in the invaded range, as well as a clear loss of tolerance of such infection by invasive-range B. sylvaticum from multiple populations throughout the invaded range (including the only population found to host the endophyte within that range). It is likely that ubiquitous infection of B. sylvaticum in Europe is maintained by a strong selection that is largely absent in the Pacific Northwest. This selective pressure may be acting through the seeds: while protection from folivores proved not to be important in previous studies (Halbritter et al. 2012), Epichloë may provide protection from seed-damaging insects or pathogens in the native range. There is, indeed, higher incidence of seed-associated insects and pathogens in the native range than the invasive (Halbritter et al. 2012). Further studies will be necessary to clarify the role of Epichloë endophytes in protection of seed.
EICA versus ERH
With regards to enemy release, EICA can be construed as a sub-case of ERH (Joshi and Vrieling 2005), though the mechanisms are distinct. Enemy Release Hypothesis can be explanatory in the absence of evolution where populations of an organism are directly controlled by co-evolved enemies (Keane and Crawley 2002; Liu and Stiling 2006), for example as in Ambrosia artemisiifolia, which seems to not have lost any defensive capabilities despite herbivore release upon invasion in France (Genton et al. 2005). Evolution of increased competitive abilities (EICA), however, is important when the release from those enemies provides selective pressures to re-allocate resources from defense to competitive traits, such as increased growth and reproduction (Blossey and Notzold 1995), or production of allelopathic chemicals (Uesugi and Kessler 2013). These two hypotheses lead to different predictions in our study system (Fig. 1).
Our experimental design allowed us to assess evolutionary change in the invaded range, such that we can effectively distinguish between the ERH and the EICA hypothesis. We found significantly increased mortality of inoculated B. sylvaticum originating from the invaded range as compared to their native range equivalents (Fig. 6). We also found reduced seedling growth rates in inoculated invasive-range plants (Fig. 5). Both of these results show a loss of tolerance for the host-specific fungal enemy in the invasive range populations tested, consistent with evolutionary loss of defensive mechanisms against this specific enemy, as predicted by the EICA hypothesis. The difference in germination rates seems to point to genetic mechanisms for increased germination in the invaded range, as well as loss of other factors controlling germination in the native range, such as seed-damaging pathogens and herbivores. These facts, taken together, are strong support for the EICA hypothesis, which predicts such a loss of defensive mechanisms to specific enemies in exchange for increased fitness in the invasive range.
Enemy release and invasion history
The story of any invasive species is unique, and while there may be unifying trends, each species has a particular history of introduction and a particular biology that influences its success. B. sylvaticum is no different; the success of this grass as an invader in the Pacific Northwest is no doubt influenced by the way in which it was introduced. During introduction, seed stock from all over the native range was planted in USDA test plots near Corvallis and Eugene (Rosenthal et al. 2008), promoting novel genetic combinations. Rapid range expansion may also have contributed to evolutionary changes (Rosenthal et al. 2008), independent of selective effects of enemy release. Such evolutionary drivers are theorized to be more important generally (Felker-Quinn et al. 2013), but it is necessary to keep in mind the individual nature of species invasions (Mitchell et al. 2006). This confluence of genotypes and brisk range expansion may have led to the rapid spread and fixation of resistance genes in the population, likely before subsequent dispersal, which is theorized to have been facilitated by logging in the region of the abandoned USDA test plots (Rosenthal et al. 2008). This argument supposes that there is selection for endophyte infection in the native range that is absent in the invaded range. If this is so, then it is unlikely to be leaf attacking insect herbivores, as these do not reduce fitness in B. sylvaticum, and insect herbivory is conspicuously elevated in the invaded range (Roy et al. 2011). Similarly, it is unlikely to be a large herbivore, as B. sylvaticum is unpalatable to most macroherbivores due to high silica content: rabbit, deer, and other macroherbivore browsing makes up an extremely small portion of total plant herbivory for this grass (Brem and Leuchtmann 2001). This appears to be true in both ranges (Roy et al. 2011). It is more likely to be a seed eating insect or seed pathogen, or an enemy affecting young seedlings, as these have stronger effects on fitness (Roy et al. 2011; Halbritter et al. 2012), and are in line with our germination results (Figs. 3a, 4).
Alternatively, the endophyte may have been lost during introduction: seeds could have been treated, either accidentally or purposefully, in ways that would have killed seed endophytes. During slow shipment or uncooled storage in the early twentieth century, seeds were likely subjected to conditions of heat (37 °C) and high humidity (~100 %) that would have led to loss of infection. Storage duration has also been shown to result in endophyte loss, with endophyte viability decreasing before seed viability (Gundel et al. 2009).
Endophytes and invasion
Vertically transmitted endophytes are commonly assumed to be mutualists, and are expected to have a positive effect on invasiveness (Richardson et al. 2000). For example, Rudgers et al. (2004) said “Specifically, vertically transmitted fungal endophytes may confer predictable advantages to invading grasses when they accompany their host to new environments (pp. 47)”. However, there is no reason to believe that all endophyte infections lead to more fit plants. In our first assay, we found greatly reduced growth rates of plants whose seeds originated in the native range (Fig. 3b), where infection is 100 %, and in our second assay, we found that endophyte infection significantly reduced growth rates of plants originating in the invaded range (Fig. 5). Given the trade-off between the costs of hosting a given endophyte and the benefits that such a symbiont can provide in a given context, it is no surprise that endophyte effects in invasion ecology are context-dependent. Our results are consistent with the published results of Brem and Leuchtmann (2002), who found that when they removed the endophyte from seeds, the resultant uninfected plants were faster growing, larger, and more competitive than infected plants.
Enemy release, in this case, is more complicated than loss of a single controlling organism upon invasion. In its native range, E. sylvatica seems to control B. sylvaticum in some ways (reducing growth rates and competitive abilities), but those detriments seem to be off-set by increased germination rates and potential protection from seed herbivores and pathogens. In the invaded range, the grass is released from control on growth and competitive abilities imposed by the fungus, likely because it is not necessary to harbor such a costly endophyte to maintain high seed viabilities in the invaded range. Whether that is through the additional release of control by a seed-damaging organism, or through novel genetic recombination that allows for high germination rates in the absence of the fungus is still to be determined.
References
Atwood JP, Meyerson LA (2011) Beyond EICA: understanding post-establishment evolution requires a broader evaluation of potential selection pressures. NeoBiota 7–25. doi: 10.3897/neobiota.10.954
Blossey B, Notzold R (1995) Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis. J Ecol 83:887–889. doi:10.2307/2261425
Bossdorf O, Prati D, Auge H, Schmid B (2004) Reduced competitive ability in an invasive plant. Ecol Lett 7:346–353. doi:10.1111/j.1461-0248.2004.00583.x
Bossdorf O, Auge H, Lafuma L, Rogers WE, Siemann E, Prati D (2005) Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 144:1–11. doi:10.1007/s00442-005-0070-z
Brem D, Leuchtmann A (1999) High prevalence of horizontal transmission of the fungal endophyte Epichloë sylvatica. Bull Geobot Inst ETH 65:3–12
Brem D, Leuchtmann A (2001) Epichloë grass endophytes increase herbivore resistance in the woodland grass Brachypodium sylvaticum. Oecologia 126:522–530. doi:10.1007/s004420000551
Brem D, Leuchtmann A (2002) Intraspecific competition of endophyte infected vs uninfected plants of two woodland grass species. Oikos 96:281–290. doi:10.1034/j.1600-0706.2002.960210.x
Bucheli E, Leuchtmann A (1996) Evidence for genetic differentiation between choke-inducing and asymptomtic strains of the Epichloë grass endophyte from Brachypodium sylvaticum. Evolution 50:1879–1887. doi:10.2307/2410746
Carroll G (1988) Fungal endophytes in stems and leaves: from latent pathogen to mutualistic symbiont. Ecology 69:2–9. doi:10.2307/1943154
Chambers KL (1966) Notes on some grasses of the Pacific Coast. Madroño 18:250–251
Clay K (1990) Fungal endophytes of grasses. Annu Rev Ecol Syst 21:275–297. doi:10.1146/annurev.es.21.110190.001423
Colautti RI, Ricciardi A, Grigorovich IA, MacIsaac HJ (2004) Is invasion success explained by the enemy release hypothesis? Ecol Lett 7:721–733. doi:10.1111/j.1461-0248.2004.00616.x
Eckblad FE, Torkelsen AE (1989) Epichloë typhina in Norway. Opera Bot 100:51–57
Enomoto T, Tsukiboshi M, Shimanuki T (1998) The gramineous plants in which Neotyphodium endophyes were found (in Japanese). J Weed Sci Technol 43:76–77
Faeth SH, Sullivan TJ (2003) Mutualistic asexual endophytes in a native grass are usually parasitic. Am Nat 161:310–325. doi:10.1086/345937
Felker-Quinn E, Schweitzer JA, Bailey JK (2013) Meta-analysis reveals evolution in invasive plant species but little support for evolution of increased competitive ability (EICA). Ecol Evol 3:739–751. doi:10.1002/ece3.488
Genton BJ, Kotanen PM, Cheptou P-O, Adolphe C, Shykoff JA (2005) Enemy release but no evolutionary loss of defence in a plant invasion: an inter-continental reciprocal transplant experiment. Oecologia 146:404–414. doi:10.1007/s00442-005-0234-x
Gundel PE, Martínez-Ghersa MA, Garibaldi LA, Ghersa CM (2009) Viability of Neotyphodium endophytic fungus and endophyte-infected and noninfected Lolium multiflorum seeds. Botany 87:88–96. doi:10.1139/B08-119
Halbritter AH, Carroll GC, Güsewell S, Roy BA (2012) Testing assumptions of the enemy release hypothesis: generalist versus specialist enemies of the grass Brachypodium sylvaticum. Mycologia 104:34–44. doi:10.3852/11-071
Harrington DP, Fleming TR (1982) A class of rank test procedures for censored survival data. Biometrika 69:553–566. doi:10.1093/biomet/69.3.553
Holmes SE, Roy BA, Reed JP, Johnson BR (2010) Context-dependent pattern and process: the distribution and competitive dynamics of an invasive grass, Brachypodium sylvaticum. Biol Invasions 12:2303–2318. doi:10.1007/s10530-009-9645-7
Joshi J, Vrieling K (2005) The enemy release and EICA hypothesis revisited: incorporating the fundamental difference between specialist and generalist herbivores. Ecol Lett 8:704–714. doi:10.1111/j.1461-0248.2005.00769.x
Kaye T, Blakeley-Smith M (2006) False-brome (Brachypodium sylvaticum). In: Boersma P, Reichard S, van Buren A (eds) Invasive Species Pac. Northwest. University of Washington Press, Seattle, pp 80–81
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170. doi:10.1016/S0169-5347(02)02499-0
Leuchtmann A, Clay K (1988) Experimental infection of host grasses and sedges with Atkinsonella hypoxylon and Balansia cyperi (Balansiae, Clavicipitaceae). Mycologia 80:291–297. doi:10.2307/3807624
Liu H, Stiling P (2006) Testing the enemy release hypothesis: a review and meta-analysis. Biol Invasions 8:1535–1545. doi:10.1007/s10530-005-5845-y
Matzek V (2012) Trait values, not trait plasticity, best explain invasive species’ performance in a changing environment. PLoS One 7:e48821. doi:10.1371/journal.pone.0048821
Meyer G, Clare R, Weber E (2005) An experimental test of the evolution of increased competitive ability hypothesis in goldenrod, Solidago gigantea. Oecologia 144:299–307. doi:10.1007/s00442-005-0046-z
Mitchell CE, Agrawal AA, Bever JD, Gilbert GS, Hufbauer RA, Klironomos JN, Maron JL, Morris WF, Parker IM, Power AG, Seabloom EW, Torchin ME, Vázquez DP (2006) Biotic interactions and plant invasions. Ecol Lett 9:726–740. doi:10.1111/j.1461-0248.2006.00908.x
Müller-Schärer H, Schaffner U, Steinger T (2004) Evolution in invasive plants: implications for biological control. Trends Ecol Evol 19:417–422. doi:10.1016/j.tree.2004.05.010
Nott H, Latch G (1993) A simple method of killing endophyte in ryegrass seeds. In: Hume D, Latch G, Easton H (eds) Proceedings of the second international symposium. Acremonium/grass Interact. AgResearch, Palmerston North, New Zealand, pp 14–15
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2013) Vegan: community ecology package. R package version 2.0-9. http://CRAN.R-project.org/package=vegan
Poorter H, Garnier E (1999) Ecological significance of inherent variation in relative growth rate and its components. In: Pugnaire F, Valladares F (eds) Handbook of functional plant ecology. Dekker, New York, pp 81–120
Powell GW, Pitt MD, Wikeem BM (2006) Effect of forage seeding on early growth and survival of lodgepole pine. J Range Manag Arch 47:379–384
Raynal G (1994) Presence in wild and grassland grasses of endophytic fungi belonging to the genus Acremonium (in French). In: ANPP—Quatr Conférence Int Sur Mal Plantes, pp 1125–1130
Richardson DM, Allsopp N, D’Antonio CM, Milton SJ, Rejmanek M (2000) Plant invasions—the role of mutualisms. Biol Rev 75:65–93. doi:10.1111/j.1469-185X.1999.tb00041.x
Rosenthal DM, Ramakrishnan AP, Cruzan MB (2008) Evidence for multiple sources of invasion and intraspecific hybridization in Brachypodium sylvaticum (Hudson) Beauv. in North America. Mol Ecol 17:4657–4669. doi:10.1111/j.1365-294X.2008.03844.x
Roy BA (2010) Brachypodium sylvaticum. Invasive Species Compend. http://www.cabi.org/isc/datasheet/9890
Roy BA, Kirchner JW (2000) Evolutionary dynamics of pathogen resistance and tolerance. Evolution 54:51–63. doi:10.1111/j.0014-3820.2000.tb00007.x
Roy BA, Coulson T, Blaser W, Policha T, Stewart JL, Blaisdell GK, Güsewell S (2011) Population regulation by enemies of the grass Brachypodium sylvaticum: demography in native and invaded ranges. Ecology 92:665–675
Rudgers JA, Koslow JM, Clay K (2004) Endophytic fungi alter relationships between diversity and ecosystem properties. Ecol Lett 7:42–51. doi:10.1046/j.1461-0248.2003.00543.x
Rudgers JA, Afkhami ME, Rúa MA, Davitt AJ, Hammer S, Huguet VM (2009) A fungus among us: broad patterns of endophyte distribution in the grasses. Ecology 90:1531–1539
Saikkonen K, Lehtonen P, Helander M, Koricheva J, Faeth SH (2006) Model systems in ecology: dissecting the endophyte–grass literature. Trends Plant Sci 11:428–433. doi:10.1016/j.tplants.2006.07.001
Schardl CL (1996) Epichloë species: fungal symbionts of grasses. Annu Rev Phytopathol 34:109–130. doi:10.1146/annurev.phyto.34.1.109
Scholthof K-BG (2007) The disease triangle: pathogens, the environment and society. Nat Rev Microbiol 5:152–156. doi:10.1038/nrmicro1596
Therneau T (2013) A package for survival analysis in S. R package version 2.37-4. http://CRAN.R-project.org/package=survival
Uesugi A, Kessler A (2013) Herbivore exclusion drives the evolution of plant competitiveness via increased allelopathy. New Phytol 198:916–924. doi:10.1111/nph.12172
Van der Meijden E (1996) Plant defence, an evolutionary dilemma: contrasting effects of (specialist and generalist) herbivores and natural enemies. Entomol Exp Appl 80:307–310. doi:10.1111/j.1570-7458.1996.tb00941.x
Van Kleunen M, Schmid B (2003) No evidence for an evolutionary increased competitive ability in an invasive plant. Ecology 84:2816–2823. doi:10.1890/02-0494
Väre H, Itämies J (1995) Phorbia phrenione (Séguy) (Diptera: Anthomyiidae) in Finland. Sahlbergia 2:119–124
Vilà M, Gómez A, Maron JL (2003) Are alien plants more competitive than their native conspecifics? A test using Hypericum perforatum L. Oecologia 137:211–215. doi:10.1007/s00442-003-1342-0
Willis AJ, Memmott J, Forrester RI (2000) Is there evidence for the post-invasion evolution of increased size among invasive plant species? Ecol Lett 3:275–283. doi:10.1046/j.1461-0248.2000.00149.x
Zabalgogeazcoa I, Vázquez de Aldana B, García Criado B, García Criado A (2000) Fungal endophytes in natural grasslands of western Spain. Book Abstr. Grassl. Conf. 4th Int. Neotyphodium/Grass Interact. Symp. University of Paderborn, Soest, Germany, p 126
Acknowledgments
Adrian Leuchtmann introduced B. Roy to Brachypodium sylvaticum and Epichloë sylvatica in Switzerland. Several individuals or agencies kindly allowed us to work on their land. In Switzerland we thank: Forstämtern, Grün Stadt Zürich, and Kreisförstern. In the US we thank: S. DeGhetto and Corvallis Parks for Bald Hill, B. Marshall and Cascade Timber for Sweet Home, T. Winters and Lane County Parks for Mt. Pisgah (HBRA), J. Reed and G. de Grassi for Jasper, D. Johnson for MacForest within the MacDonald-Dunn Forest, and J. Leroux for Owl; the remaining US sites were located in the Willamette National Forest. We are most grateful to K. Blaisdell, A. Clark, M. Cruzan, M. Davis, B. Dentinger, K. McCulloch, A. Miller, and L. Reynolds for assistance with lab work and discussions. Funding was provided by NSF DEB-0515777 and DEB-0841613, granted to B. A. Roy; R. Vandegrift was supported by a National Science Foundation Graduate Research Fellowship, DGE-0829517.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Vandegrift, R., Blaser, W., Campos-Cerda, F. et al. Mixed fitness effects of grass endophytes modulate impact of enemy release and rapid evolution in an invasive grass. Biol Invasions 17, 1239–1251 (2015). https://doi.org/10.1007/s10530-014-0791-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-014-0791-1