
Abstract
Externalizing behavior is substantially affected by genetic effects, which are moderated by environmental exposures. However, little is known about whether these moderation effects differ depending on individual characteristics, and whether moderation of environmental effects generalizes across different environmental domains. With a large sample (N = 1,441 individuals) of early adolescent twins (ages 11 and 13), using a longitudinal multi-informant design, we tested interaction effects between negative emotionality and both positive and negative aspects of three key social domains: parents, peers, and schools, on the phenotypic variance as well as the etiology of externalizing. Negative emotionality moderated some of the environmental effects on the phenotypic, genetic, and environmental variance in externalizing, with adolescents at both ends of the negative emotionality distribution showing different patterns of sensitivity to the tested environmental influences. This is the first use of gene-environment interaction twin models to test individual differences in environmental sensitivity, offering a new approach to study such effects.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Externalizing behavior (EXT) during adolescence is detrimental both to the self and to others. Childhood and adolescence EXT, characterized by disobedience, irritable mood and verbal or physical aggression, were found to increase the risk for various psychiatric disorders in adulthood (Reef et al. 2011), and for various negative criminal and psychosocial life outcomes (Huesmann et al. 2009). Thus, understanding EXT development could have important personal and societal implications. Individual differences in EXT have been associated both with environmental and genetic effects (Burt 2009; Hannigan et al. 2017). However, the extent of these genetic effects was found to vary, depending on different environmental exposures (Hicks et al. 2009). Based on findings of individual differences in sensitivity to environmental influences on the phenotypic variance in EXT (e.g., Lengua 2008; Pitzer et al. 2011; DiLalla et al. 2015; Janssens et al. 2017a), we propose that the effects of environmental exposures on the etiology of EXT will also differ according to person-based characteristics.
Meta-analyses found substantial genetic and environmental effects on EXT (Burt 2009) and its development (Hannigan et al. 2017). However, genetic and environmental effects do not work independently. Gene-environment interactions (GxE) are one type of joint effects of genes and environments. Typically, two different GxE types are investigated: 1. The moderating effects of genes on the association between an environmental factor and a trait (to be discussed later); 2. The moderating effects of specific environments on the expression of genetic variance in a trait. The latter means that while individual differences in a trait could be influenced by genetic effects, these genetic effects are moderated themselves by environmental exposure. Assessing such GxE effects is usually done with twin designs, enabling us to estimate how different factors moderate genetic and environmental variances in a trait. Indeed, Hicks et al. (2009) found that the expression of genetic variance in EXT increased as a function of environmental adversity in the parental, peer and academic domains. For example, as adolescents had more antisocial peers, the role of their genetic differences became more important in EXT etiology.
GxE findings are important as they demonstrate that the expression of genetic variance is not the same for individuals who experience different environmental exposures. However, such studies do not directly address the individuals’ sensitivity to environmental influences. Theories of individual differences in environmental sensitivity postulate that individuals differ in the degree to which their development is affected by environmental influences (Belsky 2005; Ellis et al. 2011; Pluess 2015). Such theories have gained immense support, showing that associations between environmental effects and developmental outcomes differ according to person-based characteristics (van IJzendoorn and Bakermans-Kranenburg 2015; Slagt et al. 2016).
Typically, studies of individual differences in environmental sensitivity have tested variation in the associations between environmental effects and a trait’s phenotypic variance. We propose that this notion can also be applied to traits’ genetic and environmental variance. That is, moderation effects of environmental influences on genetic variance in a trait may depend on the person’s characteristics. For example, Hicks and colleagues' (2009) finding that the expression of genetic variance in EXT increased as a function of the number of antisocial peer affiliations, could potentially exist only for individuals with reactive temperament but not for others. Such effects can be inferred as individual differences in sensitivity to environmental influences on the etiology of EXT (or any trait). Thus, using twin studies to address individual differences in environmental sensitivity helps to further understand the genetic or environmental mechanism behind moderation effects on a traits’ phenotypic variance.
Environmental effects on the phenotypic variance of EXT were also found to be moderated by person-based characteristics. Many studies have found that candidate genes (mainly dopaminergic and serotoninergic) moderate the effects of parenting (e.g., Janssens et al. 2017a), peers (e.g., DiLalla et al. 2015), and the school (e.g., De Laet et al. 2016) on EXT. These findings are, in fact, a manifestation of the first type of GxE studies mentioned above, where specific genes moderate the association between an environmental effect and a trait. However, as the downsides of the candidate gene approach are increasingly apparent (Duncan and Keller 2011), we focus on a different marker for individual differences in environmental sensitivity – temperament, and specifically, negative emotionality (NE).
Temperament, the combined individual differences in the affect, activity, attention and sensory sensitivity domains, is presumed to have a substantial genetic or neurobiological basis (Rothbart and Bates 2006; Zentner and Bates 2008). These differences appear early in life and are relatively stable across the life span (Rothbart and Bates 2006; Zentner and Bates 2008). NE is the tendency to be easily distressed and experience emotions such as fear, frustration, sadness and anger (Buss and Plomin 1984; Rothbart and Bates 2006). Thus, NE may reflect a highly reactive nervous system, which responds more strongly to environmental stimuli (Ellis et al. 2011; Slagt et al. 2016). Indeed, a meta-analysis concluded that the individuals with higher NE or with a more difficult temperament (a composite of multiple temperamental traits, including NE) are the ones more sensitive to parental influences (Slagt et al. 2016). NE was also found to moderate environmental effects on EXT specifically (e.g., Lengua 2008; Pitzer et al. 2011; Tung et al. 2018). Therefore, we aimed to test whether moderation effects of environmental influences on the expression of genetic and environmental variance in EXT are dependent on NE.
However, it is important to ask which environmental effects should be considered when investigating how and for whom environmental exposure influences EXT development. The study of individual differences in environmental sensitivity has mainly focused on the parental domain as the environmental effect of interest, both in general (Belsky and Pluess 2009; Slagt et al. 2016), and in the study of EXT (e.g., Lengua 2008; Burk et al. 2011; Pitzer et al. 2011; Rioux et al. 2015; Tung et al. 2018). While the parental role is crucial in the early years (Grusec 2011), the peer and school domains become increasingly meaningful during adolescence (Smetana et al. 2006). Indeed, these three environments were all found to have robust phenotypic associations with EXT (Reijntjes et al. 2011; Ttofi et al. 2012; Pinquart 2017), as well as moderation effects on its genetic variance (Dick et al. 2007; Feinberg et al. 2007; Hicks et al. 2009). However, most previous studies have focused on one environmental effect at a time (e.g., Lengua 2008; Pitzer et al. 2011; DiLalla et al. 2015; Janssens et al. 2017a), limiting our comprehensive understanding of environmental effects on EXT development.
Moreover, two main issues arise when examining previous findings in individual differences in environmental sensitivity, that we will review in relation to findings where temperament is the moderator and EXT is the outcome. First, there is evidence challenging the notion that only individuals high on NE are sensitive to environmental influences. For example, Lengua (2008) found that both boys with high, and low, NE showed associations between parenting practices and EXT, but in opposite directions. That is, they both were sensitive to parental influences, but in different ways. Similarly, Essex et al. (2011) found that both children high and low on NE were sensitive to the effects of teacher–child relationship on their internalizing and externalizing symptoms, but to different aspects of the relationship (conflict and closeness, respectively). This raises the possibility that individuals are not either sensitive or not sensitive across all environmental influences and behavioral outcomes (the domain-general perspective), but instead, that different individuals could be sensitive to different environmental influences or regarding different behavioral outcomes (the domain-specific perspective).
Addressing this issue requires examining individual differences in the effects of various environmental domains (e.g., parents and peers) within the same sample. We are aware of only two studies addressing the effects of both the parental and peer domains on EXT, either with a genetic moderator (Janssens et al. 2017b), or with a temperamental moderator (Tung et al. 2018). Both studies found significant effects only for some of the interactions tested, with Janssens et al. (2017b) finding dopaminergic interactions with both parental and peer effects on rule-breaking but not on aggressive behavior, and Tung et al. (2018) finding temperament interactions with negative parenting on EXT, but not with positive parenting, friendship conflict or friendship closeness.
This pattern of findings represents the second issue at hand, where many studies that investigate the moderating effects of temperament on EXT find significant interactions for some environmental aspects but not others (Burk et al. 2011; Rioux et al. 2015; Slagt et al. 2018). Tung et al. (2018) findings are an example across social domains, but there are also such instances within one domain, the parental domain. For example, Lengua (2008) found that NE moderated the effects of maternal rejection/acceptance and physical punishment on EXT, but not of inconsistent discipline. Such findings further emphasize the question whether there are some individuals who are sensitive across many environmental influences.
The investigation of environmental influences is even more complex when considering effects of both positive, and negative, environments. The possible different effects of positive versus negative aspects of an environment are represented in three models under the meta-framework of individual differences in environmental sensitivity (Pluess 2015): 1) the diathesis-stress model, in which certain individuals are more sensitive to negative environmental effects; 2) the differential susceptibility model, in which certain individuals are more sensitive to both negative and positive environmental effects; 3) the vantage sensitivity model, where certain individuals are more sensitive to positive environmental effects.
Combining these two issues highlights our insufficient understanding of which individuals may develop EXT in response to which environmental effects. Therefore, our second aim was to include representations for multiple key social domains during adolescence when investigating individual differences in sensitivity to environmental influences on EXT, on its phenotypic, genetic, and environmental variance.
The Present Study
We aimed to gain a broad understanding of individual differences in sensitivity to environmental influences on developing EXT during adolescence, by adding to the current literature in two important ways. First, we wanted to test individual differences in sensitivity to environmental influences on EXT both in the typical way, that is, on the phenotypic variance in EXT, and in a new way, using a twin design, on the etiology of EXT, that is, on the genetic and environmental variance in EXT. To do so, we expanded the currently used GxE twin model, which tests the moderating effect of one factor on the genetic and environmental variance in a trait, to estimate the interactive effects of two moderators – environmental and personal. This can teach us if the genetic or environmental contribution to the variance in a trait not only differs depending on environmental exposures (such as parenting), but rather if this dependency differs across individuals based on their personal characteristics (specifically, NE) as well.
Second, we aimed to gain a better understanding of whether the same individuals are influenced by different domains, or rather different individuals are influenced by different domains. Therefore, we examined the effects of three prominent social domains during adolescence: parents, peers, and school, and examined both their positive (e.g., parental support) and negative (e.g., conflict with parents) aspects, as EXT might be influenced both by a highly negative environment and by the lack of an important positive environment. We continued by testing which individuals demonstrated sensitivity, and whether the interaction is in the form of diathesis-stress, differential susceptibility, or vantage sensitivity.
We used a multi-informant longitudinal design with a twin early adolescent sample. First, we tested interaction effects of NE with positive and negative aspects of the parent, peer, and school environment, on the observed variance of EXT. To address the role of NE, environment, and their interaction on developmental change, we predicted EXT at age 13 by environmental measures and NE from age 11, adjusting for EXT levels at age 11. Second, we used the extended version of Purcell's (2002) GxE twin model, to examine how environmental and temperamental moderators interact to affect the genetic and environmental variation in EXT.
We expected to find interaction effects between the environment and NE on EXT development, such that different individuals (depending on their NE) would be differentially influenced by environmental effects. However, based on the findings reviewed above (Lengua 2008; Essex et al. 2011), we considered the possibility that different adolescents are sensitive to different environmental effects, or show associations in opposite directions.
Method
Participants
Families participated as part of the Longitudinal Israeli Study of Twins, a large twin study on genetic and environmental influences on social development (Vertsberger et al. 2019). Parents of twins born in Israel during 2004–2005 from Hebrew-speaking families were invited to participate. The current data were collected from mothers and their twins when the twins were 11 (Wave 1) and 13 (Wave 2). For the purposes of this study, families that had available data on EXT at age 11 or at age 13 were included, creating a total of 726 families. Mother-report data were disqualified for seven families at age 11 and one family at age 13 (disqualified from the age 11 sample as well), mainly due to mix-ups whilst filling ids. In addition, for twenty-eight adolescents, the self-report data at age 11 was disqualified mainly due to id mix-ups or adolescents appearing to have received help from a family member in reading and responding to the survey. The final sample included 1,441 individual twins from 721 families (312 families participated in both waves): 298 monozygotic (MZ) twins (from 149 families, 21%), 603 dizygotic (DZ) same-sex twins (from 302 families, 42%), 520 dizygotic opposite-sex twins (from 260 families, 36%) and twenty twins which did not have available zygosity data. Missingness analyses were conducted on our key variables and on demographic characteristics, and significant differences were found only regarding the distribution of religious and non-religious mothers between families who participated at both ages and families who participated only at age 13 (see supplementary materials, Table S1).
Participating adolescents were 51% females, aged 10.68–12.27 at Wave 1 (M = 11.16, SD = 0.22) and 12.53–14.56 at Wave 2 (M = 13.37, SD = 0.25). Mothers were 31–65 years old (M = 43.11, SD = 5.33) at Wave 1 and 33–63 at Wave 2 (M = 45.01, SD = 5.30). Sample demographics were similar to those of the general population (Vertsberger et al. 2019). Details regarding participating mothers’ education and income level can be found in supplementary Note 1.
Procedure
In both study waves mothers and twins completed a series of questionnaires as part of a larger study, from their homes, in their free time, usually in more than one sitting. In the initial phone recruiting call, mothers were told that each twin should answer the survey independently, with no interruptions. Most participants completed the questionnaires online on their home computers, whereas families who did not have a home computer or an internet connection, or preferred a paper-and-pencil version for other reasons, received printed questionnaires via postal service, which were returned after completion (22% and 13% from Wave 1 and 2, respectively). As some significant differences were found between online and paper-and–pencil participants (see supplementary Table S2), participation method was inserted as a control variable. Families were offered 3–4 movie or museum tickets in return for their participation. This study was approved by the ethics committee of the Hebrew University of Jerusalem. Parents provided informed consent for their children to participate, and the adolescents provided assent.
Measures
Externalizing Behavior
Mothers reported on children’s EXT at both waves, using a composite scale, including five items from the Conduct Problems subscale of the Strengths and Difficulties Questionnaire (SDQ, e.g., “Often fights with other children or bullies them”; Goodman 1997), and seven items adapted from the Peer Aggression and Aggression/Defiance subscales of the Infant–Toddler Social and Emotional Assessment (Carter et al. 2003) (e.g., “Hurts other children on purpose”, “Acts aggressive when frustrated”). The full item list appears in supplementary Table S3. Items were rated on a 3-point Likert scale (0 = not true, 2 = definitely true) and averaged to create a total score (αWave1 = 0.82; αWave2 = 0.79). Exploratory factor analysis (EFA) using the principal factor solution indicated a one-factor solution, according to both the scree plot and Kaiser’s criterion (eigenvalue greater than 1), accounting for 31% and 27% of the variance in age 11 and 13, respectively.
For scales with no existing Hebrew version (SDQ being an exception), English items were adapted to Hebrew using a translation-back-translation procedure (Brislin 1980).
Negative Emotionality
Mothers reported on their children’s NE at Wave 1, with the five-item Emotionality subscale (e.g., “Gets upset easily”) of the EAS questionnaire (Buss and Plomin 1984), using a 5-point Likert scale (1 = not characteristic or typical of your child, 5 = very characteristic or typical of your child). The items were averaged to create a total score (α = 0.80).
Parental Environment
Parental support and conflict with parents were measured as indicators of positive and negative parental characteristics, respectively.
Parental Support
Adolescents reported their feelings of being loved and regarded by their parent using the Regard for Child subscale (e.g., “I know my parent loves me”) of the Parent Environment Questionnaire (PEQ; Elkins et al. 1997). This subscale comprises ten items (five about mothers and five about fathers, r = 0.59), measured on a 4-point Likert scale (1 = definitely not true, 4 = definitely true), and averaged into one score (α = 0.81).
Conflict with Parents
Adolescents reported on their negative feelings and interactions with their parents using eight items of the Conflict With Parents subscale (e.g., “Me and my parent often argue”) of the PEQ (Elkins et al. 1997) measured on a 4-point Likert scale (1 = definitely not true, 4 = definitely true). Adolescents reported separately on conflict with their mothers and with their fathers (r = 0.66), and scores were averaged into one score (α = 0.89).
Peer Environment
Friend support and peer problems were measured as indicators of positive and negative peer characteristics, respectively.
Friend Support
Adolescents reported the extent to which they felt supported by their friends with the 4-item Friends subscale (e.g., “I can talk about problems with my friends”) of the Multidimensional Scale of Perceived Social Support (Zimet et al. 1988), using a 7-point Likert Scale (1 = very strongly disagree, 7 = very strongly agree) (α = 0.81).
Peer Problems
Mothers reported on their children’s problems with friendships and peers using the Peer Problems subscale of the SDQ (e.g., “Picked on or bullied by other children”; Goodman 1997). Items were averaged to create a total score (α = 0.69).
School Environment
Parents provided the name of children’s elementary school, which was used to retrieve data on schools’ characteristics, based on reports published by the Ministry of Education. Every year, the National Authority for Measurement and Evaluation in Education (RAMA) administrates surveys for teachers and students on the school’s social-emotional climate as part of the national standardized tests in select subjects (Hebrew, English, and Math). The test results and school climate surveys are published online (Ministry of Education n.d.). We used the measures of positive school climate as representing positive school characteristics and school bullying as representing negative school characteristics.
The tests are administrated to three age groups and the age group relevant to our analysis is the 5th and 6th grade combined estimation. Every school is assessed once every three years. Therefore, we retrieved relevant data from the years 2015, 2016 and 2017 (Nadolescents = 245, 304 and 318, respectively). School measures were available for 867 (60%) of all adolescents, and 494 (66%) of the adolescents who participated at age 13. School measures were missing if we did not have the school’s name, or if there were no available reports on the school. In most families the twins attended the same school, but for 9% of families with available school data the twins attended different schools (mainly gender-separated schools).
Positive School Climate
The score of general positive feeling about the school comprised three items: “I like being in school”, “Even if I could have, I wouldn’t move to another school” and “I feel good at school”. Students answered these items on a 5-point Likert scale, and results are the percentage of students surveyed in the school that chose the top two agreement scale values (4 or 5). The average of the items’ percentages is published by RAMA and used here.
School Bullying
The school’s bullying measure comprised three total scores (computed by RAMA the same as described above): 1) Students’ lack of feeling protected (based on three items, e.g., “Sometimes I’m scared to go to school, because it has students who act violently”); 2) Engagement in violent incidents (based on nine items assessing general, extreme, or indirect bullying “In the last month”, e.g., “a student pushed me”; “a student bullied me into giving them money, food or other valuables”; “a student tried to convince other students not to speak with me or be my friends”); 3) Bullying in social media (based on four items, e.g., “In the last month someone started rumors about me on social media (e.g., WhatsApp, Facebook etc.,) with the aim to hurt me”). We averaged these three total scores into one school bullying measure (α = 0.68).
Zygosity
Zygosity for same-sex twins was mainly assessed using DNA data (available for 51% of same-sex pairs). Most of the remaining sample’s zygosity was assessed using a parent questionnaire regarding physical similarity (see Vertsberger et al. 2019), which has shown 95% agreement with DNA measures (Price et al. 2000).
Data Analysis
Statistical analyses were conducted using RStudio (RStudio Team 2020) based on R 4.0.3 (R Core Team 2020). Pairwise deletion was used with correlations and listwise deletion was used with generalized linear mixed-effects modeling (GLMM). Packages used for the phenotypic moderation effects included the glmer function from the lme4 package (Bates et al. 2015) for conducting GLMM, and interactions (Long 2019) for probing and plotting interactions. The genetic analyses used the umx package (Bates et al. 2019) for univariate analyses and OpenMx 2.18.1 (Boker et al. 2011; Neale et al. 2016) for the moderation analyses.
Moderation Effects on the Phenotypic Variance in Externalizing Behavior
GLMM was used due to the dependency between twins from the same family and skewness of the dependent variable (i.e., EXT). To deal with the skewness of EXT, GLMM was fitted with an inverse gaussian distribution and the default link function \(\frac{1}{{\mu }^{2}}\). These distribution and link functions were chosen based on model fitting, according to recommendations by Lo & Andrews (2015) (see supplementary Table S4). As this distribution takes only positive values, we added 1 to the mean EXT scores, transforming their range from 0–2 to 1–3.
To test the moderation effects of NE on the associations between environmental effects at age 11 with EXT at age 13, we conducted a four-step hierarchical analysis. First, we tested the null model, where the only factor entered was the random effect of family (i.e., the nesting of twins within families). Second, we tested the change in EXT, by predicting age 13 EXT from age 11 EXT, while adjusting for sex (1: female; 2: male) and method of participation at age 11 (1: paper; 2: website). Third, we entered our moderator (NE) and six predictors (3 environmental domains x positive/negative aspects), which were all standardized to minimize scale differences. Fourth, we added the interaction terms between the environmental measures and NE.
Post-hoc analyses using the Johnson-Newman (J-N) technique (Preacher et al. 2006) were done on significant interactions to identify the values of NE for which the slope of the environmental predictor was significant. That is, which adolescents show an association between the environmental predictor and EXT. We also probed interactions to examine which pattern of environmental sensitivity they demonstrate: diathesis-stress, differential susceptibility, or vantage sensitivity. This was done by calculating the region of significance (RoS; using an online calculator, Preacher et al. n.d.) with respect to the environmental predictor (X) (Roisman et al. 2012). With the use of RoS, it is possible to identify the range of predictor values (specifically the ends, − 2 SD/ + 2 SD) for which simple slopes of high and low NE values differ from each other significantly. Diathesis-stress is supported if the simple slopes differ only in a “for worse” manner, that is, only on the negative side of the environmental scale. Vantage sensitivity is supported if the simple slopes differ only in a “for better” manner, on the positive side of the environmental scale. Differential susceptibility is supported if the simple slopes differ “for better and for worse” on both sides of the environmental scale.
Moderation Effects on the Genetic and Environmental Variance in Externalizing Behavior
Genetic analyses were conducted on EXT at age 11, as this sample was much larger than the age 13 sample, and twin analyses require large samples for sufficient power (Neale et al. 1994). Because variables included in the analyses had different variance magnitudes, we rescaled some of the variables to have the same order of magnitude throughout. The new descriptive statistics of the rescaled variables appear in supplementary Table S5. In both the univariate and multivariate moderation analyses we adjusted for the main effect of sex by inserting it as a covariate, and thus DZ same-sex and DZ other-sex twins were included as one group.
Twin designs generally compare similarity between MZ and DZ twins. The phenotypic variation in a trait is then typically decomposed onto three factors: genetic factors (A), shared environment factors (C) and nonshared environment factors (E). Details on the basic twin design and estimation of the ACE components can be found in supplementary Note 2. For the univariate analyses, we first tested the full ACE model and then dropped nonsignificant path coefficients (where the 95% confidence interval included zero) and compared the two models. The more parsimonious model was chosen if model fit did not worsen after modifying the model.
The moderation effects on the genetic and environmental variance in EXT were fit to the raw data using full-information maximum likelihood estimation, and modeled using an extended version of Purcell's (2002) GxE twin model. A detailed explanation of the basic, bivariate, form of Purcell’s (2002) model appears in supplementary Note 3. The extended model allows incorporating more than one moderator, along with the interaction between the moderators. Using such modeling enables us not only to estimate the moderating effects of either NE or environmental effects on the expression of genetic and environmental variance in EXT, but importantly, also to estimate the moderation effect of their interaction.
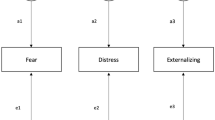
We conducted six separate analyses, where NE and one of the six environmental variables were treated as moderators. The general scheme for our moderation models is presented in Fig. 1, where M represents the currently analyzed environmental variable. Path coefficients with a dot (∙) in their subscripts are conditioned upon the moderators. For example,

Path diagram for the genetic moderation models. Diagram depicts only one twin’s part of the model, and only the A and E components, as the univariate analysis of externalizing behavior found no C component; the latent A variables would covary with the co-twin’s latent A variables according to zygosity. A refers to sources of additive-genetic variance, and E refers to sources of nonshared-environmental variance; some environmental variables had shared-environmental variance in addition to or instead of additive-genetic variance (not depicted in figure). All latent variables have a variance of 1.0 (not depicted), and all manifest variables are adjusted for the main effects of sex (not depicted). M environmental variable, NE Negative emotionality, EXT Externalizing at age 11. As explained in the main text, path coefficients with a dot (∙) in their subscripts are conditioned upon the putative moderators
That is, in the full model with both moderators and their interaction, the effect of the first genetic factor (\({A}_{1}\)) on the third variable (EXT) is estimated as the sum of its unmoderated effect \({a}_{130}\), its effect moderated by M (\({a}_{131}\)), its effect moderated by NE (\({a}_{132}\)), and finally, its effect moderated by the interaction between M and NE (\({a}_{133}\)). Similarly, the effects of the second and third genetic factors can be described as follows:
Therefore, \({a}_{13\cdot }^{2}\) represents moderated additive-genetic variance common to EXT and the environmental variable, \({a}_{23\cdot }^{2}\) represents moderated additive-genetic variance common to NE and EXT, and \({a}_{33\cdot }^{2}\) represents moderated additive-genetic variance unique to EXT. As the result of interest is the total genetic variance in EXT, we calculated the value of \(a_{{13\cdot }}^{2} \; + \;a_{{23\cdot }}^{2} \; + \;a_{{33\cdot }}^{2}\), as a function of different M and NE values. Substitution of e for a in the preceding symbols yields the analogous case for the nonshared-environmental variance components.
In the analysis of each environmental variable, we fitted 3 models. In Model 0, a “no moderation” model, all moderation coefficients (all coefficients with subscripts ending in “1”, “2”, or “3”) were fixed to zero. In Model 1, a “two moderators” model, the “first-order” moderation coefficients, that is the moderation effects of the environmental variable, M, and negative emotionality, NE (i.e., coefficients with subscripts ending in “1” or “2”, but not those ending in “3”) were freely estimated. In Model 2, the “full moderation” model, all moderation coefficients were freely estimated, adding the moderation effects of the interaction between the environmental variable and NE. In each analysis, we selected from these three models the one that had the smallest AICc (sample-size corrected Akaike’s Information Criterion).
Results
Descriptive statistics and correlations for all key measures appear in Table 1. First, EXT at age 11 and EXT at age 13 are highly correlated, indicating stability. EXT at both ages demonstrated a similar pattern of associations with our age 11 predictors: positive associations were found with NE and with the negative aspects of all three environmental domains—conflict with parents, peer problems and school bullying (the latter was significant only with EXT at age 11). In contrast, a significant negative association was found only with parental support and friend support at age 11, but not with the school’s positive aspects. This demonstrates that different aspects of adolescents’ environment at age 11, i.e., when our predictors were measured, are associated with the levels of EXT they show at age 13.
Within each environmental domain, the positive and negative aspects exhibited negative associations with each other, that is, the more parental support adolescents received, the less conflict they experienced with their parents; the more friend support adolescents received, the less peer problems their mothers reported that they have; and the more students at the school reported a positive school climate, the less they reported on bullying incidents in the school. Within the positive environmental aspects, parent and friend support were positively associated, but there were no associations with positive school climate. There were no associations within the negative environmental aspects. Finally, NE was positively associated with the parental and peer negative environmental aspects, and negatively associated with parental support.
We found two significant sex differences using independent t-tests with a Bonferroni adjusted alpha level of 0.007 (0.05/7). Boys were higher than girls on EXT at both ages (t11(1262.6) = 4.85, p11 < 0.001, d11 = 0.27; t13 (700.17) = 3.23, p13 = 0.001, d13 = 0.24; Mgirls = 1.27, 1.23; SDgirls = 0.27, 0.24; Mboys = 1.35, 1.29; SDboys = 0.33, 0.29; for age 11 and 13, respectively), while girls scored higher than boys on parental support (t(1006.4) = 4.42, p < 0.001, d = 0.27; Mgirls = 3.77, SDgirls = 0.31; Mboys = 3.68, SDboys = 0.38). Analyses were therefore adjusted for sex.
Moderation Effects on the Phenotypic Variance in Externalizing Behavior
Table 2 presents the GLMM results testing the environmental and interaction effects on EXT at age 13. Environmental predictors had no main effects on EXT at age 13 after adjusting for sex and EXT at age 11. NE had a significant effect on EXT only in the full model, where the interaction effects were included. Several significant interactions between NE and environmental variables emerged above and beyond the continuity of EXT. Specifically, parental support and peer problems predicted EXT development from age 11 to age 13, depending on adolescent’s NE during the environmental exposure. In addition, the interaction between NE and positive school climate was marginally significant, but we proceeded to examine it to see if we can identify individuals who are more sensitive than others to environmental influences, as this was our main interest. Comparing the main effects model to the interaction effects model revealed that the interactions’ model had a better model fit (χ2diff(6) = 26.78, p < 0.001).
Individual Differences in Sensitivity to Parent Effects
Parental support, but not conflict with parents, showed a significant interaction: high-NE adolescents showed less EXT when receiving more parental support, but low-NE adolescents showed more EXT when receiving more parental support (Fig. 2A.1). This pattern was supported by the J-N intervals, where the slope of parental support is significant for both high and low NE values (outside of -1.58 SD and 0.35 SD; Fig. 2A.2). The interaction between parental support and NE is compatible with the diathesis-stress pattern (RoS on X outside of 0.05 SD and 2.42 SD). That is, the difference between adolescents high and low on NE is expressed when they receive less parental support.

Significant interactions between environmental effects and negative emotionality on externalizing behavior. On the left the interaction plot is presented, and on the right the Johnson-Neyman intervals are presented, for significance of predictor slopes
Individual Differences in Sensitivity to Peer Effects
In contrast to the parent effects, a significant interaction was found between NE and the negative, but not positive, aspect of peer effects (Table 2). While high-NE adolescents showed more EXT if they had more peer problems, low-NE adolescents showed an opposite association– they had less EXT if they had more peer problems (Fig. 2B.1). This pattern was supported by the J-N intervals, with the peer problems slope being significant for both high and low NE values (outside of the NE levels of − 1.19 SD and 0.98 SD, Fig. 2B.2). Probing the peer problems-NE interaction revealed that high-NE and low-NE adolescents differed significantly in the high end of the peer problems scale (RoS on X outside of − 3.00 SD and − 0.05 SD), a pattern that is compatible with diathesis-stress.
Individual Differences in Sensitivity to School Effects
School effects demonstrated a marginally significant interaction with NE for the positive, and not negative, aspect of the school – positive school climate (Table 2). Nevertheless, we chose to probe the interaction to understand whether there was a significant association between positive school climate and EXT for some adolescents. Interestingly, low-NE adolescents that attended schools with a higher positive climate exhibited more EXT (Fig. 2C.1). This is supported by the J-N intervals (Fig. 2C.2), where the slope of positive school climate is significant only for low NE values (inside of -3.82 SD and -1.77 SD). RoS analysis found that the slope for low-NE adolescents differs in the lower end of the scale (RoS on X inside of -4.31 SD and 0.06 SD), indicating that the effect on EXT comes from low positive school climate, i.e., compatible with a diathesis-stress pattern.
Alternative Analytic Approaches
As our predictors are reported by different sources, missing values substantially reduced sample size for the full model. Therefore, we also examined separate models for each environmental domain (parents, peers, school). The conclusions remained largely the same, with the exception that the parenting main effects were now significant, and the positive school climate-NE interaction now outside of the marginally significant range (p = 0.126; see supplementary Table S6). In addition, as interpreting between-subject effects is complicated when using GLMM (Fitzmaurice et al. 2004), we also analyzed the phenotypic moderation effects using Generalized Estimating Equations (GEE). The results remained largely the same as in our main analysis, with the main effect of conflict with parents becoming significant, the peer problems-NE interaction becoming marginally significant (p = 0.082), and the school-climate-NE interaction non-significant (p = 0.135; see supplementary Table S7).
Summary of Moderation Effects on the Phenotypic Variance
High-NE adolescents showed sensitivity to aspects of both the parental and peer environment, in the expected direction: they demonstrated more EXT when exposed to non-supporting parents and when having more peer problems. Low-NE adolescents, on the other hand, showed sensitivity to aspects of the school in addition to aspects of the parent and peer environments, but in the opposite, unexpected, direction: they demonstrated less EXT when exposed to non-supporting parents, when having more peer problems and when exposed to a less positive school climate.
Moderation Effects on the Genetic and Environmental Variance in Externalizing Behavior
Table 3 presents the MZ and DZ twin correlations, as well as the univariate model’s ACE components. Twin correlations and the univariate model were not calculated for school variables, as they are not measured on a person-, or even familial, level, but rather by an entire class of students. Table 3 shows that EXT is highly heritable (66–72%) with the rest of the variance explained by nonshared environmental effects (28–34%). NE was highly heritable as well (72%), as can be expected from temperament (Zentner and Bates 2008). The parental and peer problems variables demonstrated moderate to high heritability (45–65%). The rest of the variance in the parental variables was explained by nonshared environment (38–55%), whereas peer problems showed a shared environment effect (16%) as well as a nonshared environment effect (19%). Finally, friend support was the only variable that showed no genetic effect, but showed only environmental effects, both shared (22%) and nonshared (78%). As EXT, NE and the parental variables showed only genetic and nonshared environment effects, these were the components that were estimated in the corresponding moderation models (i.e., AE models). In contrast, as friend support showed no genetic effect, and as school variables are school-level measures, only the shared and nonshared environment effects were estimated in these moderation models (i.e., CE models).
When attempting to fit the genetic moderation models (Models 1 and 2) with peer problems and positive school climate as the environmental moderators, we encountered seemingly intractable numerical optimization difficulties, such that we could not obtain a solution that satisfied OpenMx’s convergence diagnostics. Despite our best efforts a credible solution could not be found. Detailed explanation and strategies employed are provided in supplementary Note 4. Therefore, for peer problems and positive school climate we only report the results of Model 0, i.e., the model with no moderation effects. The results can be found as path diagrams in supplementary Figures S2 and S3. Peer problems had genetic correlations with both NE (rg = 0.213) and EXT (rg = 0.306) and a nonshared-environmental correlation with NE (re = 0.131), but not with EXT (re = − 0.030). As for positive school climate, as genetic effects were not estimated for this variable, its genetic correlations were not estimated, and it did not show nonshared-environmental correlations with NE (re = 0.067) or EXT (re = − 0.009). That is, there are common genetic, but not environmental, factors that cause adolescents to be higher both on EXT and on peer problems, and no common etiology for EXT and school climate.
Table 4 presents the fit indices for the moderation models of NE and the rest of the environmental measures on EXT etiology. For all environmental measures tested, a model with moderated effects had a better fit to the data than a model without any moderation effects (Model 0). Thus, the expression of genetic and environmental variance in EXT varies across different levels of NE and across different levels of environmental exposure. Furthermore, for the parental domain and school bullying the genetic and environmental variances are also moderated by the interactive effects of NE and environment. That is, the moderating effects of parents and school on EXT etiology are dependent on adolescents’ NE levels. Results of the best fitting models are presented in Fig. 3, which shows variations in the unstandardized variance estimates as a function of NE and the environmental predictor. In addition, the results will be described individually, separately for the predictors where the two moderators model had the best fit, and for the predictors where the full model, with interaction effects, had the best fit.

Moderation analyses for the genetic and environmental variance in externalizing behavior. Changes in the unstandardized variance of externalizing are presented as a function of rescaled values of environmental exposure and negative emotionality for the best-fitting model
Independent Moderation Effects for the Environment and NE
Only friend support showed moderation effects for the two predictors, but not for the interaction between them.
Friend Support
The expression of genetic variance in EXT slightly increased as NE increased, but the more pronounced difference seems to depend on friend support, where greater friend support increased genetic variance in EXT. That is, as adolescents are higher on NE and as they are exposed to more friend support, genetic differences between them become more important in EXT etiology. The expression of nonshared environment variance in EXT increased as NE increased across all values of friend support, but friend support had no effect on the expression of environmental variance. That is, as adolescents are higher on NE, the environmental differences between them become more important in EXT etiology.
Interactive Moderation Effects of the Environment and NE
Parental support, parental conflict and school bullying showed both separate and interactive moderation effects of the environment and NE on the expression of genetic and environmental variance in EXT.
Parental Support
The expression of genetic variance in EXT is the greatest on the ends of both parental support and NE: it is high both for high and low levels of parental support, and high for high and low levels of NE, but not medium levels of both predictors. That is, genetic differences between adolescents are important in EXT etiology for high-NE adolescents who experience a lot of parental support, and even more important for adolescents who are either low or high on NE, that experience low parental support. The nonshared environment variance in EXT, however, is substantial only for high-NE adolescents that are also exposed to low parental support. This indicates that environmental differences are important in EXT etiology for high-NE adolescents who experience low parental support.
Conflict with Parents
The expression of genetic variance in EXT increased for high-NE adolescents as conflict with parents decreased, and for low-NE adolescents as conflict with parents increased. Genetic differences were the most important in EXT etiology when low-NE adolescents had high levels of conflict with their parents. The EXT nonshared environment variance was high only for high-NE adolescents who also experience high levels of conflict with parents. That is, environmental differences between adolescents are important in EXT etiology when high-NE adolescents experience a lot of conflict with their parents.
School Bullying
The expression of genetic variance in EXT was high for high-NE adolescents both when school bullying was low and high. The genetic variance in EXT was even greater for low-NE adolescents in schools high on bullying. That is, when school bullying was low, genetic differences between adolescents were important in EXT etiology for high-NE adolescents, but when school bullying was high, genetic differences were important in EXT etiology for high-NE adolescents, and even more for low-NE adolescents. The expression of nonshared environment variance in EXT increased as school bullying increased, both for low-, and high-NE adolescents, however the environmental variance in EXT was greater for high-NE adolescents throughout. That is, environmental differences between adolescents are important in EXT etiology mostly for high-NE adolescents and mainly in schools with a lot of bullying.
Summary of Moderation Effects on the Genetic and Environmental Variance
Environmental differences were important in EXT etiology for high-NE adolescents in all tested models, and only showed some importance for low-NE adolescents regarding the school bullying effect. Genetic differences between adolescents were important in EXT etiology as NE and as the environmental exposure levels increased in the friend support model. However, in the parental domain and school bullying, the effect of the environment on the genetic variance in EXT depended on NE levels. Genetic differences between adolescents were more important in EXT etiology when adolescents didn’t experience parental support and attended high bullying schools, for both high-, and low-, NE adolescents. However, genetic differences were more important in EXT etiology when conflict with parents was low for high-NE adolescents, but when conflict with parents was high for low-NE adolescents.
Discussion
EXT during adolescence could have long term negative consequences to the self and to others (Huesmann et al. 2009; Reef et al. 2011), therefore, it is important to understand what influences its development. Even though environmental effects were found both on the phenotypic and genetic variance of EXT, individual differences in sensitivity to such effects were studied only on the phenotypic level. Furthermore, effects of environmental aspects on EXT are usually studied individually, thus limiting our ability to understand which individuals are sensitive to which environmental influences. We found that both adolescents with high, and with low, levels of NE demonstrated sensitivity to environmental influences on EXT, however, they presented different patterns of sensitivity. This was found both with the phenotypic and the genetic and environmental variance in EXT.
Moderation Effects on the Phenotypic Variance of Externalizing Behavior
To understand how environmental effects influence EXT development, we studied three key social domains—adolescents’ parent, peer, and school environments. For each domain, we tested both positive and negative aspects. Furthermore, as theory and empirical findings suggest that environmental effects do not influence everyone to the same degree (Pluess 2015; Slagt et al. 2016), we tested how temperament moderates the associations between environment and EXT. Specifically, we measured how the environment and NE at age 11 affect EXT at age 13, adjusting for sex effects and initial levels of EXT at age 11.
Whereas we did not find main effects, we did find some interaction effects of these environments and NE on EXT development from age 11 to age 13. Specifically, we found a significant interaction between parental support and NE, such that low levels of parental support were associated with more EXT in high-NE adolescents, and less EXT in low-NE adolescents. We also found a significant interaction between peer problems and NE, such that peer problems related to more EXT in high-NE adolescents but to less EXT in low-NE adolescents. Finally, we found a marginally significant interaction between positive school climate and NE, that revealed a positive association between positive school climate and EXT for low-NE adolescents.
While various studies have demonstrated individual differences in sensitivity to environmental influences on EXT (Lengua 2008; Pitzer et al. 2011; DiLalla et al. 2015; Rioux et al. 2015; De Laet et al. 2016; Janssens et al. 2017a; Tung et al. 2018), these studies focused on a narrow measurement of the environment, not addressing the issue of whether sensitive individuals are generally more sensitive to any environment, or rather are more sensitive to the effects of that particular environmental aspect or domain and might not be sensitive to others. This question was raised more generally in theoretical work regarding individual differences in environmental sensitivity and is questioning whether such differences are domain-general (individuals are either sensitive to many environmental influences or not/much less sensitive), or domain-specific (different individuals are sensitive to different environmental effects) (Belsky 2005; Belsky and Pluess 2009, 2013; Ellis et al. 2011; Mitchell et al. 2013; Belsky and Hartman 2014; Hartman and Belsky 2015). While theoretical and evolutionary thinking favors the domain-specific perspective, suggesting that the effects of different environmental inputs could have either costs or benefits depending on the context, thus creating individual differences in sensitivity to each of these different inputs (Belsky 2005; Markovitch and Knafo-Noam 2021), there is almost no empirical work testing this question.
However, some direct and indirect evidence for domain-specificity is emerging. Two recent studies directly tested, and supported, the notion of domain-specificity of individual differences in environmental sensitivity. That is, different individuals were found to be sensitive to different environments (Belsky et al. 2021; Markovitch and Knafo-Noam 2021). In addition, two studies tested individual differences in the effects of two environmental domains on EXT (within the same sample), thus indirectly testing if the same individuals are sensitive to different environments (Janssens et al. 2017b; Tung et al. 2018). Similar to our findings, both studies found that only some of the moderating effects they tested were significant, indicating that the “sensitive” individuals might not be more sensitive to every aspect of the environment.
Investigating the interactions to identify which adolescents are more influenced by their environment yielded interesting results. As could be expected, high-NE adolescents (the typically “sensitive” individuals) showed significant associations between EXT and both parental support and peer problems. These associations were in the expected direction, with more EXT associated with less parental support and more peer problems. In contrast, low-NE adolescents (typically “not sensitive”) also showed sensitivity to their environment, with significant associations between EXT and positive school climate, as well as with parental support and peer problems. These associations were not in an expected direction, with more EXT associated with less peer problems, more parental support and greater positive school climate.
A closer look on which adolescents demonstrated environmental sensitivity in each end of the NE scale revealed substantial differences: whereas the high-NE adolescents who were sensitive to their parental and peer environments had a NE score of above 0.35 SD and 0.98 SD, respectively, low-NE adolescents who were sensitive to their environment had a score below − 1.19, − 1.58 and − 1.77 SD for the peer, parent, and school environments, respectively. As the minimum level for high-NE is close to the mean while the maximum level of low-NE is more extreme, this means that the sensitive high-NE group includes significantly more adolescents than the sensitive low-NE group. Indeed, 19–40% of adolescents were in the high-NE group (depending on the cutoff value), and 3–13% of adolescents were in the low-NE group. This group size difference might be one of the reasons that high-NE individuals are often found as sensitive to environmental influences and low-NE are not.
Even though the sensitive low-NE group might be smaller and thus under-represented in studies, careful examination of previous findings reveals a similar pattern of results. In some cases, when probing interactions between environmental factors and temperament on development, the group which is typically “not-sensitive” also showed a significant association between the environmental exposure and the outcome (EXT, as well as internalizing problems, social competence and affect dysregulation) (Lengua 2008; Bush et al. 2010; Du Rocher Schudlich et al. 2011; Essex et al. 2011; Tung et al. 2018). As this pattern of findings is usually not part of the hypotheses, it is generally ignored or disregarded. However, as such findings add up, their accumulation suggests that it might present a real, under-researched, pattern that needs to receive theoretical and empirical attention moving forward.
In addition, we carefully suggest that further consideration should be given to the associations found in unexpected directions for low-NE adolescents. As such associations make little theoretical sense, these results might be spurious. However, this is not the first time that such results were found. Lengua (2008) found less EXT and internalizing problems in low-NE children exposed to more negative parenting practices. Such unexpected associations were also found for high-NE children. Both children high and low on NE demonstrated associations in the expected directions as well, depending on the parenting practice, NE dimension and outcome. These findings were discussed individually according to the constructs involved, but we raise the possibility that Lengua’s and our findings might point to the existence of individuals who are sensitive to environmental influences in untrivial ways. Although further evidence is needed before concluding that there is such a pattern of sensitivity, we suggest, with caution, two possible explanations for this pattern. First, because low-NE adolescents are less expressive with their emotions, negative environments might strengthen these inward tendencies, thus decreasing EXT, whereas positive environments suppress the inward tendencies, enabling adolescents to be more outward toward the world, expressed, among other ways, in EXT. Second, the same environmental input might be perceived and processed differently for individuals low and high on NE, enabling opposite developmental outcomes. Future work should address the existence of such a pattern and its underlying mechanism.
More generally, our findings demonstrate that, at least in the current context, there aren’t individuals who are sensitive to their environment and others who are not (domain-generality), but rather, that some individuals are sensitive to some aspects of the environment or in some ways, and others who are sensitive to other aspects or in other ways (domain-specificity). Our results thus add to the knowledge of two different research fields. First, they demonstrate the independent roles of each of the parent, peer, and school environments in EXT development, but only for some adolescents and not others. Such results emphasize the importance of a comprehensive investigation of environments to understand EXT development, as well as the importance of individual differences in such effects. Second, our results also contribute to the field of individual differences in environmental sensitivity in general, providing support for patterns of domain-specificity. Human development depends on environmental inputs, but the study of such inputs is complex. The effects of different environments are interrelated and often are nested within each other (Bronfenbrenner and Morris 2006). None of them can be ignored, and most interestingly, our findings might suggest that they have rather different effects on the developing person, depending on the individual’s characteristics.
Moderation Effects on the Genetic and Environmental Variance in Externalizing Behavior
Studies of individual differences in environmental sensitivity were done until now only on the phenotypic variance of traits. This is the first study to examine individual differences in environmental sensitivity using GxE twin models. This approach allows us to test individual differences not only on the observed variance of a trait, but also on its genetic and environmental variance, thus directly addressing the etiology of these individual differences. As our findings showed such interaction effects, this method could be considered as a new approach for testing individual differences in environmental sensitivity.
Specifically, we found that regarding the nonshared environment variance, variation in environmental exposure was more important in EXT etiology as NE increased. Such findings are in accordance with theoretical and empirical claims of higher environmental sensitivity in high-NE individuals (Slagt et al. 2016). These results might also be interpreted through the different models of individual differences in environmental sensitivity. The nonshared environment influences on EXT etiology were important for high-NE adolescents only on the negative ends of the parental scales, suggesting a diathesis-stress pattern, with high-NE adolescents vulnerable to negative (or lack of positive) environment. For friend support and school bullying, in contrast, the importance of the nonshared environment was high for high-NE adolescents all along the environmental scales, suggesting a pattern of differential susceptibility to both negative and positive environments.
An exception in the results of moderating effects on the nonshared environment variance in EXT was that differences in environmental exposure were somewhat important to EXT etiology, but to a lesser degree, for low-NE adolescents when school bullying levels were high. As low-NE adolescents, who are the typically “not-sensitive”, showed sensitivity to school environmental effects on both the phenotypic and environmental EXT variance, this further supports the domain-specificity perspective, demonstrating that different individuals could be sensitive to different environmental influences, especially as school effects are not frequently studied in this context. It is important to note, however, that some of our environmental predictors were found to have a meaningful heritable variation, suggesting that the variance in these “environmental” variables is at least partly attributable to heredity.
The results for the moderation effects on the expression of genetic variance in EXT are much more difficult to interpret under the framing of individual differences in environmental sensitivity. When a moderator moderates the expression of genetic variance, it is impossible to know whether these effects come from environmental suppression of genetic tendencies, causing under-expression, or rather that environmental effects facilitate genetic tendencies, causing over-expression. When testing the moderating effects of friend support on the genetic variance, the importance of genetic differences in EXT etiology increased as friend support increased, and to a lesser degree as the level of NE increased. This means that more friend support enables genetic tendencies for EXT to manifest, regardless of NE levels.
In contrast, the moderating effects of the parental domain and school bullying on the expression of genetic variance were moderated by NE. With parental support, for high-NE adolescents both positive parental environment and the lack of it enable genetic tendencies for EXT to manifest. It further seems that the lack of parental support facilitates genetic tendencies for EXT not only for high-NE adolescents, but for low-NE adolescents as well. Conflict with parents demonstrated a different pattern. Whereas high levels of conflict with parents facilitate genetic tendencies for EXT in low-NE adolescents, low levels of conflict with parents may facilitate genetic tendencies for EXT in high-NE adolescents. Finally, high levels of school bullying enable genetic tendencies for EXT to manifest in low-NE adolescents and to a lesser degree in high-NE adolescents, for which low levels of school bullying also facilitate genetic tendencies for EXT. This shows that different individuals, both those high and low on NE, are affected by their environment, but differently, further supporting domain-specificity.
The results from the genetic analyses mirror, to some extent, the results from the phenotypic analyses. Unfortunately, only one of the significant interactions in the phenotypic analysis could also be tested in the genetic analysis – the interaction between parental support and NE. While the phenotypic analysis found that both adolescents high, and low, on NE were sensitive to the influences of low parental support, the genetic analysis showed that the effect for high-NE adolescents comes from influencing both the genetic and environmental variance in EXT, and the effect for low-NE adolescents comes from influencing only the genetic variance in EXT. More generally, our results emphasize the importance of both personal and environmental factors to EXT etiology, similar to their importance for the phenotypic development of EXT.
Strengths, Limitations and Future Directions
This study has several strengths, both conceptual and methodological. First, the present study was the first to apply Purcell’s (2002) GxE twin model to study individual differences in environmental sensitivity, by using both environmental and personal moderators in the model. Even though this method allows for a new approach to study individual differences in environmental sensitivity and adds the perspective of its effects on the etiology of traits, its use is limited due to the sample size that is required to estimate such a complex model. Second, to gain a rather comprehensive understanding of environmental effects on EXT development during adolescence, we tested a wide range of environmental effects in the same sample, concerning positive and negative aspects of the parent, peer, and school domains. Third, this study used a multi-informant design, gathering information from adolescents, their parents, and official data assembled by the Ministry of Education. Fourth, this study showed longitudinal effects, of how aspects of the adolescent’s environment at age 11 affect their level of EXT at age 13, beyond the continuity of this behavior. It is important to note that in Israel, children typically attend elementary school until around age 12, then transferring to middle school. Therefore, when we measured EXT at age 13, most of our participants attended a different school than at the time of the school’s measurements. This means that at least for some adolescents, the characteristics of their previous school, and specifically the school’s positive climate, affected their current level of EXT, beyond its concurrent effect.
Future work should address several additional issues. First, the created EXT factor in our sample was not as reliable as we would like, explaining only 27–31% of the variance. This emphasizes the breadth of the construct of EXT, requiring further research to find a well-defined measure and replicate our findings with it. Second, even though we had quite a large sample size, it is not ideal for assessing many estimates within twin models. The age 13 sample was even smaller, preventing us from testing the effects on EXT etiology longitudinally. Future studies should aim to replicate our findings in a larger sample, as well as test it longitudinally. Third, although we tested a range of environmental influences on the adolescent, these do not begin to cover the range of social stressors the adolescent is exposed to. Therefore, future studies should explore individual differences in the influences of additional social domains, such as the neighborhood and romantic relationships, and of further aspects of the social domains included in the present study. Fourth, our multi-informant design enabled us to control for biases in report to some extent, but not fully (e.g. maternal reports on both NE and EXT). Future studies should aim to replicate our results with measures which are aggregated across different informants, excluding the option of single informant bias. Fifth, even though measuring the school environment at the school-level could be a more objective representation of the environment the individual is exposed to, it might not reflect their own feelings about the school. Future studies should test whether the current findings replicate when the school environment is measured at the individual level. Sixth, the current sample is relatively privileged as indicated by high levels of maternal education (see supplementary Note 1), which may lead to lower negative outcomes such as EXT. Future work should study varied samples. Finally, our results provide important support for domain-specificity in environmental sensitivity, but they are preliminary. Thus, future studies should aim to collect more evidence, on a broader variation of environments, outcomes, sensitivity markers, developmental periods, and samples.
Another possible limitation might relate to the interpretation of gene-environment interplay type. That is, even though we focus on GxE, gene-environment correlations (rGE) might also be at play. When estimating the genetic models without moderation effects, we were able to directly estimate the genetic correlation between EXT and peer problems, seemingly an “environmental” variable, and the existence of such a correlation can be seen as an example for rGE (Hasenfratz et al. 2015). However, Purcell’s (2002) moderation model estimates genetic variance in the moderators, as well as genetic correlations between the moderators and trait of interest (see supplementary Note 3), enabling to estimate GxE when there may be a genetic correlation between the moderator and trait of interest. As the moderator is typically, and in our case, regarded as an “environmental” variable, the model enables the interaction to be estimated beyond such rGE.
Conclusions
Our findings contribute to the literatures on both EXT and individual differences in environmental sensitivity. We demonstrate how, even when incorporating several environmental effects in the same analyses, different environmental domains influence EXT development above and beyond the effects of the other domains, but only when considering individual differences in temperament. We further found that personal and environmental characteristics, and in some contexts their interactions, are important for EXT etiology, as expressed in the genetic and environmental variance in EXT. Thus, our findings stress the importance of considering both a comprehensive account of the individual’s environment and person-based characteristics that might moderate these effects when studying EXT development.
The implications to the literature on individual differences in environmental sensitivity are twofold. First, our study offers a new approach to test individual differences in environmental sensitivity, by employing environmental and personal moderators in GxE twin models. Using these methods adds the possibility to understand whether the combined effects of the environment and the person affect development by influencing the genetic or the environmental variance. Second, our study joins the previous few studies (Belsky et al. 2021; Markovitch and Knafo-Noam 2021) that have found some evidence for domain-specificity of individual differences in environmental sensitivity, further supporting the possibility that individuals should not be treated as either sensitive to their environment or not, but rather that different individuals can be sensitive to different environmental influences or in different ways.
Availability of data and materials
Data is available upon request due to ethics considerations.
Code availability
Code is available upon request.
References
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Bates TC, Neale MC, Maes HH (2019) umx: A library for structural equation and twin modelling in R. Twin Res Hum Genet 22:27–41
Belsky J (2005) Differential susceptibility to rearing influence: An evolutionary hypothesis and some evidence. In: Ellis B, Bjorklund D (eds) Origins of the social mind: Evolutionary psychology and child development. Guilford, New York, pp 139–163
Belsky J, Hartman S (2014) Gene-environment interaction in evolutionary perspective: differential susceptibility to environmental influences. World Psychiatry 13:87–89. https://doi.org/10.1002/wps.20092
Belsky J, Pluess M (2009) Beyond diathesis stress: differential susceptibility to environmental influences. Psychol Bull 135:885–908. https://doi.org/10.1037/a0017376
Belsky J, Pluess M (2013) Beyond risk, resilience, and dysregulation: Phenotypic plasticity and human development. Dev Psychopathol 25:1243–1261. https://doi.org/10.1017/S095457941300059X
Belsky J, Zhang X, Sayler K (2021) Differential Susceptibility 2.0: Are the same children affected by different experiences and exposures. Dev Psychopathol. https://doi.org/10.1017/S0954579420002205
Boker S, Neale M, Maes H et al (2011) OpenMx: an open source extended structural equation modeling framework. Psychometrika 76:306–317. https://doi.org/10.1007/s11336-010-9200-6
Brislin R (1980) Translation and content analysis of oral and written material. In: Triandis H, Berry J (eds) Handbook of cross-cultural psychology, vol 2. Allyn & Bacon. Boston, MA, pp 349–444
Bronfenbrenner U, Morris PA (2006) The bioecological model of human development. In: Lerner RM, Damon W (eds) Handbook of child psychology, 6th edn. Wiley, Hoboken, NY, pp 793–828
Burk LR, Armstrong JM, Goldsmith HH et al (2011) Sex, temperament, and family context: how the interaction of early factors differentially predict adolescent alcohol use and are mediated by proximal adolescent factors. Psychol Addict Behav 25:1–15. https://doi.org/10.1037/a0022349
Burt SA (2009) Rethinking environmental contributions to child and adolescent psychopathology : a meta-analysis of shared environmental influences. Psychol Bull 135:608–637. https://doi.org/10.1037/a0015702
Bush NR, Lengua LJ, Colder CR (2010) Temperament as a moderator of the relation between neighborhood and children’s adjustment. J Appl Dev Psychol 31:351–361. https://doi.org/10.1016/j.appdev.2010.06.004
Buss AH, Plomin R (1984) Tempermant: Early developing personality traits. Lawrence Erlbaum Associates Inc, Hillsdale, New Jersy
Carter AS, Briggs-Gowan MJ, Jones SM, Little TD (2003) The Infant-Toddler Social and Emotional Assessment (ITSEA): Factor structure, reliability, and validity. J Abnorm Child Psychol 31:495–514. https://doi.org/10.1023/A:1025449031360
De Laet S, Colpin H, Van Leeuwen K et al (2016) Teacher-student relationships and adolescent behavioral engagement and rule-breaking behavior: The moderating role of dopaminergic genes. J Sch Psychol 56:13–25. https://doi.org/10.1016/j.jsp.2016.02.002
Dick DM, Viken R, Purcell S et al (2007) Parental monitoring moderates the importance of genetic and environmental influences on adolescent smoking. J Abnorm Psychol 116:213–218. https://doi.org/10.1037/0021-843X.116.1.213
DiLalla LF, Bersted K, John SG (2015) Peer victimization and DRD4 genotype influence problem behaviors in young children. J Youth Adolesc 44:1478–1493. https://doi.org/10.1007/s10964-015-0282-4
Du Rocher Schudlich TD, White CR, Fleischhauer EA, Fitzgerald KA (2011) Observed infant reactions during live interparental conflict. J Marriage Fam 73:221–235. https://doi.org/10.1111/j.l741-3737.2010.00800.x
Duncan LE, Keller MC (2011) A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am J Psychiatry 168:1041–1049. https://doi.org/10.1176/appi.ajp.2011.11020191.A
Elkins IJ, McGue M, Iacono WG (1997) Genetic and environmental influences on parent-son relationships: Evidence for increasing genetic influence during adolescence. Dev Psychol 33:351–363. https://doi.org/10.1037/0012-1649.33.2.351
Ellis BJ, Boyce WT, Belsky J et al (2011) Differential susceptibility to the environment: An evolutionary-neurodevelopmental theory. Dev Psychopathol 23:7–28. https://doi.org/10.1017/S0954579410000611
Essex MJ, Armstrong JM, Burk LR et al (2011) Biological sensitivity to context moderates the effects of the early teacher-child relationship on the development of mental health by adolescence. Dev Psychopathol 23:149–161. https://doi.org/10.1017/S0954579410000702
Feinberg ME, Button TMM, Neiderhiser JM et al (2007) Parenting and adolescent antisocial behavior and depression: Evidence of genotype× parenting environment interaction. Arch Gen Psychiatry 64:457–465. https://doi.org/10.1001/archpsyc.64.4.457
Fitzmaurice GM, Laired N, m, Ware JH, (2004) Applied longitudinal analysis. John Wiley & Sons, Hoboken, New Jersey
Goodman R (1997) The strengths and difficulties questionnaire: A research note. J Child Psychol Psychiatry Allied Discip 38:581–586. https://doi.org/10.1111/j.1469-7610.1997.tb01545.x
Grusec JE (2011) Socialization processes in the family: Social and emotional development. Annu Rev Psychol 62:243–269. https://doi.org/10.1146/annurev.psych.121208.131650
Hannigan LJ, Walaker N, Waszczuk MA et al (2017) Aetiological influences on stability and change in emotional and behavioural problems across development: a systematic review. Psychopathol Rev. https://doi.org/10.5127/pr.038315
Hartman S, Belsky J (2015) An evolutionary perspective on family studies: Differential susceptibility to environmental influences. Fam Process 55:700–712. https://doi.org/10.1111/famp.12161
Hasenfratz L, Benish-Weisman M, Steinberg T, Knafo-Noam A (2015) Temperament and peer problems from early to middle childhood: Gene-environment correlations with negative emotionality and sociability. Dev Psychopathol 27:1089–1109. https://doi.org/10.1017/S095457941500070X
Hicks BM, South SC, DiRago AC et al (2009) Environmental adversity and increasing genetic risk for externalizing disorders. Arch Gen Psychiatry 66:640–648. https://doi.org/10.1001/archgenpsychiatry.2008.554
Huesmann LR, Dubow EF, Boxer P (2009) Continuity of aggression from childhood to early adulthood as a predictor of life outcomes: Implications for the adolescent-limited and life-course-persistent models. Aggress Behav 35:136–149. https://doi.org/10.1002/ab.20300
Janssens A, Van Den Noortgate W, Goossens L et al (2017a) Externalizing problem behavior in adolescence: parenting Interacting with DAT1 and DRD4 genes. J Res Adolesc 27:278–297. https://doi.org/10.1111/jora.12271
Janssens A, Van Den Noortgate W, Goossens L et al (2017b) Adolescent externalizing behaviour, psychological control, and peer rejection: Transactional links and dopaminergic moderation. Br J Dev Psychol 35:420–438. https://doi.org/10.1111/bjdp.12184
Lengua LJ (2008) Anxiousness, frustration, and effortful control as moderators of the relation between parenting and adjustment in middle-childhood. Soc Dev 17:554–577. https://doi.org/10.1111/j.1467-9507.2007.00438.x
Lo S, Andrews S (2015) To transform or not to transform: using generalized linear mixed models to analyse reaction time data. Front Psychol 6:1–16. https://doi.org/10.3389/fpsyg.2015.01171
Long JA (2019) interactions: Comprehensive, user-friendly toolkit for probing interactions. https://cran.r-project.org/package=interactions
Markovitch N, Knafo-Noam A (2021) Sensitivity, but to Which Environment? Individual Differences in Sensitivity to Parents and Peers Show Domain-Specific Patterns and a Negative Genetic Correlation. Dev Sci 00:e13136. https://doi.org/10.1111/desc.13136
Ministry of Education (n.d.) (Almost) Everything About Schools. https://cms.education.gov.il/educationcms/applications/ims/homepage.htm
Mitchell C, McLanahan S, Brooks-Gunn J et al (2013) Genetic differential sensitivity to social environments: Implications for research. Am J Public Health 103:S102–S110. https://doi.org/10.2105/AJPH.2013.301382
Neale MC, Eaves LJ, Kendler KS (1994) The power of the classical twin study to resolve variation in threshold traits. Behav Genet 24:239–258. https://doi.org/10.1007/BF01067191
Neale MC, Hunter MD, Pritikin JN et al (2016) OpenMx 2.0: extended structural equation and statistical modeling. Psychometrika 81:535–549. https://doi.org/10.1007/s11336-014-9435-8
Pinquart M (2017) Associations of parenting dimensions and styles with externalizing problems of children and adolescents: an updated meta-analysis. Marriage Fam Rev 53:613–640. https://doi.org/10.1080/01494929.2016.1247761
Pitzer M, Jennen-Steinmetz C, Esser G et al (2011) Differential susceptibility to environmental influences: The role of early temperament and parenting in the development of externalizing problems. Compr Psychiatry 52:650–658. https://doi.org/10.1016/j.comppsych.2010.10.017
Pluess M (2015) Individual differences in environmental sensitivity. Child Dev Perspect 9:138–143. https://doi.org/10.1111/cdep.12120
Preacher KJ, Curran PJ, Bauer DJ (n.d.) Simple intercepts, simple slopes, and regions of significance in MLR 2-way interactions. http://www.quantpsy.org/interact/mlr2.htm
Preacher KJ, Curran PJ, Bauer DJ (2006) Computational tools for probing interactions in multiple linear regression, multilevel modeling, and latent curve analysis. J Educ Behav Stat 31:437–448. https://doi.org/10.3102/10769986031004437
Price TS, Freeman B, Craig I et al (2000) Infant zygosity can be assigned by parental report questionnaire data. Twin Res 3:129–133. https://doi.org/10.1375/136905200320565391
Purcell S (2002) Variance components models for gene-environment interaction in twin analysis. Twin Res 5:554–571. https://doi.org/10.1375/136905202762342026
R Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reef J, Diamantopoulou S, Van Meurs I et al (2011) Developmental trajectories of child to adolescent externalizing behavior and adult DSM-IV disorder: Results of a 24-year longitudinal study. Soc Psychiatry Psychiatr Epidemiol 46:1233–1241. https://doi.org/10.1007/s00127-010-0297-9
Reijntjes A, Kamphuis JH, Prinzie P et al (2011) Prospective linkages between peer victimization and externalizing problems in children: A meta-analysis. Aggress Behav 37:215–222. https://doi.org/10.1002/ab.20374
Rioux C, Castellanos-Ryan N, Parent S et al (2015) Differential susceptibility to environmental influences: Interactions between child temperament and parenting in adolescent alcohol use. Dev Psychopathol 28:1–11. https://doi.org/10.1017/S0954579415000437
Roisman GI, Newman D, a, Fraley RC, et al (2012) Distinguishing differential susceptibility from diathesis-stress: Recommendations for evaluating interaction effects. Dev Psychopathol 24:389–409. https://doi.org/10.1017/S0954579412000065
Rothbart MK, Bates JE (2006) Temperament. In: Lerner DR, Eisenberg N (eds) Handbook of child psychology (6th edition), vol 3. Social, emotional, and personality development. Wiley, New York, pp 99–176
RStudio Team (2020) RStudio: Integrated Development Environment for R. RStudio, PBC, Boston, MA. http://www.rstudio.com/
Slagt M, Dubas JS, Deković M, van Aken MAG (2016) Differences in sensitivity to parenting depending on child temperament: A meta-analysis. Psychol Bull 142:1068–1110. https://doi.org/10.1037/bul0000061
Slagt M, Dubas JS, van Aken MAG, Ellis BJ (2018) Sensory processing sensitivity as a marker of differential susceptibility to parenting. Dev Psychol 54:543–558. https://doi.org/10.1037/dev0000431
Smetana JG, Campione-Barr N, Metzger A (2006) Adolescent development in interpersonal and societal contexts. Annu Rev Psychol 57:255–284. https://doi.org/10.1146/annurev.psych.57.102904.190124
Ttofi MM, Farrington DP, Lösel F (2012) School bullying as a predictor of violence later in life: A systematic review and meta-analysis of prospective longitudinal studies. Aggress Violent Behav 17:405–418. https://doi.org/10.1016/j.avb.2012.05.002
Tung I, Noroña AN, Morgan JE et al (2018) Patterns of sensitivity to parenting and peer environments: Early temperament and adolescent externalizing behavior. J Res Adolesc 29:225–239. https://doi.org/10.1111/jora.12382
van IJzendoorn MH, Bakermans-Kranenburg MJ, (2015) Genetic differential susceptibility on trial: meta-analytic support from randomized controlled experiments. Dev Psychopathol 27:151–162. https://doi.org/10.1017/S0954579414001369
Vertsberger D, Abramson L, Knafo-Noam A (2019) The Longitudinal Israeli Study of Twins (LIST) reaches adolescence: genetic and environmental pathways to social, personality and moral development. Twin Res Hum Genet 22:567–571. https://doi.org/10.1017/thg.2019.94
Zentner M, Bates JE (2008) Child temperament: An integrative review of concepts, research programs, and measures. Int J Dev Sci 2:7–37. https://doi.org/10.3233/DEV-2008-21203
Zimet GD, Dahlem NW, Zimet SG, Farley GK (1988) The multidimensional scale of perceived social support. J Pers Assess 52:30–41. https://doi.org/10.1207/s15327752jpa5201_2
Acknowledgements
The authors would like to thank Lior Abramson, Dana Katsoty, Rotem Schapira and the research assistants who helped with data collection, and members of the Social Development Lab at the Hebrew University who commented on a previous version of the manuscript. The authors also thank the families in the Longitudinal Israeli Study of Twins (LIST) for their continued participation.
Funding
This work was funded by the Israel Science Foundation (grant # 1670/13 and grant # 1333/18). The first author was supported by a fellowship from the Israeli Committee for Advancing Psychometrics & the National Institute for Testing and Evaluation (ICAP & NITE).
Author information
Authors and Affiliations
Contributions
Conceptualization: NM, AKN; Methodology: AKN, NM; Formal analysis and investigation: NM, RMK; Writing—original draft preparation: NM; Writing—review and editing: NM, AKN, RMK; Funding acquisition: AKN; Supervision: AKN. All authors reviewed the results and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no relevant financial or non-financial interests to disclose.
Ethical approval
This study was approved by the ethics committee of the Hebrew University of Jerusalem.
Consent to participate
Parents provided informed consent for their children to participate; adolescents provided assent.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Markovitch, N., Kirkpatrick, R.M. & Knafo-Noam, A. Are Different Individuals Sensitive to Different Environments? Individual Differences in Sensitivity to the Effects of the Parent, Peer and School Environment on Externalizing Behavior and its Genetic and Environmental Etiology . Behav Genet 51, 492–511 (2021). https://doi.org/10.1007/s10519-021-10075-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10519-021-10075-7