
Abstract
Tilapia lake virus (TiLV) is a novel single-stranded RNA virus that is considered a threat to the universal tilapia industry. Recombinant technology proved to apply to different viruses and does not require handling live viruses. The present study developed a recombinant protein from segment 4 of TiLV with the aim of developing an immunological detection method for the virus. For this purpose, the complete coding sequence of segment 4 of the virus was optimized, amplified, and cloned in pET-28a( +), a prokaryotic expression vector, and the TiLV segment 4 construct (pET-seg4) was developed. The recombinant protein was expressed in BL21-competent Escherichia coli by isopropyl-β-D-thiogalactopyranoside (IPTG) induction. The successful expression of the recombinant protein was confirmed by SDS-PAGE. The polyclonal antibody (PAb) raised against the recombinant protein was used for the indirect enzyme-linked immunosorbent assay (ELISA) development to detect TiLV in the pooled kidney and mucus samples. The coating concentrations for the recombinant protein and the TiLV-positive samples were standardized as 3.125 µg and 6.25 µg, respectively, and the optimum polyclonal antibody dilution was found as 1:800. The diagnostic sensitivity and diagnostic specificity of the assay were 82.35% and 100%, respectively. The recombinant protein developed in this study provided a better understanding of raising a codon-optimized protein and its use in developing an indirect ELISA test for TiLV, which can also be used as a recombinant vaccine for immunizing tilapia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aquaculture is a sector growing at a faster pace than other food animal sectors of the world and plays a pivotal role in global food demand, in addition to nutritional security and socio-economic welfare. The United Nations predicts the world population to outreach 8.5 billion by 2030 (United Nations Department of Economic and Social Affairs Population Division 2019). Tilapia, which belongs to the family Cichlidae, contributes 10% (6.4 mt) of the world aquaculture production, of which 8% by Nile tilapia (FAO 2018) has the potential to supply sufficient food and nutrition in an environmentally and economically sustainable manner. The tremendous growth in aquaculture with high stocking density increased the transmission of viruses between organisms, leading to the discovery of many aquaculture viruses. TiLV is an emerging viral disease responsible for substantial economic loss and mass mortality, reaching 90% in severe cases (Fathi et al. 2017; Surachetpong et al. 2017). The emergence of TiLV infection, the first-ever significant disease of tilapia, has put the global tilapia industry at risk (Jansen et al. 2018). Eyngor et al. (2014) first reported the virus from Israel, and subsequently, the virus was reported from Ecuador (Ferguson et al. 2014), Columbia (Kembou Tsofack et al. 2017), Egypt (Fathi et al. 2017; Nicholson et al. 2017), Thailand (Surachetpong et al. 2017; Dong et al. 2017a, b), and recently in India with more than 85% mortality (Behera et al. 2018). Recently, in India, co-infection of TiLV with another novel pathogen (Tilapia parvovirus) has been reported (Rajendran et al. 2023).
TiLV is an enveloped virus possessing negative-sense, single-stranded RNA with 10 segments (Eyngor et al. 2014; Bacharach et al. 2016; Surachetpong et al. 2017) with a diameter ranging from 55 to 100 nm (Ferguson et al. 2014; Eyngor et al. 2014; Surachetpong et al. 2017). The 10 segments each have an open reading frame (ORF); each segment has a size range of 456 to 1641 nucleotides with a total genome size of 10.323 Kbp (Bacharach et al. 2016). Criollo-Joaquin et al. (2019) reported a 12% structural homology between the segment 4 hypothetical protein of TiLV and the influenza A virus neuraminidase (NA) protein. Recently, the virus has been classified as Tilapia tilapinevirus, the only species in the monotypic genus Tilapinevirus, the only genus in the family Amnoonviridae (ICTV 2018). Currently, TiLV sequences available in the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/) are from Ecuador, Egypt, India, Israel, Malaysia, Tanzania (Lake Victoria), Thailand, and Uganda (Lake Victoria). Currently, two whole-genome sequences are available, one from Israel (Bacharach et al. 2016) and the other from Thailand (Surachetpong et al. 2017). Also, the complete coding sequences of TiLV have been reported by Subramaniam et al. (2019).
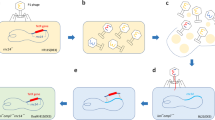
Available information about the virus is at an infant stage where most functional proteins encoded by each segment and their antigenic properties are not yet known and understood. Most of the diagnostic tests available for TiLV are based on molecular detection, cell culture isolation of the virus, or novel techniques of histopathological lesion scoring (Valsalam et al. 2024). Only a few methods based on immunoassay are available for detecting TiLV infection so far (Hu et al. 2020). In a cohabitation challenge study in red hybrid tilapia, the survivors developed protective immunity against segment 4 of TiLV (Tattiyapong et al. 2020). In a further study using bioinformatics, genetics, and biochemical tools, the segment 4 of TiLV was identified as a nucleoprotein (NP) with the essential properties for the replication of the negative-sensed RNA genome (Abu Rass et al. 2022). Fulfilling the desired characteristics on its immunogenicity and identification as a NP, the current study chose segment 4 as a target for the development of recombinant protein. The recombinant protein from the selected gene segment of TiLV can be utilized as a positive control for diagnostic test development, including ELISA and also has the potential as a vaccine candidate in further research on TiLV by testing its immunogenicity in the host. However, developing a recombinant protein requires the adequate expression of the selected heterologous protein in the prokaryotic expression host. The expression of the selected gene from segment 4 of TiLV is hampered in the prokaryotic expression system because of codon bias, a phenomenon when the codon usage of the heterologous protein differs from that of the bacterium. Hence, the current study provides the method of developing a codon-optimized recombinant protein of TiLV from its segment 4 as it shows homology with the NA protein of another virus. This method of developing the recombinant protein by biomolecular engineering provides a way of constructing the proteins artificially (using engineered codons) in the laboratory setting without the actual involvement of the virus but fulfilling various applications in diagnostics and vaccination. The developed recombinant protein is further used for the TiLV diagnosis in the fish kidney and mucus samples using the polyclonal antibody (PAb) raised against it in the current study. The ELISA-based immunological detection assays are advantageous by bypassing all upstream and downstream processing of molecular detection methods.
Materials and methods
Ethical approval, bacterial strain, and plasmid
The permission numbers for the present study by the Institute Animal Ethics Committee (IAEC) and Institutional Biosafety Committee are F.No. CIFE/IBSC/2022-23 and CIFE-03-2019, respectively. HiPurA™ Escherichia coli BL21(DE3) competent cells were procured from HiMedia, India, and were used as a host strain. Plasmid pET-28a( +) was used as an expression vector procured from Allianz BioInnovation.
Retrieval of segment 4 sequences of TiLV ORF from GenBank and sequence analysis
All available isolates of the TiLV segment 4 gene (complete coding sequence) were retrieved from the National Center for Biotechnology Information (NCBI). The different isolates of this segment were analyzed using bioinformatics tools such as standard nucleotide alignment (BLAST) and multiple sequence analysis clustalΩ (Omega) (Sievers et al. 2011). Antigenicity prediction of TiLV segment 4 was made by the PREDICTED ANTIGENIC PEPTIDES program (http://imed.med.ucm.es/Tools/antigenic.pl). For structure validation, the Ramachandran plot was generated from the amino acid sequence of TiLV-seg4 by SWISS-MODEL Interactive Workspace (https://swissmodel.expasy.org/interactive). Secondary structure composition was generated from the amino acid sequence of TiLV segment 4 by the PredictProtein program (https://predictprotein.org/).
Optimizing rare codons of E. coli within the TiLV gene to enhance protein expression
Codon usage bias was analyzed for the TiLV segment 4 gene (MK392375.1) nucleotide sequence to make it compatible for expression by the E. coli expression host. This was done using the Rare Codon Analysis tool (GenScript, USA) (https://www.genscript.com/tools/rare-codon-analysis). After codon usage analysis, a 1065 bp-long construct containing codon-optimized TiLV segment 4 gene sequence was synthesized by GenScript (Piscataway, NJ USA) company. The optimized sequence was further used to clone the pET-28a( +) expression vector and transform E. coli BL21(DE3) competent cells.
PCR of TiLV segment 4, Restriction enzyme (RE) digestion and purification
The codon-optimized segment 4 gene of TiLV was amplified using the primers in Table 1. The PCR conditions include initial denaturation for 2 min at 94 °C, followed by 35 cycles of denaturation at 94 °C (30 s), annealing at 51 °C for 60 s, and extension at 72 °C for 30 s, with a final extension step of 7 min at 72 °C. The PCR amplicon was cleaned using the GeneJET Gel Extraction Kit (Thermo Scientific, USA) and digested with NdeI and XhoI enzymes (FastDigest Thermo Scientific) in a reaction volume of 20 μL by adding segment 4 gene (up to 1 μg), 10 × FD buffer (2 μL), NdeI (1 μL), XhoI (1 μL), and nuclease-free water, and the mixture was incubated overnight (30 °C) in a water bath. After a short spin, the enzymes were inactivated at 65 °C for 5 min. The pET-28a( +) vector was digested with NdeI (1 μL) and XhoI (1 μL). The insert and the vector were purified after digestion, and the concentration of the fragments was analyzed using the NanoDrop spectrophotometer (Thermo Scientific).
Cloning in expression vector and transformation of HiPurA™ BL21(DE3) competent cells
The RE digested and purified insert (segment 4) and pET-28a( +) vector were ligated using the T4 DNA ligase (Thermo Scientific, USA) in a 20-µL total reaction volume. The reaction mixture was incubated at 16 °C overnight in a water bath and used to transform E. coli BL21(DE3) competent cells. The transformation was performed following the manufacturer’s instruction for E. coli BL21(DE3) competent cells (HiMedia, India). Briefly, the tubes containing BL21(DE3) competent cells were thawed on ice, and 100 µL was dispensed into another tube to which 50 ng of the recombinant plasmid (pET-seg4) was added, and the mixture was retained on ice for 30 min, followed by a heat shock for 45 s at 42 °C in the water bath and placed on ice for 2 min. To the mixture, 0.9 mL of SOC media was poured and incubated for 1 h at 37 °C, and 100 µL was spread on LB (Luria–Bertani)-kanamycin (50 µg/mL) plates and incubated at 37 °C overnight. The segment 4 gene was cloned into the pET-28a( +) expression vector (Allianz BioInnovation, India).
Plasmid purification and confirmation of TiLV insert in pET-28a( +) by RE digestion
The pET-seg4 plasmid was isolated using GeneJet Plasmid Miniprep Kit (Thermo Scientific, USA) as per the manufacturer’s protocol. Briefly, the positive colony from the culture plate was inoculated in LB kanamycin broth and grown overnight. The culture was harvested and resuspended in 0.2 mL of resuspension buffer; lysis solution (0.2 mL) was added and mixed by inverting gently. A neutralization solution of 0.35 mL was added and centrifuged (12,000 rpm for 5 min). The supernatant from this mixture was transferred to the GeneJet spin column and centrifuged (1 min at 12,000 rpm). The wash solution (0.5 mL) was added and centrifuged (12,000 rpm for 1 min), and the step was repeated. The column was transferred to another micro-centrifuge tube and eluted with 30 μL of elution buffer. A 20-μL reaction was prepared for RE digestion using NdeI and XhoI for the plasmid DNA (pET-seg4) (1 μg). The mixture was analyzed on 1% agarose gel and documented by the gel documentation system (GENESYS, USA).
Expression of protein in HiPurA™ BL21(DE3) cells
The hypothetical protein of the inserted gene was expressed in E. coli BL21(DE3) cells by isopropyl β-D- 1-galactopyranoside (IPTG) induction. Briefly, the E. coli BL21(DE3) cells containing the pET-seg4 were cultured in 10 mL of LB broth (50 µg/mL of kanamycin) and incubated at 37 °C overnight (180 rpm). The cells were transferred to a fresh medium and incubated at 37 °C for a few hours (h) to reach an OD value of 0.8 at 600 nm. Expression was stimulated by adding IPTG to a final concentration of 10 mM (not added to the sample, which served as non-induced control); cultures were further grown with continuous shaking (180 rpm) overnight at 15 ºC. The cells were harvested at three points: 5, 8, and 12 h post-induction by centrifugation at 6000 rpm for 10 min.
SDS-PAGE analysis of expressed protein (Laemmli 1970)
The expression of the recombinant segment 4 proteins was confirmed by SDS gel electrophoresis. The induced cells were harvested after discarding the supernatant; the cells were resuspended in cell lysis buffer. The samples were boiled (5–10 min at 100 °C) and loaded in sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE). The electrophoresis unit ran at 55 mA under constant voltage till the sample crossed stacking gel, and the current was increased to 150 mA and allowed to run through the resolving gel. Proteins within the gel were fixed by soaking in methanol solution for 1 h, then soaking in Coomassie dye overnight on the rocking platform and destained for background removal.
Recombinant protein purification
The NiNTA protein purification method was adapted from Gupta et al. (1999). Briefly, the pellet was taken from 1 L of culture and resuspended in 25 mL of buffer (300 mM sodium chloride, GuHCl 6 M, and sodium phosphate 0.1 M, pH 7.8). The content was stirred (room temperature—RT, 30 min), centrifuged (30 min, 10,000 × g at 4 °C), and the supernatant was mixed (RT, 30 min) with 50% Ni–NTA (4 mL). The content was loaded into the column and washed with buffer (300 mM NaCl, sodium phosphate 0.1 M, pH 7.8, and 8 M urea). The Ni–NTA resin-containing protein was washed with urea gradient (8 to 0 M) in buffer (sodium phosphate 0.1 M, pH 7.8, and 300 mM NaCl), and further washing was done using buffer (500 mM NaCl and sodium phosphate 0.1 M, pH 6.0) and the protein was eluted with a gradient of Imidazole (0–400 mM) in buffer (sodium phosphate 0.1 mM, pH 7.0, and 10% glycerol). The protein was dialyzed against 1 L chilled PBS (pH 7.4) in a rocking platform overnight at 4 °C with the change of PBS solution every 3–4 h. Using the BSA standard curve method, the protein concentration was measured using an EPOCH2 reader (BioTek).
PAb production against TiLV-seg4 recombinant protein and agar gel immunodiffusion test (AGID)
The study employed a male 2.0-kg New Zealand white rabbit that was maintained at the animal house of the ICAR-Central Institute of Fisheries Education in Mumbai. Freund’s complete adjuvant (FCA) and Freund’s incomplete adjuvant (FIA) were utilized for the primary and booster immunizations, respectively, with the TiLV-seg4 protein in a 1:1 (V/V) ratio. Subcutaneous administration of the antigen was used at 200 µg antigen per animal; vaccination and booster shots were administered (Leya et al. 2021), and serum was taken. For the AGID test, 1.2% agarose gel was prepared with 8% NaCl in a glass slide (Valsalam et al. 2023); the center well was filled with the recombinant protein, the surrounding wells received the serum, and the slide was incubated in a humid box at RT for 48 h.
Western blot analysis of recombinant protein
To ensure the expressed recombinant protein of segment 4 western blot was generated, the protein was separated by SDS-PAGE and transferred to nitrocellulose membrane (Sigma) using the semi-dry electroblotting (trans-blot SD semi-dry transfer cell, Bio-Rad) apparatus. Blocking was done overnight at 4 °C using the blocking buffer (5% non-fat dried milk in PBS buffer containing 0.05% Tween 20, PBST). The membrane was washed thrice in PBST and incubated with the PAb (1:500) in the blocking buffer for 1 h at RT. After repeating the washing steps, the membrane was incubated for 1 h at RT with the anti-rabbit IgG conjugated with HRP (horseradish peroxidase) (1:6000 dilution). After final washing, the membrane was treated with DAB (3,3ʹ-diaminobenzidine).
Indirect ELISA test development
For developing the protocol of indirect ELISA test (Lin 2015; Kohl and Ascoli 2017) for specific antigen detection, the standardization of the procedure was done by checkerboard titration of the PAb dilution (dilution range from 1:200 to 1:1600) and coating recombinant protein dilution (dilution range from 1.56 to 25 µg); here, the recombinant protein was coated as antigen. Further standardization was done for the detection of TiLV antigen in tissue samples by another checkerboard titration of the PAb dilution (dilution range from 1:200 to 1:1600) and the TiLV-positive tissue sample dilution (50 µg and 1.56 µg). At this step of standardization, for the validating ability of PAb for TiLV detection in infected tissues, the tissue homogenate previously detected as TiLV positive by using the reverse transcription-polymerase chain reaction (RT-PCR) was used. Further validation of the developed assay (for finding the diagnostic sensitivity and specificity) was done by testing the pooled (three in one) samples of the tilapia kidney (10 pools along with a blank negative control and goldfish kidney homogenate as tissue-negative control) and tilapia mucus (8 pools) samples by both RT-PCR and indirect ELISA.
The detailed ELISA protocol was as follows. In a 96-well ELISA plate (Greiner Bio-One), the wells were coated with the recombinant protein/tissue homogenate in the coating buffer and incubated overnight at 4 °C. The plate was washed thrice with washing buffer (PBST 0.01 M PBS, pH 7.2, 0.05% Tween 20). The unbound sites were blocked with BSA (5% bovine serum albumin, Genetix Biotech Asia Pvt. Ltd.) for 1 h at 37 °C, and washing was repeated as mentioned above. The PAb treatment was given for 1 h at 37 °C, and washing was done three times. The anti-rabbit IgG HRP conjugate treatment was given for 1 h at 37 °C. After the final washing for about four times, the colorimetric detection was done using 100 µL TMB (3,3ʹ, 5,5ʹ-tetramethylbenzidine). The data were investigated using the Statistical Package for Social Sciences (IBM SPSS 22).
Result
Sequence analysis, antigenicity prediction, and structure validation of TiLV segment 4 protein
Multiple sequence alignment (CLUSTAL Ω) of the complete coding sequence of the segment 4 gene showed 100% similarity between two Israeli isolates, while other isolates have 96–97% similarity at the nucleotide level, as shown in Supplementary Fig. 1. The antigenicity prediction of the amino acid sequence of the segment 4 gene showed an average antigenic propensity of 1.0362 and 15 antigenic determinants available in the sequence, as shown in Supplementary Fig. 2. The Ramachandran plot and secondary structure composition for the hypothetical protein validated the protein structure, as shown in Figs. 1 and 2. The desired results obtained in Figs. 1 and 2 showed the functional significance of the conformation of the selected protein to determine the structure of recombinant segment 4 protein produced in E. coli for successful expression. The Ramachandran plot showed the dense array of right-handed α helix; a moderate array of parallel, anti-parallel, and right-twisted β sheets was found with only a few amino acids showing a left-handed α helix. The result showed the antigenicity being primarily located in the favored region of α-helix and β-sheet. Most of the protein part consisted of loops followed by helices and strands. The presence of interconnecting loops confirms the possibility for the recombinant protein to be produced in the native conformation.

Ramachandran plot generated from the amino acid sequence of TiLV-seg4 generated by SWISS-MODEL Interactive Workspace

Secondary structure composition generated from the amino acid sequence of TiLV segment 4 by PredictProtein program
Codon optimization of segment 4 nucleotide sequence
Since a heterologous host was used to express the protein, codon optimization was done by deleting rare codons within the selected segment and replacing them with another codon that the expression host prefers to use for the same amino acid. A 1065-bp long codon-optimized artificial TiLV-seg4 gene was synthesized by GenScript company, as shown in Supplementary Fig. 3 (A) after codon usage analysis in the heterologous prokaryotic host (E. coli). The codon adaptation index (CAI) of the original gene in the desired expression host was 0.33, which was optimized to the CAI value of 0.97 in synthesized nucleotide intended for better expression of the desired gene. The optimized findings are shown in Supplementary Fig. 3 (B).
Cloning in pET-28a( +) vector and expression in HiPurA™ BL21(DE3) cells
The synthesized codon-optimized TiLV-seg4 was amplified using primer pair (TiLV4-F and TiLV4-R) and RE digested; the expected size of the PCR product, i.e., 1065 bp for segment 4, was visualized in 1% agarose gel, as shown in Supplementary Fig. 4. Similarly, the pET-28a( +) vector was digested and ligated with the gene segment of interest to develop the pET-seg4 gene construct. A schematic representation of the cloning process is shown in Supplementary Fig. 5. The positive clones were selected, and the clone was confirmed by RE digestion of the plasmid to release 1065 bp segment 4 of the TiLV gene from the 5369-bp pET-28a( +) vector (Fig. 3). The segment 4 gene encoding a hypothetical protein was expressed as a recombinant protein using the expression vector in E. coli BL21(DE3) competent cells. SDS-PAGE was used to analyze the induced and uninduced samples with different induction periods, and the result showed a noticeable band corresponding to 35 kDa in the induced sample with a 12-h induction period. No protein was observed at a similar position for uninduced and induced samples with a 5- and 8-h induction period (Fig. 4). The protein was purified, and the yield was measured as 20 mg/L, which was low.

Agarose gel electrophoresis (1%) showing RE digestion (Nde I and Xho I) of pET-seg4 showing the release of 1065 bp segment 4 of TiLV gene from 5369 bp pET28 vector confirming the cloning. L: 1065 bp segment 4 gene released from 5.3 kb pET-28a( +) vector; M: gene ruler 1 kb DNA ladder (Thermo)

SDS-PAGE (10% Tris-glycine) showing the expression of 35 kDa recombinant protein (segment 4); M-prestained protein marker (BLUeye-GeneDireX); UI1, 2, 3 un-induced samples of 5, 8, 12 h; I1, I2, I3 induced samples 5, 8, and 12 h post-induction with 10 mM IPTG; I4: purified protein (̴ 35 kDa)
AGID and indirect ELISA test development using the PAb raised against the recombinant protein
The immune reactivity of the raised TiLV-seg4 PAb with the recombinant protein is shown by the line of precipitation in the AGID test (Fig. 5). The successful expression of the segment 4 recombinant protein is confirmed by corresponding western blot as shown in Fig. 6. The optimized concentration of the recombinant protein per well for ELISA detection was 3.125 µg at 1:800 PAb dilution (Fig. 7). Similarly, the optimum RT-PCR positive tissue concentration for ELISA detection was estimated as 6.25 µg per well at 1:800 PAb dilution (Fig. 8). The developed assay was employed to detect TiLV in the pooled samples (kidney and mucus). The goldfish kidney homogenate as a negative tissue control showed no reactivity with PAb. The cutoff (mean of negative control + 3 × standard deviation) was calculated (Classen et al. 1987) as 0.0751, and the results showed that 8/10 pools were positive in the kidney tissue (Fig. 9) and 6/8 pools were positive in the mucus samples (Fig. 10), indicating 80% and 75% positive rate in the kidney and mucus samples, respectively. To assess the diagnostic sensitivity and specificity of the developed test, the results were compared with the RT-PCR test. The diagnostic sensitivity and diagnostic specificity of the current test were 82.35% and 100%, respectively (Table 2).

Agarose gel immunodiffusion test: The line of precipitation is seen between healthy 1 (recombinant protein) and well 2 (PAb); wells 3 and 4 are prebleed and nonspecific rabbit sera, respectively

Western blot detection of recombinant segment 4 protein (̴ 35 kDa) of TiLV using PAb. Lane M: ladder; lanes 1 and 2: segment 4 recombinant protein

Indirect ELISA detection limits of the recombinant protein of segment 4 with various dilutions of the protein and PAb. Mean ± SE

Indirect ELISA detection limits of TiLV in RT-PCR positive fish tissue extracts (pool of the kidney, liver, brain) with various tissue homogenates and PAb dilutions. Mean ± SE

Application of the indirect ELISA test for detecting TiLV in pooled (three in one) kidney samples of tilapia (number of pools = 10). The OD values along the Y-axis represent the mean absorbance, and the values above the cutoff line are considered positive. Tissue homogenate from goldfish was used as a nonspecific control for the test. Samples are represented as M ± SD

Application of the indirect ELISA test for detecting TiLV in pooled (three in one) mucus samples of tilapia (number of pools = 8). The OD values along the Y-axis represent the mean absorbance, and those above the cutoff line are considered positive. Samples are represented as M ± SD
Discussion
Disease diagnosis is a pivotal step in disease prevention, and developing a rapid diagnostic tool is a requisite for health management in aquaculture. Of the different disease prevention methods, vaccination stands out as one of the most effective and efficient methods compared to other preventive measures (Bedekar et al. 2020). Recombinant protein vaccines are one of the recent approaches in vaccinology that can elicit both humoral and cell-mediated immune responses when administered with a suitable adjuvant. Also, recombinant protein vaccines are suitable against viral diseases as they do not require handling live viruses (Kumari et al. 2018). To develop the recombinant protein, segment 4 of TiLV was selected as a model sequence as it was reported to have 12% structural homology with NA protein (transmembrane protein) of influenza A virus capsid (Criollo-Joaquin et al. 2019) and is believed to be a potential antigen of the virus. This structural homology may be due to the highly mutant viral capsid protein but maintaining a conserved domain within a subtype. Contrastingly, an analysis of the TM (transmembrane) domain by PSIPRED software showed that segment 4 does not have any transmembrane helix region (Acharya et al. 2019). Mikalsen et al. (2005) findings indicated that the viral capsid proteins of hemagglutinin esterase (HA) and NA are antigens against isavirus and influenza virus. Koren and Nylund (1997) explained that the NA played a vital role in virus replication by releasing virion particles from infected cells by breaking down the sugars that bind the mature virion particles. Eichelberger et al. (2018) suggested that NA enzymes may be an excellent option for developing vaccines against Orthomyxovirus as antibodies against NA bind to conserved domains of virus subtypes, which protect against heterologous viruses. These reports further increased the probability of segment 4 coding for a significant antigen of TiLV.
All available isolates (complete coding sequence) of segment 4 were retrieved from NCBI, and their similarities were analyzed using CLUSTAL Ω with a resultant sequence similarity of 96–87%. Phylogenetic analysis showed that the segment 4 gene of TiLV presents significant evolutionary proximity with the Dhori virus, an orthomyxovirus (Acharya et al. 2019). In a previous study by Pulido et al. (2019), it was found that the genome sequence of TiLV of the Peruvian isolate is 96.89–97.13%, similar to that of the Israeli isolates. As per the sequence analysis using programs like SWISS-MODEL, PREDICTED ANTIGENIC PEPTIDES (Kolaskar and Tongaonkar 1990), and PredictProtein, the selected gene was expected to code for a significant viral antigen and is thus suitable for cloning and production of recombinant protein. All the antigenic propensity values greater than 1.00 are considered potential antigenic determinants that can elicit an antibody response (Hasan et al. 2013), and the current study found the value for segment 4 as 1.0362, which confirms the immunogenic potentiality of the hypothetical protein. “The capability to foresee epitopes from amino acid sequence data alone is potentially beneficial, even though only a single determinant can be predicted with assurance for any given molecule” (Hopp and Woods 1981). Segment 4 showed the availability of 15 antigenic determinants, proving the future utility as a recombinant vaccine candidate.
The Ramachandran plot predicts the protein’s structural stereochemical property by ψ–ϕ main-chain torsion angles, and for a successful vaccine candidate, secondary structure characters play an essential role. The most common secondary structures of this hypothetical protein are helices and β-sheets where the antigenicity is predominantly positioned. In a previous study, a similar plot was generated for ROP16 for identification as a vaccine against Toxoplasma gondii; 84.64% of the amino acid residues were located in the favored region (Ghaffari et al. 2020). A recent study by Acharya et al. (2019) showed that all the fourteen predicted ORFs of the TiLV genome, including segment 4 ORF, consist of residues liable to be an alpha-helix and beta-sheet. The secondary structure generated using PredictProtein (Bernhofer et al. 2021) by analyzing the amino acid sequence of segment 4 showed that the hypothetical protein is composed mainly of loops followed by the secondary structures helix and strand, respectively. The loops play a chief role in the conformational changes occurring in diverse domains of the protein by connecting the helices and strands (Papaleo et al. 2016).
The segment 4 gene sequence showed a low CAI value (0.33) for expression in E. coli, which may lead to a feeble expression by the host and requires a CAI value > 0.8. Hence, the sequence was optimized, resulting in a CAI of 0.97, which has a high expression level. In a previous study by Kathwate (2022), after codon optimization, a CAI of 1.0 was achieved for good probable expression of the vaccine in E. coli K12. Rare codons in E. coli signify less than 2–8% of their corresponding codon partners (Sahdev et al. 2008). Kane (1995) observed that clusters of codons such as AUA, AGG/AGA, CUA, CGA, and CCC can decrease the quality and quantity of expressed proteins. Goldman et al. (1995) further pointed out that introducing rare codons incompatible with heterologous host genome during recombinant protein expression prompts the reduction in overall expression rate levels. The codon optimization enhances protein expression by > 1000-fold (Mauro 2018). A previous study revealed that codon optimization significantly enhances the expression of the gene that codes for the F2 domain located within the erythrocyte binding antigen (EBA-175) of Plasmodium falciparum (Yadava and Ockenhouse 2003). Apart from the CAI value, the GC content of segment 4 was also optimized to prolong the half-life of the mRNA.
In a recent study, Criollo-Joaquin et al. (2019) successfully inserted the segment 4 gene (predicted NA gene) into the expression plasmid vector pCMV. The present study used a pET-28a( +) vector for cloning and expressing segment 4 of TiLV. The pET-28a( +) vector was also used (Rodiansyah et al. 2022) for cloning and overexpression of “staphylococcal enterotoxin B gene synthetic (SEBsyn)” as a therapeutic component against cancer cells. In this study, a band of approximately 35 kDa was observed for induced samples taken 12 h post-induction. This approximate band size confirmed recombinant protein expression, as the expected protein size for segment 4 was around 39 kDa when estimated using the DNA to Protein program (http://insilico.ehu.es/translate/).
The ELISA-based immunological detection tests can be employed with the advantages of avoiding all the upstream (RNA extraction, purity check, and cDNA synthesis) and downstream (agarose gel electrophoresis) processes used for molecular detection, where the downstream processing is reduced as in the case of qRT-PCR; the cost required for testing mass scale sampling is affordable and less in ELISA than molecular testing (Valsalam et al. 2023). Various ELISA-based detection techniques were developed by previous researchers for the diagnosis of several pathogens, for example, ELISA to detect Pasteurella piscicida in culture and in “spiked” fish tissue (Bakopoulos et al. 1997), rapid detection of fish rhabdoviruses by ELISA (Dixon and Hill 1984), and rapid detection of VHSV (viral hemorrhagic septicemia virus) in fish by ELISA (Olesen and Jørgensen 1991). A recombinant protein (S8)–based indirect ELISA test for anti-tilapia lake virus IgM detection in sera was earlier developed (Hu et al. 2020). The present study portrayed the possibility of using recombinant protein from segment 4 of the virus as a possible means of diagnostic test development. Another recombinant protein from TiLV segment 8 (VP20) was tested as a vaccine candidate in tilapia, and the findings showed that the recombinant protein, along with the corresponding DNA vaccine, was more effective compared to the separate administration (Zeng et al. 2021).
The segment 5 and 6 proteins (S5196-272 and S6200-317) of TiLV were expressed in E. coli and proved to be a potential vaccine candidate for Nile tilapia (Lueangyangyuen et al. 2022). A specific 36-kDa protein band from segment 4 of TiLV was immunogenic in the survivors of TiLV-infected tilapia by Tattiyapong et al. (2020). The potential of the current recombinant protein (segment 4) needs to be further tested to prove the above findings. The ELISA test in the present study showed a very sensitive cutoff for effectively identifying the diseased fish. An indirect ELISA test earlier developed for the TiLV detection using the TiLV-Ab raised against the whole viral proteins showed 92.3% and 89.7% positive rates in the tested pools of mucus and kidney, respectively (Valsalam et al. 2023), whereas the TiLV-seg4 PAb in the current study resulted in 75% and 80% positive rates in the mucus and kidney pools, respectively, with high specificity. The study was conducted to investigate the suitability of anti-segment 4 PAb for detection of the TiLV antigen in tissue homogenate (protein concentration ranging from 50 to 1.56 µg). At present, the protocol developed is for a qualitative test; however, the assay can be upgraded for the quantitative detection (protein concentration) of the virus by establishing the reference standards of known virus copy number and recombinant protein.
Conclusion
The pET-seg4 containing codon-optimized synthetic TiLV-seg4 was successfully expressed in BL21(DE3) competent E. coli cells by IPTG induction and analyzed by SDS-PAGE. The segment 4 recombinant protein (35 kDa) of TiLV can be used as a positive control for developing various immunodiagnostic methods for virus detection and can be tested as a recombinant vaccine for immunizing tilapia. The application of the recombinant protein as a tool for the ELISA test development is shown in the present study.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Abu Rass R, Kustin T, Zamostiano R, Smorodinsky N, Ben Meir D, Feder D, Mishra N, Lipkin WI, Eldar A, Ehrlich M, Stern A (2022) Inferring protein function in an emerging virus: detection of the nucleoprotein in tilapia lake virus. J Virol 96(6):e01757-e1821
Acharya V, Chakraborty HJ, Rout AK, Balabantaray S, Behera BK, Das BK (2019) Structural characterization of open reading frame-encoded functional genes from tilapia lake virus (TiLV). Mol Biotechnol 61:945–957. https://doi.org/10.3390/jmse10010100
Bacharach E, Mishra N, Briese T, Zody MC, Tsofack JEK, Zamostiano R, Berkowitz A, Ng J, Nitido A, Corvelo A, Toussaint NC (2016) Characterization of a novel orthomyxo-like virus causing mass die-offs of tilapia. Mbio 7(2):e00431-e516. https://doi.org/10.1128/mBio.00431-16
Bakopoulos V, Volpatti D, Papapanagiotou E, Richards R, Galleotti AA (1997) Development of an ELISA to detect Pasteurella piscicida in culture and in ‘spiked’ fish tissue. Aquaculture 156:359–366
Bedekar MK, Kole S, Tripathi G (2020) Biotechnological approaches to fish vaccine. In: Genomics and biotechnological advances in veterinary, poultry, and fisheries. Academic Press, pp 407–419. https://doi.org/10.1016/B978-0-12-816352-8.00017-5
Behera BK, Pradhan PK, Swaminathan TR, Sood N, Paria P, Das A, Verma DK, Kumar R, Yadav MK, Dev AK, Parida PK (2018) Emergence of tilapia lake virus associated with mortalities of farmed Nile tilapia Oreochromis niloticus (Linnaeus 1758) in India. Aquaculture 484:168–174. https://doi.org/10.1016/j.aquaculture.2017.11.025
Bernhofer M, Dallago C, Karl T, Satagopam V, Heinzinger M, Littmann M, Olenyi T, Qiu J, Schütze K, Yachdav G, Ashkenazy H (2021) PredictProtein-predicting protein structure and function for 29 years. Nucleic Acids Res 49:W535–W540
Classen DC, Morningstar JM, Shanley JD (1987) Detection of antibody to murine cytomegalovirus by enzyme-linked immunosorbent and indirect immunofluorescence assays. Journal of Clinical Microbiology 25:600–4. https://doi.org/10.1128/jcm.25.4.600-604.1987
Criollo-Joaquin M, Motte E, Salvatierra M, Medina J, Diringer B, Sandoval G, Mialhe E (2019) Design and evaluation of the expression of a potential vaccine of DNA against lake tilapia virus (TiLV). Rev Peru Biol 26:301–310
Dixon PF, Hill BJ (1984) Rapid detection of fish rhabdoviruses by the enzyme-linked immunosorbent assay (ELISA). Aquaculture 42:1–12
Dong HT, Ataguba GA, Khunrae P, Rattanarojpong T, Senapin S (2017a) Evidence of TiLV infection in tilapia hatcheries from 2012 to 2017 reveals probable global spread of the disease. Aquaculture 479:579–583. https://doi.org/10.1016/j.aquaculture.2017.06.035
Dong HT, Siriroob S, Meemetta W, Santimanawong W, Gangnonngiw W, Pirarat N, Khunrae P, Rattanarojpong T, Vanichviriyakit R, Senapin S (2017b) Emergence of tilapia lake virus in Thailand and an alternative semi-nested RT-PCR for detection. Aquaculture 476:111–118. https://doi.org/10.1016/j.aquaculture.2017.04.019
Eichelberger MC, Morens DM, Taubenberger JK (2018) Neuraminidase as an influenza vaccine antigen: a low hanging fruit, ready for picking to improve vaccine effectiveness. Curr Opin Immunol 53:38–44. https://doi.org/10.1016/j.coi.2018.03.025
Eyngor M, Zamostiano R, Tsofack JEK, Berkowitz A, Bercovier H, Tinman S, Lev M, Hurvitz A, Galeotti M, Bacharach E, Eldar A (2014) Identification of a novel RNA virus lethal to tilapia. J Clin Microbiol 52:4137–4146
FAO (2018) The State of World Fisheries and Aquaculture 2018 (SOFIA). Meeting the sustainable development goals Rome. Available from: https://www.fao.org/documents/card/en?details=I9540EN. Accessed 14 Aug 2023
Fathi M, Dickson C, Dickson M, Leschen W, Baily J, Muir F, Ulrich K, Weidmann M (2017) Identification of tilapia lake virus in Egypt in Nile tilapia affected by ‘summer mortality’ syndrome. Aquaculture 473:430–432. https://doi.org/10.1016/j.aquaculture.2017.03.014
Ferguson HW, Kabuusu R, Beltran S, Reyes E, Lince JA, Del Pozo J (2014) Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): a case report. J Fish Dis 37(6):583–589. https://doi.org/10.1111/jfd.12142
Ghaffari AD, Dalimi A, Ghaffarifar F, Pirestani M (2020) Structural predication and antigenic analysis of ROP16 protein utilising immunoinformatics methods in order to identification of a vaccine against Toxoplasma gondii: an in silico approach. Microb Pathog 142:104079. https://doi.org/10.1016/j.micpath.2020.104079
Goldman E, Rosenberg AH, Zubay G, Studier WF (1995) Consecutive low-usage leucine codons block translation only when near the 5′ end of a message in Escherichia coli. J Mol Biol 245:467–473. https://doi.org/10.1006/jmbi.1994.0038
Gupta P, Waheed SM, Bhatnagar R (1999) Expression and purification of the recombinant protective antigen of Bacillus anthracis. Protein Expr Purif 16:369–376
Hasan A, Hossain M, Alam J (2013) A computational assay to design an epitope-based peptide vaccine against Saint Louis encephalitis virus. Bioinform Biol Insights 7:BBI-S13402. https://doi.org/10.4137/BBI.S13402
Hopp TP, Woods KR (1981) Prediction of protein antigenic determinants from amino acid sequences. Proc Natl Acad Sci 78:3824–3828. https://doi.org/10.1073/pnas.78.6.3824
Hu H, Zeng W, Wang Y, Wang Q, Bergmann SM, Yin J, Li Y, Chen X, Gao C, Zhang D, Liu C (2020) Development and application of a recombinant protein-based indirect ELISA for detection of anti-tilapia lake virus IgM in sera from tilapia. Aquaculture 520:734756. https://doi.org/10.1016/j.aquaculture.2019.734756
International Committee on the Taxonomy of Viruses (ICTV) (2018) Virus taxonomy: 2018b Release. Available from: https://ictv.global/taxonomy/taxondetails?taxnode_id=202206025&taxon_name=Amnoonviridae
Jansen MD, Dong HT, Mohan CV (2018) Tilapia lake virus: a threat to the global tilapia industry? Rev Aquac. https://doi.org/10.1111/raq.12254
Kane JF (1995) Effects of rare codon clusters on high-level expression of heterologous proteins in Escherichia coli. Curr Opin Biotechnol 6:494–500. https://doi.org/10.1016/0958-1669(95)80082-4
Kathwate GH (2022) In silico design and characterisation of multi-epitopes vaccine for SARS-CoV2 from its spike protein. Int J Pept Res Ther 28:1–15. https://doi.org/10.1007/s10989-021-10348-z
KembouTsofack JE, Zamostiano R, Watted S, Berkowitz A, Rosenbluth E, Mishra N, Briese T, Lipkin WI, Kabuusu RM, Ferguson H, Del Pozo J (2017) Detection of tilapia lake virus in clinical samples by culturing and nested reverse transcription-PCR. J Clin Microbiol 55:759–767. https://doi.org/10.1128/JCM.01808-16
Kohl TO, Ascoli CA (2017) Indirect immunometric ELISA. Cold Spring Harb Protoc 2017:396–402. https://doi.org/10.1101/pdb.prot093708
Kolaskar AS, Tongaonkar PC (1990) A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett 276:172–174. https://doi.org/10.1016/0014-5793(90)80535-Q
Koren CWR, Nylund A (1997) Morphology and morphogenesis of infectious salmon anaemia virus replicating in the endothelium of Atlantic salmon Salmo salar. Dis Aquat Org 29:99–109. https://doi.org/10.3354/dao029099
Kumari R, Kole S, Soman P, Rathore G, Tripathi G, Makesh M, Rajendran KV, Bedekar MK (2018) Bicistronic DNA vaccine against Edwardsiella tarda infection in Labeo rohita: construction and comparative evaluation of its protective efficacy against monocistronic DNA vaccine. Aquaculture 485:201–209. https://doi.org/10.1016/j.aquaculture.2017.11.052
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227(5259):680–685
Lin AV (2015). Indirect Elisa. ELISA: methods and protocols, 51–59. https://doi.org/10.1007/978-1-4939-2742-5_5
Lueangyangyuen A, Senapin S, Dong HT, Unajak S, Wangkahart E, Khunrae P (2022) Expression and purification of S5196–272 and S6200–317 proteins from tilapia lake virus (TiLV) and their potential use as vaccines. Protein Expr Purif 190:106013
Leya T, Ahmad I, Valappil RK, Kurcheti PP, Tripathi G, Sharma R, Bedekar MK (2021) Development of species-specific IgM antibodies and elevation of mucosal immune response in Labeo rohita using recombinant bicistronic nano DNA vaccine priming. Fish & Shellfish Immunol 113:185–95. https://doi.org/10.1016/j.fsi.2021.04.008
Mauro VP (2018) Codon optimization in the production of recombinant biotherapeutics: potential risks and considerations. BioDrugs 32:69–81. https://doi.org/10.1007/s40259-018-0261-x
Mikalsen AB, Sindre H, Torgersen J, Rimstad E (2005) Protective effects of a DNA vaccine expressing the infectious salmon anemia virus hemagglutinin- esterase in Atlantic salmon. Vaccine 23(41):4895–4905. https://doi.org/10.1016/j.vaccine.2005.05.025
Nicholson P, Fathi MA, Fischer A, Mohan C, Schieck E, Mishra N, Heinimann A, Frey J, Wieland B, Jores J (2017) Detection of tilapia lake virus in Egyptian fish farms experiencing high mortalities in 2015. J Fish Dis 40(12):1925–1928. https://doi.org/10.1111/jfd.12650
Olesen NJ, Jørgensen PEV (1991) Rapid detection of viral haemorrhagic septicaemia virus in fish by ELISA. J Appl Ichthyol 7(3):183–186
Papaleo E, Saladino G, Lambrughi M, Lindorff-Larsen K, Gervasio FL, Nussinov R (2016) The role of protein loops and linkers in conformational dynamics and allostery. Chem Rev 116:6391–6423
Pulido LLH, Mora CM, Hung AL, Dong HT, Senapin S (2019) Tilapia lake virus (TiLV) from Peru is genetically close to the Israeli isolates. Aquaculture 510:61–65. https://doi.org/10.1016/j.aquaculture.2019.04.058
Rajendran KV, Sood N, Rao BM, Valsalam A, Bedekar MK, Jeena K, Pradhan PK, Paria A, Swaminathan TR, Verma DK, Sood NK (2023) Widespread occurrence of tilapia parvovirus in farmed Nile tilapia Oreochromis niloticus from India. J Fish Dis. https://doi.org/10.1111/jfd.13871
Rodiansyah A, Tan MI, Nugrahapraja H (2022) Construction, cloning, and overexpression of Staphylococcal Enterotoxin B Gene Synthetic (SEBsyn) in pET-28a (+): Pre-development Bacterial-Toxin Therapy for Cancer. In 7th International Conference on Biological Science (ICBS 2021) (pp. 464–470). Series: Advances in Biological Sciences Research, Atlantis Press. ISSN: 2468–5747. https://doi.org/10.2991/absr.k.220406.065
Sahdev S, Khattar SK, Saini KS (2008) Production of active eukaryotic proteins through bacterial expression systems: a review of the existing biotechnology strategies. Mol Cell Biochem 307:249–264. https://doi.org/10.1007/s11010-007-9603-6
Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, Lopez R, McWilliam H, Remmert M, Söding J, Thompson JD (2011) Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol 7:539. https://doi.org/10.1038/msb.2011.75
Subramaniam K, Ferguson HW, Kabuusu R, Waltzek TB (2019) Genome sequence of tilapia lake virus associated with syncytial hepatitis of tilapia in an Ecuadorian aquaculture facility. Microbiol Resour Announc 8:e00084-e119
Surachetpong W, Janetanakit T, Nonthabenjawan N, Tattiyapong P, Sirikanchana K, Amonsin A (2017) Outbreaks of tilapia lake virus infection, Thailand, 2015–2016. Emerg Infect Dis 23:1031. https://doi.org/10.3201/eid2306.161278
Tattiyapong P, Dechavichitlead W, Waltzek TB, Surachetpong W (2020) Tilapia develop protective immunity including a humoral response following exposure to tilapia lake virus. Fish Shellfish Immunol 106:666–674
United Nations Department of Economic and Social Affairs Population Division (2019) World population prospects 2019: Highlights. United Nations. Retrieved from from: https://population.un.org/wpp/Publications/Files/WPP2019_Highlights.pdf
Valsalam A, Rajendran KV, Kezhedath J, Godavarikar A, Sood N, Bedekar MK (2023) Development of an indirect ELISA test for the detection of tilapia lake virus (TiLV) in fish tissue and mucus samples. Microb Pathog 315:114707
Valsalam A, Bedekar MK, Kezhedath J, Sood N, Poojary N, Namdeo MS, Shrivastava N, Rajendran KV (2024) Isolation, in vitro, and in vivo pathogenicity test of tilapia lake virus (TiLV) and development of a prognostic semi-quantitative lesion scoring system for differentiating clinical/subclinical infection in farmed tilapia (Oreochromis niloticus L.). Microb Pathog 186:106475
Yadava A, Ockenhouse CF (2003) Effect of codon optimization on expression levels of a functionally folded malaria vaccine candidate in prokaryotic and eukaryotic expression systems. Infect Immun 71:4961–4969. https://doi.org/10.1128/IAI.71.9.4961-4969.2003
Zeng W, Wang Y, Chen X, Wang Q, Bergmann SM, Yang Y, Wang Y, Li BO, Lv Y, Li H, Lan W (2021) Potency and efficacy of VP20-based vaccine against tilapia lake virus using different prime-boost vaccination regimens in tilapia. Aquaculture 539:736654
Acknowledgements
The authors acknowledge the Director, ICAR-Central Institute of Fisheries Education, Mumbai, India, for providing all the essential facilities and the DBT, GOI.
Funding
The funding was provided by ICAR-Central Institute of Fisheries Education and Department of Biotechnology (BT/PR30609/AAQ/03/939/2018), GOI, India.
Author information
Authors and Affiliations
Contributions
LF- bench works and writing, data compilation. MKB- Conceptualization, data analysis and interpretation. AG- bench works, data compilation and writing. AV- Methodology, data compilation; Writing- original draft preparation. GBP- Reviewing and Editing. RKV-Monitoring; Writing-Correction.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Handling editor: Brian Austin
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lalruatfela, Bedekar, M.K., Godavarikar, A. et al. Molecular cloning and expression of codon-optimized segment 4 hypothetical protein (35 kDa) of tilapia lake virus (TiLV) in pET-28a( +) expression vector and development of indirect ELISA test. Aquacult Int 32, 5997–6015 (2024). https://doi.org/10.1007/s10499-024-01452-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-024-01452-0