
Abstract
Objectives
To characterize a novel feruloyl esterase from Escherichia coli BL21 DE3.
Results
The gene encoding BioH was cloned and overexpressed in E. coli. The protein was purified and its catalytic activity was assessed. BioH exhibited feruloyl esterase activity toward a broad range of substrates, and the corresponding kinetic constants for the methyl ferulate, ethyl ferulate, and methyl p-coumarate substrates were: K m values of 0.48, 6.3, and 1.9 mM, respectively, and k cat /K m values of 9.3, 3.8, and 3.8 mM−1 s−1, respectively.
Conclusions
Feruloyl esterase from E. coli was expressed for the first time. BioH was confirmed to be a feruloyl esterase.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Feruloyl esterases (EC 3.1.1.73, FAEs), which are a carboxylesterase subtype, participate in the release of hydroxycinnamic acids from plant cell walls (Benoit et al. 2008; Koseki et al. 2009). FAEs are widespread in plants, fungi, and bacteria. The potential of FAEs to degrade the plant cell wall is important when designing processes for improved biomass utilization (Faulds 2010).
Sanishvili et al. (2003) reported that BioH is a member of the protein hydrolase superfamily, and they demonstrated that it possesses weak thioesterase activity by comparing the crystal structure of BioH with those of other known enzymes. Xie et al. (2007) identified BioH as the sole enzyme responsible for the hydrolysis of the dimethylbutyryl-S-methyl mercaptopropionate to the free acid during the biocatalytic conversion of monacolin J to simvastatin. Wang et al. (2010) found that with BioH efficiently resolved a variety of secondary alcohols, with enantiomeric excess (ee) values of the desired product reaching 98 % under mild conditions. Jiang et al. (2014) proposed a new method for synthesizing dihydropyran derivatives by a Escherichia coli BioH esterase-catalyzed method. Jiang and Yu (2014a, b) found that BioH possesses sufficient activity to catalyze the aldol, Knoevenagel and Baylis–Hillman reaction. Gu et al. (2015) used E. coli BioH coupled with a mandelate racemase to successfully improve the ee value toward methyl (S)-o-chloromandelate based on a strategy of sequential hydrolysis and racemization.
Escherichia coli BL21 is often used as a negative control for expressing exogenous FAEs. In this work, by overexpressing the bioH gene in E. coli strain BL21, we characterized this enzyme as a novel FAEs. This work provides a more thorough examination of the function of BioH, and it enriches our knowledge of FAEs-producing bacteria.
Materials and methods
Materials
Methyl ferulate (MFA), methyl p-coumarate (MpCA), chlorogenic acid, ethyl caffeate, and ethyl ferulate were obtained from Langde (Yancheng, China). Substrates used to assess enzymatic substrate preferences were purchased from Sigma–Aldrich or Aladdin (Shanghai, China). Molecular biology reagents, including PCR reagents, T4 DNA ligase, restriction enzymes, as well as PCR purification, gel extraction, and plasmid miniprep kits, were purchased from TaKaRa (Dalian, China). IPTG was obtained from Sangon Biotech (Shanghai, China). All other chemicals were of analytical grade.
Plasmids, bacterial strains, and media
Escherichia coli DH5α was used for DNA manipulations and propagating the pColdII vector (TaKaRa) and its derivative. E. coli BL21 served as the host for recombinant protein production. E. coli strains were cultured in lysogeny broth (LB), ampicillin (Amp) was added at 100 μg/ml when required.
Cloning of the E. coli bioH gene, and overexpression and purification of BioH
The bioH gene was PCR-amplified from E. coli BL21 genomic DNA using two synthetic primers. The forward primer (5′-GCCG GAA TTC ATG AAT AAC ATC TGG TGG CAG A-3′) contains an EcoRI restriction site (underlined), and the reverse primer (5′-GC TCT AGA CTA CAC CCT CTG CTT CAA CG-3′) contains a XbaI restriction site (underlined). The PCR-amplified bioH gene and pColdII plasmid were digested with EcoRI and XbaI, purified, and ligated by T4 DNA ligase. The ligation products were transformed into E. coli DH5α. After identifying clones containing the correct insert via restriction enzyme analysis and DNA sequencing, the plasmid containing the bioH gene was transformed into E. coli BL21 competent cells.
BL21 cells containing the correct expression plasmid were inoculated into LB/Amp medium and grown overnight at 37 °C. An aliquot of the culture was transferred into fresh LB/Amp medium and grown at 37 °C until the OD600 value reached 0.4. Then, the culture was incubated at 15 °C for 30 min, and BioH expression was induced with 0.4 mM IPTG. The culture was grown at 15 °C for 24 h, and cells were collected by centrifugation (6000×g, 6 min, 15 °C). The cell pellet was re-suspended in 20 mM sodium phosphate buffer (pH 7.4), and the cells were lysed by ultra-sonication on ice. Subsequently, unbroken cells and debris were removed by centrifugation (6000×g, 15 min, 4 °C), and the supernatant was used as the crude enzyme. The overexpressed protein was histidine-tagged, and the crude extract was filtered through a 0.2 µm cellulose filter and purified using a Ni-HisTrap HP column (GE Healthcare). After the column was equilibrated in binding buffer (20 mM sodium phosphate buffer, 500 mM NaCl, 5 mM imidazole, pH 7.4), histidine-tagged BioH was eluted using 10 column vol of a linear 0–100 % buffer B gradient (buffer B, 20 mM sodium phosphate buffer, 500 mM NaCl, 500 mM imidazole, pH 7.4). Aliquots were analyzed by SDS-PAGE, and the purified enzyme was concentrated using an ultrafiltration spin column. The enzyme was flash frozen and stored at −70 °C until further use.
Analysis of feruloyl esterase activity and kinetic parameters
Feruloyl esterase activity was measured by HPLC. Solutions A (pure methanol) and B (formic acid, 0.2 %) were applied to a reversed-phase C18 SunFire column (25 cm × 4.6 mm inner diam., 5 μm particle size) (Waters) at 35 °C. The method was as follows: 0.5 min, 10 % A, 1 ml/min; 0.1 min, 10 % A, 1 ml/min; 15 min, 100 % A, 1 ml/min; 5 min, 100 % A, 1 ml/min; 9.9 min, 10 % A, 1 ml/min. Samples were filtered through a 0.45 μm filter, and detection was at 280 nm. The retention times and spectral data of each peak were used to determine whether the substrates were hydrolyzed. The reaction mixtures contained 2 ml substrate solutions and 0.1 ml purified protein, and were shaken at 35 °C for 4 h. The reactions were terminated by boiling them for 3 min. Control reactions in which the enzyme was replaced with the same volume of buffer were used to account for any spontaneous hydrolysis of the substrates.
The kinetic parameters were determined from the initial rates of hydrolysis of 0.0625–1 mM MFA, MpCA, ethyl caffeate, and ethyl ferulate in buffer (pH 8.8) at 40 °C. Under these conditions, BioH is stable, and spontaneous substrate hydrolysis is negligible. K m and V max were calculated using a Lineweaver–Burk plot. For the calculation of k cat , the molecular mass was determined by denaturing SDS-PAGE. Samples were analyzed by HPLC as previously described.
Substrate specificity
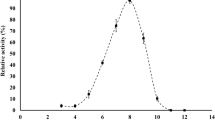
The enzymatic substrate profile of purified BioH was determined using an ester library as described previously (Liu et al. 2001). The hydrolysis of various ester substrates was measured colorimetrically using 4-nitrophenol as a pH indicator at 25 °C according to Janes et al. (1998). The decrease in absorbance at 412 nm was monitored for 30 min in a 96-well flat-bottom plate in a microplate reader. Each well contained 1 mM substrate, 4.65 mM N,N-bis(2-hydroxyethyl)-2-aminoethanesulfonic acid (BES), 0.434 mM 4-nitrophenol, 7.1 % v/v acetonitrile, and 38 μg enzyme/ml. Blank reactions without purified protein were performed for each substrate. All assays were performed in triplicate, and the average activities were quantified. The results are shown as means ± standard deviations.
Results and discussion
Cloning of the E. coli bioH gene, and overexpression and purification of BioH
The bioH gene from E. coli BL21 was cloned into the pColdII vector and overexpressed in E. coli BI21 (DE3). BioH was overexpressed in E. coli as a hexahistidine-tagged protein, and purified to homogeneity by nickel-affinity chromatography. SDS-PAGE was used to examine the purity of the enzyme, and it revealed a single protein band with an estimated molecular mass of 28 kDa (Fig. 1).

SDS-PAGE analysis of the overproduction and purification of BioH in E. coli. The gels were stained with Coomassie brilliant blue. Arrows indicate the overproduced and purified protein. Lane M low molecular weight standards, lane 1 overproduction of BioH in BL21 (DE3)/pColdII cell lysates after induction with IPTG, lane 2 fractions eluted from a histidine-affinity column
The enzyme activity profile
To investigate the substrate specificity of BioH, the purified protein was evaluated in parallel against a panel of 49 different substrates. BioH showed high preference and good enzymatic activity when feruloyl esters were used as substrates. BioH did not hydrolyze chlorogenic acid or rosmarinic acid (Table 1). The enzyme showed the highest activity toward methyl ferulate other than Tri-O-acetyl-d-glucal, which is similar to phenolic esters in the chemical structure. The broad substrates specificity of BioH was similar to two feruloyl esterases from Lactobacillus johnsonii (Lai et al. 2009) and Fae1A from Anaeromyces mucronatus (Qi et al. 2011). BioH could be classified into the feruloyl esterase base on its substrate specificity (Crepin et al. 2004).
Steady-state kinetic analysis
Purified BioH showed classical Michaelis–Menten kinetics, and linear double-reciprocal plots were obtained for hydroxycinnamic esters; the data are shown in Table 2. The best catalytic efficiencies (k cat /K m ) were obtained against ethyl ferulate and MpCA. The K m values demonstrated that the enzyme had a 10-fold higher affinity for ethyl ferulate than MpCA. Among the four tested substrates, BioH possessed the lowest affinity for ethyl caffeate.
Conclusions
A feruloyl esterase from Escherichia coli was found and expressed for the first time. We describe E. coli BioH as a novel feruloyl esterase with a wide substrate range. Since BioH was first identified via its relationship to biotin synthesis, it has subsequently been shown to possess additional enzymatic activities. Moreover, this protein could be obtained by the large-scale fermentation of E. coli at a low cost, and as a result, we believe that BioH has broad application prospects.
References
Benoit I, Danchin EGJ, Bleichrodt RJ, de Vries RP (2008) Biotechnological applications and potential of fungal feruloyl esterases based on prevalence, classification and biochemical diversity. Biotechnol Lett 30:387–396
Crepin VF, Faulds CB, Connerton IF (2004) Functional classification of the microbial feruloyl esterases. Appl Microbiol Biotechnol 63:647–652
Faulds CB (2010) What can feruloyl esterases do for us? Phytochem Rev 9:121–132
Gu JL, Ye LD, Guo F, Lv XM, Lu WQ, Yu HW (2015) Improved apparent enantioselectivity of a hydrolase by sequential hydrolysis and racemization. Tetrahedron Lett 56:1489–1491
Janes LE, Lowendahl AC, Kazlauskas RJ (1998) Quantitative screening of hydrolase libraries using pH indicators: identifying active and enantioselective hydrolases. Chem Eur J 4:2324–2331
Jiang L, Yu HW (2014a) Enzymatic Promiscuity: Escherichia coli BioH Esterase—catalysed aldol reaction and Knoevenagel reaction. Chem Res Chin Univ 30:289–292
Jiang L, Yu HW (2014b) An example of enzymatic promiscuity: the Baylis-Hillman reaction catalyzed by a biotin esterase (BioH) from Escherichia coli. Biotechnol Lett 36:99–103
Jiang L, Wang B, Li RR, Shen S, Yu HW, Ye LD (2014) Catalytic promiscuity of Escherichia coli BioH esterase: application in the synthesis of 3, 4-dihydropyran derivatives. Proc Biochem 49:1135–1138
Koseki T, Fushinobu S, Ardiansyah Shirakawa H, Komai M (2009) Occurrence, properties, and applications of feruloyl esterases. Appl Microbiol Biot 84:803–810
Lai KK, Lorca GL, Gonzalez CF (2009) Biochemical properties of two cinnamoyl esterases purified from a Lactobacillus johnsonii strain isolated from stool samples of diabetes-resistant rats. Appl Environ Microbiol 75:5018–5024
Liu AMF, Somers NA, Kazlauskas RJ, Brush TS, Zocher F, Enzelberger MM, Bornscheuer UT, Horsman GP, Mezzetti A, Schmidt-Dannert C, Schmid RD (2001) Mapping the substrate selectivity of new hydrolases using colorimetric screening: lipases from Bacillus thermocatenulatus and Ophiostoma piliferum, esterases from Pseudomonas fluorescens and Streptomyces diastatochromogenes. Tetrahedron Asymm 12:545–556
Qi M, Wang P, Selinger LB, Yanke LJ, Forster RJ, McAllister TA (2011) Isolation and characterization of a ferulic acid esterase (Fae1A) from the rumen fungus Anaeromyces mucronatus. J Appl Microbiol 110:1341–1350
Sanishvili R, Yakunin AF, Laskowskid RA, Skarina T, Evdokimova E, Doherty-Kirby A, Lajoie GA, Thornton JM, Arrowsmith CH, Savchenko A, Joachimiak A, Edwards AM (2003) Integrating structure, bioinformatics, and enzymology to discover function: BioH, a new carboxylesterase from Escherichia coli. J Biol Chem 278:26039–26045
Wang B, Tang XL, Liu J, Yu HW (2010) Escherichia coli BioH: a highly enantioselective and organic solvent tolerant esterase for kinetic resolution of sec-alcohols. Tetrahedron Lett 51:6360–6364
Xie XK, Wong WW, Tang Y (2007) Improving simvastatin bioconversion in Escherichia coli by deletion of bioH. Metab Eng 9:379–386
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Kang, L., Bai, Y., Cai, Y. et al. Discovery of novel feruloyl esterase activity of BioH in Escherichia coli BL21(DE3). Biotechnol Lett 38, 1009–1013 (2016). https://doi.org/10.1007/s10529-016-2075-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-016-2075-6