
Abstract
An inventory of the species in and around the clove agroforestry systems (AFS) on the east coast of Madagascar revealed that none of the currently grown species derive from the native Malagasy biodiversity: the herbaceous and woody plants which comprise planned agrobiodiversity, i.e. are deliberately planted by the farmer, are without exception, introduced species. The associated agrobiodiversity, including plants that grow spontaneously and are protected or tolerated by farmers, is composed of a mix of native and introduced species. This situation, which is paradoxical for an island with a very high level of endemicity is the result of a double process of species introduction. The first is associated with the waves of migration, which, during the past three thousand years or so, allowed colonisation of the island, previously devoid of human occupants. The migrant populations originating from Austronesia and then from Africa, introduced their own crops and associated growing practices and only exploited local biodiversity through gathering, which did not require planting or tending. This situation is therefore the result of a long process of farmers' strategies to manage natural resources in a context of subsistence agriculture. The second process is more recent and is associated with increasing occupation by Europeans from the 16th/eighteenth century onwards, which peaked at the end of the nineteenth century with the French colonisers, who introduced and then developed cash crops (coffee, cloves, vanilla). This long-term retrospective study sheds light on the dynamics at the origin of the biodiversity "melting pot" visible today in the AFS along the east coast of Madagascar. It shows the constant evolution and adaptation of farmers' strategies and cropping systems, but also their historical roots. The cohabitation between introduced and indigenous species, observed today in Madagascar, is found in other AFSs built around an introduced cash crop, in various tropical zones. Knowing the origins and determinants of this cohabitation could be useful to support future evolutions of these cropping systems in the current context of changing agricultural practices in response to ecological and macroeconomic pressures.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Madagascar has a rich original biological heritage with a particularly high level of endemicity. Goodman and Benstead (2005) estimated the plant species to be 83% endemic (i.e., found only in Madagascar) or native (not only originating in Madagascar). This biodiversity is threatened by human practices, particularly deforestation and climate change (Vieilledent et al. 2018). Madagascar is therefore considered to be a biodiversity hotspot (Myers et al. 2000). However, this statement does not account for the wealth of agrobiodiversity present in agricultural areas (Chirwa et al. 2008; Kull et al. 2013). According to the definition by Qualset et al. (1995), Vandermeer and Perfecto (1995), or Jackson et al. (2007), agrobiodiversity includes planned crops and livestock species (i.e., chosen and established by the farmer). Many non-domesticated resources are also available in cultivated areas, termed associated agrobiodiversity. Some of these non-domesticated resources are useful to humans. Some are non-productive species that support food production (honey plants, pollinators, symbionts, fertilising plants, and all organic control agents). This associated diversity also includes all the elements that spontaneously colonise the agroecosystem, such as microorganisms and fauna in the soil, weeds, herbivores, carnivores, and parasites, which may sometimes be considered as bio-aggressors, and have a negative economic impact on the farmer.
This agrobiodiversity is partly indigenous (native or endemic), but also includes a pool of introduced species. According to the definitions of Richardson et al. (2000), a species is introduced if humans carried the species or its propagules across major geographical barriers the species would otherwise have been unable to cross. An introduced species is considered naturalised when the biotic and abiotic barriers to its reproduction are removed. A naturalised species is said to be invasive if it has a high capacity for propagation.
Early works already established the mixture of introduced and indigenous species in the Malagasy biodiversity: Prudhomme (1902), Heckel (1910), Perrier de la Bâthie (1928; 1931/1932), Decary (1946a) drew up the first lists of plants and animals introduced into Madagascar. Based on these lists, Kull et al. (2012) estimated that about 10% of the plant species present in Madagascar today are introduced and that at least 70% of them are useful. In addition, a feature shared by the three main food crops (rice, maise, cassava) and the three main cash crops (clove, lychee, vanilla), which respectively contribute to the food security and economic stability of Madagascar (Gouzien et al. 2016), is that none are native to Madagascar. In general, as described by Kull et al. (2013) using the concept of a biodiversity melting pot, introduced species share the same spaces with indigenous species in Malagasy ecosystems, contribute to ecosystem services and rural well-being or, conversely, have a mixed or negative impact.
The main point this article addresses concerns the origin of agrobiodiversity present in Madagascar and more particularly in the cropping systems along the east coast which was analysed by the authors of a series of previous works (Arimalala et al. 2019; Michel et al. 2021; Mariel et al. 2021, 2022).
In this region, hillside cropping systems are predominantly clove-based agroforests (AFS) that are on a continuum between mono-species clove plantations, parks in which clove is often the dominant species, and complex agroforests (Arimalala et al. 2019). Fallow areas surround these clove-based systems, which vary in density and diversity, and rice cultivation in the lowlands (Mariel et al. 2021, 2022). Clove-based cropping systems are complex and rich in biodiversity (Arimalala et al. 2019; Michel et al. 2021; Mariel et al. 2022). The part played by native and introduced species, the origin of these species, and the determinants of their introduction or their maintenance in the cropping system have not been studied to date.
The purpose of this study is first to provide an inventory of the main vascular plant species that characterises agrobiodiversity in and around clove AFS to assess the proportion of introduced and native species. Second, the processes that led to this agrobiodiversity are analysed through a close examination of the literature, particularly historical documentation and iconography.
Materials and methods
The study was based on three recent botanical inventories in clove-based cropping systems in the southern part of Madagascar's Analanjorofo region. The first inventory concerned 42 fields with clove trees in the rural communes of Ambatoharanana and Ambodimanga II (Fénérive-Est district) previously analysed by Arimalala et al. (2019). In each field, a plot of approximately 900 m2, selected as being representative as possible of the entire area, was traced, and surveys of the presence/absence of plant species were made. In the study conducted by Michel et al. (2021), the same protocol was applied to woody species in a sample of 30 plots. The third data set was collected during field visits with farmers in the rural village of Vohibary (Vavatenina district) in an inventory of 17 plots (Mariel et al. 2021).
These inventories are probably not exhaustive, herbaceous (Poaceae) and pteridophytes are probably underrepresented, but are sufficiently detailed to serve as a basis for the presentstudy.
The identity (vernacular and scientific name) of each plant was established. When identification was doubtful, a sample was taken to be identified by comparison with reference specimens in Madagascar's national TAN herbarium (Tsimbazaza Botanical and Zoological Park) and in the TEF herbarium (FOFIFA's Forest Research and Natural Resource Management Department) in Antananarivo.
The species identified were assigned to three main categories based on the knowledge and practices obtained in interviews with the farmers and according to the definitions proposed by Qualset et al. (1995), Vandermeer and Perfecto (1995) and Jackson et al. (2007): (i) species planted and tended by farmers; (ii) spontaneously occurring species, conserved or tolerated by farmers in and around their plots; (iii) weeds or invasive species that are neither planted, nor favoured, nor tolerated, but unintentionally supported by farmers because they are difficult or costly to control and eradicate. This classification mainly reflects the agricultural/nutritional value of the plants expressed by the farmers in our study area, it can be refined by accounting for the non-agricultural uses of the plants (for medicinal purposes, as wood for construction, fuelwood, dye, etc.) or their level of abundance in the cultivated areas.
Within each group, the species were assigned to one of two categories according to the stratum they occupy in the field: (i) herbaceous plants that form the lower stratum, generally less than two metres high (Arimalala et al. 2019) and (ii) large, mostly woody species that form the upper stratum.
The cropping system in which they grow corresponds to the categories specified by Arimalala et al. (2019): (i) M: mono-species clove tree plantations (monoculture), with no accompanying cultivated species, but sometimes used for grazing zebu cattle; (ii) P: tree covered parkland where clove trees are associated with a herbaceous crop (rice, sugar cane, maize, etc.) and other cultivated woody plants; (iii) C: complex AFS where the clove trees are associated with frequently cultivated herbaceous and woody species and less frequently with forest species; (iv) T: fallow or tavy (slash-and-burn farming practice) area; (v) L: lowland.
The status of each species was determined according to the following three classes, based mainly on the studies of Perrier de la Bâthie (1931/1932), Decary (1946a) and Beaujard (2011): (i) EN: endemic (exclusively Malagasy) or native (presence extending beyond Madagascar); (ii) IF: introduced formerly and naturalized, i.e. capable of maintaining itself spontaneously (Richardson et al. 2000); (iii) IN: at an undetermined date and naturalised; (iv) IR: introduced recently (since the eighteenth century and the significant presence of Europeans).
Finally, the geographical area of origin of the introduced species is reported based on bibliographical sources. Bibliographical and iconographic data were sourced from a body of ancient and recent documents. The older studies were published by agronomists and botanists working in Madagascar since the early nineteenth century. These source documents are generally in French.
Results
Table 1 is a compilation of the main species identified in cropping systems with integrated clove agroforests in the Fénérive-Est and Vavatenina districts.
The first observation is that the agrobiodiversity present in and around the AFS (as illustrated in Fig. 1A–C) consists predominantly of introduced species: 18 of the 23 species or genera of herbaceous plants listed in Table 1 are introduced, only five are endemic or native. In the case of woody plants, the proportion is 42 introduced species out of 56.

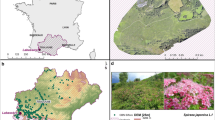
Landscape of the east coast of Madagascar : A: near the village of Ambatoharanana, lanscape combining introduced [clove (A), lychee (B), bread fruit (C), eucalyptus (D), coconut tree (E), Albizia spp (F), banana (G), rice (H)] and native [raffia (Z), bamboo (Y), traveller’s tree (X)] species (photo: E Penot, 2014, © CIRAD). B: Rice field and clove-based agroforestry systems close to Vavatenina with introduced species [clove (A), lychee (B), bread fruit (C), eucalyptus (D), rice (H), jackfruit (I), mango tree (J)] and native species [ bamboo (Y), traveller’s tree (X), raffia (Z)] (photo: E Penot, 2016). C: AFS with clove trees (Sainte-Marie Island) where introduced cultivated herbaceous species [banana (G), manioc (M), pineapple (N)], endemic herbaceous species [Stenotaphrum dimidiatum (W)] and introduced herbaceous species [Clidemia hirta (O)] were identified; the upper stratum is composed of introduced cultivated species [clove (A), coconut (E), papaya (K), mandarin (L),] and endemic species (Dracanea reflexa (V)] (photo: P Danthu, 2012)

Historical milestones of native and introduced biodiversity practices and uses along the east coast of Madagascar (based on de Flacourt 1661, reprinted 2007; Deschamps 1961; Radt 1970; Le Bourdiec 1974; Allibert 1988; Adelaar 1989; Burney et al. 2004; Campbell 2005; Beaujard 2011; Dewar and Wright 1993; Dewar 2014; Crowther et al. 2016; Pierron et al. 2017; Godfrey et al. 2019)

Some examples of plant introduction trials or use of introduced species in the early 20th century on the east coast of Madagascar. A: Experimental plantation at Ivoloina station of 18-month-old kapok trees (Ceiba pentandra, synonym: Eriodendron anfractuosum) originally intended both as fibre producers (kapok) and as support trees for pepper vines, but the excessive shade produced proved harmful to the pepper plants (Anonymous 1904). B: Coffee tree shade trials with two introduced species: Hura crepitans (native to the tropical regions of America) and Albizia falcataria. 15-month old plantation at Ivoloina station (Anonymous, 1904). C: Test tapping of specimen rubber tree introduced in the Ivoloina station at the beginning of the 20th century, but which was not followed up (Foiben-Taosarintanin'I Madagasikara collection, Antananarivo, © FTM). D: Recently introduced vanilla crop supported by an old introduced species, banana (photo: E Prudhomme, circa 1900, Nosy Be) (CIRAD, CD_PV2/170; https://numba.cirad.fr/ark:/12148/btv1b10103466n). Banana was often associated with coffee and cocoa trees on the east coast to provide shade in their early years (Deslandes 1903). E: An example of cash crop developed during French colonisation involving two species introduced to Madagascar in the 18th century: coffee trees under albizia, possibly Albizia falcataria. In the first year of planting, young coffee trees are sometimes shaded by large introduced herbaceous species (banana, maize, or cassava) used as intercrops (Deslandes 1903; François and Ledreux 1929). Coffee cultivation was profitable until the 1960s and 1970s, but then declined (photo: A Fauchère, 1903, Ivoloina station) (CIRAD, CD_PV50/2257 and CD_PV47/1986; https://gallica.bnf.fr/ark:/12148/btv1b101023802). F: Cocoa tree nursery established under permanent shelter in Ivoloina station. Note the presence of breadfruit trees (Artocarpus altilis) and eucalyptus (Anonymous 1904)
The second observation is that all the cultivated species, whether herbaceous (12 species) or woody (22 species), are introduced species. Poaceae, Myrtaceae, Annonaceae, Rubiaceae, Sapindaceae, Moraceae and Rutaceae are the most widely represented botanical families. Some, incuding rice, cassava or coconut, were introduced and naturalised a long time ago, whereas others, such as vanilla, clove, lychee or mango, were introduced as recently as the nineteenth century (Perrier de la Bâthie 1931/1932; Decary 1946a; Bouriquet 1946; Beaujard 2011).
The third observation concerns non-cultivated species for which our inventories show a balance between native and introduced species. Amongst the farmed species, 17 are native and 19 introduced, whilst the ratio for weeds is two to six. Asteraceae, Fabaceae, Myrtaceae and Poaceae are the most represented families in this portion of agrobiodiversity.
The fourth observation, a corollary of the second, is that no indigenous species are cultivated. They are disseminated in uncultivated or fallow areas, as well as in the AFSs, where they are managed by the farmer (Mariel et al. 2021). This is the case, for example, of Ravenala madagascariensis, Harungana madagascariensis or Raphia farinifera. They belong to the associated agrobiodiversity defined by Vandermeer and Perfecto (1995) and Jackson et al. (2007).
Discussion and conclusions
The results of our study (Table 1) show that the cropping systems along the east coast of Madagascar result from the cohabitation of native and introduced species creating what is defined by Kull et al. (2013) as “melting pots” of biodiversity. However, our study reveals a previously unreported detail concerning the relative composition of native and introduced species by showing that the cultivated species in the growing systems derive entirely from introduced species.
This observation, which is paradoxical for a country with a notably high degree of biodiversity endemism (Goodman and Benstead 2005), is in agreement with that of Perrier de la Bâthie (1931/1932) who noted that: “In the same way as all tropical countries where the primitive vegetation has been more or less radically destroyed by humans, Madagascar has been overrun, as the ancient vegetation regressed, by a host of plants alien to its flora. Some of these incomers, of which certain are useful to man, were introduced by him intentionally; others, commensals of the human species or domestic animals, or field parasites, followed him as they follow him everywhere.” Haudricourt (1948) put forward a different hypothesis, in which he explained the Malagasy paradox by the fact that “Madagascar has a rich flora which could have spawned useful cultivated plants, yet humans arrived quite late on this island, with his knowledge of farming and a stock of cultivated plants”. The latter view is supported by several studies involving various scientific disciplines (palaeoecology, palynology, human genetics, and linguistics). Despite divergences, these studies show that the colonisation of Madagascar by humans is relatively recent, starting at the most two or three thousand years ago (Burney et al. 2004; Pierron et al. 2017), and that humans began to significantly inhabit the island less than a thousand years ago (Fig. 2). Two major actors were involved: Austronesians, from what is now Borneo and Java, were the first to arrive, followed by the Bantus, who crossed the Mozambique Channel (Adelaar 1989; Straka 1996; Burney et al. 2004; Beaujard 2011; Pierron et al. 2017), probably arriving in successive waves (Beaujard 2007) (Fig. 2). These migrants brought with them species they already knew how to grow (Beaujard 2007), which would explain why these newcomers did not set about domesticating the local species. The Austronesian populations also imported the slash-and-burn farming practice known as tavy, which resembles the one practised by the Southeast Asian farmers (Kiener 1963; Ketterings et al. 1999). Despite being banned for a century (Bertrand and Randrianaivo 2003), tavy is still practised on the east coast but has been progressively abandoned since the middle of the twentieth century (Mariel et al. 2022). The native elements of flora and fauna were only picked (fruit, honey, medicinal plants, fibres, dyes), felled (timber, wood), fished or hunted. It was not until Madagascar became a French colony in 1896, that processes involving farming and domestication of native and endemic Malagasy species were considered (Tourte, 2019). Botanists and agronomists were then instructed to draw up “the inventory of plant and animal productions that Madagascar was likely to offer to attract farmers, whether they be colonial newcomers or long-standing Malagasy inhabitants” (Prudhomme, 1908) even if most local species were judged to be of poor quality (Deslandes 1903). The botanists then set about exploring the local plant resources and their potential for cultivation. From this resource, Perrier de la Bâthie (1924) published a list of edible or useful fruits, with a potential for domestication, as he said “Amongst the multitude of plants that cover the Grande Île, a large proportion produces fruits which are consumed or used by humans. Most of these species vary in shape, size, flavour or the other properties of fruit. So that selecting these fruits, improving them, bending the plants that produce them towards cultivation and distributing the species would seem to be as useful a work as any, likely to produce useful results not only for Madagascar but also all the tropical regions.” This list contains raffia (Raphia farinifera), but also Flacourtia ramontchi, Rubus rosaefolius and even Tristemma virusanum whose presence have been confirmed in the AFS along the east coast (Styger et al. 1999). The same survey was carried out on aromatic plants by Gatefossé (1921) and Decary (1955) who pointed to the potential of several native species of the east coast, such as Aframomum augustifolium, Vespris madagascarica (synonym: Pelea madagascarica), and Danais fragrans, whose aromatic interest may be equal to that of several introduced plants, such as nutmeg, patchouli, ylang-ylang or champak. However, the cultivation of these species was not developed anywhere in Madagascar and no market for them was ever pursued. These plants were rapidly abandoned, and are not cultivated at all today. As gathered produce, they do little more than fulfil the personal medicinal needs of local populations (Pernet et Meyer 1957) or feed small production sectors, particularly of essential oils whose development is recent, linked to the expansion of aromatherapy.
The few plant products obtained from native and endemic species that have given rise to export sectors are sourced entirely by harvesting them in their natural habitat. For instance, at the beginning of the twentieth century, this was the case for different natural rubber species belonging amongst others to the Mascarenhasia and Landolphia genera (Danthu et al. 2016), raffia palm (Raphia farinifera) (Perrier de la Bâthie 1931; Mouranche 1955), precious woods (Dalbergia spp., Diospyros spp.) (Perrier de la Bâthie 1950; Schurman et Lowry II 2009), copal, the extracted resin of Trachylobium verrucosum (Jacob de Cordemoy 1911), and in recent years, the bark of Prunus africana (Stewart 2003), which has led to the rarefaction of them all, and in some case, even threatened the species concerned.
Most of the species cultivated along the east coast today are formerly introduced plants transported there by successive human migrations, such as rice, cassava, yam, maize and coconut, which then spread across the island progressively over the centuries, (Beaujard 2011). Some originate from Africa, like coffee (Coffea arabica), Cola nitida and banana (Musa spp.). The route taken by banana is not clearly established but the species is thought to have originated in Asia and transited via East Africa (Lejju et al. 2006; Beaujard 2011; Perrier et al. 2011). Oryza sativa, Cocos nucifera, Curcuma longa, Artocarpus heterophyllus, Colocasia esculenta, Litchi chinensis, Cinnamomum verum, Saccharum officinarum are all of Asian origin, whilst others originated in Oceania (Artocarpus altilis and Spondias dulcis) or the Americas (Carica papaya, Persea americana, Annona muricata, Psidium cattleianum, Pachira aquatica, Schinus terebinthifolius) (Perrier de la Bâthie 1931/1932; Evreinoff 1950; Maistre 1964; Radt 1970; Fouqué 1972; Le Bourdiec 1974; Styger and Rakotoarimanana 1996; Beaujard 2011; Kull et al. 2012; Crowther et al. 2016). They partly arrived with the waves of human migration that form the Malagasy population today, the most well-documented being the two Asiatic waves between the third and fifth centuries. Then in the twelfth century, regular but less intense African (Bantu) migrations arrived via the west coast and also, from the ninth century on, through exchanges with Arabian sailors (Beaujard 2011; Pierron et al. 2017) (Fig. 2). Some introduction patterns, like banana, are the subject of debate. This is also the case of the coconut tree, which Beaujard (2011) considered ancient and accompanying the first Austronesians, whereas Delorme (1972) considered it to be more recent, i.e., in the eleventh and twelfth century, and brought by Arabians, without excluding the possibility of sea transport due to the natural floating capacity of coconuts (Gerbaud 2011).
A second more thoroughly documented introduction period is that related to contacts with Europeans, from the sixteenth century, but especially the eighteenth century on, marked by the establishment of colonial empires (Fig. 2). This consisted of a vast movement to transfer economically important plants between different parts of the world for the purpose of furthering the empires’ prosperity. This movement was characterised by the creation of a network of botanical gardens and acclimatisation stations, particularly in tropical areas (Brockway 1979; Bonneuil and Kleiche 1993). In Madagascar, this period was marked by the creation of the first acclimatisation gardens: Nanisana in the Highlands and Ivoloina on the east coast (Galliéni 1908). Maize and cassava were introduced into Madagascar towards the middle of the eighteenth century. Clove, vanilla and black pepper are considered to have been introduced more recently, the dates being fairly well known: clove around 1827, vanilla around 1870 and black pepper around 1900 (Rollot 1926; Perrier de la Bâthie 1931/1932; Bouriquet 1946; Decary 1946a, 1963; Maistre 1964; Raison 1972; Kull et al. 2012), although some doubt hangs over clove and vanilla, the first attempts to cultivate these species in Madagascar Campbell (2005) date to 1835 and 1825. Lychees, mango and avocado trees were first acclimatised in 1802 by the French botanist André Michaux in Tamatave (François 1927). The first cotton and sugar cane plants were acclimatised and cultivated in Sainte-Marie in the 1820s (Decary 1937), at the same time as cinnamon, camphor, star fruit and guava trees (Ledreux 1932). Eucalyptus trees were introduced extensively from 1857 on with the first plantations located along the road between Tamatave and Tananarive in 1897 (Verhaegen et al. 2011; Chauvet 1968). It should also be noted that many introductions did not lead to the development of an industrial or even local sector: there were many failures or failed attempts. Thus, the introduction of mangosteen (Garcinia mangostana), durian (Durio zibethinus) or Brazil nuts (Bertholletia excelsa) and palm oil trees (Elaeis guineensis) in the Ivoloina station at the beginning of the twentieth century did not result in any development or in rare small-scale developments (François 1927; Montagnac 1961; Sigonney 1966). The same was true for Pisdium cattleianum, Ceiba pentandra, Hura crepitans and Hevea brasiliensis (Fig. 3A–C) (Prudhomme 1899/1900; Danthu et al. 2016).
One determining factor, which had a major impact on the spread and cultivation of species of economic value on the east coast of Madagascar, was the determination of the French colonial power in the first half of the twentieth century to promote cash and export agriculture in order to fulfil the requirements of the metropolis and to achieve financial autonomy for the colonies (Jumelle 1900; Galliéni 1908; Prudhomme 1908, Perrier de la Bâthie 1931/1932; Chevalier 1946; Braudeau 1961; Jacob 1987; Danthu et al. 2014; Cocoual and Danthu 2018; Mariel et al. 2022). Farmers, a few large landowners, but more often 'hard-working peasant farmers' (Koerner 1969), grew these crops as a source of income to pay the many and very heavy taxes on rice fields or cattle (Jacob 1987). Clove, vanilla, coffee or cocoa were thus promoted and developed (Fig. 3D–F).
Among species currently not cultivated but kept by farmers in their AFSs for various uses, our study recorded the presence of indigenous plants and also introduced species. The latter include, for example, Syzygium malaccence and Syzygium cumini (fruit trees), Casuarina equisetifolia (wood, tannins, stakes for vanilla (François 1934), eucalyptus (wood), Albizia spp (A. lebbeck, A. falcataria for wood, shade for coffee trees or firewood (François and Ledreux 1929) (Fig. 3B, E), or Jatropha curcas (stakes for vanilla). Some cocoa trees and coffee trees, promoted by the colonial power, are still kept by farmers in low abundance and for self-consumption (Mariel et al. 2021, 2022; Braudeau 1961). The main native species present in these AFSs are bamboo (Decary 1962; Dransfield 2003), Harungana madagascariensis, Ficus tilifolia or Intsia bijuga. These species are a source of wood for energy or general use and often have an important place in the local pharmacopoeia (Heckel 1910; Kiener 1954; Pernet and Meyer 1957; Blaser et al. 1993). Special mention should be made of Raphia farinifera, which is one of the rare species native to the east coast of Madagascar—even if there are some doubts concerning its nativity, as Dransfield and Beentje (2003) reported that this species is actually introduced—that have been exported since 1860 (Dufournet 1938/1939). Madagascar has, in fact, a virtual world monopoly on raffia fibre, obtained by pulling off ribbon-like strips from the upper surface of the leaflets of young unfolding leaves, formerly used in horticulture and viticulture to tie vine shoots (Perrier de la Bâthie 1931; Decary 1946b; Castel 1947; Mouranche 1955; Deschamps 1961) and now much sought after for handicrafts, including top-end woven sun hats.
Some species are considered as weeds by the farmers, who acknowledge their usefulness for certain purposes. This is the case of Psiadia altissima, an endemic species with medicinal properties, used to produce dye and essential oil (Pernet and Meyer 1957; Danthu et al. 2008; Carrière et al. 2005; Andriamanantena et al. 2019). In this category, Psidium cattleianum, Grevillea banksii and Lantana camara are all introduced (Perrier de la Bâthie 1928, 1931/1932; Decary 1946a; Binggeli 2003; Carrière et al. 2005), and yet it would undervalue them to limit their definition exclusively to their invasive capacity or their impact on the native biodiversity (Kull et al. 2014). For example, Psidium cattleianum was introduced for its tasty edible fruits (Carrière et al. 2008), Litsea glutinosa (invasive but producing wood, fodder, and medicines) (Jacq et al. 2005) and Grevillea banksii is used for its wood and its shade capacity for young clove trees. Farmers mentioned three introduced species as invasive plants: Rubus moluccanus, mistakenly imported in the twentieth century because it was confused with grape vine (Binggeli 2003), Clidemia hirta whose accidental introduction in 1914 into the Ivoloina trial garden from the Buitenzorg Botanical Garden (now Bogor, Indonesia) (Perrier de la Bâthie 1928) and last, Mimosa pudica, which nonetheless has important uses in traditional pharmacopoeia (Pernet and Meyer 1957).
Ravenala madagascariensis is a particular case, this endemic species very prevalent in fallow lands and is used by the Betsimisaraka population as a foodstuff, in traditional remedies, for building huts, and for making baskets, ropes, and for wrapping food. It is a species with both an iconic representation, as well as a high commercial value (Hladik et al. 2002; Rakotoarivelo et al. 2014), but may be considered a weed if it is too abundant in cultivated areas.
Our results (Table 1) also show the presence of a large pool of native or endemic species that farmers protect, conserve, or tolerate in their cropping systems. These species have diverse uses: cover crops, fertiliser, aromatic plants, medicinal plants, dyestuffs, producers of fruit, firewood or timber, shade plants, and often have a strong symbolic and identity value.
However, our results has certain limitations. The first is the non-exhaustiveness of our inventories, as mentioned in Materials and Methods, which probably underestimates the role of associated native or introduced species and the ecosystem services they provide, as shown in other contexts by Altieri (1999) and Gaba et al. (2020). The second limitation is that, in this study, we considered biodiversity only at the species level. If we consider it at an intraspecific level, introduced biodiversity represents only part of species biodiversity, for example, only a few bread fruit (Artocarpus altilis) cultivars are cultivated in Madagascar compared to hundreds in its native range (Deslandes 1903; Ragone 1997). The same is true for yam (Dioscorea alata), Martin and Rhodes (1977) identified at least 235 yam cultivars in its native area, whereas only ten or so have been described in Madagascar (Jeannoda et al. 2003), or clove, whose varieties identified in the Moluccas (Pool 1986; Mahulette et al. 2019) do not seem to be present in Madagascar, where only one type is thought to exist (Maistre 1964). We should also mention the case of banana (Lejju et al. 2006; Perrier et al. 2011), although many varieties are found along the east coast (Deslandes 1903). The narrowness of the genetic base of introduced species can be explained by the fact that current populations are often derived from only a few spawners introduced by humans, as demonstrated by Maistre (1964) or Ragone (1997). A third limitation is that our inventories are restricted to species richness (the number of species) and do not include abundance indicators for each species.
Nevertheless, the ratio of introduced to native species in the agrobiodiversity of growing systems along the east coast of Madagascar remains very much in favour of introduced species. It derives from a long evolutionary process associated successively with the settlement history of Madagascar and subsequently with farmers' strategies for managing natural resources and the development of their production systems. In particular, repeated slash-and-burn cycles strongly disrupt the functioning of the native ecosystem by progressively depleting the soil seed bank, reducing soil fertility, and by reducing the ability of frugivorous vertebrate populations (lemurs, birds) to ensure the dissemination of seeds of native species (Razafindratsima 2014; Morelli et al 2020), or by enabling colonisation by herbaceous species (Aristida similis) or bushy species (Lantana camara, Rubus moluccanus, Psadia altissima, etc.) that will delay and limit recolonisation by trees (initiated by pioneer species such as Harungana madagascariensis or Trema orientalis) (Styger et al. 2007). Those mainly responsible for this evolution were the Austronesian and Bantu migrant populations, who brought their own seeds and associated cultivation practices with them. More recently, this evolution has also been linked to the arrival and presence of Europeans, who continued to introduce plants and created conditions which favoured the development of cash crops to supply external markets (Danthu et al. 2014; Penot and Danthu 2019).
The current cropping systems are therefore the result of the conversion of open or fallow lands previously deforested by cycles of displacement of migrant farmers who imported their crops and associated know-how (Mariel et al. 2022), relayed by the development of cash crops imposed by the colonial power and pursued by the political powers since independence. This chronology of land use may explain why the current biodiversity of SAFs along the east coast of Madagascar is essentially exogenous and contains few endemic elements, unlike other agroforestry systems which are thought to have resulted from the direct conversion of forests, as reported by Martin et al. (2020) and Osen et al. (2021) for forest-derived vanilla agroforests.
Indeed, the relative preponderance of introduced plants in the clove systems along the east coast is found in other Malagasy regions such as the cocoa-growing area of Sambirano (Descroix et al. 2016), or the vanilla-growing area of Sava (Bouriquet 1946; Kahane et al. 2008; Martin et al. 2020). This also concerns other ecological contexts across Madagascar: the Malagasy Highlands, where rice, cassava, peanuts, sugarcane, eucalyptus, and pine trees predominate among cultivated fields and hills (tanety) (Donque 1974; Chauvet 1968; Carrière and Randriambanona 2007); the northwest (where the main crops are cotton, peanuts, tobacco, ylang-ylang, cocoa); Sakalava region (maize, cassava, lima beans) (Donque 1974). The relative preponderance of introduced plants is also found in other island contexts, for example in the islands of the Mascarene Archipelago, the Seychelles or the Comoros, which have a similar prehistory and history to Madagascar: late human colonisation, European and Arab presence in the last few centuries, and then annexation to colonial empires (French or English) that directed their agriculture towards serving distant metropolises (Doumenge 1987; Lutz and Wils 1994; Tassin et al. 2006; Beaujard 2007; Sandron 2007; Allibert 2015; Halidi 2018). These contexts remain to be fully documented.
In other tropical situations, introduced plants are associated with indigenous plants that may remain numerous, fulfill many functions, including supplying local markets, especially with fruit. This is the case in countries where new products were introduced during the colonial occupation (cocoa, coffee, rubber) in old agrarian communities developing complex agroforestry systems. This is the case of AFS inclduing rubber trees in Ivory Coast (Snoeck et al. 2013) or jungle rubber in Southeast Asia, where farmers combine imported species (hevea, coffee, pepper) with many native species: tea, rattan (Calamus manan), durian (Durio zibethinus), rambutan (Nephelium lappaceum), jengkol (Archidendron pauciflorum), petai (Parkia speciosa), duku (Lansium domesticum), which are important on the local market (Penot and Ollivier 2009; Warren-Thomas et al. 2020). Examples can also be found in the Americas with coffee systems (Cerdán et al. 2012) and in Africa with cocoa trees. In particular, cocoa cultivation systems in Cameroon are rich in both introduced species (avocado, citrus, mango, etc.) and native species: safflower (Dacryodes edulis), kola nut (Cola nitida), oil palm (Elaeis guineensis), njangsang (Ricinodendron heudelotii), banana, and Ceiba pentandra, Terminalia superba, etc. (Saj et al. 2017; Jagoret et al. 2018).
In a global context in which agricultural practices are changing rapidly in response to climatic and macroeconomic pressures (linked to international markets) and where the ecological transition is an important factor, our study in Madagascar opens up new avenues of research for understanding stakeholder strategies and the way networks operate to disseminate the information and plant material that feed "melting-pots" of biodiversity. Thus, if the hypothesis of greater plasticity of non-native species to global warming is confirmed (Zettlemoyer et al. 2019), the preponderance of introduced species in cropping systems could be an influential element in the resilience of these cropping systems, with effects that could be either positive or negative. In this context, our results raise wider questions: What is the current state of farmers' knowledge and know-how? What means were used to develop and transmit them? In the current context in which natural forests are being replaced by largely anthropised areas, several issues emerge: Is the pool of knowledge and know-how related to native species under threat? To what extent is it progressively being abandoned? What could replace it and why? To what extent does the loss of local knowledge of native plants compromise future uses, which could develop to meet new challenges? What are the problems involved in preserving native and introduced species?
As we have seen, questions concerning the cohabitation of indigenous and introduced plants and the resulting hybridisation of knowledge in the cropping systems found along the east coast of Madagascar have a generic value that could be applied in many other tropical contexts.
References
Adelaar KA (1989) Malay influence on Malagasy: linguistic and culture-historical implications. Ocean. Linguist 28:1–46. https://doi.org/10.2307/3622973
Allibert C (1988) Les contacts entre l'Arabie, le Golfe persique, l'Afrique orientale et Madagascar : confrontation des documents écrits, des traditions orales et des données archéologiques récentes. In: L'Arabie et ses mers bordières. I. Itinéraires et voisinages, Maison de l'Orient et de la Méditerranée, Lyon, pp 111–126
Allibert C (2015) L’archipel des Comores et son histoire ancienne. Essai de mise en perspective des chroniques, de la tradition orale et des typologies de céramiques locales et d’importation. Afriques 6:1–49. https://doi.org/10.4000/afriques.1721
Altieri MA (1999) The ecological role of biodiversity in agroecosystems. Agric Ecosyst Environ 74:19–31. https://doi.org/10.1016/S0167-8809(99)00028-6
Andriamanantena M, Danthu P, Cardon D, Fawbush FR, Raonizafinimanana B, Razafintsalama VE, Rakotonandrasana SR, Ethève A, Petit T, Caro Y (2019) Malagasy dye plant species: a promising source of novel natural colorants with potential applications-a review. Chem Biodivers 16:e1900442. https://doi.org/10.1002/cbdv.201900442
Anonymous (1904) Rapport annuel 1903. Direction de l’Agriculture - Madagascar et Dépendances. Tananarive, Madagascar. Bibliothèque Historique du CIRAD, Nogent-sur-Marne. Cote BH_OU5008
Arimalala N, Penot E, Michels T, Rakotoarimanana V, Michel I, Ravaomanalina H, Roger E, Jahiel M, Leong Pock Tsy JM, Danthu P (2019) Clove based cropping systems on the east coast of Madagascar: how history leaves its mark on the landscape. Agrofor Syst 93:1577–1592. https://doi.org/10.1007/s10457-018-0268-9
Beaujard P (2007) East Africa, the Comoros Islands and Madagascar before the sixteenth century. On a neglected part of the World-System. Azania 42:15–35
Beaujard P (2011) The first migrants to Madagascar and their introduction of plants: linguistic and ethnological evidence. Azania 46:169–189. https://doi.org/10.1080/0067270X.2011.580142
Bertrand A, Randrianaivo D (2003) Tavy et déforestation. In: Aubert S, Razafiarison S, Bertrand A (eds). Déforestation et systèmes agraires à Madagascar - Les dynamiques des tavy sur la côte orientale, Cirad/Cite/Fofifa, Montpellier and Antananarivo, pp 9–30
Binggeli P (2003) Introduced and invasive plants. In: Goodman S, Benstead JP (eds) The natural history of Madagascar. The University of Chicago Press, Chicago and London, pp 257–268
Blaser J, Rajoelison G, Tsiza G, Rajemison M, Rabevohitra R, Randrianjafy H, Razafindrianilana N, Rakotovao G, Comtet S (1993) Choix des essences pour la sylviculture à Madagascar. Akon'ny Ala Numéro Spécial (Tome 1 and Tome 2), ESSA, Antananarivo, Madagascar.
Bonneuil C, Kleiche M (1993) Du jardin d'essais colonial à la station expérimentale 1880–1930. CIRAD, Montpellier, France
Bouriquet G (1946) Le vanillier et la vanille à Madagascar. Revue Internationale de Botanique Appliquée et d'Agriculture Tropicale 286bis:398–404. https://doi.org/10.3406/jatba.1946.1981
Braudeau J (1961) L’extension et l’amélioration de la culture du cacaoyer à Madagascar. Café Cacao Thé 5:82–101
Brockway LH (1979) Science and colonial expansion: the role of the British Royal Botanic Gardens. Am Ethnologist 6:449–465. https://doi.org/10.1525/ae.1979.6.3.02a00030
Burney DA, Pigott Burney L, Godfrey LR, Jungers WL, Goodman SM, Wright HT, Jull AJT (2004) A chronology for late prehistoric Madagascar. J Hum Evol 47:25–63. https://doi.org/10.1016/j.jhevol.2004.05.005
Campbell G (2005) An economic history of imperial Madagascar, 1750–1895. Cambridge University Press, New York
Carrière SM, Andrianotahiananahary H, Ranaivoarivelo N, Randriamalala J (2005) Savoirs et usages des recrus post-agricoles du pays Betsileo : valorisation d’une biodiversité oubliée à Madagascar. VertigO 6. https://doi.org/10.4000/vertigo.3047
Carriere SM, Randriambanona H (2007) Biodiversité introduite et autochtone : antagonisme ou complémentarité ? Le cas de l’eucalyptus à Madagascar. Bois et Forêts des Tropiques 292:5–21. https://doi.org/10.19182/bft2007.292.a20349
Carrière SM, Randrianasolo E, Hennenfent J (2008) Aires protégées et lutte contre les bioinvasions : des objectifs antagonistes ? Le cas de Psidium cattleianum Sabine (Myrtaceae) autour du parc national de Ranomafana à Madagascar. VertigO. https://doi.org/10.4000/vertigo.1918
Castel R (1947) Commerce extérieur. Madagascar L’Encyclopédie Coloniale et Maritime tome deuxième. Encyclopédie de l’Empire Français, Paris, pp 101–114
Cerdán CR, Rebolledo MC, Soto G, Rapidel B, Sinclair FL (2012) Local knowledge of impacts of tree cover on ecosystem services in smallholder coffee production. Agric Syst 110:119–130. https://doi.org/10.1016/j.agsy.2012.03.014
Chauvet B (1968) Inventaire des espèces forestières introduites à Madagascar. Université de Tananarive, Tananarive
Chevalier A (1946) L’évolution de la culture du caféier à Madagascar. Revue Internationale de Botanique Appliquée et d'Agriculture Tropicale 286:390–398. https://doi.org/10.3406/jatba.1946.1980
Chirwa PW, Akinnifesi FK, Sileshi G, Syampungani S, Kalaba FK, Ajayi OC (2008) Opportunity for conserving and utilizing agrobiodiversity through agroforestry in Southern Africa. Biodiversity 9:5–48. https://doi.org/10.1080/14888386.2008.9712881
Cocoual M, Danthu P (2018) Le giroflier à Madagascar: essai d’histoire et de géographie coloniales, 1896–1958. Revue de Géographie Historique 12. http://rgh.univ-lorraine.fr/articles/view/96. Accessed 29 February 2020.
Crowther A, Lucas L, Helm R, Horton M, Shipton C, Wright HT, Walshaw S, Pawlowicz M, Radimilahy C, Douka K, Picornell-Gelabert L, Fuller DQ, Boivin NL (2016) Ancient crops provide first archaeological signature of the westward Austronesian expansion. PNAS 113:6635–6640. https://doi.org/10.1073/pnas.1522714113
Danthu P, Penot E, Ranoarisoa KM, Rakotondravelo JC, Michel I, Tiollier M, Michels T, Normand F, Razafimamonjison G, Fawbush F, Jahiel M (2014) The clove tree of Madagascar: a success story with an unpredictable future. Bois et Forêts des Tropiques 320:83–96. https://doi.org/10.19182/bft2014.320.a20547
Danthu P, Rakotobe M, Mauclère P, Andrianoelisoa H, Behra O, Rahajanirina V, Mathevon B, Ralembofetra E, Collas de Chatelperron P (2008) Essential oil production increases value of Psiadia altissima fallows in Madagascar’s eastern forests. Agrofor Syst 72:127–135. https://doi.org/10.1007/s10457-007-9067-4
Danthu P, Razakamanarivo H, Deville-Danthu B, Razafy Fara L, Le Roux Y, Penot E (2016) The short and forgotten history of rubber in Madagascar: the first controversy between biodiversity conservation and natural resource exploitation. Bois et Forêts des Tropiques 328:27–43. https://doi.org/10.19182/bft2016.328.a31300
Decary R (1937) Les débuts de la colonisation agricole à l’Ile Sainte-Marie de Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale 17:610–618. https://doi.org/10.3406/jatba.1937.5775
Decary R (1946a) Plantes et animaux utiles de Madagascar. Annales du Musée Colonial de Marseille 4:5–234
Decary R (1946b) Plantes textiles introduites ou spontanées à Madagascar. Revue Internationale de Botanique Appliquée et d'Agriculture Tropicale 286bis:454–459. https://doi.org/10.3406/jatba.1946b.1987
Decary R (1955) Quelques plantes aromatiques et à parfum de la Flore de Madagascar. Journal d'Agriculture Tropicale et de Botanique Appliquée 2:416–422. https://doi.org/10.3406/jatba.1955.2237
Decary R (1962) Les emplois du bambou à Madagascar. Journal d'Agriculture et de Botanique Appliquée 9:65–70. https://doi.org/10.3406/jatba.1962.2657
Decary R (1963) Plantes introduites à Madagascar et toponymie. Journal d'Agriculture et de Botanique Appliquée 10:204–218. https://doi.org/10.3406/jatba.1963.2706
Delorme M (1972) Le Cocotier à Madagascar. Oléagineux 27:357–361
Deschamps H (1961) Histoire de Madagascar. Mondes d’Outre-Mer.Berger-Levrault, Paris
Descroix F, Rakotomalala JJ, Bousquet E, Rahaingosambatra DH, Chen-Yen-Su A (2016) Les cacaoyères du Sambirano - Des produits à fort potentiel qualitatif insuffisamment valorises. The cocoa of Sambirano - Products with high quality potential under-valued. In: Duchaufour H (ed) Recherche interdisciplinaires pour le développement durable et la biodiversité des espaces ruraux malgaches. FSP Parrur, Antananarivo, pp 19–55
Deslandes R (1903) Légumes et fruits à Madagascar dans la circonscription agricole de l’Est. Bibliothèque d’Agriculture Coloniale, Librairie Maritime et Coloniale, Augustin Challamel Editeur, Paris, pp 5–40
Dewar RE (2014) Early human settlers and their impact on Madagascar’s landscapes. In: Scales IR (ed) Conservation and environmental management in Madagascar. Routledge, Abington and New York, pp 44–64
Dewar RE, Wright HT (1993) The Culture History of Madagascar. J World Prehist 7:417–466
de Flacourt E (1661 reprinted 2007) Histoire de la Grande Isle Madagascar. Inalco-Karthala, Paris
Donque G (1974) L'agriculture et l'élevage à Madagascar. Cahiers d'Outre-Mer 106:105–127 and 107:271–300. https://doi.org/10.3406:caoum.1974.2692 and https://doi.org/10.3406/caoum.1974.2703
Doumenge JP (1987) Les Îles Seychelles. Annales de Géographie 533:78–102. https://doi.org/10.3406/geo.1987.20585
Dransfield J (2003) Poaceae, Bambuseae, Bamboos, Palms. In: Goodman S, Benstead JP (eds) The natural history of Madagascar. The University of Chicago Press, Chicago and London, pp 467–471
Dransfield J, Beentje H (2003) Arecaceae, Palms. In: Goodman S, Benstead JP (eds) The natural history of Madagascar. The University Of Chicago Press, Chicago and London, pp 448–457
Dufournet R (1938–1939) Le raphia à Madagascar. L’Agronomie Coloniale 251:134–151; 253:1–8; 254:43–54; 255:76–83.
Evreinoff VA (1950) Le Litchi. Fruits d'Outre-Mer 5:325–333
Fouqué A (1972) Espèces fruitières d’Amérique tropicale. Fruits 27:62–72
François E (1927) La production des fruits à Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale 7:713–724. https://doi.org/10.3406/jatba.1927.4576
François E (1934) Les produits malgaches - La vanille. La Revue de Madagascar 5:39–56
François E, Ledreux A (1929). La culture des caféiers à Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale, 9:241–249 and 314–321. https://doi.org/10.3406/jatba.1929.4727 and https://doi.org/10.3406/jatba.1929.4740
Gaba S, Cheviron N, Perrot T, Plutti S, Gautier JL, Bretagnolle V (2020) Weeds enhance multifunctionality in arable lands in South-West of France. Front Sustain Food Syst 4:71. https://doi.org/10.3389/fsufs.2020.00071
Galliéni JS (1908) Neuf ans à Madagascar. Librairie Hachette et Cie, Paris
Gattefossé J (1921) Les végétaux aromatiques de Madagascar. L’Agronomie Coloniale 46:113–121
Gerbaud P (2011) La culture de la noix de coco. FruiTrop 193:41–44
Godfrey L, Scroxton N, Crowley BE, Burns SJ, Sutherland MR, Perez VR, Faina P, McGee D, Ranivoharimanana L (2019) A new interpretation of Madagascar’s megafaunal decline: the “Subsistence Shift Hypothesis.” J Hum Evol 130:126–140. https://doi.org/10.1016/j.jhevol.2019.03.002
Goodman S, Benstead JP (2005) Updated estimates of biotic diversity and endemism for Madagascar. Oryx 39:73–77. https://doi.org/10.1017/S0030605305000128
Gouzien Q, Penot E, Jahiel M, Danthu P (2016) Le Girofle : poids dans l'économie malgache et place de Madagascar dans le marché mondial. Document de travail AFS4FOOD 18. 17p. http://agritrop.cirad.fr/582468. Assessed 5 February 2020
Halidi A (2018) Comores : colonisation, insularité et imaginaire national. Les Cahiers d'Outre-Mer 278:403–432. https://doi.org/10.4000/com.9692
Haudricourt AG (1948) L’origine des plantes cultivées malgaches. Mémoires de l'Institut Scientifique de Madagascar, Série B 1:143–145
Hladik CM, Blanc P, Hladik A (2002) L’arbre du voyageur. Des usages et de la diffusion horticole du Ravenala. Hommes & Plantes 41:18–27
Heckel E (1910) Les plantes utiles de Madagascar. Annales du Musée Colonial de Marseille 8:5–372
Jackson LE, Pascual U, Hodgkin T (2007) Utilizing and conserving agrobiodiversity in agricultural landscapes. Agric Ecosyst Environ 121:196–210. https://doi.org/10.1016/j.agee.2006.12.017
Jacob G (1987) Gallieni et « l’impôt moralisateur » à Madagascar. Théorie, pratiques et conséquences (1901–1905). Revue Française d’Histoire d'Outre-Mer 277:431–473. https://doi.org/10.3406/outre.1987.2617
Jacob de Cordemoy H (1911) Les plantes à gommes et à résines. Octave Doin et Fils Editeurs, Paris
Jacq FA, Hladick A, Bellefontaine R (2005) Dynamique d’un arbre introduit à Mayotte, Litsea glutinosa (Lauraceae): une espèce envahissante ? Revue d'Ecologie (Terre et Vie) 60:21–32
Jagoret P, Todem Ngnogue H, Malézieux E, Michel I (2018) Trajectories of cocoa agroforests and their drivers over time: Lessons from the Cameroonian experience. Eur J Agron 101:183–192. https://doi.org/10.1016/j.eja.2018.09.007
Jeannoda VH, Jeannoda V, Hladik A, Hladik CM (2003) Les ignames de Madagascar. Diversité, utilisations et perceptions. Hommes & Plantes 47:10–23
Jumelle H (1900) Le cacaoyer sa culture et son exploitation dans tous les pays de production. Auguste Challamel Editeur, Paris
Kahane R, Besse P, Grisoni M, Le Bellec F, Odoux E (2008) Bourbon vanilla: natural flavour with a future. Chron Hortic 48:23–29
Ketterings QM, Tri Wibowo T, van Noordwijk M, Penot E (1999) Farmers’ perspectives on slash-and-burn as a land clearing method for small-scale rubber producers in Sepunggur, Jambi Province, Sumatra, Indonesia. For Ecol Manage 120:157–169. https://doi.org/10.1016/S0378-1127(98)00532-5
Kiener A (1954) Note sur le Hintsy (Intsia bijuga O. Kze.). Bois et Forêts des Tropiques 38:36–42. https://doi.org/10.19182/bft1954.38.a18522
Kiener A (1963) Le « tavy » à Madagascar. Bois et Forêts des Tropiques 90:9–16. https://doi.org/10.19182/bft1963.90.a18888
Koerner F (1969) Décolonisation et économie de plantations. Situation des propriétés européennes à Madagascar. Annales de Géographie 430:654–679
Kull CA, Carrière SM, Moreau S, Rakoto Ramoarantsoa H, Blanc-Pamard C, Tassin J (2013) Melting pots of biodiversity: tropical smallholder farm landscapes as guarantors of sustainability. Environ Sci Policy Sustain Dev 55:6–16. https://doi.org/10.1080/00139157.2013.765307
Kull CA, Tassin J, Carrière SM (2014) Approaching invasive species in Madagascar. Madag Conserv Dev 9:60–70. https://doi.org/10.4314/mcd.v9i2.2
Kull CA, Tassin J, Moreau S, Rakoto Ramoarantsoa H, Blanc-Pamard C, Carrière SM (2012) The introduced flora of Madagascar. Biol Invasions 14:875–888. https://doi.org/10.1007/s10530-011-0124-6
Le Bourdiec F (1974) Hommes et paysages du riz à Madagascar: étude de géographie humaine. Association des géographes de Madagascar, Tananarive, Madagascar.
Ledreux A (1932) Le cannelier à Madagascar. La Parfumerie Moderne 3:179–187
Lejju B, Robertshaw P, Taylor D (2006) Africa’s earliest bananas? J Archaeol Sci 33:102–113. https://doi.org/10.1016/j.jas.2005.06.015
Lutz W, Wils AB (1994) People on mauritius: 1638–1991. In: Lutz W (ed) Population – development – environment. Understanding their interactions in mauritius. Springer-Verlag, Heidelberg, Germany, pp 75–97
Mahulette AS, Hariyadi, Yahya S, Wachjar A, Marzuki I (2019) Morpho-agronomical diversity of forest clove in Moluccas, Indonesia. Hayati-J Biosci 26:156–162. https://doi.org/10.4308/hjb.26.4.156
Maistre J (1964) Le clou de girofle. Les plantes à épices. G.-P. Maisonneuve & Larose, Paris, pp 77–124
Mariel J, Carrière SM, Penot E, Danthu P, Labeyrie V (2021) Exploring farmers’ agrobiodiversity knowledge and management practices: a case study of clove agroforests in Analanjirofo, Madagascar. People Nat 3:914–928. https://doi.org/10.1002/pan3.10238
Mariel J, Penot E, Labeyrie V, Herimandimby H, Danthu P (2022) From shifting cultivation (tavy) to agroforestry systems: a century of changing land use on the East Coast of Madagascar. Agrofor Syst (accepted)
Martin DA, Osen K, Grass I, Hölscher D, Tscharntke T, Wurz A, Kreft H (2020) Land-use history determines ecosystem services and conservation value in tropical agroforestry. Conserv Lett 13:e12740. https://doi.org/10.1111/conl.12740
Martin FM, Rhodes AM (1977) Intra-specific classification of Dioscorea alata. Trop Agr (Trinidad) 54:1–13
Michel I, Lobietti M, Danthu P, Penot E, Velonjara F, Jahiel M, Michels T (2021) Agroforestry innovation by smallholders facing uncertainty: the case of clove-based cropping systems in Madagascar. Eur J Agron 123:126218. https://doi.org/10.1016/j.eja.2020.126218
Montagnac P (1961) Les cultures fruitières à Madagascar. Bulletin de Madagascar 177:105–127
Morelli TL, Smith AB, Mancini AN, Balko EA, Borgerson C, Dolch R, Farris Z, Federman S, Golden CD, Holmes SM, Irwin M, Jacobs RL, Johnson S, King T, Lehman SM, Louis EE Jr, Murphy A, Randriahaingo HNT, Randrianarimanana HLL, Ratsimbazafy J, Razafindratsima OH, Baden AL (2020) The fate of Madagascar’s rainforest habitat. Nat Clim Chang 10:89–96. https://doi.org/10.1038/s41558-019-0647-x
Mouranche R (1955) Le palmier raphia de Madagascar. Bois et Forêts des Tropiques 41:8–22. https://doi.org/10.19182/bft1955.41.a18600
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. https://doi.org/10.1038/35002501
Osen K, Soazafy MR, Martin DA, Wurz A, März A, Ranarijaona HLT, Hölscher D (2021) Land-use history determines stand structure and tree diversity in vanilla agroforests of northeastern Madagascar. Appl Vegetation Sci 4:e12563. https://doi.org/10.1111/avsc.12563
Penot E, Danthu P (2019) Entre « ignorance politique », ingérence et contrôle de l’Etat: quelles politiques publiques pour la filière girofle à Madagascar, aux Comores et à Zanzibar. Éthique et Économique/Ethics and Economics 16:106–114
Penot E, Ollivier I (2009) L’hévéa en association avec les cultures pérennes, fruitières ou forestières : quelques exemples en Asie, Afrique et Amérique latine. Bois et Forêts des Tropiques 301:67–82. https://doi.org/10.19182/bft2009.301.a20407
Pernet R, Meyer G (1957) Pharmacopée de Madagascar. Institut de Recherche Scientifique. Tananarive-Tsimbazaza, Madagascar.
Perrier X, Edmond De Langhe E, Donohue M, Lentfer C, Vrydaghs L, Bakry F, Carreel F, Hippolyte I, Horry JP, Jenny C, Lebot V, Risterucci AM, Tomekpe K, Doutrelepont H, Ball T, Manwaring J, de Maret P, Denham T (2011) Multidisciplinary perspectives on banana (Musa spp.) domestication. PNAS 108:11311–11318. https://doi.org/10.1073/pnas.1102001108
Perrier de la Bâthie H (1924) Sur quelques plantes non cultivées de Madagascar à fruits comestibles ou utiles et sur la possibilité de leur culture. Revue de Botanique Appliquée et d'Agriculture Coloniale 38:652–663. https://doi.org/10.3406/jatba.1924.4224
Perrier de la Bâthie H (1928) Les pestes végétales à Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale 77:36–43. https://doi.org/10.3406/jatba.1928.4034
Perrier de la Bâthie H (1931) Le Raphia de Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale 113:4–17. https://doi.org/10.3406/jatba.1931.4947
Perrier de la Bâthie H (1931–1932) Les plantes introduites à Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale 121:719–729; 122:833–837; 123:920–923; 124:991–999; 125:48–52; 126:128–133; 127:213–220; 128:296–301; 129:372–383; 130:462–468; 131:530–543
Perrier de la Bâthie H (1950) L’Ebène de Madagascar et les arbres qui le produisent. Revue Internationale de Botanique Appliquée et d'Agriculture Tropicale 327–328:38–44. https://doi.org/10.3406/jatba.1950.6291
Pierron D, Heiske M, Razafindrazaka H, Rakoto I, Rabetokotany N, Ravololomanga B, Rakotozafy LMA, Rakotomalala MM, Razafiarivony M, Rasoarifetra B, Andriamampianina Raharijesy M, Razafindralambo L, Ramilisonina FF, Lejamble S, Thomas O, Abdallah AM, Rocher C, Arachiche A, Tonaso L, Pereda-loth V, Schiavinato S, Brucato N, Ricaut FX, Kusuma P, Sudoyo H, Ni S, Boland A, Deleuze JF, Beaujard P, Grange P, Adelaar S, Stoneking M, Rakotoarisoa JA, Radimilahy C, Letellier T (2017) Genomic landscape of human diversity across Madagascar. PNAS 114:E6498–E6506. https://doi.org/10.1073/pnas.1704906114
Pool PA, Eden-Green SJ, Muhammad MT (1986) Variation in clove (Syzygium aromaticum) germplasm in the Moluccan Islands. Euphytica 35:149–159
Prudhomme E (1899/1900) Le caoutchouc sur la côte est de Madagascar. Revue de Madagascar 1:366–384; 2:43–52
Prudhomme E (1902) Les fruits et légumes de Madagascar. Colonie de Madagascar et Dépendances, Bulletin Economique 1:1–35; 2:121–131; 3:215–241; 4:317–321
Prudhomme E (1908) L’agriculture à Madagascar. Bulletin de l'Office Colonial. Supplément au n°5:1–13
Qualset OC, Mc Guire P, Warburton ML (1995) Agrobiodiversity’ key to agricultural productivity. Calif Agric 49:45–49. https://doi.org/10.3733/ca.v049n06p45
Radt C (1970) Aperçu sur l’histoire de la canne à sucre. Journal d'Agriculture Tropicale et de Botanique Appliquée 17:141–147. https://doi.org/10.3406/jatba.1970.3056
Ragone D (1997) Breadfruit. Artocarpus altilis (Parkinson) Fosberg. Promoting the conservation and use of underutilized and neglected crops. 10. Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute, Rome, Italy
Raison JP (1972) L’introduction du manioc à Madagascar : un problème non résolu. Terre Malgache, Tany Malagasy 13:223–228
Rakotoarivelo N, Razanatsima A, Rakotoarivony F, Rasoaviety L, Ramarosandratana AV, Jeannoda V, Kuhlman AR, Randrianasolo A, Bussmann RW (2014) Ethnobotanical and economic value of Ravenala madagascariensis Sonn in Eastern Madagascar. J Ethnobiol Ethnomed 10:57. https://doi.org/10.1186/1746-4269-10-57
Razafindratsima OH (2014) Seed dispersal by vertebrates in Madagascar’s forests: review and future directions. Madagascar Conserv Dev 9:90–97. https://doi.org/10.4314/mcd.v9i2.5
Richardson DM, Pyšek P, Rejmánek M, Barbour MG, Dane Panetta F, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107. https://doi.org/10.1046/j.1472-4642.2000.00083.x
Rollot C (1926) Le manioc à Madagascar. Revue de Botanique Appliquée et d'Agriculture Coloniale 6:81–88. https://doi.org/10.3406/jatba.1926.4374
Salomon JN (1979) Une culture semi-industrielle à Madagascar: les plantes à parfum. Cahiers d’Outre-Mer 126:158–178. https://doi.org/10.3406/caoum.1979.2899
Sandron F (2007) Dynamique de la population réunionnaise (1663–2030). In: Sandron F (ed) La population réunionnaise : analyse démographique. IRD, Paris, pp 27–41
Saj S, Durot C, Mvondo Sakouma K, Tayo Gamo K, Avana-Tientcheu ML (2017) Contribution of associated trees to long-term species conservation, carbon storage and sustainability: a functional analysis of tree communities in cacao plantations of Central Cameroon. Int J Agric Sustain 15:282–302. https://doi.org/10.1080/14735903.2017.1311764
Schuurman D, Lowry PP II (2009) The Madagascar rosewood massacre. Madagascar Conserv Dev 4:98–102. https://doi.org/10.4314/mcd.v4i2.48649
Sigonney M (1966) Le palmier à huile et les perspectives de développement de l’élaéiculture à Madagascar. Terre Malgache, Tany Malagasy 1:221–232
Snoeck D, Lacote R, Kéli J, Doumbia A, Chapuset T, Jagoret P, Gohet E (2013) Association of hevea with other tree crops can be more profitable than hevea monocrop during first 12 years. Ind Crops Prod 43:578–586. https://doi.org/10.1016/j.indcrop.2012.07.053
Stewart KM (2003) The African cherry (Prunus africana): Can lessons be learned from an over-exploited medicinal tree? J Ethnopharmacol 89:3–13. https://doi.org/10.1016/j.jep.2003.08.002
Straka H (1996) Histoire de la végétation de Madagascar oriental dans les derniers 100 millénaires. In: Lourenço WR (ed) Biogéographie de Madagascar. Orstom, Paris, pp 37–47
Styger E, Rakotoarimanana JEM (1996) A la découverte des espèces ligneuses fruitières indigènes de Madagascar. Silo National des Graines Forestières, Antananarivo, Madagascar.
Styger E, Rakotoarimanana JEM, Rabevohitra R, Fernandes ECM (1999) Indigenous fruit trees of Madagascar: potential components of agroforestry systems to improve human nutrition and restore biological diversity. Agrofor Syst 46:289–310. https://doi.org/10.1023/A:1006295530509
Styger E, Rakotondramasy HM, Pfeffer MJ, Fernandes ECM, Bates DM (2007) Influence of slash-and-burn farming practices on fallow succession and land degradation in the rainforest region of Madagascar. Agric Ecosyst Environ 119:257–269. https://doi.org/10.1016/j.agee.2006.07.012
Tassin J, Rivière JN, Bruzzese E (2006) Ranking of invasive woody plants species for management on Réunion Island. Weed Res 46:388–403. https://doi.org/10.1111/j.1365-3180.2006.00522.x
Tourte R (2019) La période colonial et les grands moments des Jardins d’essais (1885–1890 – 1914–1918). L’Harmattan, Paris
Vandermeer J, Perfecto I (1995) Breakfast of biodiversity: the truth about rainforest destruction. Food First Books, Oakland, California, USA
Verhaegen D, Randrianjafy H, Montagne P, Danthu P, Rabevohitra R, Tassin J, Bouvet JM (2011) Historique de l’introduction du genre Eucalyptus à Madagascar. Bois et Forêts des Tropiques 309:17–25. https://doi.org/10.19182/bft2011.309.a20462
Vieilledent G, Grinand C, Rakotomalala FA, Ranaivosoa R, Rakotoarijaona JR, Allnutt TF, Achard F (2018) Combining global tree cover loss data with historical national forest cover maps to look at six decades of deforestation and forest fragmentation in Madagascar. Biol Cons 222:189–197. https://doi.org/10.1016/j.biocon.2018.04.008
Vlassov S (1936) Espèces alimentaires du genre Artocarpus. 1. L’Artocarpus integrifolia L. ou le jacquier. Mémoires de l'Institut Royal Colonial Belge 4:7–80
Warren-Thomas E, Nelson L, Jutong W, Bumrungsri S, Brattström O, Stroesser L, Chambon B, Penot E, Tongkaemkaew U, Edwards DP, Dolman PM (2020) Rubber agroforestry in Thailand provides some biodiversity benefits without reducing yields. J Appl Ecol 57:17–30. https://doi.org/10.1111/1365-2664.13530
Zettlemoyer MA, Schultheis EH, Lau JA (2019) phenology in a warming world: differences between native and non-natives plant species. Ecol Lett 22:1253–1263. https://doi.org/10.1111/ele.13290
Acknowledgements
This study was conducted as part of the research programmes supported by the DP Forêts et Biodiversité in Madagascar, associating the CIRAD, the FOFIFA and the University of Antananarivo. The authors would like to thank Karen Newby and Daphne Goodfellow for the translation of this article.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Danthu, P., Michel, I., Carrière, S.M. et al. Coming from elsewhere: the preponderance of introduced plant species in agroforestry systems on the east coast of Madagascar. Agroforest Syst 96, 697–716 (2022). https://doi.org/10.1007/s10457-022-00732-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-022-00732-z