
Abstract
This study evaluates the potential for pasture improvement and restoration at two silvopastoral sites. We used a total of 16 wild legume species under different management systems (rhizobial inoculation, mycorrhizal inoculation, sheep penning, and sheep penning with mycorrhizal inoculation), at two mountain sites in Sierra Nevada Natural Park (Granada, southern Spain). The first site, Soportújar, had higher soil fertility and rainfall than the second, Lanjarón. Forage yields in Soportújar ranged from 265 to 8970 kg DM ha−1, Vicia amphicarpa, Vicia monantha, Vicia disperma and Lathyrus clymenum being the most productive species. Mycorrhizal inoculation resulted in higher forage yields for Lens nigricans, V. disperma and Vicia lutea. Seed yields were low, ranging from 5.9 to 1234 kg ha−1. Forage yields in Lanjarón were lower, ranging from 46 to 1415 kg DM ha−1; and the most productive species were V. monantha, V. disperma, Lathyrus cicera and Medicago rigidula. Sheep penning alone and together with mycorrhizal treatment resulted in greater forage yields for most of the species studied, although differences were only significant for V. disperma, V. monantha and Lathyrus sphaericus. Seed yields ranged from 0.4 to 60 kg ha−1. In conclusion, we recommend V. monantha, V. disperma and L. cicera followed by V. amphicarpa and L. clymenum (in wetter more fertile sites) and M. rigidula (in drier sites), as they seem to be the best adapted to the pedoclimatic conditions of Sierra Nevada Natural Park. Increasing nutrients in the soil (by sheep penning) and promoting nutrient assimilation (by mycorrhizal inoculation) may be effective strategies for increasing pasture biomass in silvopastoral sites.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the Mediterranean basin, pine woodlands are the most fire-sensitive ecosystems (Pausas et al. 2008). The creation of grazed fuel-breaks, which involves reducing tree density by thinning to facilitate livestock grazing (Ruiz Mirazo and Robles 2012), could be an efficient way to diminish these environmental risks as these practices help to efficiently control vegetation fuel load (Ruiz Mirazo and Robles 2012). In addition, combining grazing with other techniques such as pasture seeding may increase the efficiency of the livestock activity, as they allow extension of the grazing period in such silvopastoral systems (Thavaud 2009).
Rangelands store about half of the global terrestrial carbon, making them key for global climate change mitigation, and moreover, they contain over a third of the global biodiversity hotspots; they are, however, one of the biomes most susceptible to land degradation and global climate change (Board 2005). Legumes are key components of Mediterranean rangelands; even at very dry sites and overgrazed areas, they are able to maintain their populations, especially those with low or prostrate shoots with renewal buds close to or below ground (Porqueddu and González 2006). Additionally, annual legumes are adapted to low rainfall conditions, as well as low winter temperatures and a wide range of soil conditions (Ewing 1999). Annual self-reseeding legumes have great potential for pasture improvement and restoration in semiarid areas for several reasons: (1) they are able to fix nitrogen and, consequently, to help meet the nutritional needs of other plants (Graham and Vance 2003); (2) they have a long permanence in the soil seed bank due to great hardseededness (Arianoutsou and Thanos 1996); and (3) they provide good quality fodder, mainly linked to high protein content (Porqueddu and González 2006). Nevertheless, there has been little research into wild species and forage and seed yield performance or their suitability for pasture improvement and restoration, especially in silvopastoral sites (Porqueddu et al. 2013) and fuel-break areas (Thavaud 2009).
Despite the ready availability of cultivars of some annual legumes, many of them are not suitable for silvopastoral sites, in most cases because they are of non-local provenance. In particular, their utilization in improvement and pasture restoration plans, especially in Natural Areas, should be restricted to autochthonous species and ecotypes (Conrad and Tischew 2011) and thus, there is a need to select and test local ecotypes on the target sites (Ewing 1999). In relation to this, a number of programmes have been developed to identify and improve such material in Mediterranean climate areas (Abdelguerfi et al. 1988; Porqueddu and González 2006; Porqueddu et al. 2013).
Soil fertility is usually very low in mountain areas, especially in acidic soils, where nutrients are scarce and often are not available due to a low pH (Graham and Vance 2003). In Spain, one traditional low-cost practice to increase soil nutrient content and improve pasture quality in many grasslands and silvopastoral systems (dehesas) consists of the confinement of livestock in night pens (in Spanish, “redileo”). This practice consists of fencing livestock, mainly sheep, for three consecutive nights. Usually, there is about 1.5-2 m2 per sheep and assuming that two-thirds of daily dejections occur at night, a total of 20-30 Tn of dung per hectare plus urine (rich in potassium) are added to the soil over three nights (San Miguel 2001). These fences are moved every 3 days to enclose an adjacent area.
Other strategies to increase nutrient availability are linked to cooperative microbial activities occurring in the rhizosphere that can be exploited as a low-input biotechnology to help the productivity of both agricultural and natural ecosystems (Barea et al. 2005), and these include the activities of nitrogen-fixing bacteria and mycorrhizae. Firstly, symbiotic nodulating bacteria, collectively termed rhizobia, in legume roots ensure nitrogen availability for the plant due to their capability to reduce atmospheric nitrogen to ammonia in symbiotic root nodules (Leigh 2002). Furthermore, when using local ecotypes, local endosymbionts are usually present in the soil, but if necessary, seeds should be inoculated before sowing to ensure their persistence (Villadas et al. 2016). Secondly, vesicular–arbuscular mycorrhizae (VAM) are known to be very efficient in improving growth and nitrogen content in legumes, due to their ability to improve phosphorus intake by plants, and increase the supply of other immobilized nutrients or those present in low concentrations, such as ammonium, zinc and copper (Barea et al. 1987, 2005).
In September 2005, an accidental fire burned 3417 ha in Lanjarón, Sierra Nevada Natural and National Park (southern Spain), destroying natural vegetation, mainly scrub, woody scrub and repopulated pine woodland. A restoration plan was designed, and this included a specific rehabilitation programme for pastures in a fuel-break area, which consisted of identification and field harvesting of wild forage legumes, experimental sowings of target species, and assessment of fertilization techniques for the most successful species. In this context, this study sought to test the possibilities of pasture improvement at two silvopastoral sites (a fuel-break area and an open pine plantation) in Sierra Nevada Natural Park (Granada, southern Spain), through the evaluation of forage and seed yield of a total of 16 wild legumes species under different soil conditions and using various techniques for improving nutrient availability (rhizobial inoculation, mycorrhizal inoculation, sheep penning, and sheep penning with mycorrhizal inoculation). The following questions were posed: (1) which species perform best in terms of forage and seed yield? and (2) which fertilization techniques result in the highest forage and seed yields?
Materials and methods
The trials were carried out at two silvopastoral sites in Sierra Nevada Natural Park: (1) Soportújar (Vivero de la Sombra), an abandoned forestry nursery within a pine plantation which had been used as a sheepfold in recent years, and (2) Lanjarón (Cortijo Quemado), a fuel-break area in a pine plantation (Pinus pinaster) (Fig. 1). Table 1 summarizes the main characteristics of each site at the beginning of the experiment, and Table 2 presents the monthly rainfall for each site during the experiments together with mean historical values. These two sites were selected at the request of the Natural Park managers, since this park covers a large area (86,432 ha) and soil fertility and humidity conditions are diverse. Notably, Lanjarón is a good representative of dry sites (facing S/SE) with low soil fertility, while Soportújar is a good representative of wetter sites with fertile soils (facing W/SW). It was hoped that conducting the experiments at these sites would provide specific information about the suitability of species at each type of site and this might be useful for managers, shepherds, farmers, and other stakeholders willing to collaborate with pasture management and restoration in the Natural Park.

Study sites: Soportújar (left) and Lanjarón (right)
Seeds were collected in the field at different sites in Sierra Nevada Natural Park during late spring and early summer 2013. The experiments were carried out over two growing seasons: 2013/14 and 2014/2015. In 2013/14, the aim was to determine which species performed best under rainfed conditions at both experimental sites. Most seeds were sown on both sites, the exceptions being when there were not enough seeds, in which case they only were sown on the site where they were expected to perform the best. In 2014/2015, various techniques were tested in order to increase the availability of nutrients for the plants. For this year, we generally used only the species within each genus that were best adapted, based on the previous results. Nevertheless, due to constraints on seed availability, some species that performed better than others were not sown in the second year. Table 3 lists the species sown at each site in October 2013 and October 2014 and the corresponding seed density.
In 2013/14, for both sites, the experimental layout was a randomized block design with four replicates per species. Each replicate consisted of a 2 × 1.5 m plot. Prior to the establishment of the plots, the entire experimental area (at both sites) was ploughed to create a suitable seedbed. After that, each plot was fertilized with a pelletized organic amendment composed of a mixture of turf and sheep manure (81.7% organic matter, 2.6% total organic nitrogen, 2% of P2O5, 3.9% K2O). Seeds were sown by hand in furrows at 25-cm spacing, and lightly covered with soil.
In 2014/15, in Soportújar, a one-factor random block design was used with two treatments: mycorrhizae inoculation (M) and control (C) (without inoculation). Mycorrhizae inoculation consisted of applying mycorrhizae to soil, by watering with a solution containing a commercial product called Glomigel® (Mycovitro S.L.), once seedlings were established in the plots.
In Lanjarón, the experimental layout was a randomized block design with four blocks and with four treatments per block and species: rhizobium, sheep-penning (SP), and sheep-penning + mycorrhizae (SPM), and control. The rhizobium treatment consisted of pelletization of the seeds with native rhizobial species and symbiovars that were identified and isolated by Villadas et al. (2016) in the same area for these legume species. Sheep penning was carried out as follows: in summer 2014, 300 sheep were fenced into a 300 m2 pen for 16 h a day over 3 days. We estimated that around 15 Tn of fresh manure was deposited per ha. This management resulted in an increase in soil fertility to: 19.89 meq 100 g−1 cation exchange capacity, 4.55% organic matter, 0.24% total N, 16.25 p.p.m. P2O5, and 880 p.p.m. K2O (all being higher than pre-penning values given in Table 1). Sheep-penning + mycorrhizae consisted of applying mycorrhizae to soil, using the same procedure as in Soportújar, once seedlings were established in plots that had previously been treated by sheep-penning. The control consisted of uninoculated seeds or seedlings and untreated soil.
The experimental layout was different at each site due to the aforementioned differences in site conditions. In particular, two of the treatments aiming to increase soil fertility (especially nitrogen, phosphorus and potassium), namely, sheep penning and rhizobium, were not relevant at Soportújar given the high soil fertility at this site.
Forage yield was estimated in mid-May by hand-clipping plant forage within four randomly selected 25 × 25 cm quadrats in each plot. Samples were dried in a forced-air oven at 60 °C to constant weight (48 h) to determine dry weight. Averaged data were extrapolated to obtain yields for 1 hectare. Seed yield was estimated by the same procedure as for forage yield, except that samples were not oven dried. Seeds were manually extracted from each sample and weighed. In Lanjarón, which has poor soil quality, there was very poor fructification (almost none) following a spring (March and April) drought in 2014, and thus the corresponding data are not shown. In contrast, in Soportújar, with better soil conditions and higher precipitation, fruits developed and matured and, hence, it was possible to estimate seed yield.
Forage yield data from 2013/14 were analysed using the GLM procedure in IBM SPSS, Statistics for Windows, version 23. Levene and Shapiro–Wilk tests were applied to check homoscedasticity and normality, respectively, to ensure that assumptions of the model were met. No data transformation was needed. One-way analysis of variance (ANOVA) and least significant difference (LSD) post hoc tests were performed, for each site, to assess differences between species. Seed yield data from 2013/2014 (only for Soportújar) were analysed using the non-parametric Kruskal–Wallis test, as the data did not meet the assumptions of homoscedasticity and normality, and pairwise comparison post hoc tests.
For forage and seed yield data from 2014/15, the non-parametric Kruskal–Wallis test was used both for Soportújar and Lanjarón to test for differences among species. In Soportújar, within-species differences between treatments were assessed with Student’s t test (or the Mann–Whitney U test when homoscedasticity and normality assumptions were not met), while in Lanjarón differences between treatments were assessed by one-way ANOVA or with the Kruskal–Wallis test.
Results
In spring 2014, a seed germination experiment was carried out for all the species tested on the field trials to identify methods to reduce hardseededness, and thereby, promote germination (see Ramos et al. 2016 for further details). Based on the results of that experiment, various scarification treatments were applied to the seeds, for the 2014/15 sowings only, as indicated in Table 4.
Soportújar
Forage yield
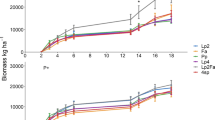
In Soportújar, for the first growing season, forage yields ranged between 265 and 7570 kg DM ha−1), with significant differences among species (H = 28.833; d.f. = 10; p = 0.001; (Fig. 2). The highest forage yields were obtained for V. monantha (7570 kg DM ha−1) and the lowest for L. sphaericus, T. glomeratum and T. cherleri (range: 265–375 kg DM ha−1) (Fig. 2). For the second growing season, significant differences were again found among species (pooled across treatments) (H = 48.338, d.f. = 7, p < 0.0001), with V. amphicarpa, V. monantha and V. disperma producing the highest yields (range: 7674–8969 kg DM ha−1), while L. nigricans, M. truncatula and V. lutea had the lowest yields (range: 633–1010 kg DM ha−1) (Fig. 2).

Forage yields (kg DM ha−1 year−1) in Soportújar for the growing seasons 2013/2014 and 2014/2015 (pooled across treatments). VM: Vicia monantha, VD: Vicia disperma, LCLY: Lathyrus clymenum, M: Medicago spp., LT: Lathyrus tingitanus, VL: Vicia lutea, MO: Medicago orbicularis, LN: Lens nigricans, TC: Trifolium cherleri, TG: Trifolium glomeratum, LS: Lathyrus sphaericus, VA: Vicia amphicarpa, LC: Lathyrus cicera, MT: Medicago truncatula. Different numbers over the horizontal brackets indicate significant differences between species. Purple bars indicate species that were sown in both growing seasons
When comparing the different treatments (control and mycorrhizal) within each species, we found significant positive responses to mycorrhizae application for L. nigricans, V. disperma and V. lutea, while this treatment had a detrimental effect on L. cicera (Table 5).
Seed yield
Seed yields were low for both growing seasons (Fig. 3). For 2013/14, seed yield was generally consistent with forage yield in Soportújar, the most productive species in terms of forage yielding the most seeds (V. monantha, 1234 kg ha−1), while the least productive (T. cherleri, T. glomeratum and L. sphaericus) gave the poorest seed yields (range: 21.6–66.7 kg ha−1) (H = 17.008; d.f. = 9; p = 0.049; Fig. 3).

Seed yield (kg ha−1 year−1) in Soportújar for the growing seasons 2013/2014 (a) and 2014/2015 (b) (pooled across treatments). VM: Vicia monantha, VL: Vicia lutea, LCLY: Lathyrus clymenum, VD: Vicia disperma, LT: Lathyrus tingitanus, MO: Medicago orbicularis, LN: Lens nigricans, LS: Lathyrus sphaericus, TG: Trifolium glomeratum, TC: Trifolium cherleri, VA: Vicia amphicarpa, LC: Lathyrus cicera, MT: Medicago truncatula. Different numbers over the horizontal brackets indicate significant differences between species. Purple bars indicate species that were sown in both growing seasons. (Color figure online)
For 2014/15, significant differences were also found among species (H = 40.044, d.f. = 7, p < 0.0001; Fig. 3). The highest seed yields were obtained for V. disperma, V. amphicarpa, and L. cicera (range: 684–817 kg ha−1) and the lowest for V. monantha, M. truncatula and V. lutea (range: 5.9–94.5 kg ha−1).
Comparisons between treatments in 2014/2015 showed that differences were only significant for V. disperma, yields being higher with mycorrhizae than the control treatment (Table 6).
Lanjarón
Forage yield
For 2013/14, forage yields in Lanjarón were very low (range: 45.9–487 kg DM ha−1), with significant differences among species (F = 3.158, d.f. = 12, p = 0.004; LSD test, p < 0.05). The most productive species were V. monantha, L. cicera, and V. disperma (more than 400 kg DM ha−1), while the least productive was M. polymorpha (45.9 kg DM ha-1) (Fig. 4).

Forage yield (kg DM ha−1 year−1) in Lanjarón for the growing season 2013/2014 and 2014/2015 (pooled across treatments). VM: Vicia monantha, LC: Lathyrus cicera, VD: Vicia disperma, MR: Medicago rigidula, VA: Vicia amphicarpa, LCLY: Lathyrus clymenum, LS: Lathyrus sphaericus, LT: Lathyrus tingitanus, VL: Vicia lutea, M: Medicago spp., VS: Vicia sativa, TC: Trifolium cherleri, MP: Medicago polymorpha. Different numbers over the horizontal brackets indicate significant differences between species. Purple bars indicate species that were sown in both growing seasons. (Color figure online)
For 2014/15, forage yields were notably higher (around three-fold higher for most of the species) than in the first growing season (range: 189–1415 kg DM ha−1). The most productive species were V. monantha and V. disperma (1415 and 1309 kg DM ha-1), while the least productive was T. cherleri (189 kg DM ha−1) (Fig. 4) (H = 61.621, d.f. = 6, p < 0.0001; p < 0.05).
Comparing treatments (growing season 2014/15), most of the species had higher yields with SP or SPM than under the control conditions, although differences were only significant for V. disperma, V. monantha and L. sphaericus (Table 7). For V. disperma and V. monantha, both SP and SPM were associated with the highest values for forage yield, while for L. sphaericus SPM seemed to boost forage yield. Although the yields for the other species did not differ significantly between treatments, high values under SP and SPM were found for M. rigidula and L. cicera, the latter also showing better performance under rhizobium (Table 7).
Seed yield
In Lanjarón, plants rarely fructified in 2013/14, and therefore, seed yield could not be quantified (see Materials and Methods). For 2014/15, seed yields were quantifiable but still very low, and there were significant differences between species (H = 19.077, d.f. = 6, p = 0.004; Fig. 5). The highest yields were obtained for L. cicera (60 kg ha−1) and V. disperma (56 kg ha−1) and the lowest (less than 3 kg ha−1) for L. sphaericus, T. cherleri and M. rigidula. Overall, seed yield results were consistent with forage yield.

Seed yields (kg ha−1 year−1) in Lanjarón for the growing season 2014/15 for seven species (pooled across treatments). LC: Lathyrus cicera, VD: Vicia disperma, VM: Vicia monantha, VA: Vicia amphicarpa, LS: Lathyrus sphaericus, TC: Trifolium cherleri, MR: Medicago rigidula. Different numbers over the horizontal brackets indicate significant differences between species
No differences between the treatments reached significance for any of the species (Table 8). For some species (L. sphaericus, M. rigidula and T. cherleri), however, seed yield was null for all (or almost all) the plots under control and rhizobium treatments, while SP or SPM were associated with a documentable seed yield.
Discussion
Our results have shown that forage and seed yield are low overall at these silvopastoral sites, especially in Lanjarón, probably attributable to low rainfall (especially in the spring) and/or to high hardseededness for most of the species (Table 4). Additionally, the proliferation of unsown species could have negatively affected the production due to competition with the sown species (Bàrberi 2002).
Among the 16 species tested, the most promising for pasture improvement and restoration within the two silvopastoral sites, in that they provided the highest forage and seed yields, were V. monantha, V. disperma and L. cicera at both sites, together with V. amphicarpa and L. clymenum at Soportújar, and M. rigidula at Lanjarón. Previous research of Robles et al. (2015) in Sierra Nevada Natural Park showed the high nutritional value of V. amphicarpa, V. disperma, and L. clymenum with high protein content and high organic matter digestibility, although they obtained low yields (412–200 kg DM ha−1). L. cicera has also been traditionally cropped in Mediterranean areas, and it has a high nutritional value, although it also contains toxic compounds, especially in the seed (White et al. 2002), and hence, grazing management is important to avoid poisoning.
The different pedoclimatic characteristics (Tables 1 and 2) of the study sites seemed to affect the production of the wild legumes. The higher forage and seed yields in Soportújar probably being attributable to higher rainfall and higher soil fertility (the experimental area having been used as a sheepfold for years, Table 1). Indeed, rainfall and certain soil parameters have been shown to be the most important factors in determining forage and seed yields in legumes (Leport et al. 1998; Siddique et al. 1999). Regarding the growing seasons, we must consider two different factors affecting forage and seed yield: (1) rainfall, not only the amount but also the distribution (Vázquez de Aldana and García-Criado 2008), and (2) scarification, seeds used in the second growing season having previously been scarified (Table 4). In Soportújar, the second growing season had an extremely dry spring (March and April) which resulted in lower yields for almost all the species (comparing results under the control treatment in the first and second growing seasons; see Fig. 2 and Table 4). In Lanjarón, the second growing season also had a drier spring than the previous year, although it was wetter than in Soportújar, but in this case, scarification was probably responsible for better crop performance, and in turn somewhat higher yields.
Compared to grain legumes in other Mediterranean areas (Leport et al. 1998; Siddique et al. 1999), seed yields were very low (most of the species producing less than 500 kg ha−1). A heat wave in mid-May (more severe during the first growing season) could have dramatically reduced fruit-setting in most of the species. Siddique et al. (1999) indicates that seed yield is positively correlated with dry matter production. In our study, this is true for most of the species but not all. For example, L. cicera had high seed yields at both sites in spite of having moderate or low forage yields. This could be related to the small size of the plant together with high seed weight (6.2 g 100 seeds−1). On the contrary, V. monantha (Soportújar) and M. rigidula (Lanjarón) had lower seed yields than expected; the former was severely attacked by aphids, while the latter seemed to be more sensitive to the heat wave and had a low rate of fruit setting. Indeed, water stress has been identified, together with defoliation, as a key factor underlying low seed production (Ewing 1999).
In our experiments, we found that forage yield for most of the species showed a positive response to organic fertilization, i.e., SP and SPM; but seed yield was not responsive to these treatments. Fertilization has been successfully applied in Mediterranean silvopastoral systems and positive responses have been found when fertilizing soils with less than 10 mg kg−1 of extractable phosphorus (as determined by Olsen’s method) (Osman et al. 1991), and extractable phosphorus levels are in this low range at Lanjarón. Research in Syria (Ewing 1999) showed that after applying phosphate fertilizer to degraded grasslands, biomass, legume seed pool and soil organic matter content were dramatically higher even 5 years after treatment. Similarly, positive effects of organic fertilizer (mature sheep manure) were observed by Robles et al. (2015) at the same Lanjarón site with wild legume species.
Mycorrhizal inoculation increased growth in some of the species (Soportújar: L. nigricans, V. disperma, V. lutea, and Lanjarón: L. sphaericus), probably by increasing the availability of phosphorus, but also by increasing the efficiency of the legume-Rhizobium symbiosis (Saia et al. 2014). In fact, many forage legumes have been shown to increase their growth as a response to mycorrhizal infection (see examples in Graham and Vance 2003 and Saia et al. 2014); however, different legume species have different degrees of dependence on mycorrhizae to grow. Regarding seed yield, only V. disperma in Soportújar responded to mycorrhizal inoculation. On the other hand, there was no significant effect associated with rhizobium inoculation for any of the species studied, probably because this type of bacteria are already present in the soil, and hence inoculation had no additional benefit (Villadas et al. 2016). Nevertheless, this technique could be of great interest for restoration after a fire event, as bacterial communities are negatively affected by fire as demonstrated by Cobo-Díaz et al. (2015), and consequently, rhizobium inoculation might be beneficial.
Conclusions
We found that pastures within fuel-break areas and open pine plantations in these semiarid Mediterranean conditions can be improved. Although low yields may be obtained in dry years, especially when soil fertility is low.
On the basis of our data, for these silvopastoral systems, we recommend V. monantha, V. disperma and L. cicera followed by V. amphicarpa and L. clymenum (at wetter more fertile sites) and M. rigidula (at drier sites), as they seem to be the best adapted to the pedoclimatic conditions of Sierra Nevada Natural Park. Increasing nutrients in the soil (through sheep penning) and, possibly, for some species, promoting nutrient assimilation (by mycorrhizal inoculation) may be effective strategies for increasing pasture biomass. Seed yield also seemed to be strongly affected by spring drought, and hence, when aiming to obtain seeds of these wild legumes for restoration programmes, deficit irrigation might be considered.
Further research is required to increase seed germination and, consequently, seedling establishment. Additionally, the improvement and development of specific cultivars and of appropriate mixtures of species for combined use in order to increase chances of plant establishment and restoration success is of great interest for these Mediterranean silvopastoral systems.
References
Abdelguerfi AJ, Chapot Y, Conesa AP (1988) Contribution ál’etude de la répartition des espécees spontanées de luzerne annuelles en Algérie, en relation avec certains facteurs du milieu. Fourrages 113:89–106
Arianoutsou M, Thanos CA (1996) Legumes in the fire-prone Mediterranean regions: an example from Greece. Int J Wildland Fire 6(2):77–82
Bàrberi P (2002) Weed management in organic agriculture: are we addressing the right issues? Weed Res 42(3):177–193
Barea JM, Azcón-Aguilar C, Azcón R (1987) Vesicular-arbuscular mycorrhiza improve both symbiotic N2 fixation and N uptake from soil as assessed with a 15 N technique under field conditions. New Phytol 106:717–725
Barea JM, Pozo MJ, Azcon R, Azcon-Aguilar C (2005) Microbial co-operation in the rhizosphere. J Exp Bot 56:1761–1778
Board MA (2005) Millennium ecosystem assessment. New Island, Washington, DC
Cobo-Díaz JF, Fernández-González AJ, Villadas PJ, Robles AB, Toro N, Fernández-López M (2015) Metagenomic assessment of the potential microbial nitrogen pathways in the rhizosphere of a Mediterranean forest after a wildfire. Microb Ecol 69(4):895–904
Conrad MK, Tischew S (2011) Grassland restoration in practice: do we achieve the targets? A case study from Saxony-Anhalt/Germany. Ecol Eng 37:1149–1157. https://doi.org/10.1016/j.ecoleng.2011.02.010
Ewing MA (1999) Annual pasture legumes: a vital component stabilizing and rehabilitating low-rainfall Mediterranean ecosystems. Arid Soil Res Rehabil 13:327–342
Graham PH, Vance CP (2003) Legumes: importance and constraints to greater use. Plant Physiol 131(3):872–877
Leigh GJ (2002) Nitrogen fixation at the millennium. Elsevier Science, London
Leport L, Turner NC, French RJ, Tennant D, Thomson BD, Siddique KHM (1998) Water relations, gas exchange and growth of cool-season grain legumes in a Mediterranean-type environment. Eur J Agron 9(4):295–303
Osman AE, Cocks PS, Russi L, Pagnotta MA (1991) Response of Mediterranean grassland to phosphate and stocking rates: biomass production and botanical composition. J Agric Sci 116:37–46
Pausas JG, Llovet J, Rodrigo A, Vallejo R (2008) Are wildfires a disaster in the Mediterranean basin?—a review. Int J Wildland Fire 17(6):713–723
Porqueddu C, González F (2006) Role and potential of annual pasture legumes in Mediterranean farming systems. Pastos 36(2):125–142
Porqueddu C, Re GA, Sanna F, Piluzza G, Sulas L, Franca A, Bullitta S (2013) Exploitation of annual and perennial herbaceous species for the rehabilitation of a sand quarry in a Mediterranean environment. Land Degrad Dev 27:346–356. https://doi.org/10.1002/ldr.2235
Ramos ME, Robles AB, Tognetti MJ, González-Rebollar JL (2016) Métodos para la reducción de la dureza seminal en leguminosas silvestres del espacio natural Sierra Nevada. In: Báez MD et al (eds) Innovación Sostenible: Hacia una agricultura de respuesta al cambio climático. SEEP, Lugo (Spain), pp 22–26
Robles AB, Ramos ME, Cabeza FM, Delgado F, González Rebollar JL (2015) Leguminosas herbáceas en la restauración forestal de zonas incendiadas del macizo Sierra Nevada: producción y calidad. In: Cifre Lompart J et al (eds) Pastos y forrajes en el siglo XXI. SEEP, Mallorca (Spain), pp 129–136
Ruiz-Mirazo J, Robles AB (2012) Impact of targeted sheep grazing on herbage and holm oak saplings in a silvopastoral wildfire prevention system in south-eastern Spain. Agroforest Syst 86:477–491
Saia S, Amato G, Frenda AS, Giambalvo D, Ruisi P (2014) Influence of arbuscular mycorrhizae on biomass production and nitrogen fixation of berseem. PlosOne 9(3):e9073
San Miguel A (2001) Pastos naturales españoles. Caracterización, aprovechamiento y posibilidades de mejora. Coedición Fundación Conde del Valle de Salazar- Mundi-Prensa, Madrid
Siddique KHM, Loss SP, Regan KL, Jettner RL (1999) Adaptation and seed yield of cool season grain legumes in Mediterranean environments of south-western Australia. Aust J Agric Res 50(3):375–388
Thavaud P (2009) Guide pratique pour l’entretien des coupures de combustible par le pastoralisme. La Cardère-l´Éphémère, Lauden
Vázquez-de-Aldana BR, García-Criado B (2008) Interannual variations of above-ground biomass and nutritional quality of Mediterranean grasslands in Western Spain over a 20-year period. Aus J Agr Res 59(8):769–779
Villadas PJ, Lasa AV, Martínez Hidalgo P, Flores-Félix JD, Martínez-Molina E, Toro N, Velázquez E, Fernández-López M (2016) Analysis of rhizobial endosymbionts of Vicia, Lathyrus and Trifolium species used to maintain mountain firewalls in Sierra Nevada National Park (South Spain). Syst Appl Microbiol 40:92–101
White CL, Handbury CD, Young P, Philips N, Wiese SC, Milton JB, Davidson RH, Siddique KHM, Harris D (2002) The nutritional value of Lathyrus cicera and Lupinus angustifolius grain for sheep. Anim Feed Sci Technol 99:45–64
Acknowledgements
This work has been funded by the National Parks Autonomous Agency (OAPN, Spanish Ministry of Agriculture, Food and Environment) through the project “Investigaciones sobre la flora forrajera natural en mejoras de pastos, restauración forestal y silvicultura preventiva con ganado: una experiencia piloto en Sierra Nevada” (Ref. 748). We would like to thank Baltasar del Pozo for managing the sheep penning, Mycovitro who donated the Glomigel, and Manuel Fernández-López and Pablo Villadas who provided the rhizobium inocula for this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ramos-Font, M.E., Tognetti-Barbieri, M.J., González-Rebollar, J.L. et al. Potential of wild annual legumes for mountain pasture restoration at two silvopastoral sites in southern Spain: promising species and soil-improvement techniques. Agroforest Syst 95, 7–19 (2021). https://doi.org/10.1007/s10457-018-0340-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-018-0340-5