
Abstract
Despite their importance as invasive species, there has been a hesitation to target grasses in classical biocontrol. This historic bias appears to be changing with multiple active research and release programs. Similarly, biocontrol workers appear to avoid targeting species with native congeners. These biases appear inappropriate as the ecological and entomological literature provide abundant evidence for sub-genus specificity for many herbivores, including those attacking grasses. The biocontrol program targeting Phragmites australis (Cav.) Trin. ex Steud (Poaceae) provides an informative example with endemic subspecies in North America and many sub-genus specific herbivores, including potential European control agents. Grasses and target weeds with congeneric native species require rigorous host range testing, similar to all other targets in current weed biological control programs. Furthermore, it appears prudent to ask petition reviewers and regulatory agencies to abandon their focus on results of no-choice studies and to distinguish between trivial feeding and demographic impacts.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Two widely-held tenets of the current practice in weed biocontrol are: avoid targeting grasses and avoid plant pests with congeneric species (Pemberton 2000). Although there are 2176 species of invasive grasses worldwide, biological control of invasive grasses has historically been avoided because of the economic importance of many grass species and an expectation that a general lack of species-specific secondary chemistry among grasses would result in a lack of host specific herbivores and hence threaten safety of non-target species (Witt and McConnachie 2003). Pemberton (1996) summarized the status of grass biocontrol: “there has never been a biological control program against a grass weed and it is unlikely that one will be started against these plants in the future. The enormous value that grass family crops (like corn, wheat, rice, barley, sugarcane, etc.) have, and also the belief that natural enemies of grasses have less specificity than those that feed on other plants have prejudiced this approach.”
There are no good underlying ecological reasons why weed biocontrol scientists have failed to target grasses (Witt and McConnachie 2003). Grasses may be less structurally complex leading to a reduced diversity of feeding niches, and they may not offer the same array of secondary chemistry as many other herbaceous species, but they have their own highly specific insect (particularly Diptera and Hymenoptera) and pathogen communities (Witt and McConnachie 2003), some of which are major pests of crops, such as in rice or sugarcane (Lou et al. 2013; Settle et al. 1996; Strong et al. 1977; Zhu et al. 2000). Furthermore, grasses through mutualistic endophytes and silicone-based defenses (Djamin and Pathak 1967; Gali-Muhtasib et al. 1992; Hartley and DeGabriel 2016; Massey et al. 2007) have major impacts on vertebrate and invertebrate herbivores comparable to their broadleaf counterparts (Moore et al. 2014). The charge for biocontrol scientists is to assess whether host-specific species with potential to affect host plant demography are available among grass herbivores and pathogens.
Multiple Asian grasses are invasive in the USA and all of them have several species (mostly pathogens) that appear to have restricted host ranges (Zheng et al. 2004) providing opportunities to implement biocontrol. In 2000 the planthopper Prokelisia marginata (van Duzee) (Delphacidae) was introduced into Washington State in an effort to reduce populations of smooth cordgrass, Spartina alterniflora Loisel (Poaceae) (Grevstad et al. 2003). This was followed in 2009 and 2010 with releases of the gall wasp Tetramesa romana Walker (Eurytomidae) and the scale Rhizaspidiotus donacis Leonardi (Diaspididae) targeting giant reed, Arundo donax L. (Poaceae) (Racelis et al. 2009) with promising early results (Goolsby et al. 2016). Research is underway to assess feasibility of targeting cogongrass (Imperata cylindrica (L.) P. Beauv.) (Poaceae) and Japanese stiltgrass Microstegium vimineum (Trin.) A. Camus (Poaceae) (Nestory 2016; Overholt et al. 2016). The importance of grasses as invasive species and monetary expenditures currently dedicated to their control (Brooks et al. 2004; Martin and Blossey 2013b; Rossiter et al. 2003) clearly warrant a continued focus on evaluating potential for biocontrol. In this paper, we discuss opportunities to develop safe and effective classical biological control programs when specificity of biocontrol agents requires sub-generic levels of host specificity. In particular, we use ongoing research to develop biocontrol for introduced lineages of common reed, Phragmites australis (Cav.) Trin ex Steud (Poaceae) in North America, to illustrate these prospects.
Host specificity of herbivores at the sub-species level
Pemberton (2000) pointed out difficulties targeting introduced plants with native congeners, but noted that risks can be reliably evaluated. In fact, specificity of herbivores frequently occurs below species level (Fritz and Simms 1992) and in a recent review Whitham et al. (2012) show that genotype specificity is a widespread phenomenon listing examples from 29 genera within 15 plant families. The ecological and entomological literature on host plant resistance to insects is replete with examples of selectivity in response to genotypic variation in structural (Karley et al. 2016) or chemical defenses (Agrawal 2011; McGuire and Johnson 2006; Moore et al. 2014) and particularly including grasses. These variations in resistance have been known for a long time (Painter 1951) and allow resistance breeding in crops, for example in rice, where attack by 32 different insect herbivores is controlled by variation in resistance among strains and inter-specific rice hybrids (Heinrichs 1986). Other plant–herbivore systems expressing varietal resistance occur in wheat, oats, barley and maize, including attack of wheat by Hessian fly (Mayetiola destructor Say; Cecidomyiidae), or wheat stem sawfly (Cephus cinctus Norton; Cephidae) and many others (Painter 1951). Intra-specific variation in herbivore resistance is a common phenomenon and well-known for many crops in the Poaceae. Consequently, if we were looking for a genotype or subspecies-specific biocontrol agent, we would have several to choose from within these grasses. We acknowledge that resistance is not always complete or permanent and herbivores show gradients in attack among different genotypes (McGuire and Johnson 2006). The underlying reasons for this host or genotype specialization, previously assumed to be based on performance trade-offs, now appears to involve complex interactions of genetic architecture and multitrophic interactions (Forister et al. 2012). But the evolutionary and ecological opportunity of sub-species level specificity is widespread and there is no a priori reason to reject grasses or species with close relatives as targets of biocontrol investigations due to beliefs that safety of non-targets is elusive in these systems.
The genus Solanum provides an interesting example with many important crops (potatoes, tomatoes, eggplant) and native species yet also problematic invasive species. In South Africa Solanum eleagnifolium Cav. and S. mauritianum Scopoli (Solanaceae) were targeted by releasing multiple agents including leaf beetles Leptinotarsa texana (Schaeffer) and L. defecta (Stål) (Chrysomelidae), a lace bug Gargaphia decoris Drake (Tingidae) and a flower bud weevil Anthonomus santacruzi Hustache (Curculionidae) (Hakizimana and Olckers 2013; Olckers 2000; Olckers and Hulley 1994). The herbivores established and recent evidence appears to suggest heavy and widespread impact on S. eleagnifolium (Winston et al. 2014). But with the exception of a single attack of eggplant at a single farm (Olckers, pers. comm. to Hariet Hinz) there are no reports of non-target feeding in the field (Olckers 2011), supporting decisions to release insects that show little feeding in host specificity tests on native relatives and crops. Despite important crop and native relatives, the pasture weed tropical soda apple, Solanum viarum Dinal (Solanaceae) is now under widespread control by the leaf beetle Gratiana boliviana Dunal (Chrysomelidae) in the USA (Diaz et al. 2014). Clearly, safety of non-target species is a function of host specificity of biocontrol agents and existence of closely related native species or important crops should not preclude investigations per se (Hinz et al. 2014).
We are only aware of two examples in weed biocontrol when sub-species level specificity was recorded. The first is rush skeletonweed (Chondrilla juncea L.) (Asteraceae) where existence of three different leaf morphotypes precluded success using the mite Aceria chondrillae Canestrini (Eriophyidae) and the rust fungus Puccinia chondrillina (Bubak & Syd.) (Uredinales). Both species show specificity at the morphotype level, limiting their ability to control all Chondrilla populations (Charudattan 2005; Winston et al. 2014; and references therein), although recent molecular work suggests a more complex and variable background of different genotypes, plant morphology and insect attack (Gaskin et al. 2013). The second example is Chrysanthemoides monilifera ssp. monilifera (L.) T. Norl. (Boneset) and C. monilifera ssp. rotundata (DC.) T. Norl. (bitou bush) (Asteraceae). Both have sub-species specific biocontrol agents in Australia and New Zealand (Winston et al. 2014). Furthermore, genotypic diversity of Lantana camara L. (Verbenaceae) has been an obstacle to achieving more widespread biological control of the species (Crawley 1989) and there are additional examples of biotype or population variation in preference of weed biocontrol agents and their ability to control populations or biotypes (Winston et al. 2014) demonstrating prevalence of herbivore specificity or preference below the species level assigned by taxonomists.
Introduced Phragmites in North America
Impacts of introduced P. australis on native biota
Introduced P. australis genotypes in North America can dominate wetland plant communities and this invasive success has been linked to shoreline development in New England (Bertness et al. 2002; Burdick and Konisky 2003; Silliman and Bertness 2004), prevalence of agriculture and nutrient loading, (Kulmatiski et al. 2010; Sciance et al. 2016), salt tolerance (Vasquez et al. 2005), ability to exploit disturbances and dispersal corridors along roadsides (Brisson et al. 2010; Jodoin et al. 2008), and hydrology changes (Hudon et al. 2005). Wetland managers have responded with aggressive treatments due to concerns over detrimental impacts of P. australis on native North American biota (Hazelton et al. 2014; Marks et al. 1994; Martin and Blossey 2013b).
However, evidence for negative impacts on native biota is mixed and few long-term data exist to separate impacts of P. australis invasion from other stressors affecting wetlands such as land use history, changes in hydrology, nutrient loading, etc. (Kettenring et al. 2012). Introduced P. australis is thought to threaten endangered species such as the pallid sturgeon (Scaphirhynchus albus) (Acipenseridae), the whooping crane (Grus americana) (Gruidae), the northern Great Plains population of the piping plover (Charadrius melodus) (Ciconiiformes), and the interior least tern (Sterna antillarum athalassos) (Laridae) in the Platte River (National Research Council 2004). Some, but not all, native wetland plants are negatively affected by P. australis encroachment (Crocker et al. 2017), as are some specialized marsh birds (Benoit and Askins 1999; Whyte et al. 2015) but not others (Parsons 2003). Similarly some turtles (Bolton and Brooks 2010), and small fish such as Fundulus spp. (Fundulidae) (Able and Hagan 2000) show population reductions in response to invasive P. australis. In contrast, several amphibians (Cohen et al. 2012, 2014; Martin and Blossey 2013a; Rogalski and Skelly 2012), some crustaceans (Able and Hagan 2000), Northern pike, Esox esox (Esocidae) (Larochelle et al. 2015), and decomposition processes and macroinvertebrate communities (Kennedy et al. 2012) appear to show no negative response to P. australis encroachment.
The anticipated negative ecological impacts, and reduced aesthetic and property values—particularly along the Atlantic Coast and the Great Lakes (Clawson and Duthinh 2015)—resulted in the designation of introduced P. australis as a noxious weed in Alabama, Vermont, Washington, Nebraska, British Columbia, and Alberta and the species is also listed as invasive or prohibited in Connecticut, Massachusetts, and South Carolina (USDA NRCS 2017). Introduced P. australis also has a long history of failed attempts to manage the species using chemical and physical means (Marks et al. 1994; Martin and Blossey 2013b). Only restoration of regular tidal inundation with full strength seawater can successfully suppress P. australis in coastal areas (Chambers et al. 1998). Short-term suppression of the species is possible using herbicides, while eradication is extremely difficult and only achievable for small populations not exceeding a few hundred square meters (Lombard et al. 2012; Quirion et al. 2018). Annual expenditures for herbicide use to treat P. australis in the USA alone reached US $4–5 million before 2010 (Martin and Blossey 2013b).
Development of biological control of invasive P. australis: research in North America
We initiated a classical biocontrol program in 1998. A literature review identified > 150 European herbivores, pathogens and inquilines with > 20 species listed as monophagous (Tewksbury et al. 2002). When detailed investigations began, the status of P. australis as native or introduced was contested, but now taxonomists recognize three distinct lineages, of which at least one is endemic to North America (Saltonstall 2002, 2016; Saltonstall et al. 2004). This raises the necessity that any biocontrol agent for invasive P. australis requires sub-species level host specificity.
The most widespread and regionally abundant of three different P. australis lineages in North America are introduced European genotypes (Saltonstall 2002, 2016) which are widespread (Fig. 1b) from Canada to the Gulf Coast and southern California (Saltonstall and Meyerson 2016). An endemic North American sub-species P. australis americanus Saltonstall, P.M. Peterson & Soreng is genetically and morphologically diverse and occurs from Canada to northern Mexico (Colin and Eguiarte 2016; Saltonstall and Meyerson 2016) (Fig. 1a). The Gulf Coast lineage P. australis berlandieri (E. Fourn.) C.F. Reed occurs in the southern USA south through Mexico into South America (Fig. 1c) but the status of the lineage as native or introduced is unresolved (Colin and Eguiarte 2016; Lambertini et al. 2012; Saltonstall and Meyerson 2016). European genotypes are considered invasive with widespread detrimental impacts (Marks et al. 1994), while populations of native genotypes have been lost (Saltonstall 2002) and are considered of conservation concern in eastern states. Hybridization between European and North American lineages, long suspected due to common garden evidence (Meyerson et al. 2008) has been confirmed in New York and Nevada (Saltonstall et al. 2014, 2016). The range of the lineages overlap (Fig. 1) poses a constraint on the Phragmites biocontrol program. Potential biocontrol agents are required to be specific to invasive genotypes and not affect demography or reduce populations of endemic sub-species to be acceptable to regulatory agencies and wetland managers (Martin and Blossey 2013b).

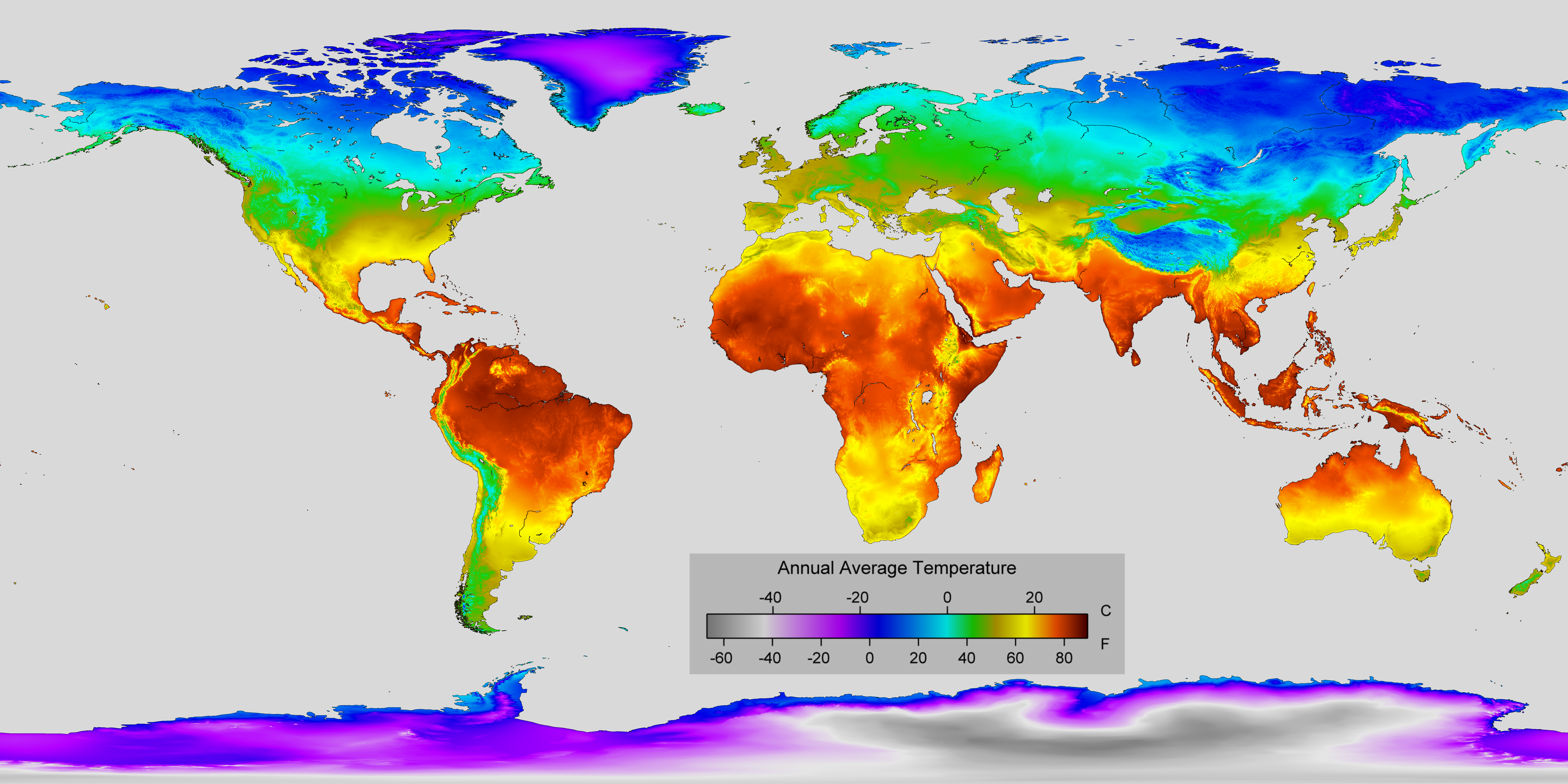
Distribution (hatched areas) of a native P. australis americanus, b introduced P. australis australis, and c P. australis berlandieri in North America. Maps drawn and updated based on Saltonstall and Meyerson (2016), USDA PLANTS database (https://plants.usda.gov), and our own records using fied surveys and submitted samples. Please note that occurrences at range margins and areas where introduced P. australis is currently advancing have great uncertainty due to sampling limitations. Expected northern and southern distribution of A. geminipuncta and A. neurica are indicated by parallel lines across North America in (a) and are based on climate conditions in their native European ranges (https://upload.wikimedia.org/wikipedia/commons/9/92/Annual_Average_Temperature_Map.png)
There is abundant evidence of strong sub-species level selectivity in the field among three native North American Phragmites herbivores and 18 European species already accidentally introduced to North America (Blossey 2003a; Blossey and Casagrande 2016). Three native Phragmites specialists (Calamomyia phragmites Felt, Cecidomyiidae; Thrypticus willestoni (Wheeler), Dolichopodidae; and Ochlodes yuma (Edwards), Hesperiidae) all occur exclusively on native P. australis americanus and have never been recorded on introduced or Gulf Coast genotypes. Ten of 18 accidentally introduced Phragmites herbivores are specialists on Phragmites and the others are polyphagous. Among these specialists Lasioptera hungarica Möhn (Cecidomyiidae), Giraudiella inclusa (Frauenfeld) (Cecidomyiidae), Lipara pullitarsus Doskocil & Chvála (Chloropidae) and Rhizedra lutosa (Hübner) (Noctuidae) are only recorded feeding on introduced P. australis, although L. hungarica also attacks hybrids in New York (Saltonstall et al. 2014). Thus seven of 13 Phragmites specialists presently known to be in North America show specificity at the sub-species level as currently defined by taxonomists and the remaining six species are found on both native and introduced P. australis.
Specialist herbivores of Phragmites in Europe
Nine European insect herbivores were chosen for further investigations as potential biocontrol agents based on their feeding niche, damage inflicted, and reported host specificity (Tewksbury et al. 2002). This list was narrowed to the four most promising agents (Häfliger et al. 2005, 2006a, b) and for two species of noctuid moths, Archanara geminipuncta (Haworth) and A. neurica (Hübner) (Noctuidae). Extensive host specificity testing is now complete (Blossey et al. 2018). We focus on these two stem mining univoltine noctuids because they are the most widespread, abundant, and with the largest additive impact on P. australis (Häfliger et al. 2006b).
Detailed life histories are well known with adults flying in July/August and eggs laid under leaf-sheaths where they overwinter until larvae hatch in early spring (Häfliger et al. 2006b). Larvae feed internally in shoot tips and upper internodes and need to change stems multiple times before pupating in lower sections of P. australis stems. In Europe, large population fluctuations are reported with outbreak cycles of 3–4 years when stem damage may reach 100% (Michel and Tscharntke 1993; Tscharntke 1990). Shoot heights and above-ground biomass reductions can be substantial, typically 20–60% (Häfliger et al. 2006a) but occasionally reaching 100% and intensity of damage may be a function of latitude (Tscharntke 1990, 1992b, 1999). Due to extensive shoot damage in Europe, A. geminipuncta is considered a keystone species influencing trophic structure and abundance of reed biota from insects to birds (Tscharntke 1989, 1992a, 1999).
Our host specificity testing followed established guidelines and was conducted in quarantine at the University of Rhode Island as well as in the native range at CABI in Switzerland (Blossey et al. 2018) with a particular focus on ability of neonate larvae to penetrate or survive in stems of other plant species. Larvae of A. geminipuncta and A. neurica were unable to survive in any of the 43 non-target test plant species, but both were able to complete development in invasive P. australis and native P. australis americanus (under no-choice conditions). We followed up with oviposition choice experiments on potted plants at CABI from 2011 to 2015, allowing mated females to choose among different European, invasive North American, and endemic North American genotypes that were offered in monospecific and mixed arrangements. Under these experimental conditions A. neurica laid 6.7% (39 of 584) and A. geminipuncta laid 6.5% (32 of 503) of their eggs on P. australis americanus with the remainder nearly evenly split among ancestral European and invasive North American P. australis (Blossey 2014; Blossey et al. 2013, 2018).
An overview of factors mitigating potential risks
Both A. neurica and A. geminipuncta lay few eggs on the endemic native sub-species P. australis americanus and their larvae are able to penetrate stems and develop successfully. We consider demographic risks for P. australis americanus to be low for the following reasons:
Leaf-sheaths
One important trait shared by native P. australis americanus across the range of the lineage are loose leaf-sheaths that fall off during fall and winter, while leaf-sheaths on invasive P. australis are retained, often for multiple years after senescence (Blossey 2003b; Ward 2010). What potential ecological benefits are derived from loose leaf-sheaths in North America is unknown, but it obviously deters oviposition by insects, such as A. geminipuncta and A. neurica. Females avoid loose leaf-sheaths, a behavioral trait which largely restricts attack to introduced P. australis (Blossey et al. 2013). Eggs attached to leaf-sheaths that fall off the plant show reduced survival rates (Blossey et al. 2018), hence choosing tight fitting leaf-sheaths for oviposition confers fitness advantages.
Larval shoot change
All Archanara larvae need to change shoots to complete development. The endemic P. australis americanus generally, but not always, occurs in small isolated populations with greatly reduced stem densities intermixed with many other wetland species. The probability for foraging larvae to successfully locate new stems is likely to be greatly reduced as stem densities decrease (Crawley and Gillman 1989), although no quantitative data on larval dispersal behavior are currently available for our system.
Demography
The goal of invasive P. australis management is to reduce negative impacts on native biota and ecosystem processes. To reach this goal it is generally assumed that management needs to greatly decrease abundance of the invasive lineage. Substantial stem mortality and reduction in rhizome growth is required to affect performance (Häfliger et al. 2006a) and subsequently demography in a species that is able to spread by seed and via rhizome growth. The low numbers of eggs laid on native stems is unlikely to result in substantial stem mortality or a demographic threat (Häfliger et al. 2006a; Myers and Sarfraz 2017).
Rapid evolution is unlikely to overcome resistance traits in P. australis americanus
We acknowledge the possibility that over evolutionary time different preferences may evolve, but the experimentally determined fundamental host range has been stable for weed biocontrol agents (Marohasy 1996; Suckling and Sforza 2014; van Klinken and Edwards 2002) even when species were re-tested decades after their introduction (Arnett and Louda 2002). Introduction of ecological novelty, for example by introduced plants, provides ample opportunity for herbivores to expand their realized host range as expressed in the field (Jahner et al. 2011) and this is often interpreted as a host shift or rapid evolution of host specificity (Secord and Kareiva 1996). But, in most cases, model organisms used to assess such host shifts, or changes in preferences based on feeding experience of larvae or adults are oligo- or polyphagous species (Anderson et al. 2013; Austel et al. 2014; Carrasco et al. 2015; Liu et al. 2005; Profitt et al. 2015). Their diet specialization is quite different from biocontrol agents that are typically monophagous in the field, and where we experimentally determine their fundamental host range.
Diets of insect herbivores change over time and both generalists and specialists may acquire new hosts (Futuyma and Agrawal 2009), and this acquisition may best be evaluated using a phylogenetically informed trait-based approach (Barrett and Heil 2012; Pearse et al. 2013; Pearse and Hipp 2009). The vast majority of phytophagous insects show “phylogenetic conservatism” retaining their association with plant taxa over millions of years (Futuyma and Agrawal 2009). Biocontrol agents passing through host range testing, as far as we can tell from decades of observation and study, appear particularly “conservative” (Blossey et al. 2001; Hinz et al. 2018; Suckling and Sforza 2014).
In the particular case of P. australis americanus we consider that this lineage is particularly safe from potential rapid evolutionary adaptations of the herbivores should introduction of A. geminipuncta and A. neurica be granted because the response to loose leaf-sheaths is behavioral, not physiological. This plant-specific trait actually adds to the safety of native genotypes. Evidence from host shifts of herbivores suggests that adaptations to process plants with different defense chemistry, or changes in female recognition of host plants that allow successful larval development are relatively “easy routes” to incorporate novel hosts (Pearse et al. 2013). However, recognition and palatability are only two of the many traits that determine the ability of herbivores to utilize plants (Barrett and Heil 2012). Larvae of both Archanara species can be reared in native P. australis americanus stems. This lineage is clearly part of the physiological host range and there appear to be few performance penalties (Blossey et al. 2018). However, as detailed above, loose leaf-sheaths can be considered a resistance trait preventing oviposition by Archanara species. This plant trait of P. australis americanus is not affected by selection operating at the herbivore level, therefore there is no pathway for rapid selection or evolutionary change operating at the level of the insects to favor genotypes that would oviposit under loose leaf-sheaths while maintaining herbivore fitness.
Ability for local management
While we believe that limited stem damage by the two Archanara species on endemic genotypes would not amount to a demographic threat, the fact that eggs overwinter under leaf-sheaths that adhere to shoots (or drop to the ground) affords a potential ability to manage a local population using controlled burns should it be necessary (Branson et al. 2015; Brix et al. 2014; Vogel et al. 2010). Land managers could burn or mow/mulch the stand, thereby locally managing the noctuid populations.
Distribution and climate
The Gulf Coast subspecies grown in pots in our common garden in Switzerland developed only extremely thin side stems. We therefore lack reliable oviposition preference data that we have for P. australis americanus. However, the European distributions of A. geminipuncta and A. neurica (Fig. 2) offer some limited evidence that these species may not be able to thrive as far south as P. australis berlandieri, a sub-species restricted to areas where average annual temperatures range from 17 to 21 °C. In Europe, neither moth is found where average temperatures exceed 15 °C and based solely on climate data, we expect the herbivores, if released, to establish in temperate North America (Fig. 1a). However, prediction of future ranges based on ancestral distributions is notoriously unreliable (Shabani et al. 2016) and species respond, including evolutionarily (Bean et al. 2012) to more than climate. Thus caution is appropriate in anticipating future distributions. Only detailed studies after release will be able to assess how well home-range climate data may predict novel range distributions.

Source: http://www.fauna-eu.org
Distribution (hatched areas) of Archanara geminipuncta (a) and A. neurica (b) in western, central and eastern Europe. Please note that occurrences at range margins have great uncertainty due to sampling limitations.
Regulatory considerations
Our case study on P. australis shows that sub-species level specificity may be more common than is often assumed, including for species in the Poaceae. But special scrutiny is certainly required, including evaluation and proper weighting of evidence collected in different venues and with different methodologies. Within the US regulatory framework, there has been a tendency to regulate based upon no-choice test results, ignoring important factors determining fundamental and ecological host ranges or demographic impacts, which in science guided decision making should override data on trivial and inconsequential cosmetic feeding (Blossey 2016a, b; Cristofaro et al. 2013; Hinz et al. 2014; Smith 2012). Our results, and those of others, have demonstrated the existence of sub-species level selectivity in herbivorous insects, extending opportunities for weed biological control. The past history of bias or hesitation to assess weed biocontrol potential based on problematic assumptions about close relatives or grasses was not based on sound understanding of ecological or evolutionary relationships.
Recent revisions to guidance documents (USDA 2016) indicate that minor probing, as Archanara larvae do on some non-target species, is considered “non-feeding”. The document further states that “arthropods may ingest enough material on certain plants to produce droppings. However, if the life span of the organism will be extended but there is little evidence of continued development, then these accepted but unsuitable species can also be discarded as potential host plants” (USDA 2016). Laboratory testing and abundant evidence from the field assures that neither of our selected species feeds and develops on any crop plant (or any species outside the genus Phragmites), further satisfying USDA/APHIS requirements that introduced species are not potential plant pests of species of economic importance, as required under the Plant Protection Act.
Furthermore, the National Environmental Protection Act requires that releases do not present a risk to the environment and the US Fish and Wildlife Service (USFWS) reviews petitions to ensure that potential biocontrol agents do not pose any risk to threatened or endangered species. While we have documented the potential for A. geminipuncta and A. neurica to occasionally oviposit (< 7% of all eggs) on endemic P. australis americanus, we believe that the potential demographic consequences are minimal, especially compared to threats the species faces from invasive P. australis encroachment (Saltonstall 2002). Furthermore, native endemic P. australis americanus is not a listed species, and while we need to safeguard any species, this will in essence reduce USFWS review to species that may be indirectly affected via food web connections or changes in wetland structure (Lampert et al. 2014). Both Archanara species are important food sources for a number of European birds as well as insect parasitoids and predators (Tscharntke 1992a, b), and P. australis is not known to possess any potentially toxic secondary chemistry. Therefore, there should be no concerns about potential harm to predators of Archanara larvae or adults, whether they be birds, bats, or other vertebrates or invertebrates.
Considering potential threats to other wetland biota, we are left with concerns regarding the potential of rapid death of extensive P. australis monocultures. This was a concern that halted herbicide management of Spartina in California to spare habitat of the endangered California rail (Lampert et al. 2014). Similar concerns over rapid defoliation of Tamarix spp. (Tamaricaceae) by the biocontrol agent Diorhabda carinulata Desbrochers (Chrysomelidae) affecting breeding habitat of the endangered southwestern willow flycatcher Empidonax traillii extimus (Tyrannidae) led to a withdrawal of release permits (Dudley and Bean 2012). Based on European evidence, we recognize the potential for outbreaking Archanara populations. However, unlike herbicide or mechanical treatments (both often followed by spring burns), with biocontrol, at least in this case, dead standing biomass will remain in place. This provides shelter and cover for wetland biota such as marsh birds or invertebrates foraging in flooded or moist soils. While there may be increased light penetration following years of noctuid outbreaks, this is desired to allow native wetland plant recruitment.
Conclusions
Invasive P. australis is one of the most widespread and successful invasive species in North America (Chambers et al. 1999; Kettenring et al. 2012; Saltonstall and Meyerson 2016) with a long history of failed management using physical and largely chemical means at enormous expense and with a dearth of information about impacts of management of wetland biota (Hazelton et al. 2014; Marks et al. 1994; Martin and Blossey 2013b; Quirion et al. 2018). Allowing continued expansion of the invasive lineage would not only jeopardize native wetland biota but also threaten the existence of endemic P. australis americanus (Saltonstall 2002). While we cannot be certain about the contribution of A. geminipuncta and A. neurica to potential reductions in invasive P. australis performance or demography, we consider risks of releasing these two species to P. australis americanus very small compared to effects of further unabated invasive P. australis encroachment or even continued unselective management using herbicide.
Although grasses such as P. australis and target weeds with many congeneric native species require rigorous host range testing, evolution has provided biocontrol scientists with an enormous diversity of herbivores—no different from other targets in current weed biological control programs. The host plant resistance literature, not often discussed among biological control researchers, provides many examples of extreme (genotype, chemotype or morphotype-based) host specificity among insect herbivores of many crops, including many grasses. Among the insect herbivores of P. australis known in North America, host specificity at the sub-species level is common in the field. Based upon extreme ovipositional preference, plant characteristics, and their European distribution, we expect that the two Archanara species under consideration for release in North America will demonstrate field-level host specificity, limiting their reproduction and restricting their impact to introduced P. australis. The native lineages are safeguarded by a number of different morphological features, particularly lack of adherence of leaf-sheaths. Successful classical biological control of the invasive sub-species of P. australis in North America would provide incentive for addressing other challenging targets in classical biological control of weeds. We argue that each weed biocontrol agent should be evaluated on its own merits and reviews should be based on clearly articulated and reliable host specificity criteria, not whether the host plant belongs to a taxonomic group (like the Poaceae) or on the existence of many close relatives. As Pemberton (2000) states, erroneous beliefs and prejudice have prevented targeting grasses with biological control. Many of these species may offer appropriate targets without jeopardizing safety of native species or ecosystems.
References
Able KW, Hagan SM (2000) Effects of common reed (Phragmites australis) invasion on marsh surface macrofauna: response of fishes and decapod crustaceans. Estuaries 23:633–646
Agrawal AA (2011) Current trends in the evolutionary ecology of plant defence. Funct Ecol 25:420–432
Anderson P, Sadek MM, Larsson M, Hansson BS, Thoming G (2013) Larval host plant experience modulates both mate finding and oviposition choice in a moth. Anim Behav 85:1169–1175
Arnett AE, Louda SM (2002) Re-test of Rhinocyllus conicus host specificity, and the prediction of ecological risk in biological control. Biol Conserv 106:251–257
Austel N, Bjorkman C, Hilker M, Meiners T (2014) Phenotypic plasticity in host plant preference of the willow leaf beetle Phratora vulgatissima: the impact of experience made by adults. Agric For Entomol 16:417–425
Barrett LG, Heil M (2012) Unifying concepts and mechanisms in the specificity of plant-enemy interactions. Trends Plant Sci 17:282–292
Bean DW, Dalin P, Dudley TL (2012) Evolution of critical day length for diapause induction enables range expansion of Diorhabda carinulata, a biological control agent against tamarisk (Tamarix spp.). Evol Appl 5:511–523
Benoit LK, Askins RA (1999) Impact of the spread of Phragmites on the distribution of birds in Connecticut tidal marshes. Wetlands 19:194–208
Bertness MD, Ewanchuk PJ, Silliman BR (2002) Anthropogenic modification of New England salt marsh landscapes. Proc Nat Acad Sci USA 99:1395–1398
Blossey B (2003a) A framework for evaluating potential ecological effects of implementing biological control of Phragmites australis. Estuaries 26:607–617
Blossey B (2003b) Morphological differences between native North American Phragmites australis genotypes and introduced invasive European genotypes In: Weinstein MP, Keough JR, Guntenspergen GR, Litvin SY (eds) Phragmites australis: a sheep in wolf’s clothing?, 6–9 January 2002, Vineland, NJ, USA, New Jersey Marine Sciences Consortium, pp 47–56
Blossey B (2014) Identification, development, and release of insect biocontrol agents for the management of Phragmites australis. US Army Corps of Engineers, ERDC/EL CR-14-2, Washington DC
Blossey B (2016a) The future of biological control: a proposal for fundamental reform. In: van Driesche R, Simberloff D, Blossey B, Causton C, Hoddle M, Marks C, Heinz K, Wagner D, Wagner K (eds) Integrating biological control into conservation practice. Wiley, Chichester, pp 314–328
Blossey B (2016b) Measuring and evaluating ecological outcomes of biological control introductions. In: van Driesche R, Simberloff D, Blossey B, Causton C, Hoddle M, Marks C, Heinz K, Wagner D, Wagner K (eds) Integrating biological control into conservation practice. Wiley, Chichester, pp 161–188
Blossey B, Casagrande R (2016) Biological control of invasive Phragmites may safeguard native Phragmites and increase wetland conservation values. Biol Inv 18:2753–2755
Blossey B, Casagrande R, Tewksbury L, Landis DA, Wiedenmann RN, Ellis DR (2001) Nontarget feeding of leaf-beetles introduced to control purple loosestrife (Lythrum salicaria L.). Nat Areas J 21:368–377
Blossey B, Casagrande RA, Tewksbury L, Hinz H, Häfliger P, Martin L, Cohen J (2013) Identifying, developing and releasing insect biocontrol agents for the management of Phragmites australis. Progress report, ERDC/EL TN-13-3 July 2013, US Army Corps of Engineers, Vicksburg, MS, USA
Blossey B, Häfliger P, Tewksbury L, Dávalos A, Casagrande R (2018) Host specificity of Archanara geminipuncta and Archanara neurica, two potential biocontrol agents for invasive Phragmites australis in North America. Biol Control (in review)
Bolton RM, Brooks RJ (2010) Impact of the seasonal invasion of Phragmites australis (common reed) on turtle reproductive success. Chelonian Conserv Biol 9:238–243
Branson D, Joern A, Sword G (2015) Sustainable management of insect herbivores in grassland ecosystem: a new perspective in grasshopper control. BioScience 56:743–755
Brisson J, de Blois S, Lavoie C (2010) Roadside as invasion pathway for common reed (Phragmites australis). Invasive Plant Sci Manag 3:506–514
Brix H, Ye S, Laws EA, Sun D, Li G, Dinga X, Yuana H, Zhaoa G, Wanga J, Peia S (2014) Large-scale management of common reed, Phragmites australis, for paper production: a case study from the Liaohe Delta, China. Ecol Eng 73:760–769
Brooks ML, D’Antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. BioScience 54:677–688
Burdick D, Konisky RA (2003) Determinants of expansion for Phragmites australis, common reed, in natural and impacted coastal marshes. Estuaries 26:407–416
Carrasco D, Larsson MC, Anderson P (2015) Insect host plant selection in complex environments. Curr Opin Insect Sci 8:1–7
Chambers RM, Mozdzer TJ, Ambrose JC (1998) Effects of salinity and sulfide on the distribution of Phragmites australis and Spartina alterniflora in a tidal marsh. Aquat Bot 62:161–169
Chambers RM, Meyerson LA, Saltonstall K (1999) Expansion of Phragmites australis into tidal wetlands of North America. Aquat Bot 64:261–273
Charudattan R (2005) Ecological, practical, and political inputs into selection of weed targets: what makes a good biological control target? Biol Control 35:183–196
Clawson B, Duthinh EC (2015) Phragmites, invasions harm riparian property values: part 1. http://msue.anr.msu.edu/news/phragmites_invasions_harm_riparian_property_values_part_1. Accessed 5 March 2017
Cohen JS, Maerz JC, Blossey B (2012) Traits, not origin, explain impacts of plants on larval amphibians. Ecol Appl 22:218–228
Cohen JS, Rainford SKD, Blossey B (2014) Community-weighted mean functional effect traits determine larval amphibian responses to litter mixtures. Oecologia 174:1359–1366
Colin R, Eguiarte LE (2016) Phylogeographic analyses and genetic structure illustrate the complex evolutionary history of Phragmites australis in Mexico. Am J Bot 103:876–887
Crawley MJ (1989) The successes and failures of weed biocontrol using insects. Biocontrol News Inform 10:213–223
Crawley MJ, Gillman M (1989) Population dynamics of cinnabar moth and ragwort in grassland. J Anim Ecol 58:1035–1050
Cristofaro M, De Biase A, Smith L (2013) Field release of a prospective biological control agent of weeds, Ceratapion basicorne, to evaluate potential risk to a nontarget crop. Biol Control 64:305–314
Crocker E, Nelson EB, Blossey B (2017) Soil conditioning effects of Phragmites australis on native wetland plant seedling survival. Ecol Evol 7:5571–5579
Diaz R, Manrique V, Hibbard K, Fox A, Roda A, Gandolfo D, Mckay F, Medal J, Hight S, Overholt WA (2014) Successful biological control of tropical soda apple (Solanales: solanaceae) in Florida: a review of key program components. Fla Entomol 97:179–190
Djamin A, Pathak MD (1967) Role of silica in resistance to Asiatic rice borer, Chilo suppressalis (Walker) in rice varieties. J Econ Entomol 60:347–351
Dudley TL, Bean DW (2012) Tamarisk biocontrol, endangered species risk and resolution of conflict through riparian restoration. BioControl 57:331–347
Forister ML, Dyer LA, Singer MS, Stireman JO, Lill JT (2012) Revisiting the evolution of ecological specialization, with emphasis on insect-plant interactions. Ecology 93:981–991
Fritz RS, Simms E (1992) Plant resistance to herbivores and pathogens. University of Chicago Press, Chicago
Futuyma DJ, Agrawal AA (2009) Macroevolution and the biological diversity of plants and herbivores. Proc Nat Acad Sci USA 106:18054–18061
Gali-Muhtasib HU, Smith CC, Higgins JJ (1992) The effect of silica in grasses on the feeding behavior of the prairie vole, Microtus ochrogaster. Ecology 73:1724–1729
Gaskin JF, Schwarzlander M, Kinter CL, Smith JF, Novak SJ (2013) Propagule pressure, genetic structure, and geographic origins of Chondrilla juncea (Asteraceae): an apomictic invader on three continents. Am J Bot 100:1871–1882
Goolsby JA, Moran PJ, Racelis AE, Summy KR, Jimenez MM, Lacewell RD, de Leon AP, Kirk AA (2016) Impact of the biological control agent Tetramesa romana (Hymenoptera: Eurytomidae) on Arundo donax (Poaceae: Arundinoideae) along the Rio Grande River in Texas. Biocontrol Sci Technol 26:47–60
Grevstad FS, Strong DR, Garcia-Rossi D, Switzer RW, Wecker MS (2003) Biological control of Spartina alterniflora in Willapa Bay, Washington using the planthopper Prokelisia marginata: agent specificity and early results. Biol Control 27:32–42
Häfliger P, Schwarzlaender M, Blossey B (2005) Biology of Platycephala planifrons (Diptera: Chloropidae) and its potential effectiveness as biological control agent for invasive Phragmites australis in North America. Biol Control 34:302–311
Häfliger P, Schwarzlaender M, Blossey B (2006a) Impact of Archanara geminipuncta (Lepidoptera: Noctuidae) on above-ground biomass production of Phragmites australis. Biol Control 38:413–421
Häfliger P, Schwarzländer M, Blossey B (2006b) A comparison of biology and host plant utilization of Archanara geminpuncta, A. dissoluta, A. neurica and Arenostola phragmitidis (Lepidoptera: Noctuidae), potential biological control agents of Phragmites australis (Arundineae: Poaceae). Ann Entomol Soc Am 99:683–696
Hakizimana S, Olckers T (2013) Should the flower bud weevil Anthonomus santacruzi (Coleoptera: Curculionidae) be considered for release against the invasive tree Solanum mauritianum (Solanaceae) in New Zealand? Biocontrol Sci Technol 23:197–210
Hartley SE, DeGabriel JL (2016) The ecology of herbivore-induced silicon defences in grasses. Funct Ecol 30:1311–1322
Hazelton ELG, Mozdzer TJ, Burdick DM, Kettenring KM, Whigham DF (2014) Phragmites australis management in the United States: 40 years of methods and outcomes. AoB PLANTS. https://doi.org/10.1093/aobpla/plu001
Heinrichs EA (1986) Perspectives and directions for the continued development of insect-resistant rice varieties. Agric Ecosyst Environ 18:9–36
Hinz HL, Schwarzländer M, Gassmann A, Bourchier RS (2014) Successes we may not have had: a retrospective analysis of selected weed biological control agents in the United States. Invasive Plant Sci Manag 7:565–579
Hinz HL, Winston RL, Schwarzländer M (2018) How safe is weed biological control? A global review of direct non-target attack. Biol Inv (submitted)
Hudon C, Gagnon P, Jean M (2005) Hydrological factors controlling the spread of common reed (Phragmites australis) in the St. Lawrence river (Québec, Canada). Ecoscience 12:347–357
Jahner JP, Bonilla MM, Badik KJ, Shapiro AM, Forister ML (2011) Use of exotic hosts by Lepidoptera: widespread species colonize more novel hosts. Evolution 65:2719–2724
Jodoin Y, Lavoie C, Villeneuve P, Theriault M, Beaulieu J, Belzile F (2008) Highways as corridors and habitats for the invasive common reed Phragmites australis in Quebec, Canada. J Appl Ecol 45:459–466
Karley AJ, Mitchell C, Brookes C, McNicol J, O’Neill T, Roberts H, Graham J, Johnson SN (2016) Exploiting physical defence traits for crop protection: leaf trichomes of Rubus idaeus have deterrent effects on spider mites but not aphids. Ann Appl Biol 168:159–172
Kennedy E, Leff LG, de Szalay FA (2012) Herbiciding Phragmites australis: effects on litter decomposition, microbial biomass, and macroinvertebrate communities. Fundam Appl Limnol 180:309–319
Kettenring KM, Blois SD, Hauber DP (2012) Moving from a regional to a continental perspective of Phragmites australis invasion in North America. AoB PLANTS. https://doi.org/10.1093/aobpla/pls040
Kulmatiski A, Beard KH, Meyerson LA, Gibson JR, Mock KE (2010) Nonnative Phragmites autralis invasion into Utah wetlands. West N Am Nat 70:541–552
Lambertini C, Mendelssohn IA, Gustafsson MHG, Olesen B, Riis T, Sorrell BK, Brix H (2012) Tracing the origin of Gulf Coast Phragmites (Poaceae): a story of long-distance dispersal and hybridization. Am J Bot 99:538–551
Lampert A, Hastings A, Grosholz ED, Jardine SL, Sanchirico JN (2014) Optimal approaches for balancing invasive species eradication and endangered species management. Science 344:1028–1031
Larochelle M, Dumont P, Lavoie C, Hatin D (2015) Varying effects of common reed invasion on early life history of Northern pike. Trans Am Fish Soc 144:196–210
Liu SS, Li YH, Liu YQ, Zalucki MP (2005) Experience-induced preference for oviposition repellents derived from a non-host plant by a specialist herbivore. Ecol Lett 8:722–729
Lombard KB, Tomassi D, Ebersole J (2012) Long-term management of an invasive plant: lessons from seven years of Phragmites australis control. North East Nat 19:181–193
Lou YG, Zhang GR, Zhang WQ, Hu Y, Zhang J (2013) Biological control of rice insect pests in China. Biol Control 67:8–20
Marks M, Lapin B, Randall JA (1994) Phragmites australis (P. communis): threats, management and monitoring. Nat Areas J 14:285–294
Marohasy J (1996) Host shifts in biological weed control: real problems, semantic difficulties or poor science? Int J Pest Manage 42:71–75
Martin LJ, Blossey B (2013a) Intraspecific variation overrides origin effects in impacts of litter-derived secondary compounds on larval amphibians. Oecologia 173:449–459
Martin LJ, Blossey B (2013b) The runaway weed: costs and failures of Phragmites australis management in the USA. Estuaries Coasts 36:626–632
Massey FP, Ennos AR, Hartley SE (2007) Herbivore specific induction of silica-based plant defences. Oecologia 152:677–683
McGuire R, Johnson MT (2006) Plant genotype and induced responses affect resistance to herbivores on evening primrose (Oenothera biennis). Ecol Entomol 31:20–31
Meyerson LA, Viola DV, Brown RN (2008) Hybridization of invasive Phragmites australis with a native subspecies in North America. Biol Inv 12:103–111
Michel R, Tscharntke T (1993) Ursachen der populationsdichteschwankungen von schmetterlingen im ökosystem schilf (Phragmites australis Trin.). Mitt Dtsch Ges Allg Angew Entomol 8:511–515
Moore B, Andrew RL, Kulheim C, Foley WJ (2014) Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol 201:733–750
Myers JH, Sarfraz RM (2017) Impacts of insect herbivores on plant populations. Annu Rev Entomol 62:207–230
National Research Council (2004) Endangered and threatened species of the Platte River. National Academies, Washington
Nestory S (2016) Biological and mechanical control of Japanese stiltgrass (Microstegium vimineum). M.S. Thesis. University of Delaware, Newark, DE, USA
Olckers T (2000) Biology, host specificity and risk assessment of Gargaphia decoris, the first agent to be released in South Africa for the biological control of the invasive tree Solanum mauritianum. BioControl 45:373–388
Olckers T (2011) Biological control of Solanum mauritianum Scop. (Solanaceae) in South Africa: will perseverance pay off? Afr Entomol 19:416–426
Olckers T, Hulley PE (1994) Resolving ambiguous results of host-specificity tests—the case of two Leptinotarsa species (Coleoptera, Chrysomelidae) for biological control of Solanum elaeagnifolium Cavanilles (Solanaceae) in South Africa. Afr Entomol 2:137–144
Overholt WA, Hidayat P, Le Ru B, Takasu K, Goolsby JA, Racelis A, Burrell AM, Amalin D, Agum W, Njaku M, Pallangyo B, Klein PE, Cuda JP (2016) Potential biological control agents for management of cogongrass (Cyperales: Poaceae) in the southeastern USA. Fla Entomol 99:734–739
Painter RH (1951) Insect resistance in crop plants. University Press of Kansas, Manhattan
Parsons KC (2003) Reproductive success of wading birds using Phragmites marsh and upland nesting habitats. Estuaries 26:596–601
Pearse IS, Hipp AL (2009) Phylogenetic and trait similarity to a native species predict herbivory on non-native oaks. Proc Nat Acad Sci USA 106:18097–18102
Pearse IS, Harris DJ, Karban R, Sih A (2013) Predicting novel herbivore-plant interactions. Oikos 122:1554–1564
Pemberton RW (1996) The potential of biological control for the suppression of invasive weeds of southern environments. Castanea 61:313–391
Pemberton RW (2000) Predictable risk to native plants in weed biocontrol. Oecologia 125:489–494
Profitt M, Khalla MA, Carrasco D, Larsson MC, Anderson P (2015) ‘Do you remember the first time?’ Host plant preference in a moth is modulated by experiences during larval feeding and adult mating. Ecol Lett 18:365–374
Quirion B, Simek Z, Dávalos A, Blossey B (2018) Management of invasive Phragmites australis in the Adirondacks: a cautionary tale about prospects of eradication. Biol Inv 20:59–73
Racelis AE, Goolsby JA, Moran P (2009) Seasonality and movement of adventive populations of the Arundo wasp (Hymenoptera: Eurytomidae), a biological control agent of giant reed in the Lower Rio Grande Basin in South Texas. Southwest Entomol 34:347–357
Rogalski MA, Skelly DK (2012) Positive effects of nonnative invasive Phragmites australis on larval bullfrogs. PLoS ONE 7(8):e44420. https://doi.org/10.1371/journal.pone.0044420
Rossiter NA, Setterfield SA, Douglas MM, Hutley LB (2003) Testing the grass-fire cycle: alien grass invasion in the tropical savannas of northern Australia. Divers Distrib 9:169–176
Saltonstall K (2002) Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc Nat Acad Sci USA 99:2445–2449
Saltonstall K (2016) The naming of Phragmites haplotypes. Biol Inv 18:2433–2441
Saltonstall K, Meyerson LA (2016) Phragmites australis: from genes to ecosystems. Biol Inv 18:2415–2420
Saltonstall K, Peterson PM, Soreng RJ (2004) Recognition of Phragmites australis subsp. americanus (Poaceae: Arundinoideae) in North America: evidence from morphological and genetic analyses. SIDA 21:683–692
Saltonstall K, Castillo HE, Blossey B (2014) Confirmed field hybridization of native and introduced Phragmites australis (Poaceae) in North America. Am J Bot 101:211–215
Saltonstall K, Lambert AM, Rice N (2016) What happens in Vegas, better stay in Vegas: Phragmites australis hybrids in the Las Vegas Wash. Biol Inv 18:2463–2474
Sciance MB, Patrick CJ, Weller DE, Williams MN, McCormick MK, Hazelton ELG (2016) Local and regional disturbances associated with the invasion of Chesapeake Bay marshes by the common reed Phragmites australis. Biol Inv 18:2661–2677
Secord D, Kareiva P (1996) Perils and pitfalls in the host specificity paradigm. BioScience 46:448–453
Settle WH, Ariawan H, Astuti ET, Cahyana W, Hakim AL, Hindayana D, Lestari AS, Pajarningsih Sartanto (1996) Managing tropical rice pests through conservation of generalist natural enemies and alternative prey. Ecology 77:1975–1988
Shabani F, Kumar L, Ahmadi M (2016) A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol Evol 6:5973–5986
Silliman BR, Bertness MD (2004) Shoreline development drives invasion of Phragmites australis and the loss of plant diversity on New England salt marshes. Conserv Biol 18:1424–1434
Smith L (2012) Host plant oviposition preference of Ceratapion basicorne (Coleoptera: Apionidae), a potential biological control agent of yellow starthistle. Biocontrol Sci Technol 22:407–418
Strong DR, McCoy ED, Rey JR (1977) Time and number of herbivore species—pests of sugarcane. Ecology 58:167–175
Suckling DM, Sforza RFH (2014) What magnitude are observed non-target impacts from weed biocontrol? PLoS ONE 9(1):e84847. https://doi.org/10.1371/journal.pone.0084847
Tewksbury L, Casagrande R, Blossey B, Häfliger P, Schwarzländer M (2002) Potential for biological control of Phragmites australis in North America. Biol Control 23:191–212
Tscharntke T (1989) Attack by a stem-boring moth increases susceptibility of Phragmites australis to gallmaking by a midge: mechanisms and effects on midge population dynamics. Oikos 55:93–100
Tscharntke T (1990) Fluctuations in abundance of a stem-boring moth damaging shoots of Phragmites australis: causes and effects of overexploitation of food in a late-successional grass monoculture. J Appl Ecol 27:679–692
Tscharntke T (1992a) Cascade effects among four trophic levels: bird predation on galls affects density-dependent parasitism. Ecology 73:1689–1698
Tscharntke T (1992b) Fragmentation of Phragmites habitats, minimum viable population size, habitat suitability, and local extinction of moths, midges, flies, aphids, and birds. Conserv Biol 6:530–536
Tscharntke T (1999) Insects on common reed (Phragmites australis): community structure and the impact of herbivory on shoot growth. Aquat Bot 64:339–410
USDA (2016) Technical advisory group for biological control agents of weeds manual, Interim edn. United States Department of Agriculture, Washington
USDA NRCS (2017) The plants database (http://plants.usda.gov/). National Plant Data Team, Greensboro, NC, USA
van Klinken RD, Edwards OR (2002) Is host specificity of weed biocontrol agents likely to evolve rapidly following establishment? Ecol Lett 5:590–595
Vasquez EA, Glenn EP, Brown JJ, Guntenspergen GR, Nelson SG (2005) Salt tolerance underlies the cryptic invasion of North American salt marshes by an introduced haplotype of the common reed Phragmites australis (Poaceae). Mar Ecol Prog Ser 298:1–8
Vogel JA, Koford RR, Debinski DM (2010) Direct and indirect responses of tallgrass prairie butterflies to prescribed burning. J Insect Conserv 14:663–677
Ward DB (2010) North America has two species of Phragmites (Gramineae). Castanea 75:394–401
Whitham TG, Gehring AA, Lamit LJ, Wojtowicz T, Evans LM, Keith AR, Smith DS (2012) Community specificity: life and afterlife effects of genes. Trends Plant Sci 17:271–281
Whyte RS, Bocetti CI, Klarer DM (2015) Bird assemblages in Phragmites dominated and non-Phragmites habitats in two Lake Erie coastal marshes. Nat Areas J 35:235–245
Winston RL, Schwarzländer M, Hinz HL, Day MD, Cock MJW, Julien MH (2014) Biological control of weeds: a world catalogue of agents and their target weeds, 5th edn. USDA Forest Service, Forest Health Technology Enterprise Team, Morgantown, WV, USA. FHTET-2014-04
Witt ABR, McConnachie AJ (2003) The potential for classical biological control of invasive grass species with special reference to invasive Sporobolus spp. (Poaceae) in Australia In: Cullen JM, Briese DT, Kriticos DJ, Lonsdale WM, Morin L, Scott JK (eds) Proceedings of the XI international symposium on biological control of weeds, Canberra, Australia, 27 April–2 May, CSIRO Entomology, GPO Box 1700, Canberra ACT 2601, pp 198–202
Zheng H, Wu Y, Ding J, Binion D, Fu W, Reardon R (2004) Invasive plants of Asian origin established in the United States and their natural enemies, vol 1. USDA Forest Service, Forest Health Technology Enterprise Team, Morgantown, WV, FHTET-2004-05
Zhu Y, Chen H, Fan J, Wang Y, Li Y, Chen J, Fan J, Yang S, Hu L, Leung H, Mew T, Teng PS, Wang Z, Mundt CC (2000) Genetic diversity and disease control in rice. Nature 406:718–722
Acknowledgements
We appreciate support and assistance we have received over the past 20 years from wetland and wildlife managers in the USA and Canada and technical support from many undergraduate students. Major funding for this work was provided by the US Fish and Wildlife Service, Army Corps of Engineers, New York State Department of Transportation, Rhode Island and New York Sea Grant programs, and Agricultural Experiment Stations of Cornell and University of Rhode Island. Patrick Häfliger and Hariet Hinz were supported by CABI with core financial support from its member countries (see http://www.cabi.org/about-cabi/who-we-work-with/key-donors/). We thank Rachel Winston for background info on biocontrol and Wade Simmons for help in preparing the figures. We appreciate helpful comments by reviewers and editors that improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editors: Mark Schwarzländer, Cliff Moran and S. Raghu
Rights and permissions
About this article

{kind=link}
Cite this article
Casagrande, R.A., Häfliger, P., Hinz, H.L. et al. Grasses as appropriate targets in weed biocontrol: is the common reed, Phragmites australis, an anomaly?. BioControl 63, 391–403 (2018). https://doi.org/10.1007/s10526-018-9871-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-018-9871-y