
Abstract
Secondary spread largely determines the distribution and success of invasive species and depends ultimately on the capacity of the invader to disperse and colonise over multiple scales. Spread of the invasive seaweed Codium fragile ssp. fragile (Codium) can occur through the dispersal of vegetative fragments, which can be buoyant or non-buoyant depending on environmental conditions. This study examined the factors influencing the dispersal, settlement, and establishment of these two types of propagules in eelgrass (Zostera marina) meadows of îles de la Madeleine, Canada, where Codium lives epiphytically on eelgrass rhizomes. To measure dispersal, ~1,400 Codium fragments were marked, released, and tracked under different hydrodynamic conditions in areas of high and low eelgrass density. Under all conditions, buoyant fragments dispersed one to two orders of magnitude further than non-buoyant fragments. Dispersal distance was positively correlated with wind speed (a proxy for surface currents in this system) for buoyant fragments and with current speed for non-buoyant fragments. For the latter, dispersal distance was also negatively correlated with eelgrass height and density. Natural deposition of drifting fragments in experimentally-manipulated eelgrass meadows was variable in space and time, but was not affected by eelgrass shoot density. Experimental disturbance of eelgrass meadows enhanced the density, biomass, and percent cover of Codium, suggesting that the exposure of eelgrass rhizomes by natural or anthropogenic disturbance promotes invasion by Codium. Our results highlight the importance of small-scale field experiments in determining the local factors affecting the spread of invasive species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The global spread of invasive species through anthropogenic means is well recognised and at times well documented. However, secondary spread after initial introduction remains less studied (Vander Zanden and Olden 2008) though it can contribute to the success of invasive species (see reviews by Parker et al. 1999; Molnar et al. 2008). The geographic range and distribution of any species is ultimately determined by environmental tolerances and biological interactions, but the rate and pattern of range expansion of an introduced species depends upon its dispersal (Schaffelke et al. 2006; Lester et al. 2007). Dispersal will not only depend on species traits (e.g., the number and characteristics of the propagules produced), but also on environmental conditions that can influence spread (e.g., distance and direction dispersed) and survival in new locations (Lodge 1993; Johnson and Padilla 1996; Johnson and Chapman 2007; Simberloff 2009). With regards to invasive species, it is therefore vital to understand the factors controlling secondary spread to ensure effective management strategies (Vander Zanden and Olden 2008; Wilson et al. 2009).
For invasive species, two complementary aspects make up secondary spread: (1) diffusive local dispersal, which increases population density and slowly expands the species’ range, and (2) long-distance dispersal (often termed “jump dispersal”) into non-adjacent areas. This latter process can rapidly expand the invaded range by creating distant satellite populations from which additional local spread can occur (Reed et al. 1988; Wilson and Lee 1989; Hengeveld 1994; Hastings et al. 2005; Mineur et al. 2010; Johnson et al. 2012). This combination of dispersal mechanisms should greatly increase both the rate of spread and the probability that the species will persist in its new environment (Johst et al. 2002; Ellner and Schreiber 2012).
Local dispersal usually occurs through the natural reproductive and dispersal mechanisms of the species in question, whereas jump dispersal in invasive species is usually interpreted as a result of anthropogenic mechanisms (Blakeslee et al. 2010; Johnson et al. 2012; Taylor et al. 2012). For example, Suarez et al. (2001) demonstrated that invasive Argentinean ants (Linepithema humile) dispersed over large distances by anthropogenic means (mostly by ships) in addition to slower local spread by natural mechanisms. Similarly, jump dispersal in the ash borer beetle (Agrilus panipenni) is associated with the transport of infested wood products (Muirhead et al. 2006), and zebra mussels (Dreissena polymorpha) can be carried between lakes and watersheds by boaters (Buchan and Padilla 1999; Johnson et al. 2001).
However, many species also have natural mechanisms for assuring the long-distance dispersal of their propagules, and thus jump dispersal can also occur in invasive species without human intervention (Forrest et al. 2000; Lyons and Scheibling 2009). In marine and aquatic systems, many species across different phyla have planktonic life stages that can disperse over long distances via currents (Norton 1992; Kinlan and Gaines 2003). Marine macroalgae in particular can employ several strategies for spread (Gaylord et al. 2002), including both sexually- and asexually-produced propagules (gametes, spores, fragments, or whole thalli) which can either settle near the parent individual (local spread) or disperse over long distances (jump dispersal). Positively-buoyant propagules (usually fragments or whole plants) are common in many macroalgae (van den Hoek 1987; Santelices 1990; Norton 1992), and thus invasive marine algae that have the ability to float may have an enhanced potential for rapid secondary spread into uninvaded areas. Although local and long-distance dispersal can thus both contribute to the successful spread of an invasive species, the exact mechanisms and factors controlling them may differ substantially. For example, conditions that promote the local spread of algae, such as short propagule viability and rapid sinking speed, may reduce the probability of jump dispersal (Gaylord et al. 2002; Harwell and Orth 2002; Higgins et al. 2003).
Finally, successful spread requires not only the dispersal of propagules, but also their survival and establishment (Lodge 1993). Determining the factors controlling both local and long-distance dispersal, as well as their settlement and survival (i.e., recruitment), is thus important for understanding the invasion dynamics of a species as well as developing effective management strategies (Johnson and Chapman 2007).
The invasive subspecies of the green alga Codium fragile (C. fragile ssp. fragile, formerly C. fragile ssp. tomentosoides and hereafter referred to as Codium) has been introduced to and is present in many temperate regions (Lyons and Scheibling 2009). It is currently spreading through îles de la Madeleine in the Gulf of St. Lawrence, Canada, especially in the large (20–40 km2) lagoons that make up a large part of the coastal zone of this archipelago (see Simard et al. 2007; Drouin 2013 for details). Whereas Codium normally attaches to hard substrata (such as rocks or shells), in this system it is almost always found attached to the rhizomes of eelgrass Zostera marina (Garbary et al. 2004; Simard et al. 2007; Drouin et al. 2011), which forms extensive meadows in the lagoons.
Codium reproduces exclusively asexually though parthenogenic gametes, isolated filaments (utricles), and fragments (Trowbridge 1998; Prince and Trowbridge 2004). Fragments range in length from small buds (ca. 1 cm in length) released from adults to entire thalli (30–40 cm) that have been dislodged from the substratum. Whereas the gametes and utricles are evidently produced as propagules, the latter more likely function fortuitously as such. Regardless, any type of fragment can attach and grow into a mature thallus if it comes into prolonged contact with a suitable substratum (Nanba et al. 2002; Scheibling and Melady 2008; Watanabe et al. 2009). Within eelgrass meadows, short-range dispersal by gametes appears to be the dominant mode of colonisation, at least on artificial substrata (Drouin 2013). However, these cannot disperse far enough (Trowbridge 1998) to explain longer dispersal within and between the lagoons. Instead, it appears that this long-distance dispersal occurs through the dispersal of fragments either directly via the settlement and attachment of fragments or indirectly via the release of gametes from fragments.
Under certain conditions, Codium fragments can be positively buoyant (Dromgoole 1982; Gagnon et al. 2011), thus increasing their dispersal potential (Gagnon et al. 2011). This state only occurs, however, under conditions of high light intensity when photosynthesis causes excess gas bubbles to accumulate in the thallus. Shifts from buoyant to non-buoyant states can occur over both short (daily) and long (seasonal) temporal scales (Gagnon et al. 2011). Therefore, both buoyant and non-buoyant fragments can be produced, depending on the light environment, and these fragments likely have differing dispersal potentials. The dispersal potential of non-buoyant fragments would be expected to depend on local bottom conditions and characteristics (e.g., current speed, topography, vegetation; Gaylord et al. 2002; Watanabe et al. 2009) whereas buoyant fragments should be more affected by surface conditions (e.g., surface currents; Biber 2007).
In addition to the dispersal potential of fragments, the spread of a species such as Codium will be determined by the number of fragments that settle and survive (i.e., the quantity of fragments which settle on a suitable substratum and grow into adult thalli). As Codium grows primarily on eelgrass rhizomes in the studied system, it is thus essential for drifting fragments to first be deposited into eelgrass meadows and then come in contact with rhizomes. Deposition is likely a function of the fragment properties (e.g., size, buoyancy), environmental conditions (e.g., currents), and structural aspects of the eelgrass meadow (i.e., density and height of plants). Attachment will then depend on processes that expose rhizomes (e.g., currents, disturbance), which are normally covered by sand or mud.
Here we examined the dispersal of buoyant and non-buoyant Codium fragments within the îles de la Madeleine lagoon system and determined how environmental factors (e.g., current speed, vegetation) affect these processes. We also measured the colonisation of fragments in eelgrass meadows where we manipulated the structure (height and density of shoots) to determine how variation in these aspects of the natural environment could affect the settlement of fragments. We hypothesized that buoyant fragments disperse further than non-buoyant fragments. In addition, we hypothesize that buoyant fragments are not affected by bottom characteristics, whereas non-buoyant fragments are, so that their dispersal is limited by high eelgrass density and height. Finally, we examined the role of disturbance on the colonisation rates of Codium. To this end, we did a manipulative experiment to evaluate the hypothesis that disturbance promotes the settlement of Codium thalli, possibly by providing more suitable substratum.
Methods
Study site
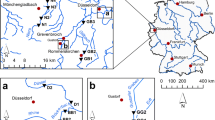
This study was done in 2008 and 2009 in the Lagune de la Grande-Entrée in the îles de la Madeleine archipelago, located in the Gulf of St. Lawrence, eastern Canada (Fig. 1). The lagoon is dominated by eelgrass (Z. marina) meadows, interspersed with bare areas of sand flats, with a salinity ranging from 28 to 32 psu and maximum temperatures reaching 22–24 °C in the summer. Codium has been present since at least 2003 (Simard et al. 2007) and mainly occurs in dense patches in shallow areas (<3 m depth) attached to eelgrass rhizomes. The lagoon is not affected by oceanic currents, so the only currents affecting fragments within the lagoons are weak tidal currents (<0.05 m s−1; Koutitonsky et al. 2002) and those caused by wind.

Location of study sites in the Lagune de la Grande-Entrée in the îles de la Madeleine archipelago. Grosse-Île: 47°37′07″N, 61°32′25″W; Salt Mine: 47°36′26″N, 61°33′55″W; Old Harry: 47°35′26″N, 61°29′10″W
Fragment dispersal
A series of dispersal experiments was done in July and August 2008 on the south shore of Grosse-Île to determine the role of buoyancy, eelgrass density, and current speed on the dispersal distance of Codium fragments in eelgrass meadows. Different methods were used to determine the dispersal of buoyant and non-buoyant fragments, but in both cases trials were conducted under unidirectional current conditions (i.e., during ebb or flood tide) in waters of 1–2 m depth.
For buoyant fragments, 10 fragments (approximately 10–15 cm long, with 1–2 branches) were released by a diver from a fixed point approximately 20 cm from the bottom to simulate tips breaking off from attached thalli (i.e., the average height of Codium thalli in the lagoon). A line was marked with rope and buoys 10 m downstream, and another diver recorded the time each fragment took to cross the line (i.e., disperse 10 m). These fragments were marked with brightly coloured string (for easier visibility in the eelgrass), which was threaded through the fragments and trimmed ≤2 mm from the thallus so as to not become entangled or interfere with the hydrodynamic characteristics of the fragments; all fragments were thus accounted for during the trials. Eighteen trials (of ten fragments each) with buoyant fragments were conducted at several locations in Grosse-Île lagoon and under different hydrodynamic conditions (i.e., some during ebb tide, and some during flood time). The trials were conducted over 2 weeks, with one or two trials being done each day, and all trials were standardised for dispersal after 1 h (i.e., dispersal distances were estimated as D = (3,600 s × 10 m)/t, where D is the distance (m) dispersed in 1 h and t is the time (s) to disperse 10 m. Current speed was measured using a Nortek Vector acoustic Doppler velocimeter placed approximately 20 cm from the bottom (i.e., at the same height at which the fragments were released). Data on hourly wind speed were obtained from Environment Canada and used as a proxy for surface current speed (note however, that this was only possible due to the very weak tidal currents our sites and may not be applicable in many other areas). A type-II regression (using the mean estimated distance for each trial) were performed with distance as the dependent variable, and current and wind speeds as the independent variables. Regression assumptions (normality and linearity by visual inspection of the variances, and homoscedasticity with Bartlett’s test) were checked prior to performing the analysis, and all assumptions were met.
For non-buoyant fragments, 50 fragments (also 10–15 cm long) were similarly released from a starting point 20 cm from the bottom, but were allowed to disperse for a fixed time (1 h). For each trial, a point of release was randomly chosen within the study area and marked, and the density and height of eelgrass along a 10-m transect downstream (from the direction of the tidal current) were measured. Fragments were again marked with coloured string, to facilitate recovery. To maximise our chances of accounting for all fragments, a 4-m-wide and 1.5-m-high plastic fence, constructed of black plastic mesh (mesh opening: 1.3 cm), was set up 10 m downstream from the release point to catch fragments. The fence was well anchored to the bottom to ensure that the fragments could not pass under it. This, however, limited the maximum dispersal distance of non-buoyant fragments to 10 m. The distances the fragments travelled over 1 h were then recorded to the nearest 50 cm. Twenty-five trials were conducted under different hydrodynamic conditions and eelgrass characteristics, with 94 % of total released fragments accounted for. Current speed was measured as above, and the density and height of eelgrass shoots in the transect were estimated from ten small circular plots (diameter = 15 cm) located randomly along the release path. A multiple regression was performed using the mean dispersal distance of each trial as the dependent variable, and current speed, eelgrass height, and eelgrass density as independent variables. As for the previous analysis, we checked the assumptions for linear regression (normality, linearity, and homoscedasticity) prior to performing the analysis and found that neither eelgrass density nor height variances were homogeneous. Data for eelgrass density and height were thus reciprocally transformed prior to conducting the regression, which corrected the heteroscedasticity.
Fragment settlement
To determine how eelgrass characteristics impacted the natural settlement (i.e., deposition) of drifting Codium fragments, we manipulated eelgrass shoot density and monitored subsequent natural colonisation at three sites (Old Harry, Grosse-Île and Salt Mine; see Fig. 1). At the beginning of the experiment in June 2008 we established four blocks of three 1-m2 circular plots at each site (6 weeks later, two more blocks were added at each site to increase the number of replicates), and within each block, plots were randomly assigned a density treatment: no eelgrass shoots, 50 % of normal shoot density, and natural shoot density (see Table 1 for natural eelgrass densities at each site). The plots at Grosse-île and Salt Mine were at 1–2 m depth, while the plots at Old Harry were at 2–3 m depth. The different densities were achieved by cutting the eelgrass at the base of the shoots, and plots were trimmed regularly to ensure density treatments were maintained throughout the experiment. The height of the eelgrass shoots was measured at each site once a month throughout the experiment, but was not manipulated; the natural density of eelgrass shoots in the control plots was also measured at the same time (Table 1). The density and biomass of drifting (i.e., unattached) Codium fragments found in each of the plots were recorded each week for 9 weeks (mid-June to mid-August) with the fragments removed each time. Since fragment settlement differed significantly among the three sites (P = 0.03), each was analysed separately to better understand the processes occurring at each site (the Salt Mine site was removed from the analyses and not examined further as almost no fragments accumulated in any of the plots throughout the summer, see “Results”). To determine the effects of eelgrass density and time on Codium fragment density, we used a general linear mixed model implemented in SAS 9.3 (PROC GLIMMIX). Since a Shapiro Wilkes test showed that the Codium fragment density was non-normal and over-dispersed, this allowed us to specify a negative binomial distribution for Codium fragment density. In addition, the maximum likelihood estimation method used in GLIMMIX allowed us to run a repeated-measures analysis despite missing values in some of the sites (due to adding sites in mid-summer), unlike a classical ANOVA approach in which these must be removed. We checked for variance homogeneity using Bartlett’s test prior to running the analyses, which show that variances were homogeneous. We then applied a post hoc Tukey’s test to determine which weeks differed significantly from each other.
Disturbance-induced colonisation
To determine the effects of disturbance on the final phase of colonisation of eelgrass meadows by Codium (i.e., the attachment of fragments and growth of thalli) six pairs of 10-m-long transects (located at 1–2 m depth) were selected and marked with sub-surface buoys in an eelgrass meadow at the Grosse-Île site in mid-August 2008. We checked carefully to confirm that there was no Codium within the transects at that time and that eelgrass height and density were relatively consistent among transects. Each pair of transects was at least 50 m from adjacent pairs, and within each pair, transects were separated by 5–10 m. For each pair, one transect was randomly assigned the disturbance treatment, and the other left unmanipulated as a control. Disturbances were created by walking through the eelgrass while pulling an anchor, which exposed rhizomes (and removed some eelgrass biomass: >50 % reduction) along a ~30-cm wide strip (giving a total disturbed area of 3 m2), simulating the disturbance by a boat propeller or an anchor. This was done as a pulse treatment, i.e., only performed once. Ten months later (mid-June 2009), the percent cover of Codium attached to rhizomes in each transect was visually estimated (we ignored any drifting or unattached fragments) Individual thalli were then collected and counted, and their average length and total biomass in each transect measured. Paired t tests were used to determine if there were differences in percent cover, density, total biomass, and mean thalli length between the treatments.
Results
Fragment dispersal
When buoyant and non-buoyant trials were compared (all buoyant fragment trials grouped together, and all non-buoyant fragment trials grouped together), buoyant fragments dispersed one to two orders of magnitude further than non-buoyant fragments after 1 h. Approximately 60 % of the non-buoyant fragments dispersed less than 3 m from the release point after 1 h, and less than 10 % reached the fence at 10 m (Fig. 2). However, estimates of the dispersal of buoyant fragments after 1 h varied from 53 to 507 m (extrapolated from a 10-m distance), with 70 % dispersing 300 m or more (i.e., 2 orders of magnitude greater than the non-buoyant fragments; Fig. 2).

Dispersal distances of buoyant and non-buoyant Codium fragments in eeglrass meadows. Values for all fragments were pooled for each trial and standardised for estimated dispersal after 1 h [n buoyant = 180 (18 trials of 10 fragments each), n non-buoyant = 1,000 (25 trials of 50 fragments each)]
Current speed (as measured 20 cm above the bottom) did not significantly affect the dispersal distance of buoyant fragments (t = −0.67, P = 0.51, R 2 = 0.027). However, the effect of wind speed (i.e., a proxy of surface current) on the dispersal of these fragments was significant and highly positively correlated (Fig. 3; t = 7.15, P < 0.0001, R 2 = 0.76).

Correlation of average hourly wind speed and the estimated dispersal distance of buoyant Codium fragments (n = 18, each point represents the mean of one trial of ten fragments)
For non-buoyant fragments, current speed influenced dispersal, but its effects on distance were affected in turn by eelgrass height and density (Table 2; significant three-way statistical interactions between current speed and eelgrass density and height). At low current speeds, dispersal distances were similar in all conditions; however, at higher current speeds, fragments dispersed further in low eelgrass density and height conditions, and an increase in either height or density (or both) decreased the effect of current speed (Fig. 4).

Effect of current speed, eelgrass density, and eelgrass height on the dispersal of Codium fragments. Trials are separated into four categories: (1) low eelgrass density (< 350 shoots·m−2) and low eelgrass height (< 20 cm), n = 10; (2) low density and high height, n = 5; (3) high density and low height, n = 5; (4) high density and high height, n = 5
Fragment settlement
Biomass and fragment density were correlated (linear regression, P < 0.01, R 2 = 0.63), and thus only density was used for statistical tests as it more accurately represents propagule pressure. Almost no fragments accumulated in any of the quadrats in the Salt Mine site throughout the summer (only 15 in total over the whole experiment) and thus there were no significant differences between treatments or over time. At both the Old Harry and Grosse-Île sites fragment density varied over time, while eelgrass density had no significant effects (Table 3). Overall, there were more fragments at the Old Harry site than at the Grosse-Île site throughout most of the summer, with order-of-magnitude differences in early summer (June; Fig. 5). Temporal patterns varied between the two sites with a steady decrease in the number of fragments over the first half of the observation period at the Old Harry site (Fig. 5A) in contrast with a mid-season peak observed at the Grosse-Île site (Fig. 5B).

Effect of time and eelgrass shoot density on the density of Codium fragments at two sites over 9 weeks in summer 2008 (mean ± SE, n = 4 per treatment per week). Different letters indicate significant differences between weeks (within the same site)
Disturbance induced colonisation
Only five of the six pairs of transects could be found in June 2009 (the sixth pair was closest to the shoreline and was most likely affected by ice scouring over the winter). All measures of abundance (percent cover, biomass, and density) of attached Codium thalli were 3–5 times greater in the disturbed transects than in the undisturbed transects (Fig. 6; t 4 = −4.20, P = 0.014; t 4 = −3.03, P = 0.039; t 4 = −3.77, P = 0.020, respectively). In fact, there were very few Codium thalli found in the non-disturbed transects. However, the average length of the attached Codium was consistently around 15 cm and did not differ significantly between disturbed and undisturbed treatments (Fig. 6; t 4 = 0.24, P = 0.829).

Percent cover, total biomass, density, and mean thalli length of attached Codium thalli in disturbed and undisturbed transects after 10 months (mean ± SE, n = 5 per treatment. Asterisks indicate a significant difference between treatments determined through paired t tests (P < 0.05)
Discussion
Our results show a clear difference in the dispersal potential of buoyant and non-buoyant fragments, with buoyant fragments potentially contributing to long-range dispersal (van den Hoek 1987) should they continue to disperse. Indeed, our results certainly underestimated the dispersal of buoyant fragments—we only measured their dispersal over a short time period, and they could theoretically continue dispersing as long as they remain buoyant (i.e., if light conditions remained high enough to ensure buoyancy; Gagnon et al. 2011) and do not wash up on the shore, as seems to be the fate of many fragments (Gagnon et al. 2011). The dispersal potential of a buoyant fragment could be several hundred kilometres in one season, although this is likely limited by varying wind speeds and surface currents, as the spread of Codium along the NW Atlantic coast seems to be approximately 50 km year−1 (Lyons and Scheibling 2009). Although we saw no effect of bottom current speed (as measured 20 cm above the bottom) on the dispersal distance of buoyant fragments, there was a strong correlation with wind speed (R2 = 0.76), which drives currents at the water surface in this system. Long-distance dispersal of buoyant fragments (over several hundred metres to kilometres) is therefore likely driven primarily by local environmental conditions and may thus vary substantially both temporally and spatially. At a larger scale (tens to hundreds of kilometres), however, large-scale climatic or current patterns probably determine overall dispersal patterns. While many buoyant fragments will likely be removed from the system by washing up on sandy shores (Gagnon et al. 2011) or sinking to unsuitable areas (e.g., sandy or muddy bottoms), the few that do disperse to suitable habitats are likely of great importance to the spread of this species. In geographic areas with strong tidal currents, wind speed and direction may have a more limited effect on fragment dispersal, and in those cases, it would likely be the speed and direction of tidal surface currents that determine the dispersal of buoyant fragments.
In contrast, non-buoyant fragments dispersed over much shorter distances, and most had stopped dispersing completely (i.e., had settled to the bottom) in less than one hour. As expected, their dispersal was correlated with bottom conditions, primarily a function of the speed of bottom currents, but modulated by biotic features. As seen in rocky habitats (Watanabe et al. 2009), individual site characteristics also had important effects on the dispersal potential of Codium fragments, though the important factors in rocky habitats seem to be exposure, slope, type and roughness of substrate, and presence of turf algae. In our case, both high eelgrass density and height reduced the effects of current, limiting dispersal and likely promoting increases in local density (but see Drouin 2013, who found no effect of eelgrass density on the recruitment of Codium to artificial concrete blocks, though this was mainly recruitment from gametes and not fragments). However, this conclusion is based on observations that did not include situations of both high shoot density and height and the highest current speeds, therefore dispersal in these conditions is more difficult to predict. However, it is likely that these variables are not independent (i.e., high shoot height or density alters the bottom current speeds) in which case our conclusion that eelgrass characteristics limit dispersal would still hold true. Since eelgrass grows quickly throughout the summer and maximum height varies between sites, there are therefore likely seasonal differences in dispersal distances of non-buoyant fragments, with less dispersal later in the season. On the other hand, release height of propagules can affect dispersal potential (Gaylord et al. 2002), and this could also increase during the growing season as Codium grows as well, thereby enhancing dispersal. Likewise, there are deeper navigation channels within the lagoons where current speeds are much higher (pers. obs.). There is little eelgrass in these areas, but fragments that drift into them could disperse much further, so these channels could potentially serve as important “pathways” for the dispersal of non-buoyant fragments at the scale of the studied lagoon. Similarly, in areas where tidal currents are much higher and the bottom habitat is less complex, dispersal of bottom fragments may be much higher and contribute to long-distance dispersal (Holmquist 1994) although this is not the case here.
In addition to dispersal, the production of propagules and their survival after settlement plays a role in successful spread of a species (Lodge 1993; Simberloff 2009). There were, however, no clear patterns of settlement of Codium fragments with both spatial and temporal variation in fragment settlement across the lagoon. Two of our sites (Old Harry and Grosse-Île) were both located adjacent (<100 m) to large Codium populations, and yet Codium settlement differed significantly between these two sites. We suggest that differences in local current patterns are driving this pattern, as Old Harry is more isolated from the rest of the lagoon, while Grosse-Île is more open. Thus fragments within Old Harry may be better retained within the area than they are at Grosse-Île. The third site (Salt Mine) was located much further away from any Codium populations (nearly 1 km) and received almost no fragments throughout the summer, suggesting that distance from a source population may also play a role at this spatial scale. Finally, differences in colonisation may also arise from variations in fragment production among sites and over time, due to differences in natural conditions (e.g., wind-driven waves) or human activities (e.g., boating), both of which can cause fragmentation (Trowbridge 1998; D’Amours and Scheibling 2007).
Changes in the physical environment due to the growth of eelgrass may have also contributed to the results we obtained: eelgrass height increased significantly in all sites over time (P < 0.01 for all sites), approximately doubling in height from June through August, and was significantly higher at the Old Harry site than at the two other sites (P < 0.01). Therefore, the ideal height for retaining fragments may have occurred earlier in the season at the Old Harry site, while at Grosse-Île the eelgrass did not reach this height until later. Meanwhile, the height of the eelgrass at Salt Mine was significantly lower than the two other sites throughout the whole summer (P < 0.01), possibly also contributing to the very low number of fragments that settled there. There were, however, no significant differences between eelgrass density treatments at any sites. Thus it appears that the eelgrass density does not influence the depositional part of the colonisation process at small scales (i.e., several metres), but that is the density and height over larger areas (i.e., tens to hundreds of metres) of the meadows that determine how far fragments disperse and where they settle.
In contrast, the exposure of eelgrass rhizomes seems to be a key element determining the recruitment of Codium. Population density appears highest in areas where shoot density changes abruptly, such as the borders between eelgrass patches and sand (pers. obs.) where eelgrass rhizomes tend to be more exposed, likely due to greater water motion or ice scouring. These areas may be prime settlement areas for Codium, thereby contributing to the observed patchiness of Codium populations in the lagoons (Drouin et al. 2011). The strong positive effect of disturbance on recruitment that we experimentally demonstrated supports this idea, and the high density of Codium in shallow areas (pers. obs) is likely due, at least partially, to higher rates of disturbance there. Disturbances likely fall into two main categories: 1) anthropogenic disturbances caused by boat traffic (anchors and propellers) in the summer (Occhipinti-Ambrogi and Savini 2003), and 2) natural disturbances caused by ice scouring in shallow waters near the coastline (Williams 2007). The large size (~15 cm) of the individual thalli observed in the disturbed sites strongly suggests that the recruits were the result of fragments settling and not other types of propagules (i.e., gametes, utricles), as Codium growth during the September–May experimental period is generally limited (Fralick and Mathieson 1973, Bégin and Scheibling 2003), especially at our sites, which are covered by ice in winter. Disturbances have been previously shown to facilitate the establishment of many marine species, including invasive species (e.g., Reed et al. 2000; Valentine et al. 2007), and Codium’s capacity for long-distance dispersal through fragmentation coupled with frequently-disturbed habitats may create ideal circumstances, thus accounting for the rapid spread of this invasive species in the Gulf of St. Lawrence. Fragments seem to take several weeks to attach to a substratum, and seem to attach and grow more quickly under static conditions (Scheibling and Melady 2008). It is thus likely that the complex structure of the eelgrass rhizomes and low current speeds in eelgrass meadows promote the settlement and reattachment of fragments, which may not occur as easily in other habitats or areas with higher current speeds.
The spread of Codium via fragments thus seems to involve at least two distinct dispersal mechanisms. Non-buoyant fragments, which usually settle within metres of the parent plants, increase the local density, leading to dense patches. In contrast, the buoyant fragments have the potential to disperse over long distances (over several kilometres), leading to the establishment of new patches or populations. This type of pattern has been previously predicted for species in which long-distance jump dispersal occurs (Hastings et al. 2005; Cannas et al. 2006; Marco et al. 2011), and a similar natural bimodal dispersal strategy has been observed in other invasive algae (e.g., Undaria pinnatifida, Sargassum muticum; Forrest et al. 2000; Valentine et al. 2007), in which sporophytes can disperse long distances. This strategy also matches the distribution of Codium we have observed in the eelgrass meadows and regional spread observed elsewhere (i.e., Carlton and Scanlon 1985; Lyons and Scheibling (2009) have both observed that spreading populations of Codium in the NW Atlantic are disjunct). For both types of dispersal, disturbances to the eelgrass, be they natural or anthropogenic, greatly increase the chances of settlement and establishment. As previous work has shown buoyancy to be positively correlated with light intensity (Gagnon et al. 2011), the relative importance of these two types of dispersal will vary temporally. In any case, these two dispersal modes likely explain the patchy distribution of Codium seen in eelgrass meadows observed over multiple spatial scales (e.g., Drouin et al. 2011).
As long-distance spread of invasive species is usually associated with anthropogenic vectors (Wilson et al. 2009), the possibility that buoyant fragments could contribute to natural long-distance dispersal (jump dispersal) in this species is of particular interest. Natural jump dispersal should, of course, occur in species with long-lived planktonic stages (Shanks et al. 2003; Kinlan et al. 2005), but may also occur in other marine and aquatic invasive species, including other algae (e.g., Forrest et al. 2000; Stewart et al. 2007; Lyons and Scheibling 2009) and the organisms associated with floating algae (Bushing 1994; Helmuth et al. 1994; Thiel 2003; Minchinton 2006; Macreadie et al. 2011; Rothäusler et al. 2012). In terrestrial systems, natural jump dispersal may also be common for many plant species (Higgins and Richardson 1999; Higgins et al. 2003; Kirk et al. 2011; Nogales et al. 2012), insects and other invertebrates (Holway 1995; Williams and Williams 1998; Li et al. 2013) and undoubtedly birds and mammals (e.g., Vogel et al. 2003; Irestedt et al. 2013). Environmental conditions can, however, greatly affect dispersal potential and colonisation. Thus, knowledge of local environmental conditions, in addition to the traits of the invasive species in question, remains necessary for predicting the spread and managing the impacts of invasive species.
References
Bégin C, Scheibling RE (2003) Growth and survival of the invasive green alga Codium fragile ssp. tomentosoides in tide pools on a rocky shore in Nova Scotia. Bot Mar 46:404–412. doi:10.1515/BOT.2003.040
Biber PD (2007) Hydrodynamic transport of drifting macroalgae through a tidal cut. Estuar Coast Shelf Sci 74:565–569. doi:10.1016/j.ecss.2007.04.019
Blakeslee AMH, McKenzie CH, Darling JA, Byers JE, Pringle JM, Roman J (2010) A hitchhiker’s guide to the Maritimes: anthropogenic transport facilitates long-distance dispersal of an invasive marine crab to Newfoundland. Divers Distrib 16:879–891. doi:10.1111/j.1472-4642.2010.00703.x
Buchan LAJ, Padilla DK (1999) Estimating the probability of long-distance overland dispersal of invading aquatic species. Ecol Appl 9:254–265. doi:10.2307/2641184
Bushing WW (1994) Biogeographic and ecological implications of kelp rafting as a dispersal vector for marine invertebrates. In: Halvorson WL, Maender GJ (eds) The fourth California islands symposium: update on the status of resources. Santa Barbara Museum of Natural History, Santa Barbara, pp 103–110
Cannas SA, Marco DE, Montemurro MA (2006) Long range dispersal and spatial pattern formation in biological invasions. Math Biosci 203:155–170. doi:10.1016/j.mbs.2006.06.005
Carlton JT, Scanlon JA (1985) Progression and dispersal of an introduced alga: Codium fragile ssp. tomentosoides (Chlorophyta) on the Atlantic coast of North America. Bot Mar 28:155–1665. doi:10.1515/botm.1985.28.4.155
D’Amours O, Scheibling RE (2007) Effect of wave exposure on morphology, attachment strength and survival of the invasive green alga Codium fragile ssp. tomentosoides. J Exp Mar Biol Ecol 351:129–142. doi:10.1016/j.jembe.2007.06.018
Dromgoole FI (1982) The buoyant properties of Codium. Bot Mar 25:391–397. doi:10.1515/botm.1982.25.8.391
Drouin A (2013) Établissement et impacts de la macroalgue non indigène Codium fragile ssp. fragile dans les herbiers marins aux îles de la Madeleine. PhD Thesis, Université Laval
Drouin A, McKindsey CW, Johnson LE (2011) Detecting the impacts of notorious invaders: experiments versus observations in the invasion of eelgrass meadows by the green seaweed Codium fragile. Oecologia 168:491–502. doi:10.1007/s00442-011-2086-x
Ellner SP, Schreiber SJ (2012) Temporally variable dispersal and demography can accelerate the spread of invading species. Theor Popul Biol 82:283–298. doi:10.1016/j.tpb.2012.03.005
Forrest BM, Brown SN, Taylor MD, Hurd CL, Hay CH (2000) The role of natural dispersal mechanisms in the spread of Undaria pinnatifida (Laminariales, Paeophyceae). Phycologia 39:547–553. doi:10.2216/i0031-8884-39-6-547.1
Fralick RA, Mathieson AC (1973) Ecological studies of Codium fragile in New England, USA. Mar Biol 19:127–132. doi:10.1007/BF00353583
Gagnon K, McKindsey CW, Johnson LE (2011) Dispersal potential of invasive algae: the determinants of buoyancy in Codium fragile ssp. fragile. Mar Biol 158:2449–2458. doi:10.1007/s00227-011-1746-z
Garbary DJ, Fraser SJ, Hubbard C, Kim KY (2004) Codium fragile: rhizomatous growth in the Zostera thief of eastern Canada. Helgol Mar Res 58:141–146. doi:10.1007/s10152-004-0173-7
Gaylord B, Reed DC, Raimondi PT, Washburn L, McLean SR (2002) A physically based model of macroalgal spore dispersal in the wave and current-dominated nearshore. Ecology 83:1239–1251. doi:10.1890/0012-9658(2002)083[1239:APBMOM]2.0.CO;2
Harwell MC, Orth RJ (2002) Long-distance dispersal potential in a marine macrophyte. Ecology 83:3319–3330. doi:10.2307/3072082
Hastings A, Cuddington K, Davies KF et al (2005) The spatial spread of invasions: new developments in theory and evidence. Ecol Lett 8:91–101. doi:10.1111/j.1461-0248.2004.00687.x
Helmuth B, Veit RR, Holberton R (1994) Long-distance dispersal of a subantarctic brooding bivalve (Gaimardia trapesina) by kelp-rafting. Mar Biol 120:421–426. doi:10.1007/BF00680216
Hengeveld R (1994) Small-step invasion research. Trends Ecol Evol 9:339–342. doi:10.1016/0169-5347(94)90155-4
Higgins SI, Richardson DM (1999) Predicting plant migration rates in a changing world: the role of long-distance dispersal. Am Nat 153:464–475. doi:10.1086/303193
Higgins SI, Nathan R, Cain ML (2003) Are long-distance dispersal events in plants usually caused by nonstandard means of dispersal? Ecology 84:1945–1956. doi:10.1890/01-0616
Holmquist JG (1994) Benthic macroalgae as a dispersal mechanism for fauna: influence of a marine tumbleweed. J Exp Mar Biol Ecol 180:235–251. doi:10.1016/0022-0981(94)90069-8
Holway DA (1995) Distribution of the Argentine ant (Linepithema humile) in Northern California. Conserv Biol 9:1634–1637. doi:10.1046/j.1523-1739.1995.09061634.x
Irestedt M, Fabre P-H, Batalha-Filho H et al (2013) The spatio-temporal colonization and diversification across the Indo-Pacific by a “great speciator” (Aves, Erythropitta erythrogaster). Proc R Soc B 280:20130309. doi:10.1098/rspb.2013.0309
Johnson CR, Chapman ARO (2007) Seaweed invasions: introduction and scope. Bot Mar 50:321–325. doi:10.1515/BOT.2007.037
Johnson LE, Padilla DK (1996) Geographic spread of exotic species: ecological lessons and opportunities from the invasion of the zebra mussel Dreissena polymorpha. Biol Conserv 78:23–33. doi:10.1016/0006-3207(96)00015-8
Johnson LE, Ricciardi A, Carlton JT (2001) Overland dispersal of aquatic invasive species: a risk assessment of transient recreational boating. Ecol Appl 11:1789–1799. doi:10.2307/3061096
Johnson LE, Brawley SH, Adey WH (2012) Secondary spread of invasive species: historic patterns and underlying mechanisms of the continuing invasion of the European rockweed Fucus serratus in eastern North America. Biol Invasions 14:79–97. doi:10.1007/s10530-011-9976-z
Johst K, Brandl R, Eber S (2002) Metapopulation persistence in dynamic landscapes: the role of dispersal distance. Oikos 98:263–270. doi:10.1034/j.1600-0706.2002.980208.x
Kinlan BP, Gaines SD (2003) Propagule dispersal in marine and terrestrial environments: a community perspective. Ecology 84:2007–2020. doi:10.1890/01-0622
Kinlan BP, Gaines SD, Lester SE (2005) Propagule dispersal and the scales of marine community process. Divers Distrib 11:139–148. doi:10.1111/j.1366-9516.2005.00158.x
Kirk H, Paul J, Straka J, Freeland JR (2011) Long-distance dispersal and high genetic diversity are implicated in the invasive spread of the common reed, Phragmites australis (Poaceae), in northeastern North America. Am J Bot. doi:10.3732/ajb.1000278
Koutitonsky VG, Navarro N, Booth D (2002) Descriptive physical oceanography of Great-Entry Lagoon, Gulf of St. Lawrence. Estuar Coast Shelf Sci 54:833–847. doi:10.1006/ecss.2001.0860
Lester SE, Ruttenberg BI, Gaines SD, Kinlan BP (2007) The relationship between dispersal ability and geographic range size. Ecol Lett 10:745–758. doi:10.1111/j.1461-0248.2007.01070.x
Li X-S, Zhang Y-L, Settele J, Franzén M, Schweiger O (2013) Long-distance dispersal and habitat use of the butterfly Byasa impediens in a fragmented subtropical forest. Insect Conserv Divers 6:170–178. doi:10.1111/j.1752-4598.2012.00199.x
Lodge DM (1993) Biological invasions: lessons for ecology. Trends Ecol Evol 8:133–136. doi:10.1016/0169-5347(93)90025-K
Lyons DA, Scheibling RE (2009) Range expansion by invasive marine algae: rates and patterns of spread at a regional scale. Divers Distrib 15:762–775. doi:10.1111/j.1472-4642.2009.00580.x
Macreadie PI, Bishop MJ, Booth DJ (2011) Implications of climate change for macrophytic rafts and their hitchhikers. Mar Ecol Prog Ser 443:285–292. doi:10.3354/meps09529
Marco DE, Montemurro MA, Cannas SA (2011) Comparing short and long-distance dispersal: modelling and field case studies. Ecography 34:671–682. doi:10.1111/j.1600-0587.2010.06477.x
Minchinton TE (2006) Rafting on wrack as a mode of dispersal for plants in coastal marshes. Aquat Bot 84:372–376. doi:10.1016/j.aquabot.2006.01.007
Mineur F, Davies AJ, Maggs CA, Verlaque M, Johnson MP (2010) Fronts, jumps and secondary introductions suggested as different invasion patterns in marine species, with an increase in spread rates over time. Proc R Soc B 277:2693–2701. doi:10.1098/rspb.2010.0494
Molnar JL, Gamboa RL, Revenga C, Spalding MD (2008) Assessing the global threat of invasive species to marine biodiversity. Front Ecol Environ 6:485–492. doi:10.1890/070064
Muirhead JR, Leung B, Overdijk C, Kelly DW, Nandakumar K, Marchant KR, MacIsaac HJ (2006) Modelling local and long-distance dispersal of invasive emerald ash borer Agrilus planipennis (Coleoptera) in North America. Divers Distrib 12:71–79. doi:10.1111/j.1366-9516.2006.00218.x
Nanba N, Kado R, Ogawa H, Komuro Y (2002) Formation and growth of filamentous thalli from isolated utricles with medullary filaments of Codium fragile spongy thalli. Aquat Bot 73:255–264. doi:10.1016/S0304-3770(02)00036-0
Nogales M, Heleno R, Traveset A, Vargas P (2012) Evidence for overlooked mechanisms of long-distance seed dispersal to and between oceanic islands. New Phytol 194:313–317. doi:10.1111/j.1469-8137.2011.04051.x
Norton TA (1992) Dispersal by macroalgae. Br Phycol J 27:293–301. doi:10.1080/00071619200650271
Occhipinti-Ambrogi A, Savini D (2003) Biological invasions as a component of global change in stressed marine ecosystems. Mar Pollut Bull 46:542–551. doi:10.1016/S0025-326X(02)00363-6
Parker IM, Simberloff D, Lonsdale WM et al (1999) Impact: toward a framework for understanding the ecological effects of invaders. Biol Invasions 1:3–19. doi:10.1023/A:1010034312781
Prince JS, Trowbridge CD (2004) Reproduction in the green macroalga Codium (Chlorophyta): characterization of gametes. Bot Mar. doi:10.1515/BOT.2004.062
Reed DC, Laur DR, Ebeling AW (1988) Variation in algal dispersal and recruitment: the importance of episodic events. Ecol Monogr 58:321–335. doi:10.2307/1942543
Reed DC, Raimondi PT, Carr MH, Goldwasser L (2000) The role of dispersal and disturbance in determining spatial heterogeneity in sedentary organisms. Ecology 81:2011–2026. doi:10.2307/177289
Rothäusler E, Gutow L, Thiel M (2012) Floating seaweeds and their communities. In: Wiencke C, Bischof K (eds) Seaweed biology. Springer, Berlin, pp 359–380. doi:10.1007/978-3-642-28451-9_17
Santelices B (1990) Patterns of reproduction, dispersal and recruitment in seaweeds. Oceanogr Mar Biol 28:177–276
Schaffelke B, Smith JE, Hewitt CL (2006) Introduced macroalgae—a growing concern. J Appl Phycol 18:529–541. doi:10.1007/s10811-006-9074-2
Scheibling RE, Melady RA (2008) Effect of water movement and substratum type on vegetative recruitment of the invasive green alga Codium fragile ssp. tomentosoides. Bot Mar 51:341–349. doi:10.1515/BOT.2008.046
Shanks AL, Grantham BA, Carr MH (2003) Propagule dispersal distance and the size and spacing of marine reserves. Ecol Appl 13:159–169. doi:10.1890/1051-0761(2003)013[0159:PDDATS]2.0.CO%3B2
Simard N, Paille N, McKindsey CW (2007) Codium fragile ssp. tomentosoides: revue de littérature et situation aux Îles-de-la-Madeleine. Can Manuscr Rep Fish Aquat Sci 2786, vii + 39 p
Simberloff D (2009) The role of propagule pressure in biological invasions. Annu Rev Ecol Syst 40:81–102. doi:10.1146/annurev.ecolsys.110308.120304
Stewart HL, Payri CE, Koehl MAR (2007) The role of buoyancy in mitigating reduced light in macroalgal aggregations. J Exp Mar Biol Ecol 343:11–20. doi:10.1016/j.jembe.2006.10.031
Suarez AV, Holway DA, Case TJ (2001) Patterns of spread in biological invasions dominated by long-distance jump dispersal: insights from Argentine ants. Proc Natl Acad Sci 98:1095–1100. doi:10.1073/pnas.98.3.1095
Taylor K, Brummer T, Taper ML, Wing A, Rew LJ (2012) Human-mediated long-distance dispersal: an empirical evaluation of seed dispersal by vehicles. Divers Distrib 18:942–951. doi:10.1111/j.1472-4642.2012.00926.x
Thiel M (2003) Rafting of benthic macrofauna: important factors determining the temporal succession of the assemblage on detached macroalgae. Hydrobiologia 503:49–57. doi:10.1023/B:HYDR.0000008486.37391.60
Trowbridge CD (1998) Ecology of the green macroalga Codium fragile (Suringar) Hariot 1889: invasive and non-invasive subspecies. Oceanogr Mar Biol 36:1–64
Valentine JP, Magierowski RH, Johnson CR (2007) Mechanisms of invasion: establishment, spread and persistence of introduced seaweed populations. Bot Mar. doi:10.1515/BOT.2007.040
Van den Hoek C (1987) The possible significance of long-range dispersal for the biogeography of seaweeds. Helgol Meeresunters 41:261–272. doi:10.1007/BF02366191
Vander Zanden MJ, Olden JD (2008) A management framework for preventing the secondary spread of aquatic invasive species. Can J Fish Aquat Sci 65:1512–1522. doi:10.1139/F08-099
Vogel P, Cosson J-F, López Jurado LF (2003) Taxonomic status and origin of the shrews (Soricidae) from the Canary islands inferred from a mtDNA comparison with the European Crocidura species. Mol Phylogenet Evol 27:271–282. doi:10.1016/S1055-7903(02)00403-7
Watanabe S, Metaxas A, Scheibling RE (2009) Dispersal potential of the invasive green alga Codium fragile ssp. fragile. J Exp Mar Biol Ecol 381:114–125. doi:10.1016/j.jembe.2009.09.012
Williams SL (2007) Introduced species in seagrass ecosystems: status and concerns. J Exp Mar Biol Ecol 350:89–110. doi:10.1016/j.jembe.2007.05.032
Williams DD, Williams NE (1998) Freshwater invertebrates from the Bermuda Islands and their zoogeographical affinities. Trop Zool 11:353–369. doi:10.1080/03946975.1998.10539371
Wilson JB, Lee WG (1989) Infiltration invasion. Funct Ecol 3:379–380
Wilson JRU, Dormontt EE, Prentis PJ, Lowe AJ, Richardson DM (2009) Something in the way you move: dispersal pathways affect invasion success. Trends Ecol Evol 24:136–144. doi:10.1016/j.tree.2008.10.007
Acknowledgments
This study was funded by the Canadian Aquatic Invasive Species Network (CAISN) of the Natural Sciences and Engineering Research Council (NSERC) of Canada, Québec-Océan, the Fondation Communautaire Gaspésie-les-Îles, and an NSERC Discovery Grant (LEJ). We greatly appreciate the logistical support of the Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ) in Îles-de-la-Madeleine, especially M. Nadeau, B. Myrand, and F. Gallien. Finally we thank C. Chevrier-Turbide and O.Rhoades for assistance in the field, and V. Jormalainen for advice on statistical methods. We are also grateful for the comments of J. Ruesink and two anonymous reviewers which greatly improved the quality of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Gagnon, K., McKindsey, C.W. & Johnson, L.E. Roles of dispersal mode, recipient environment and disturbance in the secondary spread of the invasive seaweed Codium fragile . Biol Invasions 17, 1123–1136 (2015). https://doi.org/10.1007/s10530-014-0782-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-014-0782-2