
Abstract
Nine semi-purified diets with three different levels of protein (350, 400, and 450 g/kg diet) and lipid (80, 120 and 160 g/kg diet) were fed ad libitum to Mystus gulio fry (0.16 g) in triplicate groups (20 fish/replicate) for 90 days. Flow-through fiber-reinforced tanks (50 L; water flow rate of 0.5 L/min) were used for rearing the fish. Result of the experiment showed that the dietary protein, lipid, and their interactions affect significantly (P < 0.05) the weight gain, specific growth rate (SGR), protein efficiency ratio (PER), protein productive value (PPV), energy productive value (EPV), and feed conversion ratio (FCR). Among all the dietary treatment groups, the fish fed diet containing 400 g protein and 120 g lipid/kg resulted in significantly higher (P < 0.05) weight gain (4.62 g), SGR (3.77%/day), PER (1.52), PPV (23.39%), and EPV (64.74%) and lower FCR (1.64). The whole-body protein (15.28%) and lipid (5.33%) contents were significantly higher (P < 0.05) in fish fed diets containing 400 g protein and 120 g lipid/kg diet and 350 g protein and 160 g lipid/kg diet, respectively. Second-order polynomial regression analysis indicates that the optimum protein and lipid requirements of M. gulio fry are 409–411 g protein/kg diet and 122–126 g lipid/kg diet, respectively.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Feed constitutes about 60–70% of the recurring cost in aquaculture. One of the major problems faced by fish growers around the globe is the non-availability of nutritionally balanced, cost-effective feeds. Determination of nutrient requirement is essential for formulating the quality feeds for various life stages of farmed fish (Gatlin 1995; Tucker 1998; NRC 2011). Protein is the most vital and costly nutrient, which primarily determines the feed cost. It provides both essential and non-essential amino acids, which are required for synthesis of body protein, growth, cell maintenance, development, and tissue repair of fish. The inadequate protein level in fish diets leads to poor growth, whereas the excessive dietary protein increases the cost of production and more ammonia excretion (NRC 2011). Dietary lipids are considered an important source of energy and essential fatty acids. Lipids assist in absorption of sterols and fat-soluble vitamins, serve as a source of phospholipids, and participate in the synthesis of hormones, prostaglandins, and other metabolically active compounds, which are required for the normal growth and reproduction of fish (Lovell 1989; Sargent et al. 2002; Lim and Webster 2006; Lim and Aksoy 2009; NRC 2011). Supplementation of feed with lipid is more effective than the protein or carbohydrate because lipid is an energy-dense nutrient and easily metabolized by carnivorous fish (NRC 1993). However, excessive lipid in diets might lead to decrease feed consumption and reduce growth and production of fatty fish (Hanley 1991; Sargent et al. 2002).
Long whiskers catfish, Mystus gulio, is primarily a brackish water catfish, which is also cultured in freshwater after suitable acclimatization. It is distributed along the coast of Bangladesh, India, Sri Lanka, Indonesia, Vietnam, Myanmar, Pakistan, Java, Thailand, and Malaysia (Day 1878; Kumar et al. 2019). In India, it is commonly found in the Gangetic estuary, Chilika Lake, and brackish waters of Kerala (Talwar and Jhingran 1991). The culture of this species is gaining importance in India and Bangladesh due to its high market demand and consumer preference. It is an important candidate species for aquaculture diversification because of its hardy nature, delicious taste, excellent nutritional value, and high market demand (Tripathi 1996; Kumar et al. 2019). Mystus gulio is also considered an ornamental fish with an excellent export market potential. It thrives well in oxygen-depleted water, tolerates crowding conditions, and easily accepts artificial feeds (Ali et al. 2014). This species also has great potential in modern aquaculture practices of raceway culture, recirculatory aquaculture system, and biofloc culture. Ample literature is available on reproductive biology, breeding, and seed production of M. gulio in India and elsewhere in the world. However, there is very limited information available on the nutrient requirement of this species particularly for its early stage. Therefore, the objective of the present study is to evaluate the optimal dietary protein and lipid requirements of M. gulio fry. The results of the current work will help in formulating the nutritionally balanced cost-effective nursery diets, which is essentially required for quality seed production and aquaculture of this species.
Materials and methods
Preparation of experimental diets
Nine semi-purified experimental diets (D-1 to D-9) containing 350, 400, and 450 g protein (Pr)/kg diet and 80, 120, and 160 g lipid (Li)/kg diet were formulated (Table 1). Casein and gelatin served as protein sources, dextrin and corn starch as carbohydrate sources, and equal proportions (1:1) of sunflower oil and fish oil as lipid sources. α-Cellulose was used as a filler and carboxymethyl cellulose (CMC) was added as binder. The diets were fortified with prepared vitamin and mineral mixture (Ogino 1977; Modified Lovell et al. 1984). For preparation of diets, the dry feed components were homogeneously blended, moistened with lukewarm water, and then mixed to form a stiff dough. The dough was forced through the die of a pelletizer and the resulting strands were initially air-dried and then dried at 60 °C in an air oven for 24 h. The dried strands were crumbled to pellets of suitable size and then stored in air-tight containers in a refrigerator until used. Feeds were prepared every 15 days using ingredients purchased from the market as a single batch.
Experimental design and fish maintenance
M. gulio fry (two thousand) were procured from the ICAR-Central Institute of Brackishwater Aquaculture (ICAR-CIBA), Kakdwip, West Bengal, India. The fish were acclimatized to laboratory condition in five FRP tanks (200 L) fitted with a flow-through system for the period of 2 weeks. During acclimatization, the fish were fed twice daily with a semi-purified diet (300 g/kg protein; 60 g/kg lipid) at 10% of their body weight. After acclimatization, 540 uniform-sized fry (average initial weight of 0.16 ± 0.00 g) were randomly distributed to 27 flow-through FRP tanks (50 L) in triplicates at a stocking density of 20 fish/tank. The stocked fish were batch weighed to determine the initial fish biomass in each tank. Throughout the experiment, the water volume of 40 L and the flow rate of 0.5 L/min were maintained in each experimental tank. The experiment was conducted under natural photoperiod (12-h light:12-h dark). Seasoned and well-aerated ground water was used for the experiment. All groups of fish were fed ad libitum two times a day (09:00 and 16:00 hours) for 90 days. Initially, for the first month, the feeding rate was maintained at 10% of the body weight of fish, which was subsequently reduced to 6–8% for next 2 months of the culture period depending on the feed intake. The feed was provided in two equal split dosages. After 2 h of feeding, the excreta of the fish was removed carefully from each tank by siphoning and discarded outside. Then, the unconsumed feed present if any was removed from each tank, oven-dried, and stored in separate air-tight plastic containers at 4 °C. At the end of the experiment, the fish from each rearing tank were batch weighed and the final average weight of fish was determined.
Proximate analysis of diet and fish
Before starting the experiment, 500 fish were killed with an overdose of MS 222 and the triplicate pooled samples were taken to determine initial whole-body composition. At the end of the experiment, all the 60 fishes of each treatment (20 fish/replicate × 3 replicates) were pooled separately for final whole-body composition analysis. Following the institute’s animal ethical procedure, live fish samples were killed with overdose of MS222. The killed fish were oven-dried and ground for proximate analysis in triplicates. The proximate composition of experimental diets and fish whole body were analyzed following the standard procedure (AOAC 1990). Dry matter was estimated by oven-drying the samples at 105 °C to a constant weight, nitrogen content was measured using a micro-Kjeldahl method, and crude protein was estimated by multiplying nitrogen content with a factor 6.25. The ether extract was determined by solvent extraction with petroleum ether, boiling point 40–60 °C, for 10–12 h. Total ash content of the sample was determined by incinerating the sample at 650 °C for 6 h. Gross energy (GE) of the diets and fish whole body was estimated by using a Bomb Calorimeter (Parr, model 1341; Parr Instrument Company, Moline, IL, USA). Proximate composition of the experimental diets is presented in Table 1.
Water analyses
The water quality parameters such as temperature, pH, DO, total alkalinity, total hardness, and total ammonia of the experimental tanks were analyzed once in every 15-day interval following the standard protocol (APHA 1992). The recorded values were as follows: temperature, 26.5–28.3 °C; pH, 7.6–7.8; dissolved oxygen, 6.7–7.2 mg/L; total alkalinity, 112.56–117.29 mg CaCO3/L; and total hardness,105.13–108.96 mg CaCO3/L.
Calculation of nutritional indices
The different nutritional indices were determined as follows:

Statistical analysis
Data are presented as mean ± standard error (SE). Statistical significance of data was analyzed following one-way and two-way analysis of variance (ANOVA) and Duncan’s multiple range test (DMRT) at P < 0.05. The statistical package used for data analysis was PC-SAS program for Windows, released v6.12 (SAS Institute, Cary, NC, USA (SAS 1996)). One-way ANOVA and DMRT were performed to assess the individual effect of dietary protein and lipid and the two-way ANOVA for the interactive effect of dietary protein and lipid on measured growth and nutritional parameters. Data are expressed as mean ± SE (n=3) for each Pr and Li combination (D-1 to D-9). But for the individual effect of Pr (350, 400, and 450 g/kg diet) and Li (80, 120, and 160 g/kg diet), the data are expressed as mean ± SE (n=9). The relationships between weight gain, FCR, and dietary protein and lipid levels were investigated using second-order polynomial regression models and the optimum protein and lipid requirement levels were estimated.
Results
At the end of the experiment, a survival rate of 100% was recorded in each experimental tank. It was observed that the dietary protein and lipid individually and in combination had a significant effect (P < 0.05) on growth performance and feed utilization of fish (Table 2). The weight gain, SGR, FCR, and PER were significantly better (P < 0.05) in fish fed 400 g protein/kg diet and 120 g lipid/kg diet (D-5) than in the other protein and lipid fed groups.
Feeding fish with different levels of protein and lipid significantly (P < 0.05) influenced the whole-body moisture, protein, lipid, ash, and energy contents of the fish (Table 3). The whole-body moisture content was increased significantly (P < 0.05) with an increase in dietary protein level but it was decreased (P < 0.05) with increase in dietary lipid level. The whole-body protein content was increased significantly (P < 0.05) with an increase in both dietary protein and lipid levels up to 400 g and 120 g/kg diet, beyond which there was no further improvement. In contrast to whole-body moisture content, the whole-body lipid content was significantly increased (P < 0.05) with increase in dietary lipid and decreased (P < 0.05) with increase in dietary protein content. However, the whole-body ash content was significantly increased (P < 0.05) with increase in both dietary protein and lipid levels. But, the gross energy content was significantly decreased (p < 0.05) with increase in dietary protein.
The effects of dietary protein and lipid levels on nutrient retention in M. gulio fry are presented in Table 4. Both the dietary protein and lipid individually and their combinations had significant effects (P < 0.05) on PPV, LPV, and EPV. The significantly higher (P < 0.05) PPV, LPV, and EPV values were obtained in fish fed 400 g protein/kg diet (medium level of dietary protein supplementation). Though significantly higher (P < 0.05) PPV and EPV were obtained in fish fed 120 g lipid/kg diet (medium levels of dietary lipid supplementation), the significantly higher (P < 0.05) LPV was found in 80 g lipid/kg diet fed group (lowest level of dietary lipid supplementation). Similar to the individual effects, the interactive effects of dietary protein and lipid also had significantly higher (P < 0.05) PPV and EPV in fish fed diet containing 400 g protein/kg and 120 g lipid/kg diets (D-5). However, the significantly higher (P < 0.05) LPV was recorded in fish fed 400 g protein/kg and 80 g lipid/kg diets (D-4).
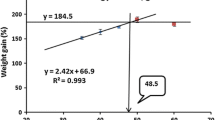
The second-order polynomial regression analysis of weight gain and FCR was used to determine the exact protein and lipid requirements of the M. gulio fry. By keeping the dietary lipid level at 120 g/kg diet (optimum), the exact protein requirement was found to be 409 g/kg diet (y = −0.0383x2 + 3.1363x − 59.5 with R2 = 0.933, where y = weight gain and x = dietary protein level) (Fig. 1) and 411 g/kg diet (y = 0.0105x2 − 0.8633x + 19.32 with R2 = 0.9257, where y = FCR and x = dietary protein level) (Fig. 2). Similarly, by keeping the dietary protein level 400 g/kg diet (optimum), the exact lipid requirement was estimated to be 126 g/kg diet (y = −0.0757x2 + 1.9121x − 7.42 with R2 = 0.9628, where y = weight gain and x = dietary lipid level) (Fig. 3) and 122 g/kg diet (y = 0.0181x2 − 0.4425x + 4.34 with R2 = 0.9274, where y = FCR and x = dietary lipid level) (Fig. 4). Therefore, from the second-order polynomial regression analysis, the exact protein and lipid requirements of M. gulio fry were found to be 409–411 g protein/kg diet and the 122–126 g lipid/kg diet, respectively.

Second-order polynomial regression of weight gain and various dietary protein levels for M. gulio fry with 120 g lipid/kg diet

Second-order polynomial regression of FCR and various dietary protein levels for M. gulio fry with 120 g lipid/kg diet

Second-order polynomial regression of weight gain and various dietary lipid levels for M. gulio fry with 400 g protein/kg diet

Second-order polynomial regression of FCR and various dietary lipid levels for M. gulio fry with 400 g protein/kg diet
Discussion
In the present study, we observed significantly higher weight gain, SGR, and PER and lower FCR of M. gulio fry at the medium protein (400 g/kg) and lipid (120 g/kg) fed group than at the lower or higher protein and lipid fed groups. It is reported by many earlier researchers that the fish fed with lower protein diet did not meet its requirement, which resulted poor nutrient gain and growth of fish, whereas the excess dietary protein is deaminated leading to deterioration of water quality (Winfree and Stickney 1981; Huang et al. 2019) and catabolized to provide energy for maintenance, thus reducing the protein conversion efficiency in fish (Lee and Putnam 1973; Mohanta et al. 2008). When the fish is fed with a diet containing less amount of protein than its optimum requirement, it needs to consume more quantity of feeds to make up the protein required for its growth and metabolism, and at high protein levels, less feed is required, and therefore, in both the cases, there is an imbalance between protein and energy leading to poor growth of fish (Rahimnejad et al. 2015 for juvenile hybrid grouper, Epinephelus fuscoguttatus × E. lanceolatus). This may be a reason for the poor growth (weight gain, SGR) and feed and nutrient utilization (FCR and PER) of M. gulio fry in both low (350 g/kg)- and high (450 g/kg)-protein diet groups. Similarly, it is reported that the high level of dietary lipid is attributed to loss of appetite and poor feed intake, which prevents the fish from taking enough nutrients (mainly the protein) to meet its optimum requirement, ultimately leading to less protein synthesis and tissue growth. On the contrary, when the dietary lipid is provided less than its requirement, the fish utilizes protein as energy source, which results in poor growth of fish and also increases the production cost (Hanley 1991; Sargent et al. 2002; Chatzifotis et al. 2010; NRC 2011). The above facts explain the poor growth, nutrient utilization, and nutrient gain of M. gulio fry in both high (160 g lipid/kg)- and low (80 g lipid/kg)-lipid diets.
The results on whole-body chemical composition of M. gulio fry indicated that the moisture content is directly proportional to dietary protein level and inversely proportional to dietary lipid level. This may be the reason for the maximum whole-body moisture content of M. gulio fry fed diet D-7 containing 450 g protein and 80 g lipid/kg diet. Similar to our findings, other researchers in their studies also reported an inverse relation between dietary lipid and the whole-body moisture content of fish (Babalola and Apata 2006 for African catfish, Heterobranchus longifilis; Lee et al. 2002 for juvenile ayu, Plecoglossus altivelis) and the direct relation between dietary protein and whole-body moisture contents (Mohanta et al. 2008). We observed maximum whole-body protein content in medium dietary protein and lipid fed group (D-5; 400 g protein and 120 g lipid/kg diet) beyond which there was no further improvement, which is similar to the results of Arslan et al. (2013) for South American catfish, surubim (Pseudoplatystoma sp.), and Zhang et al. (2016) for topmouth culter (Culter alburnus). This amply justified the higher weight gain of fish in the D-5 group than in other diet fed groups. In the current study, it is observed that the whole-body lipid content of fish and dietary lipid level are directly correlated, which is similar to the earlier researchers (Salhi et al. 2004 in black catfish, Rhamdia quelen, fry; Ali and Jauncey 2005 in African catfish, Clarias gariepinus; Zhang et al. 2016 in topmouth culter, Culter alburnus). But the dietary protein and the whole-body lipid are inversely related in M. gulio, which in agreement with Babalola and Apata (2006) in African catfish, Heterobranchus longifilis. This could be the reason for the maximum whole-body lipid content of M. gulio fry fed with D-3 diet containing lowest level of protein and highest level of lipid (350 g protein and 160 g lipid/kg diet). In this study, the whole-body ash content was increased linearly with the increase in both dietary protein and lipid levels. Therefore, maximum ash content was found in fish fed with D-9 containing 450 g protein and 160 g lipid/kg diet. Opposite to the current finding, it is reported that there is no variation in whole-body ash content of C. gariepinus due to change in dietary protein and lipid concentrations (Ahmad 2008). There was a marginal decrease in whole-body gross energy content of M. gulio fry with the increase in both dietary protein and lipid contents.
In the present study, the higher growth and nutrient utilization were recorded in M. gulio fry fed diet containing 400 g protein/kg and 120 g lipid/kg diet (D-5). This is commensurate with the higher PPV and EPV in this group as compared to other dietary protein and lipid fed groups. However, further increase in dietary protein and lipid beyond D-5 level does not improve the nutrient gain (PPV, LPV, and EPV), which agrees with the findings of earlier researchers (Li et al. 2010 in blunt snout bream, M. amblycephala; Paul et al. 2010 in Puntius gonionotus; Babalola and Apata 2006 in African catfish, H. longifilis). The proper balance between dietary protein and non-protein energy is essential for the better nutrient utilization and growth of fish (Schulz et al. 2008). An inadequate dietary protein to energy ratio may result in slower growth and low protein and energy utilization (Ai et al. 2004). Therefore, the M. gulio fry fed either higher or lower protein and/or lipid than its optimum requirement levels (400 g protein/kg and 120 g lipid/kg diet) might have an imbalance dietary protein to non-protein energy ratios which led to its poor growth and nutrient utilization.
Generally, an increase of dietary lipid level and the concomitant decrease in dietary protein level resulted in protein-sparing effect in many fish species and this effect is more prevalent at the sub-optimum level of dietary protein and a higher level of lipid (Tibbetts et al. 2005; Schulz et al. 2007). In the current study, protein-sparing effect of lipid was not observed as the best growth and nutrient utilization of M. gulio were recorded in the medium dietary protein (400 g protein/kg) and lipid (120 g lipid/kg) diet rather than the low-protein (350 g protein/kg) and high-lipid (160 g lipid/kg) diet. However, the protein-sparing effect has been documented in other catfish species such as bagrid catfish Pseudobagrus fulvidraco (Kim and Lee 2005) and African catfish C. gariepinus (Ali and Jauncey 2005).
Comparatively better growth and nutrient utilization were observed in M. gulio fry fed the diet having Pr/GE ratio of 20.31 g/MJ with a protein and lipid levels of 400 g/kg and 120 g/kg diet (D-5). The diets (D-3 and D-6) containing a high GE level of 19.23 MJ/kg had resulted in high lipid gain (LPV). It is reported that a proper balance between dietary Pr and GE is necessary to maintain high growth rate, good food conversion efficiency, and better protein utilization in fish. This also helps to minimize the excessive accumulation of lipid and glycogen in somatic tissue and liver, decrease the undesirable nitrogen waste output, and improve the quality of fish farm effluents (Tibbetts et al. 2005). The Pr/E ratios for optimum growth of several fish species ranged from 19 to 27 g/MJ (NRC 1993). The optimum dietary Pr/GE ratio (g protein/MJ) has been reported for channel catfish Ictalurus punctatus is 20.81 (Garling and Wilson 1976) which is similar to our findings. Contrary to the present finding, a higher Pr/GE ratio of 23.6 g/MJ is reported in Rhamdia quelen (Salhi et al. 2004).
Conclusions
From the results of the experiment, it is found that the dietary protein and lipid levels of 400 g and 120 g/kg diet, respectively, are adequate to realize the maximum growth potential of M. gulio fry under the given experimental condition. However, from the second-order polynomial regression analysis, it is concluded that the exact protein and lipid requirements of M. gulio fry are 409–411 g protein/kg diet and the 122–126 g lipid/ kg diet, respectively. This basic knowledge on optimum dietary protein and lipid requirement levels will be helpful in formulating the cost-effective and environment-friendly nursery diets for the culture of M. gulio fry.
Data availability
It is declared that all data and materials as well as software application or custom code support our claims and comply with field standards.
The data that support the findings of the submitted manuscript are available from the corresponding author upon reasonable request.
References
Ahmad MH (2008) Response of African catfish Clarias gariepinus to different dietary protein and lipid levels in practical diets. J World Aquacult Soc 39(4):541–548
Ai Q, Mai K, Li H, Zhang C, Zhang L, Duan Q, Tan B, Xu W, Ma H, Zhang W, Liufu Z (2004) Effect of dietary protein to energy ratios on growth and body composition of juvenile Japanese seabass, Lateolabrax japonicus. Aquaculture 230:507–516
Ali MZ, Jauncey K (2005) Approaches to optimizing dietary protein to energy ratio for African catfish Clarias gariepinus (Burchell, 1822). Aquac Nutr 11:95–101
Ali MR, Afzal M, Khan MF, Naqvi SMHM, Akhtar S (2014) Dietary protein requirement of giant river catfish Sperata seenghala (Sykes) determined using diets of varying protein level. Pak J Nutr 13:151–156
AOAC (1990) Association of Official Analytical Chemists. Official methods of analysis of the association of official analytical chemists, vol. 1, 14th edn. AOAC, Arlington, p 1102
APHA (1992) American Public Health Association. Standard methods for examination of water and wastewater, 17th edn. APHA, Washington DC
Arslan M, Dabrowski K, Ferrer S, Dietrich M, Rodriguez G (2013) Growth, body chemical composition and trypsin activity of south american catfish, surubim (Pseudoplatystoma sp.) juveniles fed different dietary protein and lipid levels. Aquac Res 44:760–771
Babalola TOO, Apata DF (2006) Effects of dietary protein and lipid levels on growth performance and body composition of african catfish Heterobranchus longifilis (Valenciennes, 1840) fingerlings. J Anim Vet Adv 12:1073–1079
Chatzifotis S, Panagiotidou M, Papaioannou N, Pavlidis M, Nengas I, Mylonas CC (2010) Effect of dietary lipid levels on growth, feed utilization, body composition and serum metabolites of meagre (Argyrosomus regius) juveniles. Aquaculture 307:65–70
Day F (1878) The fishes of India. Being a natural history of the fishes known to inhabit the seas and fresh waters of India, Burma and Ceylon. William Dowson and Sons, London, p 778
Garling DL, Wilson RP (1976) Optimum protein to energy ratio for channel catfish fingerlings Ictalurus punctatus. J Nutr 106:1368–1375
Gatlin DM (1995) Review of red drum nutrition. In: Lim CE, Sessa DJ (eds) Nutrition and utilization requirement of the red drum and relative use of dietary carbohydrate and lipid, pp 41–49
Hanley F (1991) Effects of feeding supplementary diets containing varying levels of lipid on growth, food conversion, and body composition of Nile tilapia Oreochromis niloticus (L.). Aquaculture 93:323–334
Huang QC, Qin DG, Tan BP, Du T, Yang YZ, Yang QH, Chi SY, Dong XH (2019) The optimal dietary protein level of juvenile silver sillago Sillago sihama at three dietary lipid levels. Aquac Res 51:816–827
Kim LO, Lee SM (2005) Effects of the dietary protein and lipid levels on growth and body composition of bagrid catfish Pseudobagrus fulvidraco. Aquaculture 243:323–329
Kumar P, Biswas G, Ghoshal TK, Kailasam M, Christina L, Vijayan KK (2019) Current knowledge on the biology, captive breeding and aquaculture of the brackishwater catfish Mystus gulio (Hamilton, 1822): a review. Aquaculture 499:243–250
Lee DJ, Putnam GB (1973) The response of rainbow trout to varying protein/energy ratios in a test diet. J Nutr 103:916–922
Lee SM, Kim DJ, Cho SH (2002) Effects of dietary protein and lipid level on growth and body composition of juvenile ayu Plecoglossus altivelis reared in seawater. Aquac Nutr 8:53–58
Li XF, Liu WB, Jiang YY, Zhu H, Ge XP (2010) Effects of dietary protein and lipid levels in practical diets on growth performance and body composition of blunt snout bream Megalobrama amblycephala fingerlings. Aquaculture 303:65–70
Lim CE, Aksoy M (2009) Lipid and fatty acid requirements of tilapia. Aquaculture America, Seattle, p 194
Lim CE, Webster CD (2006) Nutrient requirements. In: Lim CE, Webster CD (eds) Tilapia biology, culture and nutrition. Haworth Press, Binghamton, pp 469–501
Lovell T (1989) The Nutrients. In: Lovell T (ed) Nutrition and feeding of fish. Van Nostrand Reinhold, New York, pp 11–63
Lovell RT, Miyazaki T, Rebegnator S (1984) Requirements of α-tocopherol by channel catfish fed diets low in polyunsaturated triglycerides. J Nutr 114:894–901
Mohanta KN, Mohanty SN, Jena JK, Sahu NP (2008) Protein requirement of silver barb Puntius gonionotus fingerlings. Aquac Nutr 14:143–152
NRC (1993) Nutrient requirements of fish. National Academy Press, Washington, p 114
NRC (2011) Nutrient requirements of fish and shrimp. National Academy Press, Washington, DC. https://doi.org/10.17226/13039
Ogino C (1977) The present situation of studies on fish nutrition. Proceedings of the 7th Japan-Soviet joint symposium aquaculture, Tokyo, Japan, pp 11-18
Paul BN, Giri SS, Sarkar S, Mohanty SN, Sarangi N (2010) Effect of varying protein and lipid levels on the growth of medium carp Puntius gonionotus. Anim Nutr Feed Technol 10:81–86
Rahimnejad S, Bang IC, Park JY, Sade A, Choi J, Lee SM (2015) Effects of dietary protein and lipid levels on growth performance, feed utilization and body composition of juvenile hybrid grouper Epinephelus fuscoguttatus × E. lanceolatus. Aquaculture 446:283–289
Salhi M, Bessonart M, Chediak G, Bellagamba M, Carnevia D (2004) Growth, feed utilization and body composition of black catfish Rhamdia quelen fry fed diets containing different protein and energy levels. Aquaculture 231:435–444
Sargent JR, Tocher DR, Bell JG (2002) The lipids. In: Halver JE, Hardy RW (eds) Fish nutrition, 3rd edn. Academic Press, San Diego, pp 181–257
SAS (1996) PC-SAS Programme for windows, release v6.12. SAS institute Inc, Cary
Schulz C, Bohm M, Wirth M, Rennert B (2007) Effect of dietary protein on growth, feed conversion, body composition and survival of pike perch fingerlings Sander lucioperca. Aquac Nutr 13:373–380
Schulz C, Huber M, Ogunji J, Rennert B (2008) Effects of varying dietary protein to lipid ratios on growth performance and body composition of juvenile pike perch (Sander lucioperca). Aquac Nutr 14:166–173
Talwar PK, Jhingran AG (1991) Inland fishes of India and adjacent countries. Vol. 1 and Vol. 2. Oxford and IBH Publishing Co. Pvt. Ltd, New Delhi, Bombay and Calcutta, p 1063
Tibbetts SM, Lall SP, Milley JE (2005) Effects of dietary protein and lipid levels and (DP/DE) ratio on growth, feed utilization and hepatosomatic index of juvenile haddock Melanogrammus aeglefinus. Aquac Nutr 11:67–75
Tripathi SD (1996) Present status of breeding and culture of catfishes in South Asia. Aquat Living Resour 9(S1):219–228
Tucker JW (1998) Nutrition of larval fish. In: Tucker JR, John W (eds) Marine fish culture. Springer, Boston, pp 375–467
Winfree RA, Stickney RR (1981) Effects of dietary protein and energy on growth, feed conversion efficiency and body composition of Tilapia aurea. J Nutr 111:1001–1002
Zhang YL, Song L, Liu RP, Zhao ZB, He H, Fan QX, Shen ZG (2016) Effects of dietary protein and lipid levels on growth, body composition and flesh quality of juvenile topmouth culter Culter alburnus Basilewsky. Aquac Res 8:2633–2641
Acknowledgements
The authors are thankful to the Director, ICAR-Central Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar, India, for extending all the infrastructure facilities needed for the experiment.
Code availability (software application or custom code) As indicated in the Availability of data and material (data transparency)
Funding
Not applicable
Author information
Authors and Affiliations
Contributions
It is declared that all authors whose names appear on the submission
(1) Made substantial contributions to the conception or design of the work; or the acquisition, analysis, or interpretation of data; or the creation of new software used in the work;
(2) Drafted the work or revised it critically for important intellectual content;
(3) Approved the version to be published; and
(4)Agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
1 Rooprekha Khatua: Procurement and analysis of feed ingredients and experimental feeds, fish samples, experimental diet preparation, and feeding of fish. 2 Kedar Nath Mohanta: Experimental design, data representation, and manuscript preparation. 3 Nitish Kumar Chandan: Statistical analysis. 4 Rojalin Pattanayak: Experimental design and feed formulation. 5 Choudhury Suryakant Mishra: Collection of literature and editing of manuscript. 6 Prem Kumar: Procurement of experimental fish, acclimatization to laboratory condition, and fish maintenance
Corresponding author
Ethics declarations
Ethics approval
It is declared that:
1) The manuscript is not submitted to any journal for simultaneous consideration.
2) The submitted work is original and not have been published elsewhere in any form or language (partially or in full), unless the new work concerns an expansion of previous work.
3) The present study is not split up into several parts to increase the quantity of submissions and submitted to various journals or to one journal over time (i.e., “salami-slicing/publishing”).
4) Results are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation (including image-based manipulation).
5) The study adhered to the ICAR Guidelines for Internal Evaluation and Forwarding Research Papers to Scientific Journals and Data Management in ICAR Institutes, 2014, Indian Council of Agricultural Research, New Delhi, and the prior permission was obtained from PME, ICAR-CIFA, Bhubaneswar, prior to the start of the study.
Consent to participate
We all the authors do hereby declared that we will participate in the review process of the submitted manuscript entitled “Optimum dietary protein and lipid levels of Mystus gulio fry based on its growth, feed utilization, nutrient gain, and whole-body chemical composition” in your esteemed journal, Aquaculture International.
Consent for publication
We all the authors do hereby given our consent to publish the manuscript entitled “Optimum dietary protein and lipid levels of Mystus gulio fry based on its growth, feed utilization, nutrient gain, and whole-body chemical composition” in your esteemed journal, Aquaculture International.
Conflict of interest
The authors declare no competing interests.
Additional information
Handling Editor: Gavin Burnell
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Khatua, R., Mohanta, K.N., Chandan, N.K. et al. Dietary protein and lipid concentrations affect the growth, nutritional indices, and whole-body composition of long-whisker catfish, Mystus gulio, fry. Aquacult Int 29, 2085–2099 (2021). https://doi.org/10.1007/s10499-021-00738-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-021-00738-x