
Abstract
Classical biological control using insects has led to the partial or complete control of at least 226 invasive insect and 57 invasive weed species worldwide since 1888. However, at least ten introductions of biological control agents have led to unintended negative consequences and these cases have led to a focus on risk that came to dominate the science and practice of classical biological control by the 1990s. Based upon historical developments in the field we consider that the era of focus on benefits began in 1888 and that it was supplanted by an era in which the focus was on risks during the 1990s. This paradigm shift greatly improved the safety of biological control releases but also led to a decline in the number of introductions, probably resulting in opportunity costs. We note here the development of a third paradigm: one in which the benefits and risks of biological control are clearly and explicitly balanced so that decisions can be made that maximize benefits while minimizing risks.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The first widely acknowledged successful case of classical (also known as ‘importation’) biological control involved the iconic case of importation of the predatory beetle Rodolia cardinalis (Mulsant) and the parasitic fly Chryptochaetum iceryae (Williston) to control the cottony cushion scale, Icerya purchasi Maskall in California in 1888 (Caltagirone and Doutt 1989). Since then, classical biological control has grown with hundreds of invasive arthropods and weeds targeted, and this has resulted in the long-term suppression of numerous species of important pests and weeds (Winston et al. 2014; Cock et al. 2016). However, a small proportion of introductions have been ecologically damaging, resulting in significant non-target effects (Louda et al. 2003). Thus classical biological control has produced important environmental benefits but has also introduced ecological risks. In the last two decades biological control scientists have studied these risks intensively and have developed risk assessment protocols in an effort to make introductions ecologically safer (e.g. Follett and Duan 2000; Wajnberg et al. 2001; van Driesche and Reardon 2004; van Lenteren et al. 2003, 2006; Bigler et al. 2006; Heimpel and Mills 2017). However, a framework for balancing risks and benefits has not yet fully emerged (Sheppard et al. 2003; van Wilgen et al. 2013; Hinz et al. 2014). Such a framework includes understanding the relationship between risks and benefits in biological control and creating conceptual models that allow the comparison of anticipated benefits and risks and a means to maximize the former while minimizing the latter.
In this paper we provide a brief historical review of cases of classical biological control that either led to successful suppression of target populations or to ecological disruption or harm. We also discuss attitudes about biological control from the beginning of the practice until the present time and argue that a paradigm shift has occurred in which a focus on benefits shifted to a focus on risks in the 1990s. We end with an appeal for expanded uptake of a developing third paradigm—one in which risks and benefits are properly balanced—and discuss some ways to do this.
The historical record of classical biological control: Successes and adverse effects
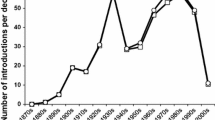
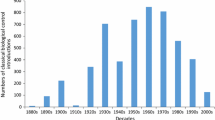
We collated cases of successful classical biological control of arthropod and weed targets from the inception of the practice to the current time. For insect pests and natural enemies we used the BIOCAT2010 database (Cock et al. 2016) and for weeds we used Winston et al. (2014). Both of these databases were supplemented with some cases covered by van Driesche et al. (2010). We did not include biological control of invertebrates other than insects, or the use of biological control of insects using agents other than insects for this analysis, but some trends involving these and other categories are discussed by Hajek et al. (2005, 2007) and Heimpel and Mills (2017). Classification of cases is slightly different for the two datasets and we tabulated successes as follows. For insects, impacts on targets that were classified as ‘partial’ successes or better in at least one country were tabulated and for weeds, impact on targets by at least one agent that were classified as ‘heavy’ either ‘regionally’ or ‘widespread’ were tabulated. This process revealed 226 successes against insect targets and 57 successes against weed targets. The top half of Fig. 1 shows the year of first release of all of these cases to illustrate the chronological pattern of biological control successes. The first success occurred in 1888 (biological control of cottony cushion scale as noted above) and the most recent one in 2011 (biological control of boxing glove cactus, Cylindropuntia fulgida Engelm., in South Africa by the cochineal insect Dactylopius tomentosus (Lamarck)).

A timeline of biological control introductions of insect agents against invasive insect pests and invasive weeds based upon the year of the first introduction of agents that provided partial or complete control of their intended targets (positive values) divided into targets that were considered primarily detrimental to economic interests or natural ecosystems. Negative values denote introduction of agents that led to unindended negative consequences as follows: 1Indian mongoose, Herpestes javanicus (É. Geoffroy Saint-Hilaire) (Seaman and Randall 1962); 2The parasitoid Cotesia glomerata (Linnaeus) (Benson et al. 2003); 3The parasitoid Compsilura concinnata (Meigen) (Boettner et al. 2000); 4Mosquitofish, Gambusia spp. (Minckley and Deacon 1968); 5The cane toad, Bufo marinus (Linnaeus) (Urban et al. 2007); 6The predatory snail Euglandina rosea (Férussac) (Civeyrel and Simberloff 1996); 7The weevil Rhinocyllus conicus (Frölich) (Rose et al. 2005); 8The seed-fly Urophora affinis (Frauenfeld) (Pearson and Callaway 2006); 9The seed-fly U. quadrafasciata (Meigen) (Pearson and Callaway 2006); 10The seed-fly Mesoclanis polana (Munro) (Carvalheiro et al. 2008)
The bottom half of Fig. 1 shows the cases of classical biological control that have had reportedly harmful ecological impacts, divided into direct effects on non-target organisms (seven agents) and indirect effects where populations of specialized agents became abundant and led to enrichment of native species leading to ecological alteration or disruption (three agents involving two weed species). Only cases of well-documented population-level non-target effects on native species are included in the figure. These are mainly cases for which manipulative field experiments were done that demonstrated such effects. We do not include some relatively well-known cases from this list for various reasons. For example, negative effects of the cactus moth Cactoblastis cactorum (Berg) on native Opuntia cacti in the Southeastern USA (Stiling et al. 2004) are not included because this species was not deliberately introduced in the United States and thus is more appropriately viewed as an invasive species (Andraca-Gomez et al. 2015). Also, the possible displacement of native coccinellids by introduced species such as Harmonia axyridis (Pallas) and Coccinella septempunctata Linnaeus within agricultural fields in the United States is not included because it is not clear whether these non-target effects extend into natural areas (Evans 2004; Harmon et al. 2007). While H. axyridis does seem to be causing negative effects in natural areas of Europe (Roy et al. 2012), it was introduced there as an augmentative biological control agent, and so this case is not included either. We also do not include negative effects of accidentally introduced biological control agents (e.g. Louda and O’Brien 2002).
Figure 1 shows two trends very clearly with respect to benefits and risks. Firstly, that successful cases of classical biological control greatly outnumber cases that produced ecological harm: the proportion of harmful cases is far less than 1%. And secondly, that introductions leading to direct negative effects appear to have ceased after the 1960s, but some cases of negative indirect effects have persisted into the 1990s. This latter pattern suggests that the biological control community has made progress in reducing risks of direct non-target impacts but that there is scope for improvement in minimizing risks of indirect non-target impacts (Wajnberg et al. 2001; Pearson and Callaway 2003; Kaser and Heimpel 2015; Kaser and Ode 2016).
Figure 1 only reflects the number of first examples of each success or harm and thus under-represents benefits. Successful classical biological control has produced enormous returns in terms of reduced crop losses, and in some cases the impacts of this on livelihoods and poverty have also been documented (Gurr and Wratten 2000; Cock et al. 2015; Naranjo et al. 2015). Environmental harm has been documented as discussed above and shown in Fig. 1, but it has not been measured in such a way that benefits and harm can be directly compared as we discuss further below.
We have divided the successful cases shown in Fig. 1 into four categories: for insect and weed targets we differentiated between introductions that were conducted for primarily economic reasons and for primarily conservation reasons. Economic targets include pests and weeds in agricultural settings as well as in natural habitats that damage commercial resources such as lumber and rangeland used for grazing livestock. Conservation targets include invasive species threatening native biodiversity. This distinction can be imperfect since biological control in agricultural settings can protect native biodiversity and biological control in natural settings can result in economic benefits (van Driesche et al. 2010). Furthermore, introduced insects or plants may be primarily agricultural pests in some countries and primarily damage ecosystems in others. For example the cottony cushion scale has primarily been targeted as a pest of fruit trees and ornamentals, but in the Galapagos Islands classical biological control was carried out to protect native plants (Hoddle et al. 2013). In this scenario, we counted the first success/harm in each system separately. In ambiguous cases, such as the release of biological control agents of aquatic weeds that can open waterways for transportation as well as protect biodiversity we classified releases as supporting conservation if benefits to particular native species were documented. Figure 1 shows clearly that classical biological control in support of conservation is a relatively recent practice. The first such introduction dates to 1962 with releases of two seed-feeding insects against the invasive shrubby weed, Hakea sericea Schrad & J.C.Wendl (Proteaceae), in South Africa, resulting in protection of plant species in the unique fynbos vegetation zone (Esler et al. 2010). However, despite this and some other early cases most introductions against targets of conservation concern occurred since the 1980s. These cases have been comprehensively reviewed by van Driesche et al. (2010).
A paradigm shift
Prior to the 1980s, the risks of biological control introductions to native biodiversity were seldom considered—particularly for biological control of insect targets. Theoretical and comparative studies focused almost entirely on benefits, posing questions such as: what is the rate of success of biological control introductions? What are the conditions under which biological control is most successful? Which traits of biological control agents determine success in biological control? (e.g. Beddington et al. 1978; Hall and Ehler 1979; Hall et al. 1980; Murdoch et al. 1985; Luck 1990; Stiling 1990, 1993; Greathead and Greathead 1992; Crawley 1989; Lane et al. 1999; Denoth et al. 2002; Kimberling 2004; Mills 2006; Stephens et al. 2013; Heimpel and Mills 2017). We call the period between 1888 and 1990 the ‘benefits era” since the emphasis of biological control scientists was on understanding how the benefits of biological control could be maximized.
Risks of biological control came to dominate the discussion of classical biological control beginning in the 1990s and from then until the present time risks of classical biological control have been extensively studied and reviewed. Unintended effects of biological control releases done before the 1950s in Hawaii caused concern (Howarth 1991; Hennemann and Memmott 2001; Messing and Wright 2006) as did effects of the released weevil Rhinocyllus conicus (Frölich) on native thistles in North America (Louda et al. 1997). These and some other cases led to a re-evaluation of classical biological control as a risk-free strategy to control invasive pests and weeds (Howarth 1991; Simberloff and Stiling 1996; Follett and Duan 2000; Wajnberg et al. 2001; Louda et al. 2003). This emphasis represented a major shift in the focus of biological control scientists from benefits to risks. We therefore consider the period from the 1990s to the present time the ‘risk era’. Although it is somewhat artificial, we denote 1991 as the first year of the risk era because of the influential review paper by Howarth (1991) titled ‘Environmental impacts of classical biological control’. Another important review paper from the 1990s was authored by Simberloff and Stiling (1996) titled ‘How risky is biological control?’. This was a particularly provocative paper in that it openly questioned the ecological safety of biological control introductions in general, stating that agents should be considered ‘guilty until proven innocent’ (pg. 1970), a sentiment that was echoed by McEvoy and Coombs (2000). Simberloff and Stiling (1996) also suggested that a ‘shift in modus operandi is needed […] in biological control’ (page 1970). We suggest that this has indeed happened and that a paradigm shift occurred around the year 1991 from an emphasis on benefits to an emphasis on risks. For instance, as a reflection of this, biological control decisions in the European Union are based on the precautionary principle, such that if an action or policy has a suspected risk of causing harm to the public or to the environment, the burden of proof that it is not harmful falls on those taking that action (Ehlers 2011).
This paradigm shift had two interrelated effects: a decline in biological control introductions and the development of novel risk-assessment protocols for biological control agents. The decline in introductions was very steep in Hawaii (Messing and Wright 2006) but occurred worldwide as well (Cock et al. 2016) although it should be noted that the decline in the last decade reported by Cock et al. (2016) is in part due to partial reporting at the time information was compiled, and also that such a decline was not seen in New Zealand (see below and Hill et al. 2013). The decline can be seen in Fig. 1 as well, where the number of successful first introductions dropped to 1.2 per year between 1998 through 2012 from 4.0 in the previous 20 years. However, despite a general decline in introductions, there was an increase in successful introductions for conservation purposes over this time period as noted above (Fig. 1). This can likely be attributed to the fact that the invasive targets in these cases were causing significant harm to populations of native species and management strategies other than biological control had either been tried and failed or were deemed to pose greater risks than biological control (or both). This illustrates the fact that biological control of species of conservation interest presents a relatively straightforward comparison of risks and benefits because the risks of doing and not doing biological control can be assessed in the same ‘currency’: declines in populations of native species (see below).
As noted above, the new emphasis on risk also led to the development of novel risk assessment protocols for biological control (e.g. van Lenteren et al. 2003, 2006; van Driesche and Reardon 2004; Wright et al. 2005; Bigler et al. 2006; Heimpel and Mills 2017). Risk assessment protocols for weed biological control were formalized in the 1970s (Wapshere 1974) and have been largely successful (Pemberton 2000; van Klinken and Edwards 2002; Paynter et al. 2004; Hinz et al. 2014; Suckling and Sforza 2014) but they had been notably absent from arthropod biological control programs (van Driesche and Hoddle 1997). Even for weed biological control programs, however, much of the emphasis of risk assessment was directed at protecting crop plants from weed biological control agents and the new risk paradigm increased the extent to which effects of such agents on native species was considered (Hinz et al. 2014). For both insect and weed biological control programs, new risk assessment protocols, along with more stringent regulations governing biological control releases have presumably contributed to fewer and safer biological control introductions, and have likely led to a steep decline in documented unintended negative side effects of biological control introductions (Fig. 1).
Lastly, we note the emergence of a new paradigm, in which the emphasis is on a balance between risks and benefits and a comparison with alternative management options, including doing nothing. We call this the ‘era of balanced benefits and risks’.
A framework that includes benefits and risks
In this section we discuss frameworks for analyzing classical biological control that include both benefits and risks. We first consider the relationship between risks and benefits and then ways that decision models can incorporate both of these aspects.
The relationship between risks and benefits in biological control
Murdoch et al. (1985) described two suites of idealized life-history traits that can lead to produce effective biological control agents. ‘Search-and-destroy’ agents are specialists that are well-adapted to locating, attacking and consuming their host (or prey) species and are able to drive their populations to low levels. Their own populations are intimately linked to that of their hosts and decline when hosts are suppressed. ‘Lying-in-wait’ agents, on the other hand, are generalists that can build up high population levels in the absence of pests and are able to suppress them when they arrive or begin to build their populations. Search-and-destroy agents illustrate how safety and efficacy can be positively correlated in specialized biological control agents (Kimberling 2004). Indeed, it is precisely the efficacy of these agents that reduces the risk that they pose since their abundance declines with that of their host. Lying-in-wait agents, on the other hand, illustrate how efficacy can trade off with safety since it is precisely the broad diet breadth (including omnivory) that increases the efficacy of these agents (Coll and Guershon 2002; Symondson et al. 2002; Lundgren 2009). Murdoch et al. (1985) did not broach the topic of safety in classical biological control or what class of biological control these idealized natural enemies would be appropriate for, but it seems clear now that search-and-destroy agents are appropriate for classical biological control while lying-in-wait agents are appropriate for conservation or natural biological control. Thus, for classical biological control, safety and efficacy will usually be positively associated. The relationship between specificity (which is linked to safety) and efficacy for idealized search-and-destroy and lying-in-wait biological control agents is provided in Fig. 2.

Hypothesized relationships between specificity and efficacy for biological control agents that follow Murdoch et al.’s (1985) “search-and-destroy” and “lying-in-wait” idealized life histories
Of course this view is oversimplified. In particular, specificity and efficacy can be uncoupled in ways that increase risk through indirect interactions. This was noted by Holt and Hochberg (2001) who used a theoretical approach to show that agents that establish at high levels without suppressing target populations can pose risks through enrichment effects. This enrichment can lead to indirect effects such as apparent competition which can be beneficial or harmful depending on whether other pest species or non-target species are affected (Kaser and Heimpel 2015; Kaser and Ode 2016). Ecological risks that flow from this kind of enrichment are exemplified by seedhead-galling tephritids introduced against spotted knapweed, Centaurea stoebe Lam. and bitou bush, Chrysanthemoides monilifera (L.) Norlindh, in the United States and Australia, respectively (Pearson and Callaway 2006; Carvalheiro et al. 2008). In both of these cases highly specialized agents established and reached high abundances themselves without substantially decreasing weed abundance. This allowed enrichment of consumers of the agents which in turn led to unintended ecological disruption through mechanisms such as apparent competition. A caveat to the spotted knapweed case is that weed suppression (efficacy) may have improved substantially after the seed bank was depleted—30 years following establishment (Story et al. 2008). The negative effects noted above will presumably diminish greatly with effective spotted knapweed suppression. Regardless, these case studies and the models of Holt and Hochberg (2001) and Kaser and Heimpel (2015) show how important it is to introduce biological control agents that are predicted to both exhibit specificity in field settings and that are expected to suppress pest or weed populations. The latter is desirable as a matter of course but it is important to recognize that efficacy can also confer safety to imported agents.
Decision models incorporating risks and benefits
Release petitions for classical biological control agents typically include a section that outlines the potential benefits to be gained by performing the release (Sheppard et al. 2003). However, only New Zealand and Australia incorporate a formal comparison between risk and benefits into their decision-making process as far as we are aware (Hunt et al. 2008; Hinz et al. 2014) although the Australian system was reportedly still primarily risk-based in the early 2000s (Sheppard et al. 2003; Harrison et al. 2005). Decisions made through New Zealand’s 1998 Hazardous Substances and New Organisms Order are based upon the core principle that biological control introductions are justified if the expected environmental benefits outweigh the expected adverse effects, and the regulatory processes provide guidance on how to make such a determination (Hill et al. 2013). This process, which includes a healthy dialogue between regulators and scientists, appears to have resulted in a balanced climate for conducting biological control which has led to approval of releases of biological control agents against 11 weed and four insect species since 1998 (Hill et al. 2013; see also Barratt and Moeed 2005; Suckling 2013). This is a higher rate of approval than seen in other countries and there is no indication of any environmental harm to date.
The implicit message of decision models that include both risks and benefits is that a goal of zero risk is unrealistic and that a more realistic goal is a net benefit in which the benefits exceed the risks of harm. This is the general approach taken in other applied sciences such as conservation biology (Wiggering et al. 2006; Maron and Cockfield 2008; McBride et al. 2010) and medicine (Blower et al. 2001; Lazo-Langer et al. 2012) where the benefits of interventions must often be weighed against possible or certain negative side effects. Also, biological control interventions should not only be compared against the options of doing nothing but against other management tactics. This approach was taken by Bigler and Kölliker-Ott (2006) who found that the environmental and health risks of using the insecticide deltamethrin to control the European corn borer, Ostrinia nubilalis (Hübner) (Lepidoptera: Crambidae), were much higher than using the parasitoid Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) in augmentative releases. Frameworks for simultaneously assessing benefits and risks of biological control introductions have been proposed by Bigler and Kölliker-Ott (2006) and Moeed et al. (2006) and were reviewed by De Clercq et al. (2011). These approaches involve qualitatively estimating the likelihood and the magnitude of both the risk and the benefits of the potential introduction. The resulting information can be used to compare risks and benefits within a decision framework. Since this process ideally includes information on the likelihood of success and the magnitude of the benefits, it should be possible to link these to the amount of risk that can be accepted. In essence, we should be more willing to accept a higher level of risk when confronted with pests or weeds with devastating rather than mild consequences and when the likelihood of successful biological control is higher rather than low. These considerations are illustrated in Fig. 3, where the magnitude of the benefit of biological control is on the x-axis and the likelihood of success on the y-axis. A line with a negative slope divides this parameter space into regions where less and more risk should be accepted below and above the line, respectively. Thus, more risk should be accepted for projects targeting potentially devastating pests that have a high probability of success than projects targeting minor pests that have a low probability of success. While the probability of success for classical biological control remains difficult to predict, a number of comparative analyses have provided useful guidance (Hawkins and Cornell 1994; Kimberling 2004; Heimpel and Mills 2017).

A conceptual model illustrating how the level of acceptable risk can be related to the magnitude of the potential benefit of biological control (proportional to the severity of damage caused by the invasive target of biological control) and the probability of success in biological control. The model suggests that little (if any) risk should be accepted for projects against relatively inconsequential pests for which the probability of success is low but that higher levels of risk should be accepted for projects targeting devastating pests for which the probability of successful biological control is high
One of the difficulties in performing a meaningful benefit/risk analysis in biological control is that the ‘currencies’ of risks and benefits are often different. While risks are typically expressed as declines in populations of native species, benefits are often expressed in monetary terms that apply to the commodity being protected. Thus, while biological control introductions can produce very favorable cost/benefit ratios (Gutierrez et al. 1999; Culliney 2005), this may not be directly relevant to environmental risks. There are ways around these difficulties however. For instance, valuation methods have been developed that can quantify externalities affected by biological control such as biodiversity protection, improved environmental quality or improved human health (Naranjo et al. 2015). Some of these analyses quantify the ecosystem services attributable to naturally occurring biological control (Costanza et al. 1997; Losey and Vaughan 2006). An extension of these analyses to classical biological control would likely reveal previously unrecognized environmental benefits of releases. Analyses such as these would surely include environmental benefits of reduced pesticide use. This approach was taken in the study done by Bigler and Kölliker-Ott (2006) described above where the risks of an insecticide to biodiversity were explicitly compared with those associated with parasitoid releases. Environmental costs associated with pesticide use can also be expressed in terms of the greenhouse gasses that are emitted during the processes of manufacturing, transporting and applying these compounds (Heimpel et al. 2013). To the extent that biological control reduces pesticide use, benefits in reduced greenhouse gas emissions can then be calculated. These and other methods could be used to capture a broader set of benefits that are produced by biological control.
Lastly, it should be noted that targeting invasive species in natural areas leads to a more straight-forward comparison between the benefits and risks of biological control introductions since in this case both are in the same currency: biodiversity. In these cases, benefits are measured in the protection of a particular set of native species from invasive species and the risks are to a different set of native species that may suffer direct or indirect non-target effects from the introduced agent. Comparisons between the number of native species protected versus endangered by biological control introductions must include an ecological valuation of these species among other factors in order to be relevant for biological control decision-making (van Driesche et al. 2016).
Conclusions
Classical biological control has protected our society and the environment from over 200 invasive insect and 50 invasive weed species over the past 130 years. Among these can be included devastating agricultural pests such as the cassava mealybug (Phenacoccus manihoti Matile-Ferrero) (Pseudococcidae) in Africa which threatened widespread food shortages (Neuenschwander 2001), and equally devastating invaders in natural areas such as the melaleuca tree in the Everglades National Park in the USA which threatens an entire ecosystem (Tipping et al. 2009). Against these successes must be weighed a much smaller number of introductions—representing less than 1% of cases—that have had negative unintended consequences. These risks began to be highlighted and taken seriously in the 1980s and during the 1990s a paradigm shift occurred in the biological control community in which risks began to be carefully monitored and models of risk assessment developed. This focus on risks was critically important for the growth and maturity of the science of biological control and the practice of classical biological control is much safer as a result. However, the pace of introductions has declined during the same time in some countries leading to likely opportunity costs. We are encouraged by the development of a second maturation step into a paradigm in which potential benefits and risks of biological control introductions are carefully balanced to arrive at solutions that maximize net social and environmental benefits. If this can be combined with other improvements such as effective dialogue and consultation between regulators, scientists and other stakeholders, and effective time-limits for key steps in the process, then the future of safe and effective classical biological control for the benefit of society will be more secure. This general approach is already being adopted in biological control regulations in some countries (notably New Zealand) but there is still a need for formal risk/benefit conceptual models that can guide decision making, and revised guidelines that take this into account to assist countries less experienced in classical biological control in their decision making. We hope this paper will contribute to this process.
References
Andraca-Gomez G, Ordano M, Boege K, Dominguez CA, Pinero D, Perez-Ishiwara R, Perez-Camacho J, Canizares M, Fornoni J (2015) A potential invasion route of Cactoblastis cactorum within the Caribbean region matches historical hurricane trajectories. Biol Invasions 17:1397–1406
Barratt BIP, Moeed A (2005) Environmental safety of biological control: policy and practice in New Zealand. Biol Control 35:247–252
Beddington JR, Free CA, Lawton JH (1978) Characteristics of successful natural enemies in models of biological control of insect pests. Nature 273:513–519
Benson J, van Driesche RG, Pasquale A, Elkinton J (2003) Introduced braconid parasitoids and range reduction of a native butterfly in New England. Biol Control 28:197–213
Bigler F, Kölliker-Ott UM (2006) Balancing environmental risks and benefits: a basic approach. In: Bigler F, Babendreier D, Kuhlmann U (eds) Environmental impact of invertebrates for biological control of arthropods. CABI Publishing, Oxon, pp 273–286
Bigler F, Babendreier D, Kuhlmann U (2006) Environmental impact of invertebrates for biological control of arthropods: methods and risk assessment. CABI Publising, Oxon
Blower SM, Koelle K, Kirschner DE, Mills J (2001) Live attenuated HIV vaccines: predicting the tradeoff between efficacy and safety. Proc Natl Acad Sci USA 98:3618–3623
Boettner GH, Elkinton JS, Boettner CJ (2000) Effects of a biological control introduction on three nontarget native species of saturniid moths. Conserv Biol 14:1798–1806
Caltagirone LE, Doutt RL (1989) The history of the vedalia beetle importation to California and its impact on the development of biological control. Annu Rev Entomol 34:1–16
Carvalheiro LG, Buckley YM, Ventim R, Fowler SV, Memmott J (2008) Apparent competition can compromise the safety of highly specific biological control agents. Ecol Lett 11:690–700
Civeyrel L, Simberloff D (1996) A tale of two snails: is the cure worse than the disease? Biodivers Conserv 5:1231–1252
Cock MJW, Day RK, Hinz H, Pollard KM, Thomas SE, Williams FE, Witt ABR, Shaw RH (2015) The impacts of some classical biological control successes. CAB Rev 10:1–57
Cock MJW, Murphy ST, Kairo MTK, Thompson E, Murphy RJ, Francis AW (2016) Trends in the classical biological control of insect pests by insects: an update of the BIOCAT database. BioControl 61:349–363
Coll M, Guershon M (2002) Omnivory in terrestrial arthropods: mixing plant and prey diets. Annu Rev Entomol 47:267–298
Costanza R, d’Arge R, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeen S, O’Neill RV, Paruelo J, Raskin RG, Sutton P, van den Belt M (1997) The value of the world’s ecosystem services and natural capital. Nature 387:253–260
Crawley MJ (1989) The successes and failures of weed biocontrol using insects. Biocontrol News Info 10:213–223
Culliney TW (2005) Benefits of classical biological control for managing invasive plants. Crit Rev Pl Sci 24:131–150
De Clercq P, Mason PG, Babendreier D (2011) Benefits and risks of exotic biological control agents. BioControl 56:681–698
Denoth M, Frid L, Myers JH (2002) Multiple agents in biological control: improving the odds? Biol Control 24:20–30
Ehlers R-U (2011) Regulation of biological control agents and the EU policy support action REBECA. In: Ehlers R-U (ed) Regulation of biological control agents. Springer, Dordrecht, pp 2–23
Esler KJ, van Wilgen BW, te Roller K, Wood AR, van der Merwe JH (2010) A landscape-level assessment of the long-term integrated control of an invasive shrub in South Africa. Biol Invasions 12:211–218
Evans EW (2004) Habitat displacement of North American ladybirds by an introduced species. Ecology 85:637–647
Follett PA, Duan JJ (2000) Nontarget effects of biological control. Kluwer Academic Publishers, Norwell
Greathead DJ, Greathead AH (1992) Biological control of insects pests by insect parasitoids and predators: the BIOCAT database. Biocontrol News Inf 13:61N–67N
Gurr GM, Wratten SD (2000) Biological control: measures of success. Kluwer, Dordrecht
Gutierrez AP, Caltagirone LE, Meikle W (1999) Evaluation of results, economics of biological control. In: Bellows TS, Fisher TW (eds) Handbook of biological control. Academic, San Diego, pp 243–252
Hajek AE, McManus ML, Junior ID (2005) Catalogue of introductions of pathogens and nematodes for classical biological control of insects and mites. USDA Forest Service, Forest Health Technology Enterprise Team, Morgantown
Hajek AE, McManus DP, Delalibera I Jr (2007) A review of introductions of pathogens and nematodes for classical biological control of insects and mites. Biol Control 41:1–13
Hall RW, Ehler LE (1979) Rate of establishment of natural enemies in classical biological control. Bull Entomol Soc Am 25:280–282
Hall RW, Ehler LE, Bisabri-Ershadi B (1980) Rate of success in classical biological control of arthropods. Bull Entomol Soc Am 26:111–114
Harmon JP, Stephens E, Losey J (2007) The decline of native coccinellids (Coleoptera: Coccinellidae) in the United States and Canada. J Ins Cons 11:85–94
Harrison L, Moeed A, Sheppard A (2005) Regulation of the release of biological control agents of arthropods in New Zealand and Australia. In: Hoddle M (ed) International symposium on biological control of arthropods. US Forest Service, Davos, pp 715–725
Hawkins BA, Cornell HV (1994) Maximum parasitism rates and successful biological control. Science 266:1886
Heimpel GE, Mills NJ (2017) Biological control: ecology and applications. Cambridge University Press, Cambridge
Heimpel GE, Yang Y, Hill J, Ragsdale DW (2013) Environmental consequences of invasive species: greenhouse gas emissions of insecticide use and the role of biological control in reducing emissions. PLoS ONE 8(8):e72293
Hennemann ML, Memmott J (2001) Infiltration of a Hawaiian community by introduced biological control agents. Science 293:1314–1316
Hill R, Campbell D, Hayes L, Corin S, Fowler S (2013) Why the New Zealand regulatory system for introducing new biological control agents works. In: XIII International Symposium on Biological Control of Weeds, vol 2011. Waikoloa, Hawaii, USA, pp 75–83
Hinz H, Schwartzlander M, Gassmann A, Bourchier RS (2014) Successes we may not have had: a retrospective analysis of selected weed biological control agents in the United States. Inv Plant Sci Manag 7:565–579
Hoddle MS, Crespo Ramirez C, Hoddle CD, Loayza J, Lincango PM, van Driesche RG, Causton CE (2013) Post release evaluation of Rodolia cardinalis (Coleoptera: Coccinellidae) for control of Icerya purchasi (Hemiptera: Monophlebidae) in the Galapagos Islands. Biol Control 67:262–274
Holt RD, Hochberg ME (2001) Indirect interactions: community modules and biological control: a theoretical perspective. In: Wajnberg E, Scott JK, Quimby PC (eds) Evaluating indirect ecological effects of biological control. CABI Press, Oxon, pp 13–38
Howarth FG (1991) Environmental impacts of classical biological control. Annu Rev Entomol 36:485–509
Hunt EJ, Kuhlmann U, Sheppard A, Qin T-K, Barratt BIP, Harrison L, Mason PG, Parker D, Flanders RV, Goolsby J (2008) Review of invertebrate biological control agent regulation in Australia, New Zealand, Canada and the USA: recommendations for a harmonized European system. J Appl Entomol 132:89–123
Kaser JM, Heimpel GE (2015) Linking risk and efficacy in biological control host-parasitoid models. Biol Control 90:49–60
Kaser JM, Ode PJ (2016) Hidden risks and benefits of natural enemy mediated indirect effects. Curr Opin Ins Sci 14:105–111
Kimberling DN (2004) Lessons from history: predicting successes and risks of intentional introductions for arthropod biological control. Biol Invasion 6:301–318
Lane SD, Mills NJ, Getz WM (1999) The effects of parasitoid fecundity and host taxon on the biological control of insect pests: the relationship between theory and data. Ecol Ent 24:181–190
Lazo-Langer JA, Rodger MA, Barrowman NJ, Ramsay T, Wells PS, Coyle DA (2012) Comparing multiple competing interventions in the absence of randomized trials using clinical risk-benefit analysis. BMC Med Res Meth 12:3
Losey JE, Vaughan M (2006) The economic value of ecological services provided by insects. BioScience 56:311–323
Louda SM, O’Brien CW (2002) Unexpected ecological effects of distributing the exotic weevil, Larinus planus (F.), for the biological control of Canada thistle. Cons Biol 16:717–727
Louda SM, Kendall D, Connor J, Simberloff D (1997) Ecological effects of an insect introduced for the biological control of weeds. Science 277:1088–1090
Louda SM, Pemberton RW, Johnson MT, Follett PA (2003) Nontarget effects-the Achilles’ heel of biological control? Annu Rev Entomol 48:365–396
Luck RF (1990) Evaluation of natural enemies for biological control: a behavioral approach. Trends Ecol Evol 5:196–199
Lundgren JG (2009) Relationships of natural enemies and non-prey foods. Springer, Dordrecht
Maron M, Cockfield G (2008) Managing trade-offs in landscape restoration and revegetation projects. Ecol Appl 18:2041–2049
McBride MF, Wilson KA, Burger J, Fang YC, Lulow M, Olson D, O’Connell M, Possingham HP (2010) Mathematical problem definition for ecological restoration planning. Ecol Modell 221:2243–2250
McEvoy PB, Coombs EV (2000) Why things bite back: unintended consequences of biological weed control. In: Follett PA, Duan JJ (eds) Nontarget effects of biological control. Kluwer, Dordrecht, pp 167–194
Messing RH, Wright MG (2006) Biological control of invasive species: solution or pollution? Front Ecol Environ 4:132–140
Mills NJ (2006) Accounting for differential success in the biological control of homopteran and lepidopteran pests. N Zeal J Ecol 30:61–72
Minckley WL, Deacon JE (1968) Southwestern fishes and the enigma of ‘endangered species’. Science 159:1424–1432
Moeed A, Hickson R, Barratt BIP (2006) Principles of environmental risk assessment with emphasis on the New Zealand perspective. In: Bigler F, Babendreier D, Kuhlmann U (eds) Environmental impact of invertebrates for biological control of arthropods. CABI Publishing, Oxon, pp 241–253
Murdoch WW, Chesson J, Chesson PL (1985) Biological control in theory and practice. Am Nat 125:344–366
Naranjo SE, Ellsworth PC, Frisvold GB (2015) Economic value of biological control in integrated pest management of managed plant systems. Annu Rev Entomol 60:621–645
Neuenschwander P (2001) Biological control of the cassava mealybug in Africa: a review. Biol Control 21:214–229
Paynter QE, Fowler SV, Gourlay AH, Haines ML, Harman HM, Hona SR, Peterson PG, Smith LA, Wilson-Davey JRA, Winks CJ, Withers TM (2004) Safety in New Zealand weed biocontrol: a nationwide survey for impacts on non-target plants. New Zeal Plant Prot 57:102–197
Pearson DE, Callaway RM (2003) Indirect effects of host-specific biological control agents. Trends Ecol Evol 18:456–461
Pearson DE, Callaway RM (2006) Biological control agents elevate hantavirus by subsidizing deer mouse populations. Ecol Lett 9:443–450
Pemberton RW (2000) Predictable risks to native plants in weed biological control. Oecologia 125:489–494
Rose KE, Louda SM, Rees M (2005) Demographic and evolutionary impacts of native and invasive insect herbivores on Cirsium canescens. Ecology 86:453–465
Roy HE, Adriaens T, Isaac NJB, Kenis M, Onkelinx T, San Martin G, Brown PMJ, Hautier L, Poland R, Roy DB, Comont R, Eschen R, Frost R, Zindel R, van Vlaenderen J, Nedved O, Ravn HP, Gregoire J-C, de Biseau J-C, Maes D (2012) Invasive alien predator causes rapid declines of native European ladybirds. Divers Distr 18:717–725
Seaman GA, Randall JE (1962) The mongoose as a predator in the Virgin Islands. J Mammol 43:544–546
Sheppard AW, Hill R, DeClerck-Floate RA, McClay A, Olckers T, Quimby PCJ, Zimmermann HG (2003) A global review of risk-benefit-cost analysis for the introduction of classical biological control agents against weeds: a crisis in the making? Biocontrol News Info 24:91N–108N
Simberloff D, Stiling P (1996) How risky is biological control? Ecology 77:1965–1974
Stephens AE, Srivastava DS, Myers JH (2013) Strength in numbers? Effects of multiple natural enemy species on plant performance. Proc R Soc London B 280:20122756
Stiling P (1990) Calculating the establishment rates of parasitoids in classical biological control. Am Entomol 1990(Fall):225–230
Stiling P (1993) Why do natural enemies fail in classical biological control programs? Am Entomol 39:31–37
Stiling P, Moon D, Gordon D (2004) Endangered cactus restoration: mitigating the non-target effects of a biological control agent (Cactoblastis cactorum) in Florida. Restor Ecol 12:605–610
Story JM, Smith L, Corn JG, White LJ (2008) Influence of seed head-attacking biological control agents on spotted knapweed reproductive potential in western Montana over a 30-year period. Environ Entomol 37:510–519
Suckling DM (2013) Benefits from biological control of weeds in New Zealand range from negligible to massive: a retrospective analysis. Biol Control 66:27–32
Suckling DM, Sforza RF (2014) What magnitude are observed non-target impacts from weed biocontrol? PLoS ONE 9(1):e84847
Symondson WOC, Sunderland KD, Greenstone MH (2002) Can generalist predators be effective biological control agents? Annu Rev Entomol 47:561–594
Tipping PW, Martin MR, Nimmo KR, Pierce RM, Smart MD, White EB, Madeira PT (2009) Invasion of a West Everglades wetland by Melaleuca quinquenervia countered by classical biological control. Biol Control 48:73–78
Urban MC, Phillips BL, Skelly DK, Shine R (2007) The cane toad’s (Chaunus [Bufo] marinus) increasing ability to invade Australia is revealed by a dynamically updated range model. Proc R Soc London B 274:1413–1419
van Driesche RG, Hoddle M (1997) Should arthropod parasitoids and predators be subject to host range testing when used as biological control agents? Agric Hum Val 14:211–226
van Driesche R, Reardon R (2004) Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice. FHTET, USDA Forest Service, Morgantown
van Driesche RG, Carruthers RI, Center T, Hoddle MS, Hough-Goldstein J, Morin L, Smith L, Wagner DL, Blossey B, Brancatini V, Casagrande R, Causton CE, Coetzee JA, Cuda J, Ding J, Fowler SV, Frank JH, Fuester R, Goolsby J, Grodowitz M, Heard TA, Hill MP, Hoffmann JH, Huber J, Julien M, Kairo MTK, Kenis M, Mason P, Medal J, Messing R, Miller R, Moore A, Neuenschwander P, Newman R, Norambuena H, Palmer WA, Pemberton R, Panduro AP, Pratt PD, Rayamajhi M, Salom S, Sands D, Schooler S, Schwarzlander M, Sheppard A, Shaw R, Tipping PW, van Klinken RD (2010) Classical biological control for the protection of natural ecosystems. Biol Control 54:S2–S33
van Driesche R, Simberloff D, Blossey B, Causton C, Hoddle M, Marks C, Heinz K, Wagner D, Warner K (2016) Integrating biological control into conservation practice. Wiley, Oxford
van Klinken RD, Edwards OR (2002) Is host-specificity of weed biological control agents likely to evolve rapidly following establishment? Ecol Lett 5:590–596
van Lenteren JC, Babendreier D, Bigler F, Burgio G, Hokkanen HMT, Kuske S, Loomans AJM, Menzler-Hokkanen I, van Rijn PCJ, Thomas MB, Tommasini MG, Zeng QQ (2003) Environmental risk assessment of exotic natural enemies used in inundative biological control. BioControl 48:3–38
van Lenteren JC, Bale J, Bigler F, Hokkanen HMT (2006) Assessing risks of releasing exotic biological control agents of arthropod pests. Annu Rev Entomol 51:609–634
van Wilgen BW, Moran VC, Hoffmann JH (2013) Some perspectives on the risks and benefits of biological control of invasive alien plants in the management of natural ecosystems. Environ Manag 52:531–540
Wajnberg E, Scott JK, Quimby PC (2001) Evaluating indirect ecological effects of biological control. CABI Press, Wallingford
Wapshere AJ (1974) A strategy for evaluating the safety of organisms for biological weed control. Ann Appl Biol 77:201–211
Wiggering H, Dalchow C, Glemnitz M, Helming K, Muller K, Schultz A, Stachow U, Zander P (2006) Indicators for multifunctional land use—Linking socio-economic requirements with landscape potentials. Ecolog Indicat 6:238–249
Winston RL, Schwartzlander M, Hinz H, Day MD, Cock MJW, Julien MH (2014) Biological control of weeds: a world catalogue of agents and their target weeds, 5th edn. USDA Forest Service, Forest Heath Technology Enterprise Team, Morgantown
Wright MG, Hoffmann MP, Kuhar TP, Gardner J, Pitcher SA (2005) Evaluating risks of biological control introductions: a probabilistic risk-assessment approach. Biol Control 35:338–347
Acknowledgements
We thank Jacques Brodeur and Russell Messing for the invitation to contribute a submission to this Special Issue, and IOBC for supporting the conference where these ideas were originally discussed. For discussion of ideas presented in the paper we thank Joe Kaser, Rob Venette, Anne Kapuscinksi, Peter Mason and Frances Homans. We also thank Russell Messing and two anonymous reviewers for useful comments on a previous version of this article and Jacqueline Nuzzo for help with formatting Fig. 1.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Russell Messing.
Rights and permissions
About this article

Cite this article
Heimpel, G.E., Cock, M.J.W. Shifting paradigms in the history of classical biological control. BioControl 63, 27–37 (2018). https://doi.org/10.1007/s10526-017-9841-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-017-9841-9