
Abstract
Acari community structure and function associated with delayed pig carrion decomposition has not been examined. In this study, 18 swine carcasses were studied in central Texas, USA, during two consecutive summers (2013 and 2014). Samples of ca. 400 g soil were collected from beneath, aside, and 5 m away from each pig carcass over 180 days. Mites from soil samples were extracted using Berlese funnels and identified to order and family levels and classified according to ecological function. In total 1565 and 1740 mites were identified from the 2013 and 2014 soil samples, respectively. Significant differences were determined for mite community structure at order and family levels temporally on carrion (e.g., day 0 × day 14) regardless of treatments and between soil regions where mites were collected (e.g., soil beneath vs. soil 5 m away from carrion). However, no significant differences were found in mite community structure at the order level between pig carrion with and without delayed Diptera colonization (i.e., treatments). Analysis at the family level determined a significant difference across treatments for both summers. Ecological function of mites did not change significantly following the delayed decomposition of pig carcasses. However, detritivores and fungivores were significant indicator groups during the pig carrion decomposition process. Furthermore, 13 phoretic mite species associated with eight forensically important beetle species were documented. Data from this study indicated that the rate of nutrient flow into the soil impacted associated arthropod communities; however, detecting such shifts depends on the taxonomic resolution being applied.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Carrion is defined as dead and decaying vertebrate animal remains, and it represents only a small part of the total detritus pool in most large-scale ecosystems (Swift et al. 1979; Parmenter and MacMahon 2009; Benbow et al. 2020). However, carrion plays significant roles in nutrient recycling that differ from plant detritus: (i) carrion is nutrient rich (low C:N ratio) whereas plant litter is usually low in nutrients and high in C:N ratio (Swift et al. 1979), and (ii) carrion decomposes perhaps up to three orders of magnitude faster than plant litter (Parmenter and MacMahon 2009). These two qualities make carrion a unique detrital pool in an ecosystem (Barton et al. 2013a).
Nutrients released from the remains into the ground are predominately recycled by soil arthropods, which are quite diverse. In fact, large numbers of microarthropods (e.g., mites and collembolans) are found in most types of soil, and soil mites usually outnumber collembolans (Coleman et al. 2004). Four important groups of mites occur frequently in soil: Oribatida, Prostigmata, Mesostigamata, and Astigmata. Oribatids are the most common mites found in soil. These mites are usually fungivores and detritivores. Mesostigmatid mites are typically predators on other small fauna or insect eggs, although some species are fungivores. Astigmatid mites are found associated with rich, decomposing nitrogen sources and are abundant in agricultural soils and stored products, whereas the Prostigmata contains a broad diversity of mites with a variety of feeding habits and strategies (OConnor 1994; Labandeira et al. 1997; Koehler 1997; Coleman et al. 2004).
Oribatids are known to influence organic litter decomposition and nutrient dynamics in forest environments (Wickings and Grandy 2011). They feed on microbial populations as well as fragmented plant detritus (Petersen and Luxton 1982). Oribatids can store and process a significant portion of the Ca input in forest litter as well (Gist and Crossley 1975). Furthermore, oribatids are also well known to serve as bioindicators in the agroecosystem (Behan-Pelletier 1999). Some families of Prostigmata are predaceous, whereas others are microbial feeders, plant feeders, or parasites (Kethley 1990). In general, the larger predaceous Prostigmata feed upon other arthropods or their eggs, whereas the smaller species are typically nematophagous (Kethley 1990). The Mesostigmata contains fewer soil-inhabiting species than do Oribatida and Prostigmata. Many Mesostigmata are parasitic on vertebrates and invertebrates (Krantz 1978). The true soil species are almost all predators; only a few species (e.g., Uropodidae) are polyphagous, feeding on fungi, nematodes and juvenile insects (Gerson et al. 2008). Many species of Mesostigmata have a close association with other insects or arthropods for dispersal purposes (i.e., phoretic relationship) (Hunter and Rosario 1988).
Astigmata are the least common of the soil mites, although they may become abundant in certain habitats (Luxton 1981). The free-living Astigmata prefer moist environments high in organic matter, and most are microbial feeders (Andrén et al. 1995). Those with chelate chelicerae are able to chew vegetable materials, fungi, and algae (Krantz and Lindquist 1979). The Astigmata are specialists in patchy or ephemeral habitats, and they are able to reproduce in a relatively short time; hence, many species can build up large populations on concentrated resource patches within days (OConnor 2009a). Astigmata are the most successful group of mites in establishing symbiotic relationships with both vertebrates and invertebrates (Houck and OConnor 1991; OConnor 1994). One of the reasons why Astigmata is the most successful group of mites is their ability to disperse via phoresy (OConnor 1982, 1994). Phoresy is a common form of commensalism and is applied to interspecific relationships in which one organism (the phoront) attaches to another (the host) for the implied purpose of dispersal (Houck and OConnor 1991). Astigmatid deutonymphs (heteromorphic deutonymphs, also known as hypopi) most commonly occur in association with beetles (Coleoptera), ants and wasps (Hymenoptera), as well as flies (Diptera) (Binns 1982; Houck and OConnor 1991). Host specific relationships are most common where beetle species exploit rich temporary habitats such as carrion (e.g., Pelzneria sp. on Nicrophorus beetles) and vertebrate dung (e.g., Rhopalanoetus sp. on scarab beetles) (Houck and OConnor 1991). Reviews on host associations and phoretic mites have been provided by Hunter and Rosario (1988) on Mesostigmata, Norton (1980) on phoretic Oribatida, OConnor (2009a, b) on Astigmata phoretic mites, Eickwort (1990) on phoretic mites with social insects and Lindquist (1975) on mites with arthropods found in forest habitats.
Mites associated with carrion have been well documented (Perotti and Braig 2009; Perotti et al. 2010) with more than 212 phoretic mite species associated with carrion insects reported (Perotti and Braig 2009). Among these, Mesostigmata is the dominant group, represented by 127 species with 25 species belonging to Parasitidae and 48 species to Macrochelidae. Most of these mesostigmatans are associated with particular species of flies or carrion beetles. Astigmata mites are more frequently found on dried remains of vertebrate carrion. Of those identified, 52 species were phoretic on scavengers such as hide beetles (Coleoptera: Trogidae), skin beetles (Coleoptera: Dermestidae) and moths (Lepidoptera) (Perotti and Braig 2009). Barton et al. (2014) examined and compared the changes in abundance, species richness, and composition of mite and beetle assemblages sampled on kangaroo (Macropus giganteus) carcasses in a grassy eucalypt woodland near Canberra, Australia. A majority of mites were phoretic, with the mesostigmatid genera Uroseius (Uropodidae), Macrocheles (Macrochelidae) and Parasitus (Parasitidae) the most abundant taxa (excluding astigmatid mites). Abundance and richness patterns of mites and beetles were very different, with mites reaching peak abundance and richness at weeks 6 and 12, and beetles at weeks 1 and 6. Mesostigmatid mite assemblages experienced a delay in peak abundance and richness relative to beetle assemblages, suggesting differences in dispersal and reproductive traits of arthropods contribute to the contrasting diversity dynamics of carrion arthropod communities, and further highlight the role of carrion as a driver of diversity and heterogeneity in the ecosystem (Barton et al. 2014). These data are important in forensic entomology investigations.
The first study including modern forensic acarology was reported by Brouardel in 1879, in a case where a newborn child was found mummified and the time of death was independently estimated based on caterpillars (Lepidoptera) and mites present (Perotti 2009). Mégnin was consulted regarding the mite specimens and calculated back the number of generations that would have been required to account for the number of mites present on the corpse. Mégnin concluded that the estimate for the time of death was around 7–8 months before the autopsy (Mégnin 1894). Since then, few studies have mentioned mites associated with carrion (Braack 1986; Goff 1989). In Europe, Saloña et al. (2010) collected arthropods from soil at a body recovery site and determined dominant necrophagous fauna included mites from the families Melicharidae (formerly Ascidae): Proctolaelaps euserratus Karg [misidentified as Proctolaelaps epuraeae (Hirschmann)], and Laelapidae: Hypoaspis (Gaeolaelaps) aculeifer (Canestrini). They also reported mites from the families Acaridae: Sancassania berlesei (Michael), Ascidae: Zerconopsis remiger (Kramer), Urodinychidae: Uroobovella pulchella (Berlese) and Macrochelidae: Glyptholaspis americana (Berlese) for the first time in the Iberian Peninsula. Mašán et al. (2013) reported an unusual mesostigmatid mite from the family Melicharidae, P. euserratus, found in association with decaying matter of animal and human decomposition in various European countries including Slovakia, Spain and the UK. This mite species is thus considered a potential marker for later stages of decomposition, namely butyric fermentation and dry decomposition.
Saloña-Bordas and Perotti (2014) reported a case of a hanged corpse in Spain. They recovered four species of phoretic mites, namely Poecilochirus carabi s.s. G. & R. Canestrini (Parasitidae), Poecilochirus (Physoparasitus) davydovae Hyatt (Parasitidae), Pelzneria crenulata (Oudemans) (Histiostomatidae), and Pelzneria necrophori (Dujardin) (Histiostomatidae) on carrion beetles (Silphidae: Necrodes and Nicrophorus) and rove beetles (Staphylinidae: Creophilus maxillosus [L.]), which were used for interpreting the case. Szelecz et al. (2018) presented a case of human remains found in a Swiss forest where they isolated 391 mites belonging to four Acari orders (Astigmata, Mesostigmata, Prostigmata and Oribatida). Among these, the Acaridae, Sancassania berlesei, Cosmoglyphus (formerly Sancassania) oudemansi, and an unidentified Histiostomatidae were found in their immature stages. The authors suggested that the large numbers of Sancassania found in the deutonymphal stage are strong indicators of movement or relocation of the remains (Szelecz et al. 2018). Recently, deutonymphs of Lardoglyphus zacheri Oudemans were found attached to larder beetles (Coleoptera: Dermestidae) colonising experimental pig carcasses in northern Spain for the first time (Salona-Bordas and Perotti 2020). In Asia, Silahuddin et al. (2015) documented 11 families of mites associated with rabbit (Oryctolagus cuniculus [L.]) carcasses in Malaysia. Similarly, Hanifah et al. (2015) found Macrocheles scutatiformis (Macrochelidae) on dung beetles (Phaeochroops freenae Kuijten) and soil beneath monkey and rabbit carcasses placed in a secondary forest in Malaysia. Recently, a review of mites in Malaysia has suggested the use of mites as biogeographical markers or as forensic indicators (Azmiera et al. 2020a).
In the USA, OConnor (2009b) reviewed forensically relevant Astigmata. These mites include Acaridae (genera Acarus, Tyrophagus, and Sancassania), Lardoglyphidae (Lardoglyphus), and Histiostomatidae (Spinanoetus, Pelzneria, Myianoetus, Histiostoma). Pimsler et al. (2016) reported that the muscid fly, Synthesiomyia nudiseta (van de Wulp) (Diptera: Muscidae), was collected during three indoor medicolegal forensic entomology cases in Texas, USA. In each case, mites were found in association with the sample and subsequently identified as Myianoeus muscarum (L.) (Acariformes: Histiostomatidae). This report suggested that this mite is of potential value in forensic investigations, as it lends new insights into the community structure of colonizers on human remains in indoor environments.
Many abiotic and biotic factors could contribute to the delay of insect access to carrion and consequently suppress the decomposition process (Campobasso et al. 2001). Abiotic factors that delay insect colonization on carrion include weather and location origins such as season, temperature, precipitation, and high altitudes (e.g., highlands or high-rise buildings), which could deter immediate insect access and oviposition activities (Campobasso et al. 2001; Mahat et al. 2009; Heo et al. 2017). Another abiotic factor includes burial activities or being hidden, which could happen either naturally (e.g., carcass hidden by scavengers), or artificially (e.g., in criminal cases where human cadavers have been wrapped, buried or hidden in concealed containers) (Anderson et al. 2001). Third, periodicity could be another abiotic factor, for example, animals that die during the night when blow flies are not active, and oviposition is likely to occur in the next morning (Introna et al. 1998; Reibe and Madea 2010).
Because no previous studies have examined the mite community associated with carrion experiencing delayed Diptera colonization, the objective of this study was to examine the impact of delayed Diptera colonization of carrion on the soil mite community structure and function.
Materials and methods
Site description and experimental design
Detailed methodologies have been published in Heo et al. (2019). In general, swine carcass (Sus scrofa L.) decomposition was studied at a site belonging to the Field Laboratory, Texas A&M University (Burleson County, College Station, TX, USA; 30°33′18.54″N, 96°25′38.71″W, 68 m a.s.l.). The soil at the study site was characterized as loamy and clayey (Weswood-Coarsewood). The soil surface layer is reddish/brown silt loam, moderately permeable, and moderately alkaline (Jurena 2007). Vegetation at the study site is considered part of the blackland prairie ecoregion (http://www.texasalmanac.com). Common vegetation found at the study site includes Johnsongrass (Sorgum halepense L.) (dominant cover plant, covered approximately 75% of the study site), oak (Quercus spp.), annual sunflower (Helianthus annuus L.), thistles (Cirsium spp. Mill.) and others.
Studies were conducted in two consecutive summers during June 2013 and 2014. In total nine pig carcasses purchased from a local pig farmer in Anderson, Texas, were obtained for each year replicate. Sex and weight of each pig carcass were determined prior to placement in the field. The animals were deceased at the time of acquisition; therefore, the Texas A&M University Institutional Animal Care and Use Committee required no animal use protocol. The carcasses were double bagged and transported within 1 h after death to the study site. Carcasses were randomly placed minimally 20 m apart along three transects. All carcasses were oriented with heads to the north and dorsal side towards the east. The placement of pig carcasses in the field was calculated by using a Latin square design (the arrangement of treatment groups was different between years and each location was only used once). During each field season, in order to delay carrion decomposition by preventing fly colonization, three random carcasses were enclosed in individual 1.8 m3 Lumite® screen (18 × 14 mesh size) portable field cages (BioQuip Products, Rancho Dominquez, CA, USA) for 7 days, this treatment was designated as Post-7 group. Another three random carcasses were enclosed in a similar manner as above but were enclosed for 14 days, thus were designated as Post-14 group, while all insects were allowed access to the remaining three carcasses, which served as controls. All carcasses were covered with hand-made, anti-scavenging cages (0.6 m height × 0.9 m width × 1.2 m length) constructed of steel frames enclosed with poultry netting. All cages were then properly labelled according to their designation.
Climatological data including temperature and precipitation were recorded. NexSens DS1923 micro-T temperatures loggers (Fondriest Environmental, Alpha, OH, USA) were placed at the study site 0.3 m above the ground on the exclusion cages to measure local ambient temperature every 60 min for 40 days continuously. Temperature data were converted into accumulated degree hours (ADH) based on Higley and Haskell (2009). The minimum development temperature threshold was set as 10 °C as that is the minimum used for blow flies and beetles common on vertebrate carrion during the summer months in Texas, USA. Precipitation during the study period was recorded daily with a rain gauge attached to a wooden stake approximately 1.3 m above the ground and 1 m north from one of the pig carcasses.
Soil samples at the study site were taken 5 days (day −5) prior to the placement of carrion and served as the baseline. During the experiment, soil samples were collected from beneath and at the side of pig carcasses (designated as soil lateral, which is approximately 30 cm distant from the location of soil beneath the carrion), along with a control sample which was located 5 m north from the carrion on days 0, 7, 14, 21, 40, 90 and 180 to determine the soil mite community structure and function over time. Soil samples of approximately 400 g were collected using a plastic trowel and a steel tin can with 170 g capacity that were disinfected using Lysol between uses. After collection, each soil sample was placed in a Ziploc bag and labelled. These soil samples were kept in a cooler box (70 cm long, 40 cm wide, 45 cm high) filled with ice (ca. 4 °C) and transported to the laboratory. For each soil sample, approximately 20 g was then separated from each individual sample, transferred to a freezer bag, and stored in a freezer (Kenmore, USA) at −20 °C for soil chemistry analysis. The remaining soil sample was then placed in a Berlese funnel for soil arthropod extraction. After each soil sample was transferred to the Berlese funnel, the interior of the sample bag was washed with 70% ethanol to detach arthropods on the plastic surface due to electrostatics. The ethanol-arthropod mixture was then transferred directly into the mason jar containing 100 ml 70% ethanol, which was used to collect and preserve extracted specimens.
Berlese funnels were used to process the samples for 72 h to extract soil arthropods. The bucket used was a plastic utility pail (25 cm diameter, 25 cm height, 18 cm diameter at base), with a round-shaped hardware cloth (gauze size 23, mesh size 0.6 cm) placed at the bottom of the bucket. Soil samples were placed on three pieces of filter paper (9 cm diameter, qualitative P8, Fisherbrand) located on top of the hardware cloth located at the base of a funnel (20 cm diameter, 18 cm high, and 1.4 L volume). A 40 W, 470 lumens, soft white in color (Philips A19) light bulb was used to dry the soil sample. Soil arthropods moving away from the drying soil would fall into the funnel and be collected in the mason jar assigned to the sample.
Samples collected in each mason jar were examined for the presence of soil arthropods under a dissecting stereo microscope (EMZ-8TR, Meiji, Japan). The content of the mason jar was shaken gently before being poured into a plastic Petri dish (9 cm diameter, BD Falcon). A piece of grid paper (divided into 16 squares equally) was placed at the bottom of the Petri dish for the purpose of mite counting and sampling protocols. All extracted soil arthropods were counted, recorded and identified to the lowest taxonomical rank possible using Triphehorn and Johnson (2005) and Stehr (1987). Once counted, soil arthropods and the excess ethanol were transferred to a 400 ml Pyrex beaker. A piece of Grade 3 Whatman filter paper (15 cm diameter) labelled with specimen details was placed inside the funnel in order to trap the soil arthropods while the excess ethanol was removed via filtration through a Büchner funnel. After filtration, a piece of wax paper (Reynolds) was used to wrap the filter paper, along with the trapped arthropods, sealed and then stored in a freezer (−20 °C) as records.
Mites recovered from the soil samples were first counted and divided into two morphological categories, namely Oribatida and non-oribatids. To identify the mites to family, subsampling of mites was performed. Four distinct non-overlapping numbers were generated using a random number generator online (https://www.random.org/) for every sample. The mites that fell randomly into these four numbers on the 16-square grid paper were then picked up using a loop modified from a minuten pin and transferred into a small Petri dish (35 × 10 mm) filled with several drops of specimen clearing fluid (Bioquip). Mite specimens were immersed in the fluid for 24 h at room temperature placed in a fume chamber. The clearing duration could be extended depending on the degree of clearing. This clearing fluid is principally a lacto-phenol solution with glacial acetic acid. After clearing, the specimens were transferred to a glass slide (size 76 × 25 mm; Hamilton Bell) with a small drop of polyvinyl alcohol (PVA) mounting medium (Bioquip). The PVA medium contained Elvanol and lactophenol and is usually used as a substitute for Hoyer’s solution. A square cover slip (22 × 22 mm; Bioquip) was then placed on top of the mite specimens (up to 10 mites per slide). The slide was labelled accordingly, air-dried for 3–4 days and stored in a microscope slide box (76 × 25 mm; Bioquip) in an upright position. When encountered, phoretic mites were collected from macroarthropods (e.g., underneath the elytra of a beetle) and processed as in the above methods.
Mites mounted on slides were examined under a light microscope with 40 × magnification (Nikon, Japan) and identification was carried out to family level using Krantz and Walter (2009). Species confirmation of Astigmata and other groups of mites were made by relevant taxonomists.
Statistical analyses
Soil arthropod community data (separated by order, family, and function) were analysed using the statistical program JMP Pro v.11.0.0 (SAS Institute, Cary, NC, USA) for ANOVA and Tukey–Kramer honestly significant difference (HSD) post-hoc test. In addition, R project for statistical computing (R v.3.0.2) was employed to analyse soil arthropod community data using the vegan package (Oksanen et al. 2013). Vegan contains the methods of multivariate analysis (e.g., permutational analysis of variance or PERMANOVA) needed in analysing ecological communities, and tools for diversity analysis. Bonferroni corrections were used to test for significance of pair-wise comparisons without an increased probability of rejecting the null hypothesis when it was actually true (type I error) (Cabin and Mitchell 2000).
Indicator species analysis (ISA) was performed to describe which family or functional group was the best indicator among the soil mite communities based on the abundance data, with 0 representing no indication and 100 being a perfect indication for each grouping. As the threshold for statistical significance was considered α = 0.05.
Results
Weather data
Detailed weather data have been published in Heo et al. (2019). In 2013, the mean temperature was 30.6 ± 7.8 °C, with maximum 47.7 ± 4.5 °C and minimum 15.5 ± 0.0 °C. Total accumulated degree hour (ADH) for the 2013 trial was 29,209.70 (base temperature of 10 °C). Total precipitation recorded from the rain gauge throughout the study period was 39.1 mm. In 2014, the mean temperature was 29.3 ± 6.5 °C, with maximum 43.0 ± 1.8 °C and minimum 19.0 ± 0.0 °C. Total ADH for the 2014 trial was 28,080.67. There were 13 rain events recorded during the study period. Total precipitation recorded from the rain gauge throughout the study period was 171.45 mm. The ambient temperature and ADH in summer 2013 were significantly greater than in summer 2014 (p = 0.0004 and 0.014, respectively). However, for precipitation, although summer 2014 showed higher precipitation compared to summer 2013 (surplus of 132.33 mm), a two-tailed t test showed no significance between these two trials (p = 0.27).
Year effect
There was a year effect (F = 0.9326, df = 1, p = 0.001) on morphospecies (i.e., community structure through basic identification) of mites between the two trials. Furthermore, when ecological function of mites was analyzed for year effect, the results showed that there was a significant difference between years (F = 11.175, df = 1, p = 0.001) as well. Hence, data for soil mite community structure were analyzed separately according to year.
Replicate effect
There was no replicate effect (F = 0.9326, df = 1, p = 0.46) when analyzed based on morphospecies of mites or ecological function (F = 1.7254, df = 1, p = 0.14). Therefore, all data in the replicates were pooled and analyzed together.
Total orders collected
In the 2013 trial, in total 1565 mite specimens were mounted on slides. The most dominant group mounted on slides was the order Mesostigmata (36.7%) followed by the cohort Astigmata (36.5%) (Table 1).
Similarly, 1740 mite specimens were mounted on slides in 2014 and the most dominant group was the order Mesostigmata (54.0%) followed by the suborder Oribatida (24.3%) and cohort Astigmata (18.5%). Note that no species of the suborder Endeostigmata were collected from the soil samples in 2014. This could be due to simple rarity, mis-sampling in the field, or random effects during the sub-sampling process in the laboratory.
Total families collected
For the suborder Oribatida, specimens were not identified to family level. The results presented below were the families identified from non-Oribatida mites. In 2013, in total 26 families were identified from all the non-oribatid mites. One family was identified in the suborder Endeostigmata, two families in the cohort Astigmata, 10 families in the order Mesostigmata, and 13 families in the suborder Prostigmata. The most dominant family was Acaridae (Astigmata) (46.5%), followed by Macrochelidae (11.1%) and Ascidae (8.95%) (Mesostigmata) (Table 2). In 2014, 17 families were identified from all the non-oribatid Acari. One superfamily was identified as Eupodoidea (Prostigmata), one family in the cohort Astigmata, nine families in the order Mesostigmata, and seven families in the suborder Prostigmata. The most dominant family was Macrochelidae (31.5%), followed by Acaridae (25.1%) and Ascidae (14.1%). Note that most Acaridae are detritivores or necrophages whereas most known Macrochelidae and Ascidae are predators in soil. Only some of the mite specimens have been identified to genera and species. Hence, statistical analysis at the genus-species level was not performed.
Total ecological function identified
Five major ecological functional groups of mites were identified in this study. These include herbivores, predators, detritivores, fungivores, and nectarivores/pollinivores (as defined by Walter and Proctor 2013). The most abundant functional group during pig carrion decomposition was the detritivores (62.4%), followed by the predators or parasites (33.1%). In 2014, the most abundant functional group was the predators/parasites guild (50.8%), followed by the detritivores (46.8%) (Table 3).
PERMANOVA of soil mite community structure by order
PERMANOVA was performed on the mites identified at this taxonomic level (including superorder/order/suborder/cohort) to determine the effects of independent variables. In 2013, day of carrion decomposition (day) and soil region (region) were both significantly different (p = 0.001) with no significant interactions. For 2014, the results showed day and region were significantly different (p = 0.001). However, there were interactions between day × treatment (p = 0.016) and day × region (p = 0.008). Treatment (no delay vs. delay of Diptera colonization) was consistently not significant for both years (Table 4).
Abundance of soil mite (by order) in soil beneath the pig carrion
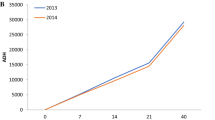
In 2013, a significant difference was found on day 14 (control × post-14 and control × post-7, both with p = 0.0044) for Prostigmata. No significant difference was found for the other orders. However, in 2014, significant differences between treatments were found on day 7 (control × post-7, p = 0.019; control × post-14, p = 0.0066) and day 40 (control × post-7, p = 0.014; control × post-14, p = 0.0023) for Mesostigmata, whereas for Prostigmata, a significant difference (p = 0.047) was found on day 180. There was no significant difference detected among treatments for both Astigmata and Oribatida (Fig. 1).

Soil mite community abundance (by order) beneath the pig carrion according to treatments in summer 2013 and 2014 at Snook, TX, USA. PMI (D) = postmortem interval (in days)
PERMANOVA of soil mite community structure by family
PERMANOVA showed day, treatment and region were significantly different with no other significant interactions detected in 2013. Similar results were demonstrated in 2014 trial. However, several significant interactions were detected, such as day × treatment (p = 0.011), day × position (p = 0.001), and treatment × position (p = 0.025) (Table 5).
As there was a significant effect in day, treatment and region for both years, further analyses were carried out. For soil regions, all soil regions were significantly different from each other (p < 0.001), indicating soil mite community structure changes according to region (Table 6). As for day of decomposition, most of the pairwise comparisons between days were significantly different (Table 7). In 2013 treatment effect was significantly different for control × post-7 (p = 0.035), whereas in 2014, post-7 × post-14 was significantly different (p = 0.02) (Table 8).
Indicator species analysis (ISA) showed three families of soil mites namely Phytoseiidae, Pygmephoridae and Acaridae were the significant indicators for 2013 whereas Ascidae, Cunaxidae, Acaridae and Erythraeidae were significant indicators for 2014 (Table 9).
Abundance of soil mites (by family) in soil beneath the pig carrion
There was no significant difference in soil mite abundance between treatments in soil beneath (p = 0.16). However, there was a day effect (p = 0.0004) detected. In this study, Acaridae (Sancassania spp.) was the dominant family observed in all treatment groups. For control carcasses, Acaridae was first observed from day 7, its abundance peaked on day 40, and subsequently reduced in abundance on day 180, and was succeeded by Macrochelidae (Macrocheles spp.), which peaked in abundance on day 180. Conversely, Acaridae was dominant for post-7 group from day 40 until day 180 (Figs. 2 and 3).

Soil mite community abundance (by family) beneath the pig carrion according to treatments in summers 2013 and 2014 at Snook, TX, USA. PMI (D) = postmortem interval (in days)

Soil mite community abundance of Macrochelidae and Acaridae beneath the pig carrion according to treatments in summers 2013 and 2014 at Snook, TX, USA. PMI (D) = postmortem interval (in day)
Soil mite function
PERMANOVA results on functional groups assigned to mites collected indicate a significant difference in day and soil region (both p = 0.001) without a significant interaction (Table 10). Treatments had no significant effects on soil mite community function (p = 0.40) in 2013; however, it was significant in 2014. For indicator species analysis, two significant functional groups in 2013 were detected namely fungivores and detritivores. However, no functional groups of soil mites were significant in 2014 (Table 11).
Abundance of soil mites (by functional groups) in soil beneath the pig carrion
The major functional groups of mites for all treatments were the detritivores, followed by the predators. Detritivores became dominant on day 14, 40 and 90 for control carcasses; day 40, 90 and 180 for post-7 carcasses and almost every sampling day for post-14 group. Although no significant difference was detected on the predator/parasite group over time, post-7 had a higher abundance of predators at soil beneath on day 21. Fungivores were in higher abundance under the control carcasses on day 14, and then re-populated on day 90. For fungivores, a significant difference was observed on day 14 (between control × post-7 and control × post-14, both p = 0.018). Detritivores in control and post-14 group increased to their peak abundance on day 40 whereas post-7 group continued to increase in detritivore abundance even on day 180. In 2014, the predators were the major functional group of mites for all treatments, followed by detritivores. There were significant differences in predator abundance between treatments in soil beneath on day 7 (control × post-7, p = 0.0029; control × post-14, p = 0.0013) and day 40 (control × post-7, p = 0.020; control × post-14, p = 0.0023). Detritivores increased their abundance on day 14 or 21 and dominated the soil beneath even up to day 180. Fungivores were observed to increase in abundance during the later stage of carrion decomposition (Fig. 4).

Soil mite abundance (by functional groups) beneath the pig carrion according to treatments over decomposition days in summers 2013 and 2014 at Snook, TX, USA
Beetles associated with phoretic mites in 2013 and 2014 trials
Phoretic mites were found in association with a total of six families and eight species of Coleoptera in both 2013 and 2014. These coleopterans families include Trogidae (one species), Dermestidae (three species), Cleridae (one species), Tenebrionidae (one species), Histeridae (one species) and Silphidae (one species). Most of the phoretic mites were found underneath the elytra of the host beetle, and a few of them were found attached on the ventral surfaces or sternites of the beetle hosts (e.g., silphid beetles). A total of nine families and 13 species of phoretic mites were identified from all beetle hosts collected in both trials. The phoretic mite families included Histiostomatidae (one species), Halolaelapidae (one species), Melicharidae (one species), Lardoglyphidae (two species), Acaridae (two species), Winterschmidtiidae (one species), Ologamasidae (one species), Parasitidae (two species) and Macrochelidae (one species) (Table 12).
Discussion
Results from this study demonstrated delayed blow fly colonization on carrion impacted soil mite community structure. Soil mite community structure was changed significantly by year. This year effect could be due to the significant change in ambient temperature as well as ADH between summers 2013 and 2014. The soil mite community structure and the associated food web shifted as a result of the abiotic environment (Whitford 1989). Temperature has been known to affect the development, survival, and reproduction of insects and mites (Ratte 1984; Broufas et al. 2007; Heo et al. 2019). It has been documented that temperature, soil acidity, humidity, amount and quality of food and environmental disturbance affect the life cycle of oribatid mite species (Luxton 1975; Maraun and Scheu 2000). Precipitation was much higher in 2014 compared to the previous year (Heo et al. 2019). This additional soil moisture could affect soil mite community structure as past study had demonstrated that water could affect collembolans and soil mites significantly in summer (Chikoski et al. 2006). In dry soils with water potential below −1.5 MPa, most bacteria, protozoa, and nematodes are not active (these taxa entered a state called anhydrobiosis, some fungivorous mites even entered cryptobiosis state) (Whitford 1989). For instance, supplemented summer rainfall resulted in an increase in vegetation cover, leading to an increase in the abundance of the insect herbivores, Auchenorrhyncha (Hemiptera) (Masters et al. 1998). Rainfall is also demonstrated to affect the trophic position of some oribatid species, probably due to changes in microbial activity and community composition (Melguizo-Ruiz et al. 2017).
Soil mite community structure associated with the carrion was significantly different over time and soil regions. Bornemissza (1957) found different stages of decomposition were correlated with the animal communities occupying them, and these communities were contrasted with the community dwelling in the leaf litter and soil. The various stages of decomposition influenced the underlying soil differently, with the greatest effect being observed during the ‘black putrefaction’ and ‘butyric fermentation’ stages (Bornemissza 1957). The current study determined similar results where day 7 (in between active and advanced decomposition stage) of control carcasses was significantly different with regard to soil mite community structures compared to post-7 and post-14 carcasses. As for soil regions, the current study agreed with Bornemissza (1957) where soil mite community beneath the carrion was significantly different between soil beneath and soil lateral (soil at a distance of ca. 30 cm away from beneath the carrion), and between soil beneath and soil 5 m away from carrion. Following these observations, we suggest ‘cadaver decomposition islands’ (CDIs) should be divided into several zones (i.e., beneath the carrion, or aside the carrion) (Heo et al. 2019) as each zone is uniquely inhabited by a soil mite community that differs in structure and function. The change in the soil mite community structure according to soil regions may be due to different concentrations of soil nutrients derived from the decomposition process, similar to the resource concentration hypothesis (Root 1973) where the decomposers (i.e., microorganisms such as bacteria and fungi, mites, earthworms, collembolans) are attracted to resources, and recycle the organic matter or perform carbon mineralization as part of their normal functions in the ecosystem (Brussaard 1997). From a nutrient flow viewpoint, soil mites are envisioned as accelerating (or delaying) nutrient release from decomposing organic matter. They may do so directly, by feeding upon organic matter and associated microflora, or indirectly, by channeling and mixing the soil, improving the quality of substrate for microfloral and other consumers (Crossley 1977).
Interpretation of soil mite community responses to delayed colonization of swine carrion was dependent on taxonomic resolution implemented. Based on the results obtained, family-level, unlike order-level, assessment of the soil mite communities demonstrated more consistent results across parameters examined. Bowman and Bailey (1997) recorded similar phenomena with freshwater macro-invertebrate communities. They determined genus-level identification did not provide a strikingly different description of community structure than higher levels (e.g., family, order) of taxonomic identification. Similarly, Olsgard et al. (1997) examined the relationship between taxonomic resolution in analyses of a microbenthic community along an established pollution gradient, and they found that higher taxonomic levels are more likely to reflect a contamination gradient than are analyses based on species abundances.
Hawlena et al. (2012) demonstrated that grasshoppers (Orthoptera) stressed by spider predators (Araneae) have a higher body C:N ratio than other grasshoppers raised without spiders. This change in element content does not slow grasshopper decomposition but disturbs belowground community function, decelerating the subsequent decomposition of plant litter. This legacy effect of predation on soil community function appears to be regulated by the amount of herbivore protein entering the soil. Similarly, the absence of a necrophagous dipteran community at the initial phase of decomposition in this study changes the cadaveric resource quality, which subsequently changes the soil mite community structure and function.
In this study, we observed general trends in population dynamics of soil mites associated with pig carrion decomposition. For Mesostigmata (e.g., Macrocheles), the population abundance was somewhat similar to a bell curve over day of decomposition (see Fig. 1). For Oribatida, the abundance pattern along the course of decomposition was in an inverted bell-shaped whereas for Astigmata (e.g., Sancassania) it was an exponential growth phase followed by a stationary phase, and eventually a decline phase (similar to a sigmoidal curve). The bell curve dynamic demonstrated by mesostigmatans indicate arrival (via phoresy), in situ reproduction, and dispersal (via phoresy) along the carrion decomposition process. The inverted bell-shaped curve exhibited by oribatid mites rendered a contrasting pattern from that of mesostigmatans where oribatid populations decline during the active decomposition stage or when the influx of decompositional fluid occurred. When the decomposition process is completed or at the skeletonization stage, the population of oribatids recovers gradually (a phenomenon known as resilience). As for the astigmatans, the population increases exponentially (via phoresy and in situ reproduction) when the carrion is available as a food source, the population then enters a stagnant phase when the resources become depleted over time, and the population eventually declines due to the death or dispersal of mites to other habitats via phoresy. Similar trends have been observed in Silahuddin et al. (2015) and Azmiera et al. (2020b) for Mesostigmata, Astigmata, and Oribatida. These trends of mite population dynamics and succession could explain their ecological roles in carrion ecosystem and be a useful tool in mPMI estimation.
In forensic acarology, Macrochelidae were normally found at the early stage, whereas Acaridae were found at the later stage of decomposition (Che Kamaruzaman et al. 2018; Szelecz et al. 2018; Azmiera et al. 2020b). It has been recorded that Macrocheles muscaedomesticae (Scopoli) arrived at a human corpse coinciding with the occurrence of one of its major carrier fly species, Fannia scalaris, whereas Macrocheles glaber (Müller) were collected from a corpse concurrent with the arrival of beetles attracted by odors from the corpse, and Macrocheles perglaber Filipponi and Pegazzano, which have a preference for phoresy on Scarabaeidae and Geotrupidae beetles, were also sampled from a corpse. Regarding distribution, M. muscaedomesticae is known from urban and rural areas and poultry farms, M. glaber from outdoors, particularly the countryside, whereas M. perglaber is known from outdoor, rural and remote, potentially mountainous locations (Che Kamaruzaman et al. 2018). Another species, Macrocheles scutatiformis Petrova was reported from soil beneath monkey and rabbit carcasses in Malaysia, and some were phoretic on a dung beetle and a Chrysomya rufifacies larva (Hanifah et al. 2015). Other forensically important Mesostigmata mites include P. euserratus (Mašán et al. 2013), P. davydovae (Saloña-Bordas and Perotti 2014), Macrocheles matrius (Szelecz et al. 2018), Uroseius sp. (Moraza and Pérez-Martínez 2019), Macrocheles merdarius, Poecilochirus austroasiaticus, and Poecilochirus subterraneus (Medina et al. 2013; Pérez-Martínez et al. 2019). The current study found a species of Macrochelidae (Macrocheles sp. [natalie group]) phoretic on a carrion beetle, Necrophorus marginatus (Silphidae), which arrived at the pig carcass 48 h after placement at the study site.
Another interesting astigmatan species, Sancassania sp. (Acaridae), was found abundantly as deutonymphs in this study, especially toward the end stage of carrion decomposition, and remained at or nearby the carcass placement site even after a year. Similarly, in a forensic case reported by Szelecz et al. (2018), both S. berlesei and Cosmoglyphus oudemansi were extracted from sample sites associated with bones, indicating movement or relocation of the remains. Also, S. berlesei was recorded from two human corpses in Belgium that had been discovered between 3- and 4-months postmortem (Leclercq and Verstraeten 1988). Clearly, the presence of Sancassania could indicate a later stage of decomposition or serve as a locality indicator due to its widespread distribution in synanthropic habitats such as animal farms (OConnor 2009b; Szelecz et al. 2018; Azmiera et al. 2020b). Other reported astigmatans of forensic importance include Acarus siro (Mégnin 1894), Tyrophagus putrescentiae (Early and Goff 1986), Histiostoma sp. (Azmiera et al. 2019), Histiostoma sachsi (Leclercq and Verstraeten 1988), Myianoetus diadematus (Russell et al. 2004), Myianoetus muscarum (Pimsler et al. 2016), Spinanoetus sp. (OConnor 2009b), Pelzneria sp. (OConnor 2009b; Saloña-Bordas and Perotti 2014), Rhizoglyphus howensis (Hani et al. 2018) and L. zacheri (Saloña-Bordas and Perotti 2020). The present study recorded Sancassania phoretic on various beetle hosts such as Dermestidae, Tenebrionidae and Histeridae. Moreover, Hexanoetus sp. was found abundantly on trogid beetles, Omorgus sp., whereas L. zacheri and L. anglolensis were both phoretic on dermestid beetles in Texas, USA.
Future research should attempt to address limitations associated with the current research. As previously mentioned, identification to genus or species level of mites was not extensive. It is important to pin down the species of mites that are important in carrion ecology and its application in forensic acarology. Second, a longer period of observation time (e.g., a year) is needed in order to offer a complete picture about carrion decomposition and the impacts on soil mite communities. Also, it would be ideal to increase the sample size (e.g., number of carcasses) to increase the statistical power and confidence interval. Similar studies could be carried out on different types of soil, in different types of habitat or landscape, different ecoregions or climatic regions as well.
Soil mite community structures are sensitive to the years of field trial, days of decomposition, and soil regions, but its sensitivity to treatment effect depends on the taxonomic resolution (i.e., family is more sensitive than order). It is evident that there was a shift in soil mite community structure and function in response to delayed vertebrate decomposition. The mite community structural shift could be due to abiotic factors such as ambient temperature, and soil moisture. Furthermore, we suggest systematic zoning of cadaver decomposition islands (CDIs) as small distances away from the soil beneath the carrion could have significant difference in soil mite community structure and function. There are still many unexplored areas in soil mite diversity associated with carrion decomposition, the relationship between phoretic mites and their insect hosts (e.g., beetles), and factors governing the dynamics of mite populations warrant further investigations. It is necessary to study soil mites and other soil organisms (e.g., soil microbes, protozoa, nematodes, collembolans, isopods) in carrion ecology for the benefits of its future applications in forensics (e.g., forensic acarology and soil biology).
References
Anderson GS (2001) Insect succession on carrion and its relationship to determining time of death. In: Byrd JH, Castner JL (eds) Forensic entomology: the utility of arthropods in legal investigations. CRC Press, Boca Raton, pp 143–175
Andrén O, Bengtsson J, Clarholm M (1995) Biodiversity and species redundancy among litter decomposers. In: Collins HP, Robertson GP, Klug MJ (eds) The significance and regulation of soil biodiversity. Springer, Netherlands, pp 141–151
Azmiera N, Mariana A, Heo CC (2019) First report of Histiostoma sp. (Astigmata: Histiostomatidae) phoretic on a forensically important blowfly, Chrysomya villeneuvi (Diptera: Calliphoridae). Trop Biomed 36(4):1099–1104
Azmiera N, Mariana A, Pimsler ML, Heo CC (2020a) Review of mites found on various animal hosts and at different localities in Malaysia. J Med Entomol 57:1354
Azmiera N, Mariana A, Heo CC (2020b). The abundance and biodiversity of forensically important mites found at four different ecoregions in Peninsular Malaysia. Submitted for review
Barton PS, Cunningham SA, Lindenmayer DB, Manning AD (2013) The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia 171(4):761–772
Barton PS, Weaver HJ, Manning AD (2014) Contrasting diversity dynamics of phoretic mites and beetles associated with vertebrate carrion. Exp Appl Acarol 63(1):1–13
Behan-Pelletier VM (1999) Oribatid mite biodiversity in agroecosystems: role for bioindication. Agric Ecosyst Environ 74(1–3):411–423
Benbow ME, Receveur JP, Lamberti GA (2020) Death and decomposition in aquatic ecosystems. Front Ecol Evol 8:17
Binns ES (1982) Phoresy as migration—some functional aspects of phoresy in mites. Biol Rev 57(4):571–620
Bornemissza GF (1957) An analysis of arthropod succession in carrion and the effect of its decomposition on the soil fauna. Aust J Zool 5(1):1–12
Bowman MF, Bailey RC (1997) Does taxonomic resolution affect the multivariate description of the structure of freshwater benthic macroinvertebrate communities? Can J Fish Aquat 54(8):1802–1807
Braack LEO (1986) Arthropods associated with carcasses in the northern Kruger National Park. S Afr J Wildl Res 16(3):91–98
Broufas GD, Pappas ML, Koveos DS (2007) Development, survival, and reproduction of the predatory mite Kampimodromus aberrans (Acari: Phytoseiidae) at different constant temperatures. Environ Entomol 36(4):657–665
Brussaard L (1997) Biodiversity and ecosystem functioning in soil. Ambio 26(8):563–570
Cabin RJ, Mitchell RJ (2000) To Bonferroni or not to Bonferroni: when and how are the questions. Bull Ecol Soc Am 81:246–248
Campobasso CP, Di Vella G, Introna F (2001) Factors affecting decomposition and Diptera colonization. Forensic Sci Int 120(1):18–27
Che Kamaruzaman NA, Mašán P, Velásquez Y, González-Medina A, Lindström A, Braig HR, Perotti MA (2018) Macrocheles species (Acari: Macrochelidae) associated with human corpses in Europe. Exp Appl Acarol 76(4):453–471
Chikoski JM, Ferguson SH, Meyer L (2006) Effects of water addition on soil arthropods and soil characteristics in a precipitation-limited environment. Acta Oecol 30(2):203–211
Coleman DC, Crossley DA, Hendrix PF (2004) Fundamentals of soil ecology. Elsevier Academic Press, Burlington
Crossley DA Jr (1977) The roles of terrestrial saprophagous arthropods in forest soils: current status of concepts. In: Mattson WJ (ed) The role of arthropods in forest ecosystems. Springer, Berlin, pp 49–56
Early M, Goff ML (1986) Arthropod succession patterns in exposed carrion on the island of O’ahu, Hawaiian Islands, USA. J Med Entomol 23(5):520–531
Eickwort GC (1990) Associations of mites with social insects. Ann Rev Entomol 35(1):469–488
Gerson U, Smiley RL, Ochoa R (2008) Mites (Acari) for pest control. Wiley, Oxford
Gist CS, Crossley DA Jr (1975) The litter arthropod community in a southern Appalachian hardwood forest: numbers, biomass and mineral element content. Am Midl Nat 93(1):107–122
Goff ML (1989) Gamasid mites as potential indicators of postmortem interval. Prog Acarol 7:443–450
Hani M, Thieven U, Perotti MA (2018) Soil bulb mites as trace evidence for the location of buried money. Forensic Sci Int 292:e25–e30
Hanifah AL, Ahamad M, Omar B, Yusoff AT, Narainasamy VV, Takaku G (2015) First report of Macrocheles scutatiformis (Acari: Macrochelidae) found on insects and soil beneath animal carcasses in Malaysia. Sains Malays 44(5):681–686
Hawlena D, Strickland MS, Bradford MA, Schmitz OJ (2012) Fear of predation slows plant-litter decomposition. Science 336(6087):1434–1438
Heo CC, Latif B, Kurahashi H, Tan SH, Chew WK, Nazni WA, Omar B (2017) Preliminary observations of necrophagous dipteran activities and a new host record of parasitoids on a high-rise building. Pol J Entomol 86(2):165–180
Heo CC, Teel PD, Banfield MM, Tomberlin JK (2019) Soil arthropod community responses to carrion with delayed insect access. Food Webs 20:e00118
Higley L, Haskell N (2009) Insect development and forensic entomology. In: Byrd JH, Castner JL (eds) Forensic entomology. The utility of arthropods in legal investigations, 2nd edn. CRC Press, Boca Raton, pp 389–405
Houck MA, OConnor BM (1991) Ecological and evolutionary significance of phoresy in the Astigmata. Ann Rev Entomol 36(1):611–636
Hunter PE, Rosario RMT (1988) Associations of Mesostigmata with other arthropods. Ann Rev Entomol 33(1):393–417
Introna F, Campobasso CP, Di Fazio A (1998) Three case studies in forensic entomology from southern Italy. J Forensic Sci 43(1):210–214
Johnson NF, Triplehorn CA (2005) Borror and DeLong’s Introduction to the Study of Insects. Thompson Brooks/Cole, Belmont
Jurena MR (2007) Soil survey of burleson county, Texas. Natural Resources Conservation Service, Washington DC
Kethley J (1990) Acarina: prostigmata (Actinedida). In: Dindal DL (ed) Soil biology guide. Wiley, New York, pp 667–756
Koehler HH (1997) Mesostigmata (Gamasina, Uropodina), efficient predators in agroecosystems. Agric Ecosyst Environ 62(2):105–117
Krantz GW (1978) A manual of acarology, 2nd edn. Oregon State University Book Stores, Corvallis
Krantz GW, Lindquist EE (1979) Evolution of phytophagous mites (Acari). Ann Rev Entomol 24(1):121–158
Krantz GW, Walter DE (2009) A manual of acarology, 3rd edn. Texas Tech University Press, Lubbock
Labandeira CC, Phillips TL, Norton RA (1997) Oribatid mites and the decomposition of plant tissues in Paleozoic coal-swamp forests. Palaios 12(4):319–353
Leclercq M, Verstraeten C (1988) Entomologie et médecine légale: datation de la mort: acariens trouvés sur des cadavres humains. Bull Ann De La Soc R Belge D’entomol 124:195–200
Lindquist EE (1975) Associations between mites and other arthropods in forest floor habitats. Can Entomol 107(04):425–437
Luxton M (1975) Studies of the oribatid mites of a Danish beech wood soil. II. Biomass, calorimetry and respirometry. Pedobiologia 15:161–200
Luxton M (1981) Studies on the astigmatid mites of a Danis beech wood soil. Pedobiologia 22:29–38
Mahat NA, Zafarina Z, Jayaprakash PT (2009) Influence of rain and malathion on the oviposition and development of blowflies (Diptera: Calliphoridae) infesting rabbit carcasses in Kelantan. Malaysia Forensic Sci Int 192(1–3):19–28
Maraun M, Scheu S (2000) The structure of oribatid mite communities (Acari, Oribatida): patterns, mechanisms and implications for future research. Ecography 23:374–382
Mašán P, Perotti MA, Saloña-Bordas MI, Braig HR (2013) Proctolaelaps euserratus, an ecologically unusual melicharid mite (Acari, Mesostigmata) associated with animal and human decomposition. Exp Appl Acarol 61(4):415–429
Masters GJ, Brown VK, Clarke IP, Whittaker JB, Hollier JA (1998) Direct and indirect effects of climate change on insect herbivores: Auchenorrhyncha (Homoptera). Ecol Entomol 23(1):45–52
Medina AG, Herrera LG, Perotti MA, Ríos GJ (2013) Occurrence of Poecilochirus austroasiaticus (Acari: Parasitidae) in forensic autopsies and its application on postmortem interval estimation. Exp Appl Acarol 59(3):297–305
Mégnin P (1894) La Faune des Cadavres. Application de l’Entomologie à la Médecine Légale. (The fauna of corpses. Application of entomology to forensic medicine). G. Masson and Gauthier-Villars et Fils, Paris
Melguizo-Ruiz N, Jiménez-Navarro G, Zieger SL, Maraun M, Scheu S, Moya-Laraño J (2017) Complex effects of precipitation and basal resources on the trophic ecology of soil oribatid mites: implications for stable isotope analysis. Eur J Soil Biol 82:98–107
Moraza ML, Pérez-Martínez S (2019) The genus Uroseius Berlese (Acari: Mesostigmata: Uropodina: Trachytidae) in the Iberian Peninsula with description of a new species associated with animal remains. Syst Appl Acarol 24(5):929–944
Norton RA (1980) Observations on phoresy by oribatid mites (Acari: Oribatei). Int J Acarol 6(2):121–130
OConnor BM (1982) Evolutionary ecology of astigmatid mites. Ann Rev Entomol 27(1):385–409
OConnor BM (1994) Life-history modifications in astigmatid mites. In: Houck MA (ed) Mites: ecological and evolutionary analyses of life-history patterns. Springer, New York, pp 136–409
OConnor BM (2009a) Cohort astigmatina. In: Krantz GW, Walter DE (eds) A manual of acarology, 3rd edn. Texas Tech University Press, Lubbock, pp 565–657
OConnor BM (2009b) Astigmatid mites (Acari: Sarcoptiformes) of forensic interest. Exp Appl Acarol 49(1–2):125–133
Oksanen JF, Blanchet G, Kindt R, Legendre P, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2013) Vegan: Community Ecology Package. R package version 2.0-9. Accessed http://CRAN.R-project.org/package=vegan.
Olsgard F, Somerfield PJ, Carr MR (1997) Relationships between taxonomic resolution and data transformations in analyses of a macrobenthic community along an established pollution gradient. Mar Ecol Prog Ser 149:173–181
Parmenter RR, MacMahon JA (2009) Carrion decomposition and nutrient cycling in a semiarid shrub-steppe ecosystem. Ecol Monogr 79(4):637–661
Pérez-Martínez S, Moraza ML, Saloña-Bordas MI (2019) Gamasina mites (Acari: Mesostigmata) associated with animal remains in the mediterranean region of Navarra (Northern Spain). Insects 10(1):5. https://doi.org/10.3390/insects10010005
Perotti MA, Braig HR (2009) Phoretic mites associated with animal and human decomposition. Exp Appl Acarol 49(1–2):85–124
Perotti MA, Braig HR, Goff ML (2010) Phoretic mites and carcasses: Acari transported by organisms associated with animal and human decomposition. In: Amendt J, Goff ML, Campobasso CP, Grassberger M (eds) Current concepts in forensic entomology. Springer, Netherlands, pp 69–91
Petersen H, Luxton M (1982) A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 39(3):288–388
Pimsler ML, Owings CG, Sanford MR, OConnor BM, Teel PD, Mohr RM, Tomberlin JK (2016) Association of Myianoetus muscarum (Acari: Histiostomatidae) with Synthesiomyia nudiseta (Wulp)(Diptera: Muscidae) on human remains. J Med Entomol 53(2):290–295
Ratte HT (1984) Temperature and insect development. In: Hoffmann KH (ed) Environmental physiology and biochemistry of insects. Springer, Berlin, pp 33–66
Reibe S, Madea B (2010) How promptly do blowflies colonise fresh carcasses? A study comparing indoor with outdoor locations. Forensic Sci Int 195(1):52–57
Root RB (1973) Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecol Monogr 43(1):95–124
Russell DJ, Schulz MM, OConnor BM (2004) Mass occurrence of astigmatid mites on human remains. Abh Ber Naturkundemus Görlitz 76:51–56
Saloña MI, Moraza ML, Carles-Tolrá M, Iraola V, Bahillo P, Yélamos T, Outerelo R, Alcaraz R (2010) Searching the soil: forensic importance of edaphic fauna after the removal of a corpse. J Forensic Sci 55(6):1652–1655
Saloña-Bordas MI, Perotti MA (2014) First contribution of mites (Acari) to the forensic analysis of hanged corpses: a case study from Spain. Forensic Sci Int 244:e6–e11
Saloña-Bordas MI, Perotti MA (2020) First record of Lardoglyphus zacheri (Acari, Lardoglyphidae) in the Iberian Peninsula and new observations on its insect carriers. Syst Appl Acarol 25(3):412–419
Silahuddin SA, Latif B, Kurahashi H, Heo CC (2015) The importance of habitat in the ecology of decomposition on rabbit carcasses in Malaysia: implications in forensic entomology. J Med Entomol 52(1):9–23
Stehr FW (1987) Immature insects, vol 1. Kendall/Hunt Publishing, Dubuque
Swift MJ, Heal OW, Anderson JM (1979) Decomposition in terrestrial ecosystems. Studies in ecology, vol 5. University of California Press, Berkeley and Los Angeles
Szelecz I, Lösch S, Seppey CV, Lara E, Singer D, Sorge F, Tschui J, Perotti MA, Mitchell EA (2018) Comparative analysis of bones, mites, soil chemistry, nematodes and soil micro-eukaryotes from a suspected homicide to estimate the post-mortem interval. Sci Rep 8(1):1–14
Walter DE, Proctor HC (2013) Mites: ecology, evolution & behaviour. Springer, Dordrecht, The Netherlands
Whitford WG (1989) Abiotic controls on the functional structure of soil food webs. Biol Fertil Soils 8(1):1–6
Wickings K, Grandy AS (2011) The oribatid mite Scheloribates moestus (Acari: Oribatida) alters litter chemistry and nutrient cycling during decomposition. Soil Biol Biochem 43(2):351–358
Acknowledgements
The authors thank Roy Norton and Hans Klompen for their assistance in mite identification. Also, we would like to acknowledge Tawni Crippen (USDA-ARS), Aaron Tarone, Mickey Eubanks, Meaghan Pimsler, Jonathan Cammack, Brandon Lyons, and Le Zheng for their support both in the field and in the laboratory. Thanks also go to undergraduate student helpers: Michael Banfield, Stephanie Thornton, Jocelyne Soh, Adrianna Tirloni, Loren Williams (Penn State University), Katherina Kang, Aaron Guererro, and Caitlin Evers. Also, we would like to extend our gratitude to the Ministry of Higher Education, Government of Malaysia and Universiti Teknologi MARA, who provided funding to the first author.
Funding
The funding for this research project is supported by Texas A&M AgriLife. The first author is supported by the scholarship provided by the Ministry of Higher Education, Government of Malaysia.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Heo, C.C., Teel, P.D., OConnor, B.M. et al. Acari community in association with delayed pig carrion decomposition. Exp Appl Acarol 85, 223–246 (2021). https://doi.org/10.1007/s10493-021-00676-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-021-00676-6