
Abstract
The predator Aphidoletes aphidimyza (Rondani) (Diptera: Cecidomyiidae) and the generalist entomopathogenic fungus Metarhizium brunneum Petch (Hypocreales: Clavicipitaceae) are effective biological control agents that can co-occur in pest management programmes. We exposed larvae to M. brunneum, on leaves and in soil, to explore possible outcomes of combining the two natural enemies. The number of emerging adults and their longevity were negatively affected by fungus in the soil. When exposed on leaves, adult emergence was unaffected but longevity was reduced. In choice experiments, females chose to lay eggs on fungus-free leaves: more control leaves had eggs and there were more eggs per leaf in the control. In no-choice experiments, the same frequency of females laid eggs in the treatment and control, but significantly more eggs were laid on the control leaves. Gravid A. aphidimyza can perceive M. brunneum and respond by choosing fungus-free oviposition sites, thus reducing contact between them.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Although the use of multiple natural enemies can have positive outcomes for biological control programmes (Labbe et al. 2009; Mohammed and Hatcher 2017), the reverse can also be true. Negative outcomes can occur if the entomopathogen infects and kills the beneficial arthropod or, at sub-lethal levels, if it negatively affects development time, fecundity or behaviour of beneficial arthropods (Poprawski et al. 1998). From an evolutionary perspective, pathogens and their hosts are constantly coevolving with each other (Roy et al. 2006). When a beneficial arthropod faces a significant risk of mortality due to a pathogen, then changes in behaviour may be selected that enable the beneficial arthropod to avoid pathogen contact and consequently reduce the risk of infection (Ormond et al. 2011). It has been shown that some beneficial arthropods do change their behaviour in response to pathogen presence. For example, some parasitoid species can discriminate between healthy and fungus-infected hosts (Rännbäck et al. 2015) and some predators detect and avoid leaves contaminated by a fungal pathogen (Meyling and Pell 2006).
Aphidoletes aphidimyza (Rondani) (Diptera: Cecidomyiidae) and Metarhizium species (Hypocreales: Clavicipitaceae) are commercially available biological control agents that may be used together to target the same or different pests, in horticultural and ornamental crops in both the greenhouse and the field (Meadow et al. 1985; Poprawski et al. 1999; Ansari et al. 2008). Aphidoletes aphidimyza is an aphid specialist used to control aphids in pest management programmes (Markkula et al. 1979). Metarhizium brunneum Petch is known as a generalist pathogen that infects a large number of insect species. However, differences in virulence among different Metarhizium species and Metarhizium brunneum sub-species have been demonstrated, as well as differences in virulence between aphid adults and nymphs, and among different aphid species (Jandricic et al. 2014). Lately, Metarhizium has been the focus of numerous studies since its generalist nature has the potential to negatively impact non-target arthropods. These studies have especially focused on the mode of application, since this fungus can be sprayed on to foliage or inoculated into the soil (Van Lenteren et al. 2003).
An aphidophagous predator may locate prey by responding to physical aspects of the habitat, or in response to semiochemicals produced by the host plant (Hodek 1993). Adult Aphidoletes aphidimyza feed on honeydew, and females use aphid honeydew as a cue to find the best oviposition sites for resulting offspring (Choi et al. 2004). This decision is of paramount importance for offspring survival and resulting levels of pest control because A. aphidimyza is only predacious in its larval stage. Females can locate a single aphid-infested plant amongst 75 un-infested plants in a greenhouse (Titi 1974), and lay eggs according to aphid density (Choi et al. 2004). Females are also able to recognize the presence of conspecifics in an oviposition site due to the production of an oviposition deterring pheromone (ODP) which deters females from laying eggs in the same location as other females. However, the role that the presence of an entomopathogen might have on the selection of oviposition sites by A. aphidimyza females has not been investigated.
It is known that A. aphidimyza adult longevity can be influenced by a number of different factors, such as the honeydew type (Watanabe et al. 2014) and prey density at the larval stage (Madahi et al. 2013). When combined in soil with entomopathogenic nematodes, adult emergence of A. aphidimyza was reduced (Powell and Webster 2004). Our previous greenhouse study revealed that the number of emerging A. aphidimyza adults was significantly decreased when they emerged from M. brunneum-treated soil compared with the control (Azevedo et al. 2017). Both the presence of conspecifics and the presence of neem insecticides also affect the A. aphidimyza females’ choice of oviposition site (Spollen and Isman 1996). However, how the presence of an entomopathogenic fungus on the oviposition site may influence A. aphidimyza females’ oviposition behaviour has not previously been investigated and is the focus of this study. Since A. aphidimyza larvae crawl on the leaf searching for potential prey and then drop into the soil to pupate, we assessed the effect of M. brunneum on larval mortality and adult longevity by exposing the larvae to fungus-treated leaves or fungus-inoculated soil. We also compared A. aphidimyza oviposition behaviour in the presence or absence of M. brunneum on oviposition sites, i.e. leaves with aphid colonies. We hypothesized that this could greatly impact both offspring survival and biological control success if the two natural enemies were used together. Besides addressing the lack of studies combining A. aphidimyza and entomopathogenic fungi, this research also enhances our understanding of the interactions between two biological control agents and the possibilities for their combined use in biological control programs.
Materials and methods
Plant material and host rearing
Pepper plants, Capsicum annuum L. (Solanaceae), were grown from seed under greenhouse conditions, and used for aphid rearing and experimental work. Two-month-old plants were transferred into cages and placed in a climate-controlled room (20 °C, L:D 16:8 h). The green peach aphid, Myzus persicae Sulzer (Homoptera: Aphididae), which is considered to be the most suitable prey for A. aphidimyza (Markkula and Tittanen 1985), was the host chosen for this study and its culture was obtained regularly from EWH BioProduction ApS (Tappernoje, Denmark).
Predator culture
Cohorts of A. aphidimyza were reared for us by EWH BioProduction at 23 ± 0.5 °C, 50–75% RH and L:D 16:8 h regime using the methods of Sigsgaard et al. (2012). Using cohorts ensured that all individuals tested were of the same age and had been reared under the same conditions. The company also mass-produces this species, complying with IOBC quality control guidelines for beneficial arthropods (Van Lenteren et al. 2003). We maintained the A. aphidimyza we received under exactly the same conditions prior to experimentation. Only third instar larvae were used in the longevity experiment but for the oviposition experiment we used newly-emerged adults obtained from pupae. Males and females were held together for 72 h prior to the oviposition experiment, and only visibly gravid females were selected for use.
Microbial inoculum
Metarhizium brunneum (KVL 12–19), which is the same genotype as the commercially-produced isolate GranMet/BIPESCO 5, was used in all experiments. Cultures were grown on 4% Sabouraud dextrose agar (SDA; Merck, Sweden) in 9 cm triple-vented Petri dishes at 20 ± 1 °C for 20 days. Conidia were harvested by flooding the cultures with sterile 0.05% Triton-X 100 (VWR, Sweden), and scraping with a sterile Drigalski spatula to dislodge conidia. The resulting suspension was transferred to a 50 ml stock tube. The concentration of the stock suspension was measured in a haemocytometer (Fuchs-Rosenthal 0.0625 mm2, depth 0.200 mm, VWR, Sweden). To assess conidial viability, germination tests were made. Suspensions were only used if germination was determined to be > 95%. Stock suspensions of conidia were refrigerated and used the same day they had been prepared.
Inoculation of soil and leaves
Soil was obtained from the University of Copenhagen experimental farm at Bakkegaarden, which has been managed as an organic farm for at least ten years. The soil was sieved through a 3 mm mesh and, for each experiment, 200 g was placed into a 10–15 l plastic bag. This was inoculated with a conidial suspension in 0.05% Triton X-100 to achieve a concentration of 5 × 106 conidia per gram of soil. The bag was closed and the contents kneaded thoroughly to distribute the conidia evenly throughout. The same volume of water containing 0.05% Triton X-100 was added to control soil. Soil was maintained at room temperature. Individual leaves, each with approximately 100 aphids, were detached from infested pepper plants and immediately submerged, sequentially, into 100 ml of 0.05% Triton-X 100 (control) or 100 ml of a 1 × 107 conidia per ml suspension of M. brunneum, for the control and treated groups, respectively. The leaves were left to air dry on tissue paper for 1 h. Control leaves were dipped, in the same way, into 0.05% Triton X-100 solution without conidia. Leaves were used in experiments within 1 h of dipping after pilot tests have showed that both the leaves and aphids could withstand the setup.
Longevity assessment
Effects of M. brunneum-treated soil on the longevity of A. aphidimyza
Soil (65 g) was placed into containers (155 ml) with the base covered with 5 ml water agar (1.5%) to ensure stable RH during experiments (95–97% RH). Mature larvae of A. aphidimyza (n = 10–30 per container) were placed into 22 containers containing soil treated with M. brunneum (n = 315 larvae in total) and 21 control containers (n = 305 larvae in total). The containers were closed with perforated lids and incubated at 23 ± 0.5 °C, 50–75% RH, in a L:D 16:8 h regime. After emergence, each A. aphidimyza midge was transferred individually into new containers (30 ml) containing a piece of filter paper dipped in a 1:10 water:organic honey solution as food. The base of each container was covered with 3 ml of 1.5% water agar to maintain a constant RH, and a little soil on the agar surface to prevent midges becoming stuck to the agar. Containers were closed with a perforated lid to allow ventilation, and incubated at the same temperature and light conditions as before. The filter paper with honey solution was changed every third day. For mycosis verification, dead midges were surface sterilized in 70% alcohol for 5 s, then rinsed in deionized water for an additional 5 s after which time they were incubated in sealed Petri dishes under moist conditions. Mycosis of M. brunneum on infected midges could be observed two days after death as evidenced by the hardening of the cadaver, emerged mycelia and green-coloured conidia. The number of days larvae spent in the soil, the day of emergence, sex, the number of days the emerging adults survived, and the number of dead with mycosis were recorded.
Effect of M. brunneum treated leaves on the longevity of A. aphidimyza
Fungus-inoculated aphid-infested leaves were placed individually in containers (155 ml). The base of each container was covered with 5 ml of water agar (1.5%) to ensure stable RH during experiments (95–97% RH), and untreated soil (65 g) to prevent larvae becoming stuck to the agar. Leaf petioles were fixed in the soil. A total of 49 containers were set up, each with mature larvae (n = 5) placed on the apex of the inoculated leaf. For the control, 44 containers were set up using triton-inoculated leaves, with the same number of larvae per replicate (n = 5). All containers were closed with perforated lids and incubated at 23 ± 0.5 °C, 50–75% RH, and a L:D 16:8 h regime. After 24 h, which was sufficient time to allow the mature larvae enter into the soil, the leaves were removed from the containers. After emergence, the same process was followed as the soil experiment, and the same variables were recorded.
Oviposition assessment
Aphidoletes aphidimyza adults
A cohort of adult midges was obtained by placing pupae (30) into each container (155 ml), five containers in total. The base of each container was coated with 5 ml water agar (1.5%) that was then covered with 1 cm of natural soil. Containers were closed with perforated lids and incubated at 23 ± 0.5 °C, 50–75% RH, and a L:D 16:8 h regime for 48 h. After this period, containers were opened and the first midges that had already emerged were discarded. The containers were incubated for a further 72 h. This way we ensured that all midges emerging during this period were mated.
Choice and no-choice experimental set-up
Individual leaves, each with approximately 100 aphids, were detached from infested pepper plants and immediately submerged, sequentially, into 100 ml of 0.05% Triton-X 100 (control) or 100 per ml of a 1 × 107 conidia per ml suspension of M. brunneum, for the control and treated groups, respectively. The leaves were left to air dry on tissue paper for 1 h. Each leaf petiole was inserted into a tube with water (5 cm high and 0.5 cm wide). This tube was placed in a 30-ml plastic container filled with 15 ml of water agar (1.5%). The water in the tube ensured that the petiole would remain turgid for longer and the soil placed on the surface of the water agar prevented adults from becoming stuck to the agar.
An A. aphidimyza female, gravid in appearance, and three males, all taken from the same container and aged less than 72 h, were placed in a transparent plastic container (10 cm × 16 cm × 16 cm), sealed with perforated lids, containing either one (no choice experiment) or two (choice experiment) pepper leaves. Each leaf constituted an oviposition site. Each oviposition site consisted of a pepper leaf with M. persicae aphids, treated or not with M. brunneum immediately before the trial. A total of 82 replicates of the choice experiment were performed, in which each container had two pepper plant leaves (one treated with M. brunneum and one untreated), and a total of 109 replicates of the no-choice experiment were performed, in which each container had a single leaf (55 treated and 54 control). After 48 h, the number of eggs laid by the females on each leaf were counted. The experiment was done at 23 ± 0.5 °C, 50–75% RH, and a L:D 16:8 h regime.
Data analysis
To analyse the effects of M. brunneum-treated soil on the length of time that it took for the A. aphidimyza to emerge from the soil, we compared both the mean emergence time across treatment groups (using a t test), and the proportion of midges that emerged across treatments (using a two-sample test for equality of proportions). To analyse the effects of M. brunneum-treated soil on the longevity of A. aphidimyza, a linear mixed effects model was used with the treatment group (M. brunneum or control) and the number of days that each larva spent in soil were included as fixed effects (along with an interaction term), and the container was included as a random effect. The model was estimated twice, once with the number of days spent in the soil taken as a continuous variable, and once with it taken as a factor, and the model with the number of days spent in the soil taken as a factor was selected on the basis of the Akaike information criterion. This model was then validated and the significance of each of the fixed effects and random effects was assessed using approximate likelihood ratio tests, with p < 0.05 considered statistically significant. Least-squares means were calculated to contrast (a) the longevity of the treatment group with the control group and (b) the longevity of midges in relation to how long each had spent in the M. brunneum treated soil.
To analyse the effects of M. brunneum-treated leaves on the length of time that it took for the A. aphidimyza to emerge from the soil, we compared the proportion of midges that emerged across treatments using a two-sample test for equality of proportions. To analyse the effects of M. brunneum-treated leaves on the longevity of A. aphidimyza, a linear mixed effects model was constructed, with the container included as a random effect and the treatment group (M. brunneum or control) as a fixed effect. The model was validated and we established the significance of both the fixed and random effects. Least-squares means were calculated to contrast the longevity of the treatment group with the control group.
For the oviposition study, a two-sample test for equality of proportions was used to test whether the proportion of eggs laid on treated leaves differed from the proportion laid on control leaves, and a Wilcoxon rank sum test was conducted to test whether the distribution of eggs was different across treatment groups. Both tests were performed separately for the choice and no-choice experiments. All analyses were carried out using R: A language and environment for statistical computing (R Core Team 2016).
Results
Longevity assessment
Effects of M. brunneum treated soil on the longevity of A. aphidimyza
There was no significant difference (t237 = 0.145, p = 0.89) in the length of time A. aphidimyza midges took to emerge from the fungus-treated soil (11.0 days; SE: 0.1) and untreated soil (11.0 days; SE: 0.1). The proportion of midges that emerged from the soil in the control containers (71%) was significantly higher than the proportion that emerged from the soil in the fungus-treated containers (53%) (\(\chi_{1}^{2}\) = 20.755, p < 0.001).
Based on our linear mixed effects model, we found that there was a statistically significant effect of both the treatment group (\(\chi_{1}^{2}\) = 15.660, p < 0.001) and the number of days that each larva spent in soil (\(\chi_{4}^{2}\) = 15.687, p = 0.003) on the longevity of the A. aphidimyza midges, but that their interaction was not significant (\(\chi_{4}^{2}\) = 5.727, p = 0.220). On average, our model predicted that A. aphidimyza midges that emerged from untreated soil would live 1.6 days longer (95% CI 0.8–2.4 days) than those that emerged from fungus-treated soil (Fig. 1, right panel). Aphidoletes aphidimyza midges that emerged from untreated soil lived longer than those that emerged from fungus-treated soil, no matter how long they had spent in the soil: overall, midges that spent a longer time in the soil did not live as long as those that emerged sooner and spent less time in the soil (Fig. 2).

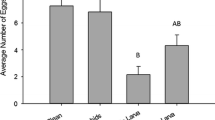
Effect of Metarhizium brunneum inoculated on leaves (left panel) and in soil (right panel) on the longevity of Aphidoletes aphidimyza adults. The least-squares means are shown (black dots) with 95% confidence intervals (black bars)

Effect of Metarhizium brunneum inoculated in soil on the longevity of Aphidoletes aphidimyza adults. The black circles indicate the least-squares means for the longevity of midges inoculated in the soil with M brunneum and the black triangles indicate the least-squares means for the control. The black bars indicate 95% confidence intervals
Metarhizium brunneum was pathogenic to A. aphidimyza midges when they were exposed to fungus applied to the soil in their final larval stage. Amongst the midges that emerged from fungus-treated soil and subsequently died, 15% supported mycosis by M. brunneum after incubation. No dead midges with mycosis were found in the control group.
Effect of M. brunneum treated leaves on the longevity of A. aphidimyza
There was no significant difference between the proportion of midges that emerged following fungus-exposure on leaves (61%) and the proportion that emerged from the control containers (57%) (\(\chi_{1}^{2}\) = 0.75791, p = 0.384). However, based on our linear mixed effects model, we found that there was a statistically significant effect of the treatment group (\(\chi_{1}^{2}\) = 18.360, p < 0.001) on the longevity of the A. aphidimyza midges. Our model estimated that the mean longevity of midges exposed to fungus on leaves when they were larvae was 6.4 days (SE: 0.3 days), which was 1.9 days (95% CI 1.1–2.8 days) shorter than the longevity of midges exposed to the untreated leaves (mean: 8.2 days; SE: 0.3 days) (Fig. 1, left panel). Additionally, 23% of A. aphidimyza midges previously exposed to fungus-treated leaves supported mycosis by M. brunneum after death. No infected midges were found in the control group.
Oviposition assessment
No-choice experiment
There was no significant difference in the percentage of leaves with eggs present between the treatment and the control (75% of fungus-treated leaves c.f. 83% of untreated leaves; \(\chi_{1}^{2}\) = 0.79, p = 0.37). The median number of eggs found on the fungus-treated leaves was 24, and on the untreated control leaves was 25. A Wilcoxon rank sum test did not reject the hypothesis that the distribution of eggs on the fungus treated and control leaves had the same mean (W = 1230, p = 0.42).
Choice experiment
Aphidoletes aphidimyza eggs were found on significantly more untreated leaves (93%) than fungus-treated leaves (65%) (\(\chi_{1}^{2}\) = 17.58, p < 0.001). Fewer eggs were found on the fungus-treated leaves (median: 9) than on the untreated control leaves (median: 21). A Wilcoxon rank sum test rejected the hypothesis that the distribution of eggs on the fungus treated and control leaves had the same distribution (W = 2075.5, p < 0.001).
Discussion
Our previous greenhouse study showed that M. brunneum, applied in soil did not influence A. aphidimyza emergence rate. However, after nine days (the end of the experiment), the number of midges was four times higher in M. brunneum-free cages, indicating an effect on longevity (Azevedo et al. 2017). In the present study, we found that M. brunneum negatively affected A. aphidimyza larval mortality when applied in soil, and longevity, when applied both in the soil and on the leaves. In our study, predators were exposed to inoculated leaves before aphids showed any signs of infection, and we did not incubate aphids to assess the infection level. However, another study assessing isolates of entomopathogenic fungi against greenhouse aphid found that M. brunneum F52, the same isolate as the one we used, had a great effect on aphid reproduction and mortality (Jandricic et al. 2014). These authors suggested that there was a barrier to effective aphid control using entomopathogenic fungi because the aphids needed to be killed before they become reproductive in order to prevent population growth (Jandricic et al. 2014). Aphidoletes aphidimyza is an important aphid natural enemy and a single larva can consume up to 190 nymphs during its development when in high aphid densities (Madahi et al. 2015). In our study, we found that the midges were susceptible to infection when exposed as larvae to M. brunneum, both on leaves and in soil. However, we also demonstrated that A. aphidimyza females actively avoided M. brunneum and chose pathogen-free oviposition sites to lay their eggs when they had a choice.
Powell and Webster (2004) observed that, when applied in soil, an entomopathogenic nematode caused over 50% larval mortality in A. aphidimyza populations under laboratory conditions and over 30% under greenhouse conditions, indicating a moderately harmful effect. In the current study, the emergence rate of A. aphidimyza midges was 1.4 times higher in the control that when exposed to M. brunneum applied in soil. Aphidoletes aphidimyza adults start mating on their first day of adult life (van Lenteren et al. 2002). However, we allowed the adults to mate for 72 h because we found more gravid females at that age. If A. aphidimyza adults are infected while emerging from soil inoculated with fungal inoculum, their fitness (i.e. mating success) may be severely affected if death occurs within the first three days. However, if adults have sufficient time to mate and females to oviposit before infection, their fitness may not be significantly affected by the fungal inoculum. We found that midges that had not been exposed to M. brunneum lived more than 1.5 days longer than those exposed to fungal inoculum. Furthermore, our longevity study showed that A. aphidimyza midges lived, on average, for 6.4 days following fungus exposure as larvae compared with 8.2 days in the control, which would provide enough time to mate and oviposit when adults.
To assess the side effects of M. brunneum on longevity of A. aphidimyza, we exposed the larvae in environments where fungal inoculum is commonly applied in biological control programmes, i.e. on the leaves or in the soil. Furthermore, A. aphidimyza spends its larval stages on leaves seeking prey, and then drops into the soil to pupate where it stays until emergence. This means that both places are important habitats in its life cycle. In both experiments we observed a decrease in adult longevity following exposure to fungal inoculum at the larval stage. Unexpectedly, we observed that when the soil was inoculated with M. brunneum, the time spent in soil during the pupal stage not only increased larval mortality but also the adult midges’ longevity. Interestingly, when applied on the leaves, M. brunneum did not affect larval mortality, although more adults became infected. Jackson and Jaronski (2009) found, after seven days, a cumulative larval mortality of over 50% when Tetanops myopaeformis were exposed to soil treated with M. brunneum F52. We observed no difference in vulnerability between females and males to the fungal inoculum. In fact, this could affect mating success since most of the males emerge before the females (van Lenteren et al. 2002) and this would shorten the mating period.
Aphidoletes aphidimyza was found to be susceptible to infection by M. brunneum, with up to 23% of dead midges showing mycosis even though exposure happened during the larval stage. Since pathogens and hosts are constantly coevolving with each other (Roy et al. 2006), it is not surprising that hosts have the ability to perceive and respond to the risk posed by a pathogen. Our results imply that A. aphidimyza females were able to recognize the presence of M. brunneum, and consequently it influenced where females laid their eggs and how many eggs were laid. However, this only happened when a choice was given. In a no-choice situation, the proportion of leaves with A. aphidimyza eggs was not significantly different, indicating either that females could not differentiate where to lay their eggs or that the urge to lay eggs overcame pathogen avoidance behaviour. Despite this, the number of eggs was higher on fungus-free leaves than fungus-treated leaves. These findings add to previous findings that A. aphidimyza females can also perceive the risk of other predators, such as Chrysoperla carnea and Coccinella septempunctata, and lay fewer eggs in their presence (Ruzicka and Havelka 1998). Females are also able to recognize the presence of conspecific eggs and larvae, and lay fewer eggs in the presence of conspecifics (Ruzicka and Havelka 1998; Guo et al. 2014). The parasitoid Trybliographa rapae also had its oviposition behaviour affected by M. brunneum: it was able to perceive the presence of the fungus and laid more eggs in healthy larvae than in infected ones (Rännbäck et al. 2015).
The present study examined, for the first time, the potential impact of M. brunneum on the larval and soil-dwelling stages of the predatory gall midge A. aphidimyza and on its oviposition behaviour, under laboratory conditions. The experimental settings simulated a worst-case scenario because A. aphidimyza was exposed to very high levels of fungus. Our experiments revealed that, even when A. aphidimyza was exposed to M. brunneum in its larval stage, they were still able to pupate, emerge as adults and live for a considerable time.
Aphidoletes aphidimyza females were not able to differentiate between neem insecticides-treated and control leaves and no difference was found in the number of eggs laid by females on both leaves, even though females exposed to the neem insecticide had a greater mortality than the control (Spollen and Isman 1996). Our study shows that A. aphidimyza females are likely to have adapted traits to avoid pathogens on the oviposition site. The findings reported here showed that A. aphidimyza females are able to perceive the risk posed by M. brunneum and react by choosing a pathogen-free site for their offspring. For biocontrol purposes, the combination of both biological agents can be considered acceptable since in a situation where Metarhizium brunneum was sprayed on the vegetation, A. aphidimyza female is able to choose an oviposition site where the pathogen did not reach, therefore diminishing the risk of infection for its offspring.
References
Ansari M, Shah F, Butt T (2008) Combined use of entomopathogenic nematodes and Metarhizium anisopliae as a new approach for black vine weevil, Otiorhynchus sulcatus, control. Entomol Exp Appl 129:340–347
Azevedo AGCd, Steinwender BM, Eilenberg J, Sigsgaard L (2017) Interactions among the predatory midge Aphidoletes aphidimyza (Diptera: Cecidomyiidae), the fungal pathogen Metarhizium brunneum (Ascomycota: Hypocreales), and maize-infesting aphids in greenhouse mesocosms. Insects 8:44
Choi MY, Roitberg BD, Shani A, Raworth DA, Lee GH (2004) Olfactory response by the aphidophagous gall midge, Aphidoletes aphidimyza to honeydew from green peach aphid, Myzus persicae. Entomol Exp Appl 111:37–45
Guo H, Meng L, Wang Y, Zheng L, Li B (2014) Oviposition behavior of the predatory midge Aphidoletes aphidimyza in response to aphid patch quality. J Insect Behav 27:816–825
Hodek I (1993) Habitat and food specificity in aphidophagous predators. Biocontrol Sci Technol 3:91–100
Jackson MA, Jaronski ST (2009) Production of microsclerotia of the fungal entomopathogen Metarhizium anisopliae and their potential for use as a biocontrol agent for soil-inhabiting insects. Mycol Res 113:842–850
Jandricic S, Filotas M, Sanderson J, Wraight S (2014) Pathogenicity of conidia-based preparations of entomopathogenic fungi against the greenhouse pest aphids Myzus persicae, Aphis gossypii, and Aulacorthum solani (Hemiptera: Aphididae). J Invertebr Pathol 118:34–46
Labbe R, Gillespie D, Cloutier C, Brodeur J (2009) Compatibility of an entomopathogenic fungus with a predator and a parasitoid in the biological control of greenhouse whitefly. Biocontrol Sci Technol 19:429–446
Madahi K, Sahragard A, Hossieni R (2013) Prey density dependent life table of Aphidoletes aphidimyza Rondani (Dip.: Cecidomyiidae) feeding on Aphis craccivora Koch (Hem.: Aphididae) under laboratory conditions. J Plant Prot Res 53:253–262
Madahi K, Sahragard A, Hosseini R (2015) Predation rate and numerical response of Aphidoletes aphidimyza feeding on different densities of Aphis craccivora. Biocontrol Sci Technol 25:72–83
Markkula M, Tittanen K (1985) Biology of the midge Aphidoletes and its potential for biological control. In: Hussey NW, Scopes N (eds) Biological pest control: the glasshouse experience. The Glasshouse Experience, Blandford, pp 78–81
Markkula M, Tiitanen K, Hamalainen M, Forsberg A (1979) The aphid midge Aphidoletes aphidimyza (Diptera, Cecidomyiidae) and its use in biological control of aphids. Entomol Fenn 45:89–98
Meadow R, Kelly W, Shelton A (1985) Evaluation of Aphidoletes aphidimyza [Dip.: Cecidomyiidae] for control of Myzus persicae [Hom.: Aphididae] in greenhouse and field experiments in the United States. Entomophaga 30:385–392
Meyling NV, Pell JK (2006) Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol Entomol 31:162–171
Mohammed AA, Hatcher PE (2017) Combining entomopathogenic fungi and parasitoids to control the green peach aphid Myzus persicae. Biol Control 110:44–55
Ormond EL, Thomas AP, Pell JK, Freeman SN, Roy HE (2011) Avoidance of a generalist entomopathogenic fungus by the ladybird, Coccinella septempunctata. FEMS Microbiol Ecol 77:229–237
Poprawski TJ, Legaspi JC, Parker PE (1998) Influence of entomopathogenic fungi on Serangium parcesetosum (Coleoptera: Coccinellidae), an important predator of whiteflies (Homoptera: Aleyrodidae). Environ Entomol 27:785–795
Poprawski T, Parker P, Tsai J (1999) Laboratory and field evaluation of hyphomycete insect pathogenic fungi for control of brown citrus aphid (Homoptera: Aphididae). Environ Entomol 28:315–321
Powell JR, Webster JM (2004) Interguild antagonism between biological controls: impact of entomopathogenic nematode application on an aphid predator, Aphidoletes aphidimyza (Diptera: Cecidomyiidae). Biol Control 30:110–118
Rännbäck L-M, Cotes B, Anderson P, Rämert B, Meyling NV (2015) Mortality risk from entomopathogenic fungi affects oviposition behavior in the parasitoid wasp Trybliographa rapae. J Invertebr Pathol 124:78–86
R Core Team (2016) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Roy H, Steinkraus D, Eilenberg J, Hajek A, Pell J (2006) Bizarre interactions and endgames: entomopathogenic fungi and their arthropod hosts. Annu Rev Entomol 51:331–357
Ruzicka Z, Havelka J (1998) Effects of oviposition-deterring pheromone and allomones on Aphidoletes aphidimyza (Diptera: Cecidomyiidae). Eur J Entomol 95:211–216
Sigsgaard L, Steinwender BM, Dimitrova D, Hansen EW, Eilenberg J (2012) Technical protocol for tests of direct and indirect side effects of new BCA formulations on non-target invertebrates. http://inbiosoil.uni-goettingen.de/fileadmin/INBIOSOIL/Deliverables/3.1%20Defined%20technical%20protocol%20for%20test%20of%20side%20effects%20....pdf. Accessed 10 Nov 2016
Spollen KM, Isman MB (1996) Acute and sublethal effects of a neem insecticide on the commercial biological control agents Phytoseiulus persimilis and Amblyseius cucumeris (Acari: Phytoseiidae) and Aphidoletes aphidimyza (Diptera: Cecidomyiidae). J Econ Entomol 89:1379–1386
Titi AE (1974) Auswirkung von der räuberischen gallmücke Aphidoletes aphidimyza (Rond.)(Itonididae: Diptera) auf blattlauspopulationen unter glas. Zeitschrift für Angewandte Entomologie 76:406–417
van Lenteren JC, Schettino M, Isidoro N, Romani R, Schelt J (2002) Morphology of putative female sex pheromone glands and mating behaviour in Aphidoletes aphidimyza. Entomol Exp Appl 102:199–209
van Lenteren JC, Babendreier D, Bigler F, Burgio G, Hokkanen H, Kuske S, Loomans A, Menzler-Hokkanen I, van Rijn P, Thomas M (2003) Environmental risk assessment of exotic natural enemies used in inundative biological control. BioControl 48:3–38
Watanabe H, Katayama N, Yano E, Sugiyama R, Nishikawa S, Endou T, Watanabe K, Takabayashi J, Ozawa R (2014) Effects of aphid honeydew sugars on the longevity and fecundity of the aphidophagous gall midge Aphidoletes aphidimyza. BioControl 78:55–60
Acknowledgements
The study was part of the EU project INBIOSOIL (Grant agreement 282767), funded by means of the 7th Framework Programme of the European Union. The authors would like to thank Erik W. Hansen and Desislava Dimitrova, EWH BioProduction for the cohorts; the laboratory technician Kristian Hanse; Judith K. Pell for the support with the proofreading. We also thank two anonymous reviewers for comments that helped us improve a previous version of this manuscript. This study was supported by the EU FP7 INBIOSOIL project and Science without Borders—Brazil (1260-13-8).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Marta Montserrat.
Rights and permissions
About this article

Cite this article
de Azevedo, A.G.C., Stuart, R.M. & Sigsgaard, L. Presence of a generalist entomopathogenic fungus influences the oviposition behaviour of an aphid-specific predator. BioControl 63, 655–664 (2018). https://doi.org/10.1007/s10526-018-9889-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-018-9889-1