
Abstract
A field study was conducted at Hetian oasis, southern Xinjiang, northwestern China, to investigate root distribution and morphology affected by interspecific interactions between jujube trees (Ziziphus jujuba Mill.) and cotton plants (Gossypium hirsutum L.). The treatments comprised (1) sole cotton, (2) 5-year-old sole jujube trees, (3) 7-year-old sole jujube trees, (4) 9-year-old sole jujube trees, (5) intercropping of cotton/5- year-old jujube trees, (6) intercropping of cotton/7-year-old jujube trees, and (7) intercropping of cotton/9-year-old jujube trees. Roots in each plot were sampled vertically by auger to a depth of 100 cm at 20-cm intervals and horizontally up to 150 cm away from the base of the trees at 30-cm intervals. The results showed that all jujube/cotton intercropping systems exhibited advantages of intercropping, with a land equivalent ratio > 1. The roots of both intercropped cotton and jujube had lower root length density (RLD) and root diameter (RD) at all soil depths compared with those of sole-cropped cotton and jujube trees. Interspecific competition with jujube trees altered the root development and morphology of the cotton plants. The roots of the trees spread underneath the crop and the roots of intercropped cotton tended to have a shallower distribution in the soil profile. A greater distance from the trees resulted in less influence on the RLD and RD of intercropped cotton. Older jujube trees had more developed roots and consequently higher RLD and RD. These findings may partly explain the interspecific competition effects in jujube tree/cotton agroforestry systems. However, the mechanisms underlying the root interactions between cotton and jujube require further investigation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Maximizing the use of available resources (positive interactions), while minimizing resource competition (negative interactions) between trees and crops, is central to improving the yield and overall productivity in agroforestry systems (Zamora et al. 2007). Agroforestry provides the opportunity to maintain both food and fibre production as well as increase biodiversity (Jose et al. 2004). It offers a number of services and environmental benefits and plays a key role in a multifunctional and sustainable agriculture landscape (Jose 2009). However, when two plant species grow together, interspecific interactions between them inevitably occur, and this is especially true in agroforestry systems (Zhang et al. 2013, 2015, 2016). Plants compete with each other for the available resources, with each plant trying to fulfill its needs (Cao et al. 2012). In tree-based alley cropping systems, resource competition between trees and crops certainly will occur, at least for the following two reasons. First, there is overlap in the growth period of the tree and crop (Ding and Su 2010). This means that there will be competition for light. Second, there is overlap in the belowground root distribution (Ding and Su 2010), and this involves unavoidable root interaction. The coarse root distribution of most perennial woody trees is largely within the top 40 cm of the soil profile (Zhang et al. 2013). Root interactions play an important role in tree-based intercropping systems. In tree-crop experimental systems, i.e., black walnut (Juglans nigra L.)-maize (Zea mays L.) and red oak (Quercus rubra L.)-maize, Jose et al. (2000a, b) found that competition for belowground resources (water or nitrogen) between the trees and crops seemed to be critical in defining the productivity and sustainability of the systems. In the same research, Gillespie et al. (2000) reported that crop yield reductions of up to 40% occurred due to severe root competition when the trees were 11 years old, while shading did not have a major influence on crop yield. In a windbreak/crop alley intercropping system in Nebraska, USA, Hou et al. (2003) found that belowground soil water competition between the crop (soybean [Glycine max (L) Merr.]) and windbreak (One row consisted of green ash (Fraxinus pennsylvanica L.) and Austrian pine (Pinus nigra Arnold). The other row consisted of green ash and eastern redcedar (Juniperus virginiana L.).) probably played a leading role in yield suppression.
Plant root systems have the inherent capability to adjust to prevailing environmental conditions through their physiological and morphological plasticity (Zamora et al. 2007). Many previous studies have shown that interspecific competition in tree-based alley intercropping systems may reduce root length density (RLD) and aboveground biomass. For instance, in a hybrid walnut (Juglans regia × nigra L.)-durum wheat (Triticum turgidum L. subsp. durum) alley cropping system, Cardinael et al. (2015) found that the RLD of intercropped tree roots was much lower than in the tree-only plots, but the roots occupied a greater volume of soil. Livesley et al. (2000) found that the root length of both trees (grevillea (Grevillea robusta) and senna (Senna spectabilis)) and an arable crop (maize) decreased to a certain degree. Similarly, Zhang et al. (2013, 2015, 2016) and Wang et al. (2014) found that jujube trees grown with wheat (Triticum aestivum L.) and Persian walnut trees (Juglans regia L.) grown with wheat in an arid climate had a lower RLD and RD than trees grown in a monospecific stand due to competition from the crops. In studies of jujube tree/winter wheat–summer maize intercropping, Yang et al. (2016) reported that the mean grain yield of the intercropped winter wheat and summer maize was reduced by 35.6 and 35.2%, respectively, compared with their corresponding sole cropping. Rao et al. (1991) concluded that roots in the upper soil profile caused intense belowground competition for resources and resulted in decreased crop productivity. Jose et al. (2004, 2006) reported that the spatial distribution of the roots of component species plays an important role in underground resource competition. Thus, knowledge of the spatial distribution of tree/crop root systems is necessary to determine the degree of competitive resource sharing among system components (Zhang et al. 2015). Moreover, Jose et al. (2000b) found that maize yields were reduced by 35 and 33% when intercropped with black walnut and red oak (Quercus spp.), respectively. In a Leucaena/maize intercropping system, Chamshama et al. (1998) found that the mean height and biomass of maize were reduced by 43 and 75%, respectively. Productivity is central to alley cropping systems, and many researchers hold the view that, in most situations, aboveground biomass is affected more by belowground root interactions than by aboveground shading (Singh et al. 1989; Ong et al. 1991; Schroth 1999; Zhang et al. 2013, 2015, 2016; Wang et al. 2014). The aforementioned studies have shown that the productivity of many agroforestry systems is limited by interspecific underground competition for resources, resulting in the current research on root interactions being a major component of alley cropping studies.
Alley cropping is a major form of agroforestry in China and involves cultivation of woody perennials, usually for fruit or nut production, in rows, while crops are cultivated between the rows of trees (Zhang et al. 2013, 2015). Agroforestry has been practiced for several thousands of years in China (Huang et al. 1997). To achieve better yield and benefit, tree-based intercropping systems are largely favored by the local farmers (Zhang et al. 2013, 2015, 2016). Jujube tree/cotton agroforestry systems play an important role in agricultural production in Hetian old oasis, Southern Xinjiang Uygur Autonomous Region, northwestern China. Xinjiang is a semi-arid region where rainfall is limited, but light is abundant and irrigation water is available, which provides excellent growing conditions for high quality fruit and fibre crop production, e.g., jujube and cotton. Growing cotton as a companion crop between jujube tree rows is a common and profitable practice, especially during the young stage of the jujube trees (younger than 10 years), and was applied to an area of approximately 32,000 ha in Hetian old oasis (Xingjiang) in 2011(Zhang et al. 2014). In this agroforestry system, the jujube trees are grown simultaneously with cotton for 180–200 days.
In recent decades, agroforestry has been rapidly developed in order to increase overall production by harnessing the potential of the various resources involved. Thus, a reasonable assessment of the current state and the potential of agroforestry is necessary for better management in the future. Although jujube tree/cotton agroforestry is one of the most popular tree-based alley cropping systems in Hetian old oasis (Zhang et al. 2014), root distribution information on these systems has been limited to descriptive and qualitative accounts. It is therefore necessary to carry out a more comprehensive study of the influence of trees on the crops in this region. In the present study, we experimentally compared three ages of jujube trees grown with a cotton crop to examine the competitive interactions between the two species and the likely response mechanisms in order to guide policy decisions in tree-crop system management. Jujube trees and cotton were selected for the study because of their importance as the main economic crops in southern Xinjiang.
Considering the growing interest in jujube trees and cotton for alley cropping systems in northwestern China and the limited information on their growth, biomass, yields and root distribution, specifically in response to belowground competition, our study was conducted to test the following key questions:
-
1.
How does the interspecific underground competition in the tree-cotton associations alter the yield and biomass?
-
2.
How will root length density and root diameter respond to belowground resource competition?
-
3.
Will root spatial distribution change in response to competition?
We hypothesized that (1) the yield and biomass will decrease with interspecific underground competition in the tree-cotton associations; (2) the root length density and root diameter will be adversely affected by belowground competition; and (3) root spatial distribution will be altered in response to belowground competition.
Materials and methods
Experimental site
The study was conducted in 2014 at the Hetian Agricultural Scientific Research Institute (73°37′N, 34°20′E), Agro-Tech Extension and Service Center of Hetian Prefecture, Xinjiang Uygur Autonomous Region, China. The annual mean temperature of this region is 13.7 °C, and cumulative temperatures above 0 and 10 °C are 4646 and 4064 °C, respectively. The annual precipitation is 37.1 mm, potential evaporation is 2636 mm, and the region has a typical arid climate. The soil at the site is classified as an Arenosol according to the classification system of the Food and Agriculture Organization (FAO). Arenosols are sandy textured soils that lack any significant soil profile development. They exhibit only a partially formed surface horizon (uppermost layer) that is low in humus, and they are bereft of subsurface clay accumulation. The study is part of a larger experiment that includes a randomized complete block design with 27 plots allocated to jujube tree-wheat alley cropping (Zhang et al. 2013, 2016; Wang, et al. 2014) and 27 plots allocated to jujube-cotton alley cropping. The characteristics of the physical and chemical properties of the soil have been described in previous papers (Zhang et al. 2013, 2015, 2016). The intercropping was designed as a replacement series.
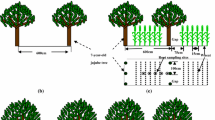
The following seven treatments were compared in this study: sole-cropped jujube trees (5, 7 or 9 years old with a north–south direction), intercropped jujube/cotton (the jujube trees were 5, 7 or 9 years old with a north–south direction), and sole-cropped cotton. The planting patterns are shown in Fig. 1. In plots with 5-year-old jujube, the row spacing was 200 cm, and there was 100-cm spacing between trees within a row. When cotton was planted between the jujube rows, the cotton strip was 120 cm wide (3 rows), and there was a 60-cm-wide bare strip between the cotton and the jujube trees. The jujube trees occupied 40% of the intercropped area, and cotton occupied 60%. In plots with 7-year-old jujube, the row spacing was 600 cm, and there was 100-cm spacing between trees within a row. When cotton was planted between the jujube rows, the cotton strip was 480 cm wide (9 rows), and there was a 60-cm-wide bare strip between the cotton and jujube trees. The jujube trees occupied 20% of the intercropped area, and cotton occupied 80%. In plots with 9-year-old jujube, the row spacing was 300 cm, and there was 200 cm between trees within a row. When cotton was planted between the jujube rows, the cotton strip was 180 cm wide (4 rows), and there was a 60-cm-wide bare strip between the cotton and the jujube trees. The jujube trees occupied 40% of the intercropped area, and cotton occupied 60%. There was 60 cm between cotton rows. Each plot included two 10-m-long rows of jujube. Thus, the 5-year-old jujube plots were 2 m × 10 m, the 7-year-old jujube plots were 6 m × 10 m, and the 9-year-old jujube plots were 3 m × 10 m. Each treatment was replicated three times. Plastic film mulching was applied to all plots according to farmers’ practices due to the limitation of heat, especially in early spring.

Schematic diagram showing the planting patterns for (a) sole-cropped 5-year-old jujube trees, (b) sole-cropped 7-year-old jujube trees, (c) sole-cropped 9-year-old jujube trees, (d) 5-year-old jujube/cotton, (e) 7-year-old jujube/cotton and (f) 9-year-old jujube/cotton in intercropped plots. Distances between rows and gap sizes (cm) are indicated. There was a distance of 30 cm between adjacent root sampling sites
A basal fertilizer was applied at a rate of 300 kg ha−1 N and 150 kg ha−1 P in 2014. The same amount (300 g ha−1) of the growth regulator mepiquat chloride (MC) was applied to all plots according to farmers’ practice to control cotton growth. The plots were irrigated ten times at 15-day intervals. Each irrigation application consisted of 60 mm (600 m3 ha−1). A top-dressed fertilizer consisting of 30 kg ha−1 N and 15 kg ha−1 P was applied at the time of irrigation. These irrigation practices were recommended to farmers by local agronomists.
Collection of root samples
The vertical root distribution was determined using the method of Böhm (1979). Soil cores were collected using an auger (5.5 cm diam.) on September 20, 2014. Both the cotton and trees were mature. The soil cores were collected in 20-cm increments to a maximum depth of 100 cm. The cores were collected 30, 60, 90, 120, and 150 cm from the tree row in each plot so that spatial changes in the horizontal distribution of the roots could be determined (Fig. 1). The specific sampling and measuring methods for roots have been described in a previous paper (Zhang et al. 2013).
Collection of plant samples
The cotton cultivar was an early-mature variety called ‘XLZ46’ and was sown by hand on 22 April 2014. The harvest date of the two crops was October 15, 2014. Immediately before harvesting, the stunting and mortality rates of the cotton plants were determined along a 5-m-long section of each cotton strip. The plants were then cut at ground level. The samples were air dried, threshed by hand, and then weighed to determine the yield and total aboveground biomass (i.e., lint + straw).
The jujube sprouted at the beginning of May and flowered after 30 days. The trees were pruned annually. The jujube fruits in each plot were harvested on October 15, 2014. The fruit were air-dried and then weighed. A total of 100 fruits were randomly subsampled to measure the water content. The dry matter of the entire final jujube yield was calculated based on the total fresh weight and a water content of 38.1% (Zhang et al. 2014).
Calculation of the land equivalent ratio (LER)
The LER is the ratio of the area under a pure stand to the area under intercropping needed to produce an equal amount of yield at the same management level (Vandermeer 1989; Rao et al. 1990, 1991; Cao et al. 2012). The LER indicates the environmental resource use efficiency of intercropping compared with sole cropping (Mead and Willey 1980; Dhima et al. 2007). The LER is calculated using the following equation:
where Yj and Yc are the yields of sole-cropped jujube and sole-cropped cotton, respectively, and Yjc and Ycj are the yields of jujube and cotton in the intercropped systems, respectively. A ratio greater than 1.0 indicates that intercropping is advantageous whereas a ratio less than 1.0 indicates that intercropping is disadvantageous (Zhang et al. 2013, 2015, 2016; Wang et al. 2014).
Statistical analysis
Analysis of variance (ANOVA) was conducted using SPSS Version 19.0 for Windows. Means (n = 3) were compared using least significant difference (LSD) tests at the 5% level.
Results
Land equivalent ratio, yield and aboveground biomass
All land equivalent ratios (LERs) of jujube tree/cotton intercropping were greater than one regardless of whether the jujube trees were 5, 7 or 9 years old. The highest efficiency of land use in this experiment was obtained for cotton/7-year-old jujube tree intercropping, with an LER value of 1.36, cotton/5-year-old jujube intercropping had an LER value of 1.23, and cotton/9-year-old jujube tree intercropping had an LER value of 1.18 (Table 1).
Intercropping with 5-, 7-, and 9-year-old jujube trees significantly reduced the cotton yield on a net area basis by 11% (p = 0.033), 20% (p = 0.026), and 29% (p = 0.001), respectively (Table 1). The above-ground biomass of cotton was significantly reduced by 5-, 7-, and 9-year-old jujube trees by 13% (p = 0.040), 17% (p = 0.043), and 30% (p = 0.001), respectively. Intercropping significantly reduced the fruit yield of 5-, 7-, and 9-year-old jujube trees by 30% (p = 0.001), 29% (p = 0.012), and 25% (p = 0.012), respectively (Table 1).
Spatial distribution of cotton root length density
Intercropping reduced the RLD of cotton at all distances from the jujube trees (Fig. 2a–d). Averaged across all soil depths (0–100 cm), the RLD of cotton intercropped with 5-year-old jujube was 69, 74, 75, 79 and 81% that of sole-cropped cotton at respective distances of 30, 60, 90, 120 and 150 cm from the trees (Fig. 2a, b, p < 0.05). In other words, the RLDs of intercropped cotton were lower than those of the sole-cropped treatment (Fig. 2a, b). For instance, the 140 cm/125 cm3 contour of cotton intercropped with 5-year-old jujube trees was below the 40-cm depth in the sole-cropped treatment but above the 40-cm soil depth in the intercropped treatment (Fig. 2a, b). The same pattern was observed in cotton intercropped with 7- and 9-year-old jujube. In all treatments, the roots were concentrated in the upper 40 cm of the soil (Fig. 2b–d), and the RLD declined rapidly with increasing depth. The RLD of intercropped cotton (Fig. 2b–d) was lower in the two cotton border rows on both sides compared with the middle rows. The horizontal distribution of cotton RLD in plots with 9-year-old trees was different from that in plots with either 5- or 7-year-old trees (Fig. 2b–d). The RLD of intercropped cotton was reduced by trees at most of the soil depths. The largest reductions in the RLD of intercropped cotton were 40–80 cm from the tree row (Fig. 2b–d, p < 0.05). The roots of intercropped cotton tended to be distributed more shallowly than those of sole-cropped cotton.

Spatial distribution of root length density (RLD, cm 125 cm−3) of sole- and intercropped cotton: (a) cotton grown alone, (b) cotton intercropped with 5-year-old jujube trees, (c) cotton intercropped with 7-year-old jujube trees, and (d) cotton intercropped with 9-year-old jujube trees
Spatial distribution of root length density of jujube
The distribution of jujube RLD is presented as contour diagrams in Fig. 3a–f. The RLD of intercropped jujube trees was lower than that of sole-cropped jujube. Averaged across the 0–100 cm depth, the RLDs of the 5-, 7- and 9-year-old intercropped jujube trees were 14 (p < 0.05), 12 (p < 0.05), and 9% (p > 0.05) lower, respectively, than those of monocropped trees of the same age. For example, the 50 cm/125 cm3 contour of 5-year-old jujube trees was below the 40-cm depth in the sole-cropped treatment but above the 40-cm soil depth in the intercropped treatment (Fig. 3a, b). The same patterns were also observed for 7- and 9-year-old jujube trees. The 70 cm/125 cm3 RLD contour of 7-year-old trees was below the 40-cm depth in the sole-cropped treatment but above the 40-cm depth in the intercropped treatment (Fig. 3c, d). For 9-year-old trees, the 80-cm/125 cm3 RLD contour was below the 40-cm depth in the sole-cropped treatment whereas in the intercropped treatments, it was above the 40-cm depth (Fig. 3e, f). In all treatments, jujube roots were concentrated in the upper 60 cm of soil (Fig. 3a–f). By and large, there was little difference in the horizontal distribution of jujube roots between the intercropped treatments and the monocropped treatments (Fig. 3a–f). In the intercropped treatments, the jujube roots grew beneath the cotton rows, and the RLD beneath the cotton was lower than that in the corresponding position in the monocropped jujube treatments (Fig. 3b, d, f).

Spatial distribution of root length density (RLD, cm 125 cm−3) of sole and intercropped jujube: (a) jujube trees grown alone for 5 years, (b) 5-year-old jujube trees intercropped with cotton, (c) jujube trees grown alone for 7 years, (d) 7-year-old jujube trees intercropped with cotton, (e) jujube trees grown alone for 9 years, and (f) 9-year-old jujube trees intercropped with cotton
Spatial distribution of cotton root diameter
The spatial distribution of cotton RD in the sole- and intercropped treatments is shown in Fig. 4. Intercropped cotton had a lower RD than monocropped cotton at the corresponding sampling positions. Intercropping led to thinner cotton roots. Averaged across the 0–100-cm depth, the RD of sole-cropped cotton was 0.26 mm. In comparison, the RD of cotton intercropped with 5-, 7- and 9-year-old jujube trees was 0.23 (p > 0.05), 0.21 (p < 0.05) and 0.17 mm (p < 0.05), respectively. The RD of cotton was lower in the intercropped treatments than in the sole-cropped treatment at all depths, especially in the 0–60-cm depth (Fig. 4a–d). Specifically, compared with sole-cropped cotton, intercropping with 5-, 7- and 9-year-old jujube trees reduced the RD of cotton by 11.5 (p > 0.05), 19.2 (p < 0.05) and 42.3% (p < 0.05), respectively (Fig. 4a–d). This indicated that the 9-year-old trees reduced the RD of cotton more than the 5- and 7-year old trees. The average RDs were 0.35 mm in the top 60 cm of the soil profile of sole cotton, but the average RD values of cotton intercropped with 5-, 7- and 9-year-old jujube trees were 0.33 (p < 0.05), 0.30 (p < 0.05) and 0.25 mm (p < 0.05), respectively, in the same soil depth range. The RDs of intercropped cotton were lower than those of sole cotton at most of the soil depths (Fig. 4a–d).

Spatial distribution of root diameter (RD mm) of sole and intercropped cotton: (a) cotton grown alone, (b) cotton intercropped with 5-year-old jujube trees, (c) cotton intercropped with 7-year-old jujube trees, and (d) cotton intercropped with 9-year-old jujube trees
Spatial distribution of jujube root diameter
The distribution of the RD of the jujube trees is presented in Fig. 5. Intercropping led to thinner tree roots and reduced the RD of 5-year-old jujube trees. In the intercropped treatments, the average RD of 5-year-old jujube trees was much larger than that of cotton at all depths (Fig. 5a, b, p < 0.05). However, a general increasing trend in the RD was also observed in the 7- and 9-year-old intercropped jujube trees. Averaged across all depths (0–100 cm), the respective RDs of the 5-, 7- and 9-year-old jujube trees were 2.63, 3.31 and 4.69 mm in the sole-cropped treatment compared with 2.36, 2.60 and 3.40 mm in the intercropped treatment. Averaged across the 0–60-cm depth, the respective RDs of the 5-, 7- and 9-year-old jujube trees were 3.91, 4.49 and 5.39 mm in the sole-cropped treatments compared with 3.53, 3.85 and 4.71 mm in the intercropped treatments. Overall, the RD of the intercropped trees was lower than that of sole-cropped trees at most of the soil depths, especially the 0–60-cm soil depth (Fig. 5a–f).

Spatial distribution of root diameter (RD mm) of sole and intercropped jujube: (a) jujube trees grown alone for 5 years, (b) 5-year-old jujube trees intercropped with cotton, (c) jujube trees grown alone for 7 years, (d) 7-year-old jujube trees intercropped with cotton, (e) jujube trees grown alone for 9 years, and (f) 9-year-old jujube trees intercropped with cotton
There were also significant differences in the vertical distribution of RD in the 5-year-old jujube tree/cotton intercropping treatment. For example, the 1.8-mm contour of 5-year-old trees was below the 60-cm depth in the sole-cropped treatment but above the 60-cm depth in the intercropped treatment (Fig. 5a, b). However, vertical differences in RD between sole and intercropped 7- and 9-year-old trees were not apparent (Fig. 5c–f).
Discussion
Land equivalent ratio, yield and aboveground biomass
An LER of 1 indicates that no yield advantage is obtained, while a value of 1.1 indicates a 10% yield advantage. In the case of an LER of 1.1, 10% more land would be needed for the sole-cropped crop to provide the same yield as the agroforestry system (Nerlich et al. 2013). In our research, all LERs were greater than 1, indicating that intercropping of cotton/jujube trees, irrespective of age, resulted in significant yield advantages. Similar results were also observed in semiarid Machakos, Kenya, where the total LERs were 1.09 for leucaena (Leucaena leucocephala)/maize systems and 1.28 for senna (Senna siamea syn. Cassia siamea)/maize systems (Jama et al. 1995). In another parallel study, Droppelmann et al. (2000) reported that the LERs for Acacia saligna (Labill.) H. Wendl/sorghum (Sorghum bicolor (L.) Moench.) intercropping at a high planting density were clearly higher than 1. The results show that there was a significant decrease in intercropped cotton yield and aboveground biomass and fruit yield of the intercropped trees compared with the respective sole-cropped cotton and jujube trees. Schroth (1995) reported that trees can compete with crops through their root systems, and this may lead to decreased yield. In a black walnut/maize alley cropping system, black walnut and maize yields declined by 33 and 35%, respectively (Jose et al. 2000b). Our results are similar to previous reports on tree-crop associations (Zhang et al. 2013, 2015, 2016; Wang et al. 2014). The above results answer our first question and indicate that biomass and yield will decrease with belowground interspecific competition in tree/crop mixtures. The results also support previous reports from the same study site, i.e., that tree-crop associations have the potential benefits of higher productivity and sustainability compared with sole crops (Zhang et al. 2013, 2015, 2016; Duan et al. 2017). Intercropping systems may be particularly useful in southern Xinjiang where relatively little arable cropping is currently practiced (Wang et al. 2014).
Interspecific competition and root spatial distribution
The acquisition of belowground resources is based on the morphology and distribution of the plant root systems, which can exhibit variations resulting from adjustments to temporal environmental conditions (Zamora et al. 2007). In a recently published review, Kumar and Jose (2017) hold the view that occurrence of two or more tree species close to one another may favour diminished lateral spread and/or deeper root penetration of the woody components. The outcomes of the experiment also answer our second and third questions on root length density, root diameter and root distribution, which were affected by belowground competition, and there were significant differences in RLD and RD between the cotton and jujube trees. Belowground interspecific interactions in the jujube/cotton intercropping system appeared in response to competition. The RLDs and RDs of both intercropped species decreased compared to the monocrops. Intercropped cotton had a lower RLD and RD than monocropped cotton in the three intercropping systems, and the older the jujube trees, the greater the decrease in RLD and RD and the stronger the inhibition of cotton root growth (Figs. 2 and 4). Although there appeared to be mutual competition in the root growth of the two crops, the competitiveness of cotton was far below that of jujube. This means that plants with high fine root length densities are likely to be more competitive than those with lower root length densities (Zhang et al. 2013, 2015). Similar results were also obtained in jujube tree/wheat (Zhang et al. 2013, 2016; Wang et al. 2014) and walnut tree/wheat (Zhang et al. 2015) intercropping systems by our research group. In a pecan (Carya illinoensis K. Koch)–cotton (Gossypium hirsutum L.) alley cropping system in southern United States, Zamora et al. (2007) found that both the RLD and total root length of cotton varied in response to interspecific competition with pecan for above- and belowground resources. In the same experiment, competition for belowground resources (water) (Wanvestraut et al. 2004) and aboveground resources (light) (Zamora et al. 2006) significantly reduced the growth of cotton roots in the non-barrier treatment compared to the barrier and monoculture treatments. Wanvestraut et al. (2004) pointed out that this rooting pattern of pecan indicated intense competition for water with the cotton plants in the non-barrier treatment. Monoculture and barrier plants had higher total fine root length compared to the non-barrier plants. Zamora et al. (2007) discovered that belowground competition for resources between pecan and cotton in the non-barrier treatment resulted in respective reductions of 25 and 33% in total root length (359 cm) compared to that of the barrier (477 cm) and monoculture (539 cm) treatments. Allen et al. (2005) suggested that wetter soil in the barrier treatment provided a greater advantage for the barrier plants to grow rapidly, resulting in expansion and greater development of their root systems compared to non-barrier plants. Bolte and Villanueva (2006) found that the sum of fine roots in mixed stands of European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) was lower than in pure stands of these species. Several authors hold the same view, i.e., that the interactions between trees and crops may lead to increased capture of a limiting growth resource (Ong and Huxley 1996; Ashton 2000; García-Barrios and Ong 2004) and may lead to lower RLD and RD of crops. In addition, in our study, the roots of intercropped jujube trees spread into the root zone beneath the cotton (Figs. 2b–d, 3b, d, f, 4b–d, 5b, d, f) and tended to have a more shallow distribution in the soil profile compared to monocropped jujube trees. The reason was that the perennial plant roots occupied a greater soil volume. Similar results have also been observed in jujube tree/wheat (Zhang et al. 2013, 2016; Wang et al. 2014) and walnut tree/wheat (Zhang et al. 2015) intercropping systems. Furthermore, we found that the RLD and RD of intercropped cotton were reduced more in the two cotton border rows on both sides compared with the middle rows. When two types of plant are intercropped in agroforestry systems, the tree roots extend to considerably greater distances and depths than do the crop roots, and they exhibit a rapid decline in root mass, number and length with increasing soil depth (Stone and Kalisz 1991; Schroth 1999). Crop root development was adversely affected adjacent to hedge-rows, as belowground interspecific interactions occurred in response to strong competition (de Wit 1960). In another companion study on root morphological plasticity of jujube/wheat intercropping systems, Wang et al. (2014) found that roots of intercropped wheat and jujube exhibited plasticity in their horizontal distribution owing to the intercropping. The shorter the distance from the jujube trees, the smaller the RLD and RD of intercropped wheat. Jose et al. (2000a) also found that the fine root biomass of both black walnut and red oak decreased as distance and depth increased in non-barrier treatments compared to barrier treatments in a tree-crop interaction experiment, and the fine root biomass increased at a distance of 1.1 m within the 0–30-cm soil layer for black walnut in the barrier treatment. Other studies have also found that the root density of Douglas fir decreased more gradually with depth in mixtures than in pure stands in a Douglas fir mixed forestry system (Hendriks and Bianchi 1993). In a jujube tree/winter wheat–summer maize agroforestry system, Yang et al. (2016) found that root density was significantly higher (p < 0.05) 5.5 m from the tree rows than at 3.5 and 2.5 m, in the 0–40-cm layer for winter wheat and the 0–10-cm layer for summer maize. These findings are also in agreement with the research results of Zamora et al. (2007) in a pecan tree/cotton intercropping, where they observed a decreasing trend in the RLD of cotton grown at a further distance from pecan trees. We believe that annual and perennial plants have different ecological niches in the soil; however, we believe this is only one of the reasons for the root spatial distribution observed in our tree/crop alley cropping system. Of course, the mechanisms of belowground competition require further investigation in jujube/cotton intercropping systems.
Density effect
In addition to the tree age effect, belowground interspecific competition derived from plant density is another key factor. Atkinson et al. (1978) demonstrated that the spatial distribution of tree root activity is influenced by the spacing between trees and associated crops. Imo and Timmer (2000) suggested that interspecific interactions in agroforestry systems are not constant and may be affected by several factors such as planting densities. Cao et al. (2012) also concluded that resource complementarity, competitive ability and severity of competition may all be affected by the density of each component in the mixture. The author found that Ginkgos were stronger competitors than crop species at high tree densities. This kind of root competition may be an asymmetric interaction, especially when the intercropped trees are old and when the tree density is high (Yin and He 1997). In our study, although the cotton plant density in the three tree-crop systems was uniform, the fact that the three tree age-based alley cropping systems had different plant density should not be neglected. Therefore, from an experimental point of view, the same tree density may be more meaningful for further research on interspecific root competition in the future.
Conclusions
In all three cotton/jujube intercropping systems examined, the roots of the trees spread beneath the cotton plants, and there were relatively lower RLDs and RDs at all soil depths than in the sole-cropped jujube tree plots. The roots of intercropped cotton tended to have a more shallow distribution in the soil profile and a lower RLD and RD compared with sole cotton. Moreover, the roots of intercropped jujube trees spread beneath the cotton but occupied a comparatively smaller soil space compared with the sole-cropped trees. Decreased soil exploration and apparent root competition led to decreases in yield and aboveground biomass. This provides direct evidence for the hypotheses that the lower biomass accumulation and yield in the tree-crop associations resulted from interspecific competition, lower RLD and RD in the soil and altered root spatial distribution. The roots of intercropped cotton were reduced more in the two cotton border rows on both sides compared with the middle rows. Jujube tree/cotton intercropping can achieve greater combined production than sole cropping. Understanding the dynamic processes of the interactions between jujube trees and cotton over time is important for farmers in Hetian oasis to evaluate their management options.
References
Allen SC, Jose S, Nair PKR, Brecke BJ, Nair VD, Graetz DA, Ramsey CL (2005) Nitrogen mineralization in a pecan (Carya illinoensis K. Koch)–cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Biol Fertil Soils 41(1):28–37
Ashton PS (2000) Ecological theory of diversity and its applications to mixed species plantation systems. In: Ashton MS, Montagnini F (eds) The silvicultural basis for agroforestry systems. CRC Press, Boca Raton, pp 61–77
Atkinson D, Johnson MG, Mattam D, Mercer ER (1978) The effect of orchard soil management on the uptake of nitrogen by established apple trees. J Sci Food Agric 30:129–135
Böhm W (1979) Methods of studying root systems. Springer, Berlin
Bolte A, Villanueva I (2006) Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur J For Res 125:15–26
Cao FL, Kimmins JP, Wang JR (2012) Competitive interactions in Ginkgo and crop species mixed agroforestry systems in Jiangsu, China. Agrofor Syst 84:401–415
Cardinael R, Mao Z, Prieto I, Stokes A, Dupraz C, Kim JH, Jourdan C (2015) Competition with winter crops induces deeper rooting of walnut trees in a Mediterranean alley cropping agroforestry system. Plant Soil 391:219–235
Chamshama SAO, Mugasha AG, Klovstad A, Haveraaen O, Maliondo SMS (1998) Growth and yield of maize alley cropped with Leucaena leucocephala and Faidherbia albida in Morogoro, Tanzan. Agrofor Syst 40:215–225
de Wit CT (1960) On competition. Verslaen van landbouwkd Onderz 66:1–82
Dhima KV, Lithourgidis AS, Vasilakoglou LB, Dordsas CA (2007) Competition indices of common vetch and cereal intercrops in two seeding ratio. Field Crop Res 100:249–256
Ding SS, Su PX (2010) Effects of tree shading on maize crop within a poplar-maize compound system in Hexi Corridor oasis, northwestern China. Agrofor Syst 80:117–129
Droppelmann KJ, Ephrath JE, Berliner PR (2000) Tree/crop complementarity in an arid zone runoff agroforestry system in northern Kenya. Agrofor Syst 50:1–16
Duan ZP, Gan YW, Wang BJ, Hao XD, Xu WL, Zhang W, Li LH (2017) Interspecific interaction alters root morphology in young walnut/wheat agroforestry systems in northwest China. Agrofor Syst. https://doi.org/10.1007/s10457-017-0133-2
García-Barrios L, Ong CK (2004) Ecological interactions in simultaneous agroforestry systems in the tropics: management lessons. Agrofor Syst 61:221–236
Gillespie AR, Jose S, Mengel DB, Hoover WL, Pope PE, Seifert JR, Biehle DJ, Stall T, Benjamin TJ (2000) Defining competition vectors in a temperate alley cropping system in the midwestern USA. 1 Production physiology. Agrofor Syst 48:25–40
Hendriks CMA, Bianchi FJJA (1993) Root density and root biomass in pure and mixed forests of Douglas fir and beech. Neth J Agric Sci 43:321–331
Hou QJ, Brandle J, Hubbard K, Schoeneberger M, Nieto C, Francis C (2003) Alternation of soil water content consequent to root-pruning at a windbreak/crop interface in Nebraska, USA. Agrofor Syst 57:137–147
Huang W, Kanninen M, Xu Q, Huang B (1997) Agroforestry in china: present state and future potential. Ambio 26:394–398
Imo M, Timmer VR (2000) Vector competition analysis of a Leucaena-maize alley cropping system in western Kenya. For Ecol Manag 126:255–268
Jama BA, Nair PKR, Rao MR (1995) Productivity of hedgerow shrubs and maize under alley cropping and block planting systems in semiarid Kenya. Agrofor Syst 31:257–274
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76:1–10
Jose S, Gillespie AR, Seifert JR (2000a) Defining competition vectors in a temperate alley cropping system in the Midwestern USA. 2. Competition for water. Agrofor Syst 48:41–49
Jose S, Gillespie AR, Seifert JR, Mengel DB, Pope PE (2000b) Defining competition vectors in a temperate alley cropping system in the mid-western USA. 3. Competition for nitrogen and litter decomposition dynamics. Agrofor Syst 48:61–77
Jose S, Gillespie AR, Pallardy SG (2004) Interspecific interactions in temperate agroforestry. Agrofor Syst 61:237–255
Jose S, Williams RA, Zamora DS (2006) Belowground ecological interactions in mixed-species forest plantations. For Ecol Manag 233:231–239
Kumar BM, Jose S (2017) Phenotypic plasticity of roots in mixed tree species agroforestry systems: review with examples from peninsular India. Agrofor Syst. https://doi.org/10.1007/s10457-016-0012-2
Livesley SJ, Gregory PJ, Buresh RJ (2000) Competition in tree row agroforestry systems. 1. Distribution and dynamics of fine roots length and biomass. Plant Soil 227:149–161
Mead R, Willey RW (1980) The concept of a land equivalent ratio and advantages in yields from intercropping. Exp Agric 16:217–228
Nerlich K, Graeff-Hönninger S, Claupein W (2013) Agroforestry in Europe: a review of the disappearance of traditional systems and development of modern agroforestry practices, with emphasis on experiences in Germany. Agrofor Syst 87:475–492
Ong CK, Huxley P (1996) Tree-crop interactions: a physiological approach. CAB International, Wallingford, p 386
Ong CK, Corlett JE, Singh RP, Black CR (1991) Above and belowground interactions in agroforestry systems. For Ecol Manag 45:45–57
Rao MR, Sharma MM, Ong CK (1990) A study of the potential of hedge row intercropping in semi-arid India using a 2-way systematic design. Agrofor Syst 11:243–258
Rao MR, Sharma MM, Ong CK (1991) A tree-crop interface design and its use for evaluating the potential of hedge row intercropping. Agrofor Syst 13:143–158
Schroth G (1995) Tree root characteristics as criteria for species selection and system design in agroforestry. Agrofor Syst 30:125–143
Schroth G (1999) A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agrofor Syst 43:5–34
Singh RP, Ong CK, Saharan N (1989) Above and below ground interactions in alley-cropping in semi-arid India. Agrofor Syst 9:259–274
Stone EL, Kalisz PJ (1991) On the maximum extent of tree roots. For Ecol Manag 46:59–102
Vandermeer J (1989) The ecology of intercropping. Cambridge University Press, New York
Wang BJ, Zhang W, Ahanbieke P, Gan YW, Xu WL, Li LH, Christie P, Li L (2014) Interspecific interactions alter root length density, root diameter and specific root length in jujube/wheat agroforestry systems. Agrofor Syst 88:835–850
Wanvestraut RH, Jose S, Nair PKR, Brecke BJ (2004) Competition for water in a pecan (Carya illinoensis K. Koch) – cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Agrofor Syst 60:167–179
Yang LL, Ding XQ, Liu XJ, Li PM, Eneji AE (2016) Impacts of long-term jujube tree/winter wheat–summer maize intercropping on soil fertility and economic efficiency—a case study in the lower North China plain. Eur J Agron 75:105–117
Yin R, He Q (1997) The spatial and temporal effects of Paulownia intercropping: the case of northern China. Agrofor Syst 37:91–109
Zamora DS, Jose S, Nair PKR (2006) Interspecific competition in a pecan–cotton alleycropping system in the southern United States: production physiology. Can J Bot 84:1686–1694
Zamora DS, Jose S, Nair PKR (2007) Morphological plasticity of cotton roots in response to interspecific competition with pecan in an alley cropping system in the southern United States. Agrofor Syst 69:107–116
Zhang W, Ahanbieke P, Wang BJ, Xu WL, Li LH, Christie P, Li L (2013) Root distribution and interactions in jujube tree/wheat agroforestry system. Agrofor Syst 87:929–939
Zhang DS, Zhang LZ, Liu JG, Han S, Wang Q, Evers J, Liu J, van der Werfc W, Li L (2014) Plant density affects light interception and yield in cotton grown as companion crop in young jujube plantations. Field Crops Res 169:132–139
Zhang W, Ahanbieke P, Wang BJ, Gan YW, Li LH, Christie P, Li L (2015) Temporal and spatial distribution of roots as affected by interspecific interactions in a young walnut/wheat alley cropping system in northwest China. Agrofor Syst 89:327–343
Zhang W, Wang BJ, Gan YW, Duan ZP, Hao XD, Xu WL, Lv X, Li LH (2016) Competitive interaction in a jujube tree/wheat agroforestry system in northwest China’s Xinjiang province. DOI, Agrofor Syst. https://doi.org/10.1007/s10457-016-9962-7
Acknowledgements
This work was financially supported by the NSFC (Project No. 31460335 and 31560376), by the Project Funded by China Postdoctoral Science Foundation, by Scientific Research Funds for High Calibre Researchers of Shehezi University (Project No. RCZX201422), by the Chinese Ministry of Science and Technology (Project No. 2009BADA4B03) and by the Chinese Ministry of Agriculture (Project No. 201003043-01). The authors also thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Zhang, W., Wang, B.J., Gan, Y.W. et al. Competitive interaction in jujube tree/cotton agroforestry system in Xinjiang province, northwestern China. Agroforest Syst 93, 591–605 (2019). https://doi.org/10.1007/s10457-017-0153-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-017-0153-y