
Abstract
In the southern region of Bahia, a large portion of the Atlantic Forest was occupied by the cacao–cabruca system, which is implemented after the complete removal of the understory vegetation without altering the canopy. The objective of this study was to determine the nitrogen concentration in the soil solution in two micro-basins; one with the cacao–cabruca system and one in the Atlantic forest in the southern region of the state of Bahia. Samples were collected weekly during two periods, from September to December 2012 and from April to June 2013, using sample extractors installed in the micro-basins at 15, 45 and 90 cm. The inorganic forms in the soil solutions were analyzed through ion chromatography, total nitrogen was analyzed using spectrophotometry and mineralization and nitrification rates were analyzed using the laboratory incubation method. Among the nitrogen forms analyzed in the cacao–cabruca soil solution, the dissolved organic nitrogen prevailed among the rain classes in the three depths. In the forest, nitrate predominated at 15 cm, while the organic nitrogen prevailed in the other depths. The highest mineralization and nitrification rates were recorded in the forest. Of the inorganic nitrogen forms analyzed in the soil, ammonium concentrations showed higher rates than nitrate in both areas. Low inorganic nitrogen concentrations in the cabruca soil solution are associated with low mineralization and nitrification rates. Thus we can conclude that even if some studies point towards the environmental efficiency of this system, there are differences in the N forms in the forest and cacao–cabruca areas.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The soil solution is an important section of the ecosystem because it is where part of the chemical processes of soil occurs, and it is easily altered according to use (Marques et al. 1996). The composition of the soil solution is the result of several processes such as the entry of elements via direct precipitation and internal precipitation, domestic production of nutrients by organic matter decomposition, weathering, loss by biological nutrient intake or leaching. Each process depends on several factors such as the depth of the soil, mobility of solutions and nutrient requirements of the plants. The low amount of nutrients available in the soil and the consequent drop in the concentration of these nutrients in the soil solution may be an indication that these nutrients are being quickly absorbed by the roots. Therefore, it is important that the supply of nutrients via internal precipitation and mineralization occur to supply this loss (Laclau et al. 2003).
Any type of land use affects the water cycle, no matter how much and how this use will depend upon the water. Studies in hydrography basins must consider their characteristics and the regions where they are installed since the exchange of energy and matter depend on the characteristic physiographic gradient of each basin (Espíndola et al. 2000).
This means that replacing the original forest cover by agricultural activities or the uncontrolled growth of urban areas may lead to serious changes in the processes that take place in a basin, such as the loss of soil nutrients. In southern Bahia, the cacao–cabruca system (Theobroma cacao) achieved a prominent position due to its economic importance, and much of the Atlantic Forest of the region was taken over by this type of cultivation. This system is put into place after the removal of the understory vegetation without any changes to the canopy, which favors the conservation of the native species of the forest (Alvim et al. 1986; Smith Dumont et al. 2014).
In spite of the reduction of cacao–cabruca production due to the Witches’ Broom, a fungus (Moniliophtera perniciosa) that swept through the south of Bahia in the 1990s, this agroforestry system is still very common in the Atlantic Forest areas of southern Bahia (Dawoe et al. 2010; Hartemink 2005; Ntiamoah and Afrane 2008). Some studies suggest that this planting system is sustainable because it maintains part of the canopy intact, so there are no changes to the structure and the functioning of these forest ecosystems (Barreto et al. 2010; Gama-Rodrigues et al. 2010; Silva Moço et al. 2009). Recent work developed at the same areas have shown that in rainy season dissolved organic carbon (DOC) export from cacao–cabruca system was three times higher than preserved forest (Costa et al. 2017). On the other hand, studies in shaded cacao farms have been showing that this traditional agroforests did not affected C storage when large tree are maintained (Schroth et al. 2013). Although some studies have been carried out demonstrating the influence of cocoa cultivation on C dynamics, little is known about the impacts of this cultivation on the soil nitrogen dynamics, which highlights the importance of understanding the consequences of such a system on the nutrient cycling of these watersheds and how they respond to this disturbance.
Thus, this study aims to determine the nitrogen concentration in the soil solution in micro-basins with the cacao–cabruca agroforestry system and the ombrophilous forest system, and to verify the influence of the cacao–cabruca system on nitrogen availability in the soil and the effects of this system on mineralization rates and net nitrification rates in soils. Our main hypothesis is that nitrogen availability is soil solution and net mineralization and nitrification will be similar between forest and cacao–cabruca system.
Materials and method
Study area
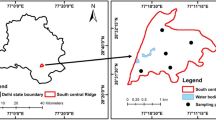
This study was conducted in two micro-basins; one in a preserved forest (PF) and the other in a cacao–cabruca system (CC), in southern Bahia, Brazil (Fig. 1). The climate is tropical wet (Af) according to the Köppen classification system, or hot and humid with no well-defined dry season. The average annual temperature is around 24 °C and the annual precipitation rates exceeds 1300 mm.

Source Modified from Márcio S. Inada
Location of study areas of the preserved forest and of the cacao plantation
The preserved area is a remnant of the Atlantic Forest classified as a Sub Montana Ombrophylous Dense Forest (IESB 2005) with a predominance of secondary vegetation, located in the Sierra State Park of Conduru (PESC). The vegetation consists of a native field that is quite similar to the herbaceous and shrub marshes. The PESC region has an extensive river system that, associated with the limitations of the relief for mechanization, favored the occupation of the land by properties, mostly with less than 100 ha. In addition, water is easily supplied to the properties for both domestic and agricultural use (Semarh 2012).
The geological formation is predominantly Precambrian rocks that form the local crystalline substrate, covered by tertiary-quaternary sediments. The soils range from sandy-clay to loamy-sandy, with some hydromorphic soils (Brazil, MMA 2004) and it is classified as typically petroplinthic Dystrophic Latosol (Oxisol) (IESB 2005).
The micro-basin with the cacao–cabruca system is inserted in the central corridor of the Atlantic Forest whose primary vegetation was the Ombrophilus Dense Forest. The soil of the area is classified as typical Dystrophic Argisol to moderate Eutrophic soil with medium clayey texture. The area has around 5% of slopes and an average of 70% of shading due to the cacao trees (Theobroma cacao L.), jackfruit trees (Artocarpus heterofolia), Cajá pulp trees (Spondias lutea), coral trees (Erythrina sp), Trumpet trees (Cecropia leucocoma) and a jequitibá tree (Cariniana sp), among other species (Sambuichi et al. 2012). In the last 15 years there has been no record of any management of the soil in this area.
Experimental design
Samples were collected weekly during two consecutive days, one day for each study area, between September 25 and December 7, 2012, and between April 2 and June 12, 2013, totaling 22 campaigns in the cabruca and 21 campaigns in the forest.
Analytical procedure
Soil solution
The soil solution samples were collected using tension lysimeters installed at depths of 15, 45 and 90 cm. In each basin, 12 extractors were installed in the surrounding of stream channel in a 1 ha plot, 1 m apart from each other, and two replicates of each depth were positioned on the right bank and two on the left bank of the creek. The soil solution was removed from the extractors by means of manual pressure. The samples were filtered using glass fiber filters (0.7 μm) previously calcined at 450 °C, placed in 100 ml polyethylene bottles and frozen for later analysis.
Dissolved inorganic nitrogen (DIN), nitrite (NO −2 ), nitrate (NO3 −) and ammonium (NH4 +) forms were analyzed using ion chromatography (DIONEX ICS 1000). Total dissolved nitrogen (TDN) was analyzed by spectrophotometry after the digestion of the sample using potassium persulfate (Grasshoff et al. 1983). Dissolved organic nitrogen (DON) forms were determined by calculating the difference between total inorganic (DIN) and total dissolved (TDN) forms.
Soil
To help characterize the areas, soil samples were collected at depths that ranged between 0 and 20, 20–40, 40–60, 60–80 and 80–100 cm using an auger-type pitcher “three”, followed by analyses of the amounts of phosphorus (P), aluminum (Al3+), calcium (Ca2+), magnesium (Mg2+), potassium (K+), CEC (ion exchange capacity) and soil particle size.
The soil samples used to characterize the areas were dried at 60 °C for further chemical and particle size analysis. The P, K+, Ca2+ and Mg2+ were analyzed using the extraction method for ion exchange resin, and Al3+ was analyzed using KCl 1 mol L−1 extraction. Regarding particle size, the sand was determined using the weighing method and the clay was determined by densitometry (with dispersant sol. sodium hydroxide and sodium hexametaphosphate, and/or dispersion in water); all analyses were performed in the Esalq/USP laboratory.
Three soil samples were also collected at 0–10 cm to determine the net mineralization and nitrification rates. To determine the net mineralization and nitrification rates, we used the modified laboratory incubation method according to Piccolo et al. (1994). The soil was extracted with 50 ml of KCl 2 N in about 10 g of moist soil. These portions were placed in jars with lids for 24 h, after which they were filtered using a Whatman 42 filter, preserved with 0.1 μM of HgCl and stored in amber bottles. The extraction was performed again after 5 days of soil incubation. The NO3 − and NH4 + concentrations in the soil extract were analyzed using spectrophotometry (Grasshoff et al. 1983). To determine the moisture of the soil, on the same day of collection, a sub sample of about 10 g was separated and dried at 60 °C until it reached constant weight.
The rates were calculated according to the following formulas:
where tn is the incubation time (5 days) and t0 is the original time (collection date).
Statistical analysis
The results were submitted to the nonparametric Kruskal–Wallis test to analyze possible significant differences (p < 0.05) between the analyzed nutrients and depths, the rain classes and the relationship between the mineralization and nitrification processes.
Results
The minimum and maximum precipitation figures recorded every collection week ranged between 0.1 and 13.5 mm in the cabruca and 0.2 and 8.7 mm in the forest. The rates were similar in most weeks in both collection areas, as shown in Fig. 2. In the cabruca, the highest rate was recorded in week 4, which corresponds to October 2012. In the forest, the highest rate was recorded in week 22, which corresponds to June 2013.

Precipitation registered between September/2012 and June/2013 in the Cocoa Cultivation (CC) and in the Forest (PF) and in southern Bahia
Physical and chemical composition of the soil
According to the results of the particle size analysis, sand fragments predominated in both areas, although the cabruca area showed a higher percentage of clay compared to the forest area. Consequently, the soil texture classes were characterized as medium-sandy in the forest and as medium-clay in the cabruca area.
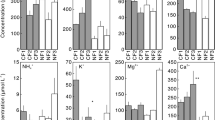
The largest concentrations of phosphorus (P), calcium (Ca), magnesium (Mg), sum of exchangeable bases (SB) and CEC saturation for bases (V) were recorded in the first centimeters of the soil in the cacao-cabruca system. The greatest acidity potential (H + Al) was observed in the cabruca soil at 40–60 and 60–80 cm. The lowest pH values and the highest CEC and V values were also observed at these depths (Table 1).
The soil solution sample volumes are related to the volume of rainfall in the weeks of collection and can influence the concentration of some forms of nitrogen. Rain classes were established in relation to the volume of rain in the collectors installed in the collection areas (Fig. 3).

Frequency of weeks collected from the rain of classes in the areas of Preserved Forest (PF) and Cocoa Cultivation (CC) in southern Bahia
The histogram represents the frequency of collection weeks related to the rain classes. The forest and the cabruca areas showed a higher frequency at 0–2 mm, which suggests that most collection weeks had low rain levels. In the forest area, the class with the lowest frequency was 4–6 mm. The class with the lowest frequency in the cabruca was 6–8 mm (Fig. 3).
DIN and DON concentrations in the soil solution
In the forest, the NO3 − concentrations predominated only at 15 cm and in all classes except at 6–8 mm. At 45 and 90 cm, the DON concentrations were higher than the DIN concentrations. At 45 cm, DON concentration variations were observed between classes, with significant differences (p < 0.05) between classes 2–4 and 4–6 and 2–4 mm and 6–8 mm (Fig. 4).

Weekly concentrations of DIN and DON (μM) in the soil solution of the Preserved Forest by class of rain
NH4 + concentrations showed close values between classes at 15 cm, except in the class 4–6. At 45 and 90 cm, lower NH4 + concentrations are observed in the 6–8 mm class, which is similar to the concentrations of the cabruca (Fig. 4).
Of all nitrogen forms analyzed in the soil solution of the cabruca, DON was the most common form in the rain classes in all three depths. Comparing the concentrations of DIN in the different classes, 6–8 mm showed the lowest concentrations of NH4 + in all three depths and higher concentrations in the 0–2 mm class (p < 0.05), except at 90 cm. This shows that higher concentrations were mostly observed during the weeks with the lower volumes of rainfall (Fig. 5).

Weekly concentrations of DIN and DON (μM) in the soil solution of the of Cocoa Cultivation by class of rain
NO3 − concentrations were lower than NH4 + in the first 15 cm in the four classes of rain. At 45 cm, the concentration was superior to NH4 + in classes 2–4 and 4-6 mm. At 90 cm, NO3 − concentrations showed similar values between classes. For each nitrogen form, no differences (p > 0.05) were observed between rainfall classes.
Comparing the DIN forms concentrations in the soil solution between the study areas showed that the averages of NH4 + concentrations are similar between the areas in the three depths. In the case of NO3 −, the highest concentrations were observed in the forest (Fig. 6a, b) and differences were found between the depths regarding the NO3 − concentrations in the forest and NH4 + concentrations in the cabruca, with decreasing concentrations as the depth increased (p < 0.05).

Mean concentrations of N forms in the depths 15, 45, 90 cm in the Preserved Forest (a) and Cacao–Cabruca system (b). Different letters indicate statistically different values (p < 0.05)
In the forest, the highest average of DIN and DON concentrations was observed at 15 cm, predominantly NO3 −. In the cacao-cabruca system, DON concentrations were higher than DIN, and the highest average was registered at 45 cm with a decrease at 90 cm.
Net mineralization and nitrification rates in soil
Of the inorganic N forms analyzed in the soil, Fig. 7 shows that N-NH4 + predominated in both study areas, with differences (p < 0.05) found between the concentrations of N-NH4 + and N-NO3 − in both areas.

N-NH4 + e N-NO3 − concentrations in the soil of Preserved Forest (PF) and Cocoa Cultivation (CC) in southern Bahia
The highest mineralization and nitrification rates were recorded in the forest, with differences (p < 0.05) observed between the mineralization rates of the forest and cabruca areas. The mineralization rate in the forest was higher than the nitrification rate, while the reverse was observed in the cabruca, with a predominance of nitrification (Fig. 8).

Rates of mineralization and net nitrification in the soil of Forest (F) and Cocoa Cultivation (CC) in southern Bahia
Discussion
Of the nitrogen forms analyzed, DON was the predominant form found in the cacao-cabruca system and NO3 − in the forest. The predominance of the NO3 − observed in the forest may be the result of minerals and organic matter transferred from the litter that covers the first few centimeters of soil, which is one of the factors that favors the highest NO3 − concentrations in the soil solution within the first 15 cm. Tropical soils are highly weathered and acidic and present high exchangeable aluminum concentrations, which reduce their fertility and increase nutrient loss by leaching. In this case, the organic matter decomposition has a fundamental role in soil fertility by increasing the CEC and releasing essential nutrients to the vegetation such as P, K, Na, Mg, and N (Baker et al. 2007).
When the conditions of the soil are favorable, such as good drainage and aerobic conditions, the development of aerobic organisms allows the oxidation of ammonium to nitrite, which, in turn, is oxidized to nitrate. Low concentrations of nitrite and ammonium and high nitrate concentrations are characteristic of Tropical Rainforests, where there is no nitrogen limitation for nitrification (Neill et al. 1995; Neu et al. 2011; Peterson et al. 2001).
Nitrification is an important process that influences the chemical composition of the soil solution, mainly in the upper layers of the soil. In the deeper layers, the biological activity becomes limited, which influences the mineralization and nitrification process and decreases nitrogen concentrations in these layers (Ranger et al. 2001). This pattern can be observed in the forest, where the DIN concentration in the soil solution, especially NO3 −, decreased as depth increased. Sudduth et al. (2013) observed that in non-affected hydrographic basins, the vegetation contributes significantly to the removal of nitrate in the ecosystems resulting in a decrease of nitrate concentrations in the soil below the area of the roots.
The lowest NO3 − concentrations in the soil solution in the cacao-cabruca system compared to the forest can be related to the exhaustion of some essential nutrients over time. In the cacao-cabruca systems, which are old, the litter layer is a major source of nutrients to the ecosystems but it is insufficient to cover the loss of nutrients (Dawoe et al. 2010; Selle 2007; Vitousek 1982). Although the litter layer was analyzed during this study, the nutrients in the soil solution are related to the quality of organic matter resulting from the decomposition of the layer of litter. Dawoe et al. (2010) in areas of cacao agroforestry found that decomposition may decrease after the conversion of the forest into these systems.
Similar behavior was observed in areas of silviculture, where concentrations of inorganic forms were lower in the soil solution when compared to the forest areas. This may be related to the low amount of nutrients available in the soil and the low levels of nutrients released by the weathering of soil minerals, thus emphasizing the importance of nutrient input through mineralization of the litter layer for ecosystems, especially in silviculture areas where litter is scarce (Laclau et al. 2003).
Low mineralization rates in the cacao–cabruca areas may be related to the low biogeochemical cycling rates of the cacao leaves and the low nutritional quality in deciduous leaves, which are part of the decomposed and mineralized organic matter (Gama-Rodrigues et al. 2010). Nitrogen absorbed by plants through the soil solution in the form of NO3 − or NH4 +, is incorporated into the biomass and subsequently returns to the soil via litter (Vitousek 1984).
The nitrification rate recorded in the forest in this study was similar to rates observed in this same area, and the averages ranged between 0.26 and 1.67 μg N g−1 day−1 (Silva, D.M.L. unpublished data). Piccolo et al. (1994); Neill et al. (1995, 1997, 1999) compared forest areas with grasslands and recorded higher mineralization and nitrification rates in the forest, thus concluding that when the forest is turned into pasture, the changes that affect these rates reflect the nitrogen biogeochemical and the fertility of the grassland over time. These rates in the grassland tend to decline with age, that is, as the older pastures dominate the deforested areas in the Amazon, the total N cycling in the soils of this region tends to decrease (Neill et al. 1999). In the cacao–cabruca system, differences between mineralization and nitrification rates were observed compared to the forest rates, and may be an indication that concentrations also decreased over time since this is an old system in southern Bahia.
Clay soils with high total organic carbon concentrations have high nitrification and mineralization rates. Low rates of these processes were recorded in pastures in the Amazon region where there is a predominance of clay soils, thus suggesting that other factors influence the nitrogen concentrations in the soil after the pasture is established (Neill et al. 1997). In this study, low nitrification and mineralization rates were also recorded in the cacao–cabruca system, although the soil of the region is characterized by Argisol, which is considered a fertile soil due to its clay content. The cabruca soil had a higher clay content than the forest area and higher concentrations of some nutrients that are important for fertility such as P, Ca2+ and Mg2+, high potential acidity (H + Al) and base saturation. The increase in Ca2+ and Mg2+ levels in this system may indicate the use of correctors which are used to reduce acidity and increase fertility. Thus, it is likely that soil fertility in the cacao-cabruca system is maintained by the application of these corrective substances. Differences between conservation areas and cacao systems were also observed in Oxisols in a basin of the river Una (southern Bahia) that had higher concentrations of base saturations, Ca2+ and Mg2+ in the cacao area (Barreto et al. 2010).
As for the inorganic nitrogen forms in the soil surface, N-NH4 + concentrations were higher than N-NO3 −. The N-NH4 + concentrations in the soil were similar in both areas, although the net mineralization rate in the forest was higher than in the cabruca. Thus, we can infer that the conditions that favor this process, namely microbial activity associated with environmental conditions such as temperature, moisture and aeration, may not be efficient in the cacao–cabruca system and may result in higher availability of N-NH4 + in the ecosystem. Zaia et al. (2012), in an Oxisol in southern Bahia, recorded lower rates of these processes in the cabruca when compared to a natural forest, but the cacao–cabruca system showed a greater ability to store organic carbon, total nitrogen, microbial biomass and N mineralization, which contributes to the improvement of soil quality.
Soil moisture can be an important way to control inorganic nitrogen forms and mineralization and detention rates of N in the soil. The ammonification process is usually less sensitive to environments with dry soils when compared to nitrification. Dry periods may promote mineralization of N, which can increase ammonium concentration (Peterjohn and Correll 1984; Luizão et al. 1992; Neill et al. 1995). In this study, the ammonium concentrations were higher during the weeks with a low rain class (0–2 mm).
Conclusion
Nitrogen distribution in soil solution was distinct between two areas, mainly in the upper 15 cm layer of soil with nitrate being predominant in the forest and the organic forms predominant in cocoa. This may be a result of the decrease of net mineralization and nitrification in cacao–cabruca system compared to forest. On the other hand, at higher depths, NO3 − decrease in soil solution probably due to biological activities and the nutrients removal by plants. Thus, despite the initial hypothesis, the cacao–cabruca system with no soil management in the last 15 years, modified the nitrogen distribution in soil solution at the upper soil layers.
References
Alvim R, Nair PKR (1986) Combination of cacao with other plantation crops: an agroforestry system in Southeast Bahia, Brazil. Agroforestry Syst 4:213–220
Baker JM, Ochsner TE, Venterea RT, Griffis TJ (2007) Tillage and carbon sequestration—What do we really know? Agric Ecosyst Environ 118:1–5
Barreto PAB, Gama-Rodrigues EF, Fontes AG, Polidoro JC, Machado RCR, Baliga VC (2010) Distribution of oxidizable organic C fractions in soils under cacao agroforestry systems in Southern Bahia, Brazil. Agrof Sys 81:213–220
Costa END, Souza JC, Pereira MA, Souza MFL, Souza WFL, Silva DML (2017) Influence of hydrological pathways on dissolved organic carbonfluxes in tropical streams. Ecol Evol 7:228–239
Dawoe EK, Isaac ME, Quashie-as J (2010) Litterfall and litter nutrient dynamics under cacao ecosystems in lowland humid Ghana. Plant Soil 330:55–64
Espíndola ELG, Vila JSV, Marinelli CE, Abdon MM (2000). A bacia hidrográfica do Rio Monjolinho. Rima, São Carlos – SP
Gama-Rodrigues EF, Nair PKR, Nair VD, Gama-Rodrigues AC, Baligar VC, Machado RCR (2010) Carbon storage in soil size fractions under two cacao agroforestry systems in Bahia, Brazil. Environ Manag 45:274–283
Grasshoff K, Erhardt M, Kremling K (1983) Methods of seawater analysis. Verlag Chemie, Weinheim
Hartemink AE (2005) Nutrient stocks, nutrient cycling, and soil changes in cacao ecosystems, A review. Adv Agron 86:227–253
Instituto de estudos sócio-ambientais do sul da Bahia - IESB. Plano de Manejo do Parque Estadual da Serra do Conduru (PESC), 2005
Laclau JP, Ranger J, Nzila JD, Bouillet JP, Deleporte P (2003) Nutrient cycling in a clonal stand of Eucalyptus and an adjacent savanna ecosystem in Congo 2. chemical composition of soil solutions. For Ecol Manage 180:527–544
Luizão RC, Bonde TA, Rosswall T (1992) Seasonal variation of soil microbial biomass—the effect of clearfelling a tropical rainforest and establishment of pasture in the central Amazon. Soil Biol Biochem 24:805–813
Marques R, Ranger J, Gelhaye D, Pollier B, Ponette Q, Goedert O (1996) Comparison of chemical composition of soil solutions collected by zero-tension plate lysimeters with those from ceramic-cup lysimeters in a forest soil. Eur J Soil Sci 3:407–417
Neill C, Piccolo MC, Steudler PA, Melillo JM, Feigl BJ, Cerri CC (1995) Nitrogen dynamics in soils forests and active pasture in the western Brazilian Amazon Basin. Soil Biol Biochem 27:1167–1175
Neill C, Piccolo MC, Cerri CC, Steudler PA, Melillo JM, Brito M (1997) Net nitrogen mineralization and net nitrification rates in soils following deforestation for pasture across the southwestern Brasilian Amazon Basin landscape. Oecologia 110:243–252
Neill C, Piccolo MC, Melillo JM, Steudler PA, Cerri CC (1999) Nitrogen dynamics in Amazon forest and pasture soil measured by 15 N pool dilution. Soil Biol Biochem 31:567–572
Neu V, Neill C, Krusche AV (2011) Gaseous and fluvial carbon export from an Amazon forest watershed. Biogeochemistry 105:133–147
Ntiamoah A, Afrane G (2008) Environmental impacts of cacao production and processing in Ghana, life cycle assessment approach. J Clean Prod 16:1735–1740
Peterjohn WT, Correll DL (1984) Nutrient dynamics in an agricultural watershed, observations on the role of A Riparian Forest. Ecology 65:1466–1475
Peterson BJ, Wollheim WM, Mulholland PJ, Webster JR, Meyer JL, Tank JL, Marti E, Bowden WB, Valett HM, Hershey AE, Mcdowell WH, Dodds WK, Hamilton SK, Gregory S, Morrall DD (2001) Control of nitrogen export from watersheds by headwater streams. Science 292:86
Piccolo MC, Neill C, Cerri CC (1994) Net nitrogen mineralization and net nitrification along a tropical forest-to-pasture chronosequence. Plant Soil 162:61–70
Ranger J, Marques R, Jussy JH (2001) Forest soil dynamics during stand development assessed by lysimeter and centrifuge solutions. For Ecol Manage 144:129–145
Sambuichi RHR, Vidal DB, Piasentin FB, Jardim JG, Viana TG, Menezes AA, Mello DLN, Ahnert D, Baligar VC (2012) Cabruca agroforests in southern Bahia, Brazil: tree component, management practices and tree species conservation. Biodiver Conse 21:1055–1077
Schroth G, Bede LC, Paiva AO, Cassano CR, Amorim AM, Faria D, Mariano-Neto E, Martini AMZ, Sambuichi RHR, Lôbo RN (2013) Contribution of agroforests to landscape carbon storage. Mitig Adapt Strat Glob Change 20:1175–1190
Selle GL (2007) Nutrient cycling in forest ecosystems. Biosci J 23:29–39
Semarh (2012). Parque Estadual da Serra do Conduru, importância do Parque Estadual da Serra do Conduru. http://www.semarh.ba.gov.br/conteudo.aspx?s=PESERRACandp=PARQEST. Accessed 04 March 2012
Silva Moço MK, Gama-Rodrigues EF, Gama-Rodrigues AC, Machado RCR, Baligar VC (2009) Soil and litter fauna of cacao agroforestry systems in Bahia, Brazil. Agroforest Syst 76:27–138
Smith Dumont E, Gnahoua G-M, Ohouo L, Sinclair F-L, Vaast P (2014) Farmers in Cote d’Ivoire value integrating tree diversity in cocoa for the provision of ecosystem services. Agroforestry Sys 88:1047–1066
Sudduth EB, Perakis SS, Bernhardt ES (2013) Nitrate in watersheds, Straight from soils to streams? J Geophys Res Biogeosci 118:1–12
Vitousek PM (1982) Nutrient cycling and use efficiency. Am Nat 4:553–572
Vitousek PM (1984) Litterfall, nutrient cycling, and nutrient limitation in tropical forest. Ecology 65:285–290
Zaia FC, Gama-Rodrigues AC, Gama-Rodrigues EF, Moço MKS, Fontes AG, Machado RCR, Baligar VC (2012) Carbon, nitrogen, organic phosphorus, microbial biomass and N mineralization in soils under cacao agroforestry systems in Bahia, Brazil. Agroforest Syst 86:197–212
Acknowledgements
We wish to thank FAPESB—Fundação de Amparo a Pesquisa do Estado da Bahia and CNPq (PPP0040/2011) and Universidade Estadual de Santa Cruz (PROPP 0220.1100.899). We also thank to Marcio S. Inada for given map with site collects, to Divanete Souza and Veet Pramad and to all technicians at UESC and “Sítio Pachamama” for helping in field assistance. We are grateful to TIKINET and Cipriana Leme for the English revision of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
de Souza, J.C., Pereira, M.A., da Costa, E.N.D. et al. Nitrogen dynamics in soil solution under different land uses: Atlantic forest and cacao–cabruca system. Agroforest Syst 92, 425–435 (2018). https://doi.org/10.1007/s10457-017-0077-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-017-0077-6