
Abstract
Molecular markers facilitate the quantification of parasitization of pest species and the distinction of related parasitoid species. We designed new markers that allow the distinction of often misidentified Diadegma semiclausum and D. fenestrale parasitization in field-collected diamondback moth (Plutella xylostella) larvae. The markers were applied to study if cornflower (Centaurea cyanus) companion plants increase parasitization of the diamondback moth by Diadegma parasitoids in cabbage fields, as they do in the Mamestra brassicae—Microplitis mediator pest-parasitoid pair. Among 1708 P. xylostella larvae analyzed, we found a high parasitization rate (72.2 % total, 41.7 % by D. semiclausum, 24.9 % by D. fenestrale, 5.6 % by both), but no significant effects of cornflower presence. Our results highlight the need for species-specific markers and the specific action of companion plants. To increase natural control in crop fields, habitat management needs to be tailored to—and acts on—specific target species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Various cabbage species are major food crops worldwide (Li 1970; Liu et al. 2000; Talekar and Shelton 1993). They are attacked by a wide array of herbivores, which are controlled by predators and parasitoids attacking all pest life-stages. Worldwide, the main cabbage pest is the diamondback moth (DBM) Plutella xylostella (Linnaeus, 1758) (Plutellidae), which is parasitized by Diadegma semiclausum (Hellen, 1949) and D. fenestrale (Holmgren, 1860) (Ichneumonidae) among others. Another important pest is the cabbage moth Mamestra brassicae (Linnaeus, 1758) (Noctuidae), which is parasitized by the hymenopteran endoparasitoids Microplitis mediator (Haliday, 1834) (Braconidae). DBM became resistant to almost all synthetic insecticides by around 1980 (Sun et al. 1986) and causes control costs over 1.4 billion US dollars per year, crop loss not included (Furlong et al. 2013). Studies conducted in Malaysia showed that insecticide application against DBM reduced rates of parasitization by parasitoids and that the recovery of natural parasitoid populations in combination with the controlled release of different parasitoids (D. semiclausum and D. collaris) led to a continued suppression of P. xylostella and an increased cabbage yield (Talekar and Shelton 1993). A recent study demonstrated that crop loss may increase in pesticide treated fields if the targeted pest is resistant to the pesticide used but its parasitoid is not (Bommarco et al. 2011).
There are two main strategies to control crop pests with natural enemies. Natural enemies can either be released (“inundative” or “inoculative biocontrol” (Landis et al. 2000)) or naturally occurring resident pest enemies (e.g. parasitoids) are strengthened by appropriate habitat management (“conservation biological control” (Pfiffner and Wyss 2004)), e.g. by adding companion plants that attract natural enemies (Belz et al. 2013) and provide crucial, otherwise lacking food sources such as floral or extrafloral nectar (Pfiffner et al. 2009). The latter approach requires a detailed understanding of the ecology of the involved crop, pest and natural enemy species as well as their interactions to ensure that the applied habitat management selectively benefits the natural enemies and not the pest.
It is often challenging to correctly determine the species of Lepidopteran pest larvae and their corresponding endoparasitoids. Traditionally, crop pest larvae were collected and maintained in the laboratory until parasitoids emerged and their species could be determined. This is very laborious and may give biased parasitization rates as host larvae have different mortality rates when parasitized by varying parasitoids. Alternatively, parasitization rates were assessed by dissection, which limits the number of individuals that can be analyzed per day and does not allow for reliable species identification in many cases. In Diadegma the available key based on adult morphology (Azidah et al. 2000) produces unreliable distinction between D. semiclausum and D. fenestrale and males can usually not be distinguished at all (Klaus Horstmann, personal communication). Furthermore, the available molecular marker supposedly specific for D. semiclausum (Traugott et al. 2006) turns out not to be species-specific as it amplifies both D. semiclausum and D. fenestrale (own data, not shown). In this study, we therefore developed species-specific molecular markers to unambiguously identify the species of the collected pest larvae and of parasitizing Diadegma wasps.
It has been shown in laboratory studies that the cornflower Centaurea cyanus L. attracts M. mediator, a larval parasitoid of the cabbage moth, with its odour (Belz et al. 2013; Géneau et al. 2013) and increases its longevity and fecundity (Géneau et al. 2012). It has also been shown that cornflowers increase parasitization rates of M. brassicae by M. mediator and increase cabbage yield in the field (Balmer et al. 2014). We thus applied our new molecular markers to DBM samples from that same study (Balmer et al. 2014) and tested whether cornflowers planted as companion plants in cabbage fields also increase the parasitization of DBM by its larval parasitoids D. semiclausum and D. fenestrale.
Materials and methods
Development of species-specific primers for P. xylostella, D. semiclausum and D. fenestrale
Parasitization of P. xylostella was determined by diagnostic multiplex PCR, targeting parasitoid DNA. For primer design, we amplified and sequenced a segment of the mitochondrial cytochrome c oxidase subunit I (COI) gene for several pest and parasitoid species collected in the field in Switzerland using universal primers LCO1490 and HCO2198 (Folmer et al. 1994). Additionally, we included published COI sequences of related species from other countries, which we retrieved from NCBI GenBank (Table 1) to ensure that the primers are species-specific and do not cross-amplify other species. To ensure the correct assignment of D. semiclausum and D. fenestrale, we used DNA from a laboratory-reared female D. fenestrale, which had been identified by a taxon specialist (Klaus Hostmann, Würzburg, Germany), as a reference. Because of the extensive sequence variation in P. xylostella COI and the importance of P. xylostella as pest of Brassicaceae, we used sequences from >200 P. xylostella individuals from 13 regions in Europe, Asia, Africa, Australia and North America—both field collected samples and data from GenBank (accessions GU094717.1, DQ076332.1-DQ076341.1, DQ076345.1-DQ076411.1, EF380067.1, EF380069.1, EF380071.1, EF380073.1, EF380075.1, EF380077.1, EF380079.1, EF380081.1, EF380083.1, EF380085.1, EF380087.1, EF380089.1, EF380091.1, EF380093.1, FJ412899.1, GU092596.1, GU092597.1, GU094715.1, GU094716.1, HQ683348.1, JN410809.1-JN410813.1). In contrast, D. semiclausum and D. fenestrale exhibited no COI sequence variation in 70 and 61 individuals from five and four different countries, respectively. Species specific primers were designed in COI regions that exhibited no variation within the target species but variation between the target species and other species. For Diadegma only one sequence per species was used as we did not find any intraspecific variation. DNA for primer design was extracted using the NucleoSpin Tissue XS kit (Macherey–Nagel, Düren, Germany) following the manufacturer’s protocol. Each 10 μl PCR reaction contained 5 μl of 2X Qiagen multiplex PCR master mix (Qiagen, Hilden, Germany), 0.2 μl of each primer [10 μM], 3.8 μl of ultrapure water, and 0.8 μl of DNA template. Sequencing was performed on an ABI 3130xl Sequencer (Applied Biosystems, Foster City, USA).
Development of multiplex PCR assay
The newly developed primers for P. xylostella, D. semiclausum, and D. fenestrale were combined in a multiplex PCR (Table 1). PCR conditions were optimized to detect parasitoid DNA inside P. xylostella samples. To this end, we amplified dilution series with samples containing different mixes of purified single-species DNA of the three species (undiluted DNA concentrations were between 137 and 185 ng µl-1). The mixes tested contained controls without DNA, 1:100 dilutions of each parasitoid mixed with the host, only the two parasitoids, the three-way mix, and the same mixtures but using 1:1,000 parasitoid dilutions. All predicted bands were readily detectable in all samples. Consistently good results were achieved in 10 μl reactions containing 5 μl of Qiagen multiplex PCR master mix (Qiagen, Hilden, Germany), 0.1 μl of each P. xylostella primer [10 μM], 0.21 μl of each D. fenestrale primer [10 μM], 0 575 μl of each D. semiclausum primer [10 μM], 2.43 μl of ultrapure water, and 0.8 μl of DNA template. The PCR program for the amplification had the following conditions: Activation of hot start Taq for 5 min at 95 °C followed by 36 cycles of denaturation at 94 °C for 30 s, annealing at 58.7 °C for 1 min, extension at 72 °C for 1 min, and final extension at 72 °C for 7 min. This protocol was used for all field samples.
Experimental setup in the field
Eight commercial organic white cabbage (Brassica oleracea) fields in Switzerland were used for the experiment in 2009 (Table 3). In each field we marked one 12 m × 12 m area in which we planted one cornflower per m2 between cabbage heads and one equally sized area without additional cornflowers at least 50 m apart (Fig. 1). Fifty meters were chosen as a compromise to space the plots as far apart within the same field as possible to account for parasitoid mobility and the limited sizes of the smaller of the available fields, which are generally not very large in Switzerland. The central 8 m × 8 m of both areas served as sampling plots. In the sampling plots, sixteen equally spaced cabbage heads were marked as sampling points, which were checked for lepidopteran pest species and sampled on 3–4 dates, depending on larval densities per field (Table 4). Samples analyzed in this study were collected simultaneously and on the same fields as the M. brassicae samples analyzed in a parallel study (Balmer et al. 2014). No pesticides were applied in the experimental areas during the study period.

Experimental setup. Experiments were conducted on commercial white cabbage field (light grey area). Per field, two sampling plots (solid boxes) were set up, one of which was inside a larger area with additionally cornflowers (Centaurea cyanus) planted as companion plants between cabbage heads (dotted area). n = 8 fields
Field sampling
All larvae (in all life-stages) of P. xylostella, M. brassicae, P. brassicae and P. rapae found on each screened cabbage head were collected individually in 1.5 ml microfuge tubes (Vaudaux-Eppendorf) frozen at −80 °C for taxonomic analysis as reported before (Balmer et al. 2014). To avoid an influence of the collection of all larvae per head on insect densities recorded on the next date, the points of sampling were shifted by one plant (cabbage head) between sampling dates. Thus, on subsequent sampling dates, insects were collected first from the predefined sampling point, then from the cabbage head to the left of it, then to the right, then the sampling point again.
DNA extraction and PCR amplification of field samples
2166 lepidopteran pest larvae were examined in the present study. Individual larvae were crushed in 1.5 ml microfuge tubes (Vaudaux-Eppendorf) in 300 μl extraction buffer, containing 5 % Chelex® 100 (Bio-Rad Laboratories, Cressier, Switzerland), 1 M Tris HCl pH8, 0.5 M EDTA pH8, 6 mg ml-1 proteinase K, and three SiLibeads Typ ZY zircon beads (Sigmund Lindner GmbH, Warmensteinach, Germany) using a Savant FastPrep® FP120 bead beater (Qbiogene, Irvine, USA) for 15 s at a speed of 5.5 m s-1. Crushed samples were incubated overnight at 56 °C. After incubation the samples were heated to 95 °C for 10 min and stored at −80 °C. For PCR the tubes were centrifuged for 3 min at 6,000 rpm and the supernatant used as DNA template. Large larvae were cut and extracted in multiple tubes, using a maximum of 1 cm of larva per tube and extractions combined again in the end.
All mulitplex PCRs were performed in 96-well plates containing three positive controls with DNA of P. xylostella, D. semiclausum or D. fenestrale and a negative control with only water and the PCR mix on a Veriti 96well Thermal Cycler (Applied Biosystems, Foster City, USA) using the conditions described above. To verify the pest species and check for parasitization, 5 μl of each PCR product were mixed with 1 μl of loading dye (Qiagen, Hilden, Germany) and analyzed by gel electrophoresis on a 1.5 % agarose gel containing 1x Gel Red (Biotium, Hayward, USA). For all species the bands from a few individuals were cut out, purified using the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany) and sequenced on an ABI 3130xl Sequencer (Applied Biosystems, Foster City, USA) to verify that each band length corresponded to the correct species. We analyzed all bands in five field samples: one with only a P. xylostella band, one each with a P. xylostella and a D. semiclausum or D. fenestrale band, respectively, and two field samples with all three bands (i.e. doubly parasitized). All bands yielded the expected sequences.
Data analysis
Differences in parasitization rate between plots with and without cornflower companion plants were assessed with a generalized linear mixed-effects model (glmm) with binominal data distribution (parasitization yes/no) and field identity as random factor. Glmms were calculated with function glmmPQL in R version 2.15.1 (R Development Core Team 2009), which produces t and p values as output. Sequential time points were pooled per plot to integrate over the entire growing season. Differences in P. xylostella abundances between plots with and without cornflower companion plants were assessed by paired t tests.
Results
Species-specificity of developed primers
All primers were designed in a way that they do not amplify DNA from closely related parasitoid species and other common pest species. Furthermore, PCR products of each taxon are of different sizes, so that the targeted species are easily distinguishable on agarose gels and multiple parasitizations can readily be detected (Fig. 2). The optimized multiplex PCR reliably amplified DNA of P. xylostella and of single and double parasitizations by D. fenestrale and D. semiclausum within a single P. xylostella larva.

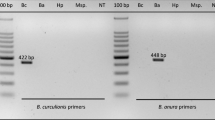
1.5 % agarose gel image of multiplex PCR on partial COI. Lane 1 100 bp ladder, lanes 2–4 DNA from laboratory reared individuals: lane 2 Plutella xylostella (163 bp), lane 3 P. xylostella parasitized by Diadegma fenestrale (249 bp), lane 4 P. xylostella parasitized by D. semiclausum (392 bp), lane 5 P. xylostella field sample parasitized by D. fenestrale and D. semiclausum, lane 6 Mamestra brassicae parasitized by Microplitis mediator, lane 7 negative control with reaction mix and distilled water. See materials and methods for details on the multiplex PCR
Parasitization rates
Among the 2166 lepidopteran larvae analyzed by PCR, 1708 were identified as P. xylostella (the others were identified as early instars of M. brassicae by M. brassicae-specific markers (Balmer et al. 2014)), of which 1234 (72.2 %) were found to be parasitized: 426 (24.9 %) only by D. fenestrale, 715 (41.9 %) only by D. semiclausum and 95 (5.6 %) by both Diadegma species (Fig. 3). Parasitization rates of P. xylostella larvae by either species were not significantly different between plots with cornflowers and those without (D. fenestrale, p = 0.24, df = 1699, t value = 1.18; D. semiclausum, p = 0.63, df = 1699, t value = −0.48; double parasitization, p = 0.38, df = 1699, t value = 0.88). In general, D. semiclausum parasitized more larvae than D. fenestrale. The rate of double parasitization by both species was significantly lower than expected if parasitization by the two species were independent (Chi2= 254.2, df = 1, p < 0.001) indicating mutual avoidance or suppression of the species. The proportion of the rarer species was at least 17 % in all eight fields and the proportion of double infections varied between 1.0 and 11.5 %. The abundance of P. xylostella did not differ between plots with and without cornflower (p = 0.48, df = 7, t = 0.75).

Mean percentage (±95 % CI of the binomial distribution) of Plutella xylostella larvae parasitized only by Diadegma fenestrale (‘Dfe’), only by D. semiclausum (‘Dse’) or by both species (‘Dfe&Dse’) per treatment (solid circles without cornflowers, open circles with cornflowers as companion plants). Significance of differences between treatments was assessed by Welch two-sample t tests
Discussion
This study had three aims: (1) to develop molecular markers to reliably distinguish the two congeneric larval parasitoid species D. semiclausum and D. fenestrale in all life-stages, (2) to quantify their relative importance for parasitization of P. xylostella, and (3) to investigate how the addition of specifically chosen non-crop plants can reduce the damage caused by crop pests without insecticide application.
Our laboratory tests using DNA from a wide range of host and Diadegma species and our in silico analyses of the corresponding DNA sequences showed that the newly developed markers reliably determine the host species and distinguish D. semiclausum and D. fenestrale within parasitized DBM larvae. The analysis of >2,000 field collected pest larvae shows that the two species and their host can efficiently be identified in one single PCR step.
Earlier studies had assumed that primarily or only D. semiclausum was present in our region (Pfiffner et al. 2009). Our new molecular markers show that D. fenestrale plays an important role in parasitizing P. xylostella as well. This highlights the importance of reliable markers for species determination in parasitized pest samples from the field and for a better ecological understanding of pest and beneficial insect networks. Such markers are especially important for species that are difficult to distinguish morphologically like D. semiclausum and D. fenestrale. The challenge of reliably identifying these Diadegma species using the available morphological keys (Azidah et al. 2000) was evident from the fact that using our new molecular markers we detected a total of 14 misidentifications by two colleagues among 131 adult Diadegma samples we received for a related study. Mostly D. fenestrale were falsely identified as D. semiclausum. The published records indicate that D. fenestrale was reared for inundative biocontrol in England and was also shipped to New Zealand (Hardy 1938). But most reports indicate that only D. semiclausum has been introduced in different countries as a biocontrol agent (Furlong et al. 2012; Sun et al. 1986; Talekar and Shelton 1993). The apparent difficulty to correctly discriminate the two species suggests that D. fenestrale may have been introduced along with (or instead of) D. semiclausum on multiple occasions.
Our results show a very high parasitization rate (72.2 %) of P. xylostella by both Diadegma species. However, in contrast to what was simultaneously found for parasitization of M. brassicae by M. mediator on the same fields (Balmer et al. 2014), we did not find significantly increased parasitization in plots with cornflowers as companion plants. We believe that these results should serve as a call for caution that habitat management can indeed have very species-specific effects. Our laboratory experiments have previously demonstrated that C. cyanus has beneficial effect on M. mediator populations, while the effects on Diadegma spp. were much less pronounced (Géneau et al. 2012). Our field experiment thus corroborates our laboratory-based results that C. cyanus benefits M. mediator but not Diadegma species. Laboratory experiments appear to be valuable to find appropriate companion plant species for conservation biological control, but these plants have very species-specific effects. An alternative explanation for the lack of a treatment effect is always that the experimental design was inappropriate for demonstrating existing effects. In our case, it could be argued that 50 meters between plots with and without cornflower, as in our study, is not enough given the high mobility of Ichneumonid larval parasitoids. However, within the exact same set-up we did find significant effects on parasitization by M. mediator with comparable mobility (Balmer et al. 2014) and in an earlier study with egg parasitoids we demonstrated significant effects on parasitization by egg parasitoids over distances that would intuitively seem too close to efficiently demonstrate treatment effects (Balmer et al. 2013). Furthermore, our experimental design with paired treatment blocks replicated on independent fields is certainly a strong one. We would therefore argue that it is most plausible that indeed our cornflower treatment had no significant effect.
Pest population densities can vary substantially between sampling years. In the study area, P. xylostella was much more frequent in 2009 than in 2010 (unpublished data). The situation was the opposite for M. brassicae where 2009 showed low, and 2010 high densities. The variability in population size between years is in line with a study conducted in Kenya, which showed strong inter-year variability of population sizes (Lohr et al. 2007). Prediction of future pest population sizes would allow the appropriate companion plants, targeting the most prevalent pest species, to be sown among the crops. Combining multiple companion plants, optimized for different parasitoids, may increase total parasitization and ensure that the populations of the most relevant parasitoids are boosted. Parasitization rates were generally high despite the apparent variability of the host population size between years, indicating the presence of alternative hosts ensuring survival of the parasitoid species.
We found only 5.6 % doubly parasitized larvae despite both Diadegma species parasitizing at least 17 % of hosts each in every single field. 5.6 % is significantly less than would be expected if both species parasitized at random, i.e. without being influenced by parasitization by the other species. This suggests that the two Diadegma species (or at least one of them) either actively avoid already parasitized larvae, presumably to increase the chances that their own offspring can finish the full development until eclosion, or that they can eliminate other parasitoids from co-parasitized hosts.
Our results further suggest that parasitization rates of P. xylostella by Diadegma spp. are close to saturation. We found 72.2 % of the pest larvae to be parasitized and this is certainly an underestimation because our PCR method may miss some parasitizations and because especially young larvae may have been collected before being parasitized. The high natural parasitization rates suggest that larval parasitoids may not be well suited as biocontrol agents because they are obviously under strong natural selection not to kill their hosts before they have completed their own development.
Artificial releases of parasitoids could further increase parasitoid population size and parasitization rate. Gichini et al. (2008) showed that releasing low numbers of parasitoids (D. semiclausum) increased parasitization of brassicaceous pests in Kenya from year to year. However, in contrast to Kenya, where D. semiclausum was introduced as an exotic parasitoid, the species is already established and occurs naturally in the region studied here. Since parasitization rates are high, even in a year with high populations of P. xylostella, additional releases may not have strong effects on pest parasitization rates. Other Diadegma species that do not occur in Europe are known to attack P. xylostella (Azidah et al. 2000). It is possible that other species would be more effective at inactivating P. xylostella than D. semiclausum and D. fenestrale. However, it is not foreseeable how the release of other species would impact the ecological balance (De Clercq et al. 2011) and the high parasitization rates found in this study do not support such an approach.
On theoretical grounds, egg parasitoids such as Trichogramma spp. (or predators), might be a more efficient alternative to larval parasitoids as the parasitization of an egg results in its death, without the pest first developing into a larva and causing damage to the crop.
In conclusion, we were unable to show an effect of cornflowers on parasitization rates of P. xylostella. This is in contrast to increased parasitization of M. brassicae by M. mediator, for which cornflowers were specifically chosen (Belz et al. 2013; Géneau et al. 2012, 2013) and to positive effects cornflowers have on predators (Ditner et al. 2013). We would argue that these combined findings primarily highlight that habitat management effects are very species-specific. They need to be tailored to specific tasks and target organisms. There is mounting evidence that flowering plants can influence parasitization rates if they have been shown to specifically benefit the respective parasitoid species (Balmer et al. 2013, 2014). Further experiments should investigate if an approach using multiple companion plants targeting different beneficial species would be able to suppress pest damage further. The species-specific molecular markers for Diadegma spp. developed here should greatly facilitate detailed population studies of Diadegma spp..
References
Azidah AA, Fitton MG, Quicke DLJ (2000) Identification of the Diadegma species (Hymenoptera: Ichneumonidae, Campopleginae) attacking the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Bull Entomol Res 90:375–389
Balmer O, Pfiffner L, Schied J, Willareth M, Leimgruber A, Luka H, Traugott M (2013) Noncrop flowering plants restore top-down herbivore control in agricultural fields. Ecol Evol 3:2634–2646
Balmer O, Géneau CE, Belz E, Weishaupt B, Förderer G, Moos S, Ditner N, Juric I, Luka H (2014) Wildflower companion plants increase pest parasitation and yield in cabbage fields: experimental demonstration and call for caution. Biol Control 76:19–27
Belz E, Kölliker M, Balmer O (2013) Olfactory attractiveness of flowering plants to the parasitoid Microplitis mediator: potential implications for biological control. BioControl 58:163–173
Bommarco R, Miranda F, Bylund H, Björkman C (2011) Insecticides suppress natural enemies and increase pest damage in cabbage. J Econ Entomol 104:782–791
De Clercq P, Mason PG, Babendreier D (2011) Benefits and risks of exotic biological control agents. BioControl 56:681–698
Ditner N, Balmer O, Beck J, Blick T, Nagel P, Luka H (2013) Effects of experimentally planting non-crop flowers into cabbage fields on the abundance and diversity of predators. Biodivers Conserv 22:1049–1061
Folmer Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotech 3(5):294–299
Furlong MJ, Wright DJ, Dosdall LM (2013) Diamondback moth ecology and management: problems, progress, and prospects. Annu Rev Entomol 58:517–541
Géneau CE, Wäckers FL, Luka H, Daniel C, Balmer O (2012) Selective flowers to enhance biological control of cabbage pests by parasitoids. Basic Appl Ecol 13:85–93
Géneau CE, Wäckers FL, Luka H, Balmer O (2013) Effects of extrafloral and floral nectar of Centaurea cyanus on the parasitoid wasp Microplitis mediator: olfactory attractiveness and parasitization rates. Biol Control 66:16–20
Gichini G, Löhr B, Rossbach A, Nyambo B, Gathu R (2008) Can low release numbers lead to establishment and spread of an exotic parasitoid: the case of the diamondback moth parasitoid, Diadegma semiclausum (Hellen), in East Africa. Crop Prot 27:906–914
Hardy JE (1938) Plutella maculipennis, curt., its natural and biological control in England. Bull Entomol Res 29(04):343–372
Landis DA, Wratten SD, Gurr GM (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu Rev Entomol 45:175–201
Li HL (1970) Origin of cultivated plants in southeast Asia. Econ Bot 24:3–8
Liu SS, Wang XG, Guo SJ, He JH, Shi ZH (2000) Seasonal abundance of the parasitoid complex associated with the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) in Hangzhou, China. Bull Entomol Res 90:221–231
Lohr B, Gathu R, Kariuki C, Obiero J, Gichini G (2007) Impact of an exotic parasitoid on Plutella xylostella (Lepidoptera: Plutellidae) population dynamics, damage and indigenous natural enemies in Kenya. Bull Entomol Res 97:337–350
Pfiffner L, Wyss E (2004) Use of sown wildflower strips to enhance natural enemies of agricultural pests. Ecological engineering for pest management. Advances in habitat manipulation for arthropods. CSIRO Publishing, Collingwood, p 256
Pfiffner L, Luka H, Schlatter C, Juen A, Traugott M (2009) Impact of wildflower strips on biological control of cabbage lepidopterans. Agric Ecosyst Environ 129:310–314
R Development Core Team (2009) R: a language and environment for statistical computing. Vienna, Austria
Sun CN, Wu TK, Chen JS, Lee WT (1986) Insecticide resistance in diamondback moth. In: Talekar NS, Griggs TD (eds) Diamondback moth management. Proceedings of the first international workshop, Taiwan, 11-15 March, 1985, pp 359-371
Talekar NS, Shelton AM (1993) Biology, ecology, and management of the Diamondback moth. Annu Rev Entomol 38:275–301
Traugott M, Zangerl P, Juen A, Schallhart N, Pfiffner L (2006) Detecting key parasitoids of lepidopteran pests by multiplex PCR. Biol Control 39:39–46
Acknowledgments
We thank all farmers for making their fields available, K. Oude-Lensferink, J. Preukschas, A. Moesch, D. Eglin, G. Förderer, M. Fürst and C. Gerber for help with field work, in the laboratory and during analyses, F. Ronco, for help with statistics, and Sebastian Gygli for proof-reading the manuscript. The project was supported by the Singenberg Foundation, the Bristol Foundation, the Federal Office for the Environment, the Parrotia-Foundation, the Werner Steiger Foundation, the Ernst Göhner Foundation, the Stiftung zur internationalen Erhaltung der Pflanzenvielfalt, Schöni Swissfresh AG and the Spendenstiftung Bank Vontobel.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Stefano Colazza.
Rights and permissions
About this article

Cite this article
Juric, I., Salzburger, W., Luka, H. et al. Molecular markers for Diadegma (Hymenoptera: Ichneumonidae) species distinction and their use to study the effects of companion plants on biocontrol of the diamondback moth. BioControl 60, 179–187 (2015). https://doi.org/10.1007/s10526-014-9637-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-014-9637-0