
Abstract
In extensive low input farming and in agroforestry systems, the importance for biodiversity of managed productive fields with respect to unmanaged marginal habitats that occupy a low proportion of farm surface, is still poorly understood, contrasting with the well-known key importance of marginal habitats in intensive systems. We analyzed the importance of open and wood pastures and marginal habitats for species richness of Iberian dehesas in Central-Western Spain. We sampled 155 plots classified into 9 general habitat categories: wood pastures (n = 41 plots); open pastures dominated by annual plants (n = 11), by perennial plants (n = 15) and co-dominated by annuals and perennial plants (n = 16); shrublands (n = 19); agricultural crops (n = 12); herbaceous strips (n = 10); woody strips (n = 11); and water bodies (n = 10). In each plot we measured the abundance and species richness of four taxonomic groups: vascular plants, bees, spiders, and earthworms. We detected 431 plant species (37 ± 2.5 CI95 in 100 m2 on average), 60 bee species (3.1 ± 1.1 in 600 m2), 128 spider species (7.4 ± 1.2 in 1.5 m2) and 18 earthworm species (2.5 ± 1.0 in 0.27 m2) in 145 sampling plots. Wood pastures supported fewer species of spiders and earthworms at the plot level, but more plants and earthworm species at the landscape level than open pastures. The low proportion of shared species among habitats and among plots within each habitat type, and the high proportion of species found in unique plots or habitats indicated that every habitat contributes to farm biodiversity. Overall, our extensive survey confirms the hypothesis that the high diversity of dehesas depends on the coexistence within farms of a wide mosaic of habitats, including marginal habitats, which seemed to harbor a disproportionately high number of species as compared to their small extent. Results support policy measures for the maintenance of farm keystone structures such as linear features, small wood/shrub patches and ponds, and reveal that these measures should not be exclusively applied to more intensive farming systems.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Agricultural intensification has led to a recent widespread decline in farmland biodiversity across many different taxa (Benton et al. 2003; Kleijn et al. 2011), but farmland still hosts many species that depend on appropriate agricultural management for their survival (Kleijn and Sutherland 2003; Opermmann et al. 2012). For instance, it is estimated that roughly 50 % of plant and animal species in Europe depend on agricultural habitats (Kristensen 2003). Biodiversity protection globally depends increasingly on maintaining biodiversity in human-dominated landscapes (Fahrig et al. 2011), and it has been argued that measures targeted to reduce inputs in high-intensity farms should be the most cost-effective policy option (Primdahl et al. 2003). Nevertheless, assessments of the ecological effectiveness of agri-environmental schemes granted in Europe (EC 2005) which aimed at increasing biodiversity on intensive farmland have given mixed results (Whittingham 2007; Kleijn et al. 2011). The same can be said of extensification measures such as organic farming (e.g. Bengtsson et al. 2005; Hole et al. 2005; Gabriel et al. 2013), set-aside (Kleijn and Baldi 2005) or the conversion of some production lands into ‘more-natural’—unmanaged or extensively managed—lands (Flohre et al. 2011). The reasons for these general failures seems to be complex and taxa-specific, including non-linear relationships between field-scale land-use intensity and diversity (Kleijn et al. 2008), landscape-scale effects on field-scale land-use changes (Concepción et al. 2008) and even regional-scale effects of land use history or regional species pools (e.g., Tscharntke et al. 2005, 2012).
Conserving what is left, rather than trying to recover what was lost, has been proposed as a more effective approach to biodiversity conservation in low-intensive farming areas that still retain significant amounts of semi-natural vegetation (Kleijn et al. 2011; Concepción et al. 2102, Opermmann et al. 2012). These criteria are mostly accomplished by extensive pasturelands, which consist of mixtures of grassland, scrub and/or woodland used for raising livestock (Paracchini et al. 2008). Indeed, these kinds of farmland dominate current lists and maps of European High Nature Value Farming Systems, which are low input farms managed extensively, that still conserve high biodiversity (Opermmann et al. 2012).
Habitat heterogeneity at multiple spatial scales has been revealed as key for biodiversity conservation in farmland (Benton et al. 2003; Concepción et al. 2008, 2012). At the farm scale, which is the main management unit for decision-making, heterogeneity can be either intrinsic to the main land uses (e.g. scattered trees over grasslands in silvopastoral systems) or be the result of the diversity of land uses and/or the existence of marginal habitats (i.e. unmanaged habitats that occupy a low proportion of farm surface, including hedges, woodlots, roadsides or boundary strips interspersed among the main fields). Both sources of heterogeneity are important in agroforestry systems which, as part of multifunctional working landscapes, have been shown to play a major role in conserving and even enhancing biodiversity from farms to landscapes in both tropical and temperate regions of the world (review in Jose 2012). In these systems, scattered trees generate fine-scale mosaics within plots (i.e., gradients of available resources availability from beneath the canopy to interstitial areas among trees; Moreno et al. 2013) causing positive effects on biodiversity disproportionate to tree cover (reviews in Manning et al. 2006; Marañón et al. 2009; Díaz et al. 2013). Scattered trees are still prominent features of many human-modified landscapes around the world (Gibbons et al., 2008). Indeed, 46 % of the world agricultural land (10.1 M km2) includes scattered trees or woodlots (Zomer et al. 2009), and 5.2 M km2 are home to different wooded silvopastoral systems in the Mediterranean, North and South America, Central Asia, South Africa and South Australia (Calama et al. 2010; Pulido, unpublished).
Oak dehesas and montados are agroforestry systems that cover over 4.5 million hectares in the Iberian Peninsula (Moreno and Pulido 2009). These systems maintain outstanding levels of biodiversity (Díaz et al. 1997 and Díaz et al. 2013), to the point of being considered as habitats to be protected under the European Habitats Directive, the cornerstone of Europe’s nature conservation policy that protects over 200 habitat types important for biodiversity (EEC 1992). This low input system typically combines within farms livestock rearing (ca. 03–0.6 Livestock Unit per ha), cereal cropping, cork and firewood harvesting and game production, and is the source of both high-quality food products and ecosystem services such as carbon sequestration, recreation, watershed maintenance or biodiversity conservation (Campos et al. 2013). High levels of plant, arthropod, earthworm and vertebrate diversity from plot to landscape scales, including several endangered species, have been attributed to intimate mixtures of forest and open habitat types at several spatial scales in oak dehesas and montados (as Díaz et al. 2013). This finding has been tested mainly for the local effects of scattered trees on plants, birds and invertebrates (e.g. Marañón 1986) and for the landscape-scale effects of the coexistence of dehesas with forest patches (e.g.; Carrete and Donázar 2005) in few case studies. Even fewer works have been done on the role of marginal habitats or landscape elements (but see Martín and López 2002). Biodiversity conservation is key for the long-term sustainability of dehesa systems, due to the economic and amenity value given by both landowners and the society to such diversity (Campos et al. 2013). Hence, explicit long-term strategies should be designed to promote management practices aimed at maintaining such biodiversity. Such measures must rely on large-scale studies analyzing the patterns of dehesa diversity and more knowledge on the processes underlying them is urgently needed.
In this work we analyze the relative importance of wood pastures (defined as herbaceous pastures with scattered trees), open pastures (without trees) and marginal habitats of dehesa farms for the species richness of four key taxonomic groups: vascular plants (primary producers), bees (pollinators), spiders (predators), and earthworms (decomposers). These groups were selected because their limited mobility allows attributing community composition mostly to local-scale effects of tree-based gradients and within-farm habitat heterogeneity. We surveyed species richness and abundance in the two main habitat types, open pastures and wood pastures (pastures with scattered trees), and in marginal habitats (linear features, water bodies, shrub patches). The effect of trees was assessed by comparing species richness of wood pastures with open pastures both at plot and habitat levels. We expected more species in wooded main habitats and an absence of certain species of open pastures due to the negative effects of woody vegetation on these species. The importance of marginal habitats was analyzed by computing the proportions of shared species among habitat types and, as marginal habitats are expected to increase species richness by supporting species not found in open and wood pastures (Díaz et al. 2013), by comparing the estimated species richness in landscapes with or without them.
Materials and methods
Study area
The study was conducted in a typically agricultural region of Central Western Spain (Tierras de Granadilla district; 235 km2 from latitude 40º7′ to 40º14′N and longitude 6º0′ to 6º21′W). According to Corine Land Cover (EEA 2010), the landscape in the district is dominated by oak dehesas (38.7 % of the land) and open pastures (18.5 %) devoted to livestock breeding, olive plantations (15.0 %), shrublands (12.5 %), dense forests (9.4 %), and herbaceous crops (3.1 %). Dehesas are mostly dominated by scattered Quercus ilex trees, with Quercus suber and Quercus pyrenaica in lower numbers.
Ten dehesa farms (485 ha on average, ranging from 150 to 835 ha) were randomly selected, and then each area-based habitat (at least 5 m wide and covering 400 m2) and linear feature (at least 0.5 m wide and 30 m long) was mapped according to a standardized protocol developed by the European EBONE and BioBio projects (Bunce et al. 2011). Habitats were defined on the basis of the dominant Raunkiaer life forms, edaphoclimatic conditions, and management. Vegetation can be formed by a single life-form class or more often by a complex mixture of life forms; hence a new habitat is mapped and separated from adjacent or surrounding habitats if a change of more than 30 % cover of a life form is recorded.
In total, we defined nine General Habitat Categories (GHC): six area-based habitats and three linear features. Area-based GHC were: (1) Wood pastures (typically 10–30 mature oak trees per ha; 30–60 cm of stem diameter and 6–8 m height, on average; <30 % of canopy cover); (2–4) Open pastures dominated either by annual species (vegetative period from October to May), by perennial species (dried in summer for 3–4 months), or by a mix of both; (5) Shrub encroached pastures (typically 40–80 % of shrub cover and 1.5–3 m height; mostly Cistus spp., Retama sphaerocarpa, Genista hirsuta and Cytisus spp., but also Thymus spp, and Lavandula sp with or without presence of a sparse tree layer); (6) Agricultural crops, either herbaceous (usually cereal) or woody (olive, vineyards and fruit trees). Linear features were: (7) Water (seasonal streams and artificial ponds); (8) Herbaceous lines and (9) Wood lines (either tree species, usually oaks, but also Fraxinus angustifolia and Salix atrocinerea and/or shrub species (Flueggea tinctoria, Rosa canina, Rubus ulmifolius, Crataegus monogyna, Arbutus unedo, Cistus spp. and Cytisus spp.). Table S1 (supplementary material) gives a complete list and details on plots and habitat sizes.
Sampling protocol
Within each farm, plots were randomly selected for monitoring biodiversity. The number of plots selected was roughly proportional to the area occupied by each GHC: Wood pastures (WoodPast; n = 41 plots), Annual plant open pastures (AnnPast; n = 11), Perennial plants open pastures (PerenPast; n = 15), Mix open pastures (MixPast; n = 16), Shrubs (n = 19), Agricultural crops (AgriCrop; n = 12), Woody lines (WoodLine; n = 11), Herbaceous lines (HerbLine; n = 10) and Water bodies (Water; n = 10). In each plot (n = 145) four taxa were monitored, attending to the four major ecological functions which are relevant for farming: vascular plants (primary production), wild bees and bumblebees (pollination), spiders (predation), earthworms (organic matter decomposition). These four biological groups are relatively easy to monitor, provide relevant information on general environmental conditions and are sensitive to management practices (Herzog et al. 2013).
Vegetation was recorded in subplots of 10 m × 10 m in area-based habitats, and of 10 m × 1 m in linear features in spring 2010. All species were carefully identified and recorded in situ. the whole subplot area. Cover was visually estimated for each species, using 5 % categories. Species with less than 5 % cover were given a nominal cover of 1 %. Bare ground includes leaf litter and rock. Total cover may be over 100 % if several layers are present (Dennis et al. 2012). Surveyed area was always randomly located by the center of the plot, and for water bodies (water courses and ponds) surveys were done along the perimeter.
Bees and bumblebees (hereafter ‘bees’) were sampled along a slow walked transect of 100 m × 2 m per plot (alternatively, two 50 m × 2 m transects) with a handheld net (solid wire hoops with diameter of 46 cm). Captured specimens were immediately transferred into a killing jar charged with ethyl acetate to minimize damages and facilitate taxonomic identification. Transects were repeated three times throughout the season (from early May to mid-July 2010).
Spiders were sampled in 5 circular subplots (0.357 m diameter) placed beforehand at random on the target vegetation within the plot. Spiders were caught using a motorized leaf blower (inverted to allow suction) and immediately separated in situ from the litter and dust and fixed in 70 % diluted alcohol to avoid any possible predation among collected individuals. Sampling was repeated three times throughout the season (from late April to late July 2010). For each date, 5 subplots were newly selected within the same plot.
Earthworms were sampled in three separated quadrats per plot (30 cm × 30 cm) combining the extraction with an expellant solution (diluted AITC solution [allyl isothiocyanate]; 0.1 g/l; 2 l per quadrat poured twice at 5 min intervals) for 30 min and the subsequent hand-sorting for 30 min more. Samples were taken from mid-March to late April of 2010. Earthworms were fixed with diluted alcohol and stored at 4 °C before identification. All individuals were identified to species, except for the genera Andrena (bees), Scytodes and Tegenaria (spiders), and Allium, Conyza, Pohlia and Riccia (plants). For more details on sampling protocols see Dennis et al. (2012).
Data analysis
To facilitate the statistical treatment of a high number of habitats differing in extension and number of sampled plots, habitats were first grouped in the 9 GHC aforementioned. Second, the three categories of open pastures (annual-plants, perennial-plants and mix) were grouped under a single group named Open pastures to be compared against Wood pastures. Finally, GHC were further grouped into two categories, Pastures (wood pastures and open pastures) and Marginal habitats (the remaining 5 GHCs that occupy small areas in the farms).
For each biological group, the sum of species observed over sub-sampling squares and/or dates gives the species richness at each plot. Mean species richness at plot level (Splot) was calculated as the average number of species over the plots of each GHC. Species richness at GHC level (Shabitat) was calculated as the sum of species observed over all sampled plots within the category. Splot represents the α diversity for each GHC (or group of GHC), while Shabitat represents the γ diversity for each GHC. Comparison of both values gives estimation on the spatial heterogeneity or β diversity within each GHC. Pooling species recorded in the whole of plots, regardless of the habitat type, gives γ diversity at dehesa landscape scale.
Values were standardized following the rarefaction (interpolation) and extrapolation (prediction) approaches proposed by Chao (2005) and Colwell et al. (2012) to make comparable values produced with different sampling efforts. EstimateS 9, and open source software (Colwell 2013), was used to compute two estimators of species richness, Coleman and Chao-2, from counts of individuals of each species in single samples or set of samples. Whereas Chao-2 follows the extrapolation approach to estimate the total species richness, including species not present in any sample, Coleman uses rarefaction to estimate the species richness for a sub-set of samples (the lowest number of samples among the units compared). We also used EstimateS 9 to compute the compositional similarity among plots (within each GHC) and among GHCs. We selected a widely used estimator of shared species (Sorensen index), as well as an estimator of the number of shared species that takes shared but unobserved species into account (Chao-Sorensen Estimator; Chao et al. 2005; see Supplementary Material).
Mean values of abundance (number of individuals per surface unit) and species richness at plot level (Splot index) were compared by mean of generalized linear mixed models (GLMMs), with farm as random factor, and GHC (and further GHC groups) as the unique fixed factor. Tree and shrub cover were included as covariates to better discern the role of trees and shrubs for the four taxonomic groups. Comparison of species richness at the habitat level (Shabitat) was based on the comparison of the species accumulation curves and associated confidence intervals (Colwell et al. 2012). As these authors noted non-overlap of 95 % confidence intervals can be used as a simple but conservative criterion of statistical significance (p < 0.05). Mean values of similarity among plots were compared by means of generalized linear models (GLMs) with GHC as the unique factor and Sorensen and Chao-Sorensen indexes as response variables.
Results
Overall species richness
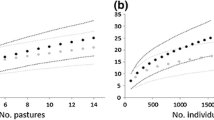
The number of species recorded per plot (Splot) was 37.0 ± 2.5 for plants (in 100 m2), 3.1 ± 1.1 for bees (in 200 m2 transect × 3 times), 7.4 ± 1.2 for spiders (in 0.5 m2 × 3 times) and 2.5 ± 1.0 for earthworms (in 0.27 m2). Pooling the data from the 145 sampled plots (species richness at dehesa landscape scale), the total number of species found was 431 for plants, 60 for bees, 128 for spiders and 18 for earthworms. Considering that 91 species of plants, 39 of bees, 34 of spiders and 7 of earthworms were recorded in just one plot, and that the species accumulation curves did not reach the asymptote (Fig. 1 and Supplementary Fig. S1), the actual species richness for dehesas in the study area is probably above the values found. Accordingly, the estimated richness for the entire 10 dehesa farms (Chao2 mean ± S.D.) was 504 ± 20 species for plants, 140 ± 40 for bees, 161 ± 14 for spiders and 25 ± 7 for earthworms.

Species accumulation curves (±95 % C.I.) for the four biological groups where data of all measured plots are randomly accumulated (n = 145). Rarefaction is indicated by solid lines, extrapolation by dashed lines, and the reference samples (measured) by the transition from solid to dashed line
Species richness per habitat type
Among the nine GHC types, differences in the mean number of species per plot (Splot) were significant for plants and spiders, marginally significant for bees and non-significant for earthworms (Tables 1, 2). Plant Splot was higher for shrub plots (48.9 ± 3.1 Confidence Interval at 95 %) followed by the four types of pastures (from 39.3 ± 2.8 to 37.4 ± 3.0) and agricultural crops (36.1 ± 2.7). Spider richness was affected by shrub cover (p < 0.001; Table 1), and Splot was higher in shrub plots (9.2 ± 1.2), with similar values for marginal habitats and open pastures dominated by perennial species, and significant lower values for other open values and wood pastures (6.1 ± 1.0, Table 2). Spiders and earthworms were more abundant in pastures (included herbaceous lines) and near water bodies and less abundant in woody habitats (wood pastures, wood lines and shrub plots). Bees were more abundant in wood lines and open pastures dominated by annual species, and Splot was marginally higher in wood lines (5.0 ± 1.2) than in remaining habitats (from 2.0 ± 0.7 to 3.5 ± 1.1; Table 2). The estimated percentages of shared species (Chao-Sorensen index) among two different plots was usually below 60 %, with mean percentages of 57.1 % for plants, 52.8 % for bees, 43.8 % for spiders and 51.1 % for earthworms. There were significant differences among GHCs for both Sorensen and Chao-Sorensen indexes (Table 3).
The overall richness recorded per GHC (Shabitat) was 5.2, 5.5, 6.5 and 3.2 times higher than Splot for plants, bees, spiders and earthworms respectively (Table 2). Moreover, Chao2 index values were 39 % for plants, 100 % for bees, 75 % for spiders and 34 % for earthworms higher than Shabitat, (Table 2). According to mean values and associated standard deviations of the Chao2 index, estimated plant richness was significantly higher in wood pastures and shrubs (380 ± 24 and 346 ± 24 species, respectively), and significantly lower in herbaceous lines (150 ± 24) and around water bodies (183 ± 18) (Table 2; see also Fig. 3a in the Supplementary Material). These differences were also confirmed by Coleman-rarefied richness (Table 2). For spiders, again wood pastures showed the highest Chao2 richness (129 ± 23 species), despite of their low Splot (6.1 ± 1.0). Richness was also high in shrubs and woody lines, especially as estimated by Coleman richness index. As for plants, herbaceous lines and around water bodies were significantly poorer than the rest of GHCs. Agricultural crops and wood lines showed higher richness of earthworms (Chao2 richness 23.8 ± 12.4 and 16.6 ± 5.1 species, respectively) than the remaining habitats. Uncertainty for the Chao2-estimated bee richness was high and differences among GHCs were less clear.
Wood pastures versus open pastures
Abundance and species richness of earthworms and spiders measured at plot level (Splot) was significantly higher in open pastures than in wood pastures (Tables 1, 2), which was confirmed by the significance of tree cover as covariate (species richness decreased with tree cover). Tree cover had also a negative effect on bee abundance. On the contrary, pooling plots estimated richness (Chao2) of plants and earthworms species were significantly higher in wood than in open pastures (Figs. 2, 3). Differences for plants, earthworms and spiders were also confirmed by Coleman-rarefied index (at n = 40) (Fig. 3).

Comparison of species accumulation curves (±95 % C.I.) for wood pastures and open pastures. Rarefaction is indicated by solid lines, extrapolation by dashed lines, and the reference samples (measured, n = 41 for wood pastures and n = 42 for open pastures) by the transition from solid to dashed line

Species richness (±95 % C.I.) measured and estimated by extrapolation (Chao2 index) and rarefaction (Coleman index; set at n = 40) for different groups of GHC
Main habitats versus marginal habitats
Regarding the comparison among all types of pastures and marginal habitats, differences for Splot were significant for plants, spiders and earthworms (Table 1). While for plant species measured richness was significantly higher in pastures than in marginal habitats, for earthworms the opposite was found. Results vary for estimated richness at the habitat level (Shabitat; Fig. 3). Marginal habitats were significantly richer than pastures for plants and earthworms (higher Chao2 and Coleman-rarefied indexes), and for bees and spiders (only with Coleman-rarefied index).
Shared species among habitats
Comparing the whole list of species recorded for each GHC type (all sampled plots pooled), the percentages of shared species was low (below 60 % for bees and spiders and 65 % for plants; Table 4). Only for earthworms the percentage of shared species was high (mostly above 65 %). Some specific combinations produced very low rates of shared species (<40 %). These were herbaceous lines with wood pastures and wood lines for plants; perennial pastures with water bodies and wood lines, and wood lines with herbaceous lines for bees; wood pastures and shrubs with herbaceous lines for spiders. Wood pastures, the most widespread GHC comprising 63 % of the dehesa area, only shared on average 56.5 % of their species with the rest of GHCs. Also linear features and water bodies shared only ca. 56 %, on average, of their species with the rest of GHCs (Table 4).
As a consequence of the low percentage of shared species among GHCs, when different GHCs were pooled (from the most ubiquitous and extended habitats up to the most rare and restricted ones) new additional species were provided by each new GHC (Fig. 4). Wood pastures only harbored 58 % of the species recorded (four taxa averaged) or 67 % of the number of species estimated (Fig. 4). Open pasture harbored 66 species of plants, 21 of bees, 24 of spiders and 1 of earthworms, that were not present in wood pastures (Figs. 3, 4). Marginal habitats (shrubs, agricultural crops, water bodies and linear features, which all together occupy less than 10 % of the total area) provided a good number of exclusive species that were not found in the main fields (wood plus open pastures). These exclusive species amount up to 26.2 % of the plants, 65 % of the bees, 50 % of the spiders and 55.6 % of the earthworm species (Table 5).

Relative accumulation of species with different habitats types compared with the relative accumulation of surface occupied by those habitats: Wood pastures (WoodPast;), Annual-plant open pastures (AnnPast), Perennial-plants open pastures (PerenPast), Mix open pastures (MixPast), Shrubs, Agricultural crops (AgriCrop), Woody lines (WoodLine), Herbaceous lines (HerbLine) and Water bodies (Water). For this curves with absolute values see Fig. S4 in the Supplementary Material
Discussion
The Mediterranean Basin is a recognized biodiversity hotspot (Myers et al. 2000) due to a complex biogeographical history, its transitional nature from large dry-tropical to wet-temperate biomas, and a long history of multiple land uses (Blondel et al. 2010). Among these land uses, extensive silvopastoral systems such as Iberian dehesas and montados are also renowned as biodiversity-rich systems (Bugalho et al. 2011; Díaz et al. 2013). Our study confirmed this high biodiversity, with an estimated richness of 504 species of plants, 140 of bees, 161 of spiders and 25 of earthworms for the dehesas of the study area (4875 ha). High levels of biodiversity of vascular plants, butterflies, birds and other vertebrates have been also found by other authors, a fact that presumably results from the intimate mixtures of forest and open habitat types at several spatial scales (reviewed by Díaz et al. 2013). Our study reports for the first time the diversity of these four key taxonomic groups measured through a systematic survey. Our approach allows us to compare different habitat types as well as to examine the relative contribution of different habitats to the landscape-scale biodiversity.
The importance of wood pastures
Wood pastures, in both natural and cultural landscapes, are still common worldwide. For these systems, scattered trees have been identified as ‘keystone structures’ due to the disproportionate ecological values and ecosystem services, including biodiversity, they provide relative to the small area they occupy in landscapes (Mills et al. 1993; Manning et al. 2006). Trees are essential sources of food and shelter, and they generate multiple resource gradients in part associated to the differential use by livestock (Moreno et al. 2013). Fine-grained ecotones created by scattered trees (from beneath the canopy to the interstial open pastures) are key for high species and niche densities of wood pastures (Bergmeier et al. 2010). Indeed, they have a disproportionate value for different taxa, as reported by Fischer et al. (2010) for birds and bats in an Australian livestock grazing landscape. These authors found that, compared to treeless sites, bird richness doubled with the presence of one tree and when 3–5 trees were present, bat richness tripled and bat activity increased by a factor of 100. Bat species richness reaches a nearly asymptotic value at roughly 5 trees/ha, while bird species richness keeps increasing slightly even above 100 trees/ha. Other studies done in Iberian dehesas have reported that plant and bird species richness are similar or higher in open woodlands than in adjacent dense forest and/or shrublands (Diaz et al. 2013).
Here, we also found more species of vascular plants, bees, spiders and earthworms in wood pastures as a whole than in open pastures (differences not significant for bees). Differences were mostly confirmed when extrapolation and rarefaction approaches were used (Figs. 2, 3). Bermeier et al. (2010) reported similar results for vascular plants, birds, snails and beetles for other European wood pastures. An increase in invertebrate richness and abundance has been reported when moving from open grassland to agroforestry conditions for carabid beetles in Northern Ireland (Cuthbertson and McAdam 1996) and for spiders and staphylinid beetles in Scotland (Dennis et al. 1996). Burgess (1999) also reports on the benefits of silvoarable systems, relative to common monocultures, in terms of the number of birds and mammals. Gillet et al. (1999) determined a plant species richness optimum at 30 % of tree cover for Swiss Jura Mountains wood pastures.
Contrary to our expectations, at plot scale we did not find higher species richness in wood pastures than in adjacent open pastures for any of the four biological groups studied. Furthermore, abundance and species richness of spiders and earthworms were negatively affected by tree cover, and were significantly higher in open pasture plots than in wood pasture plots. However, the similarity among plots, either measured or estimated (Table 3), was higher in the open than in wood pastures for vascular plants, spiders and earthworms (not for bees). Thus, the higher overall species richness found here for wood pastures is explained by the higher spatial heterogeneity (β diversity) of wood pastures. Comparing species richness at plot and habitat levels, we found that heterogeneity (Shabitat − Splot) was higher for earthworms, followed by spiders, bees and plants. This is consistent with the sedentary behavior and high isolation of populations of soil-living animals (Costa et al. 2013). Azcárate et al. (2002) showed that the presence of many species in Mediterranean grasslands is determined by dispersal (production of numerous small seeds) rather than by competitive ability, which could explain the lower importance of spatial heterogeneity for plant species richness compared to other taxa here studied.
The exception found for bees could be tentatively explained by the relative increase of anemophilous grasses (pollen dispersed by wind) and the concomitant loss of legumes and forbs (mostly entomophilous with pollen dispersed by insects) in the neighborhood of trees (Marañon et al. 2009) caused by the positive effect of trees on soil nitrogen availability (Gallardo et al. 2000; Moreno and Obrador-Olán 2007). Both abundance and species richness of bees were higher in pastures dominated by annual plant species (rich in forbs and legumes) than in pastures dominated by perennial species (rich in grasses).
The conservation of the habitat mosaic
Biological diversity is unevenly distributed across space (Gaston 2000). Patterns of biodiversity varies across spatial scales and between taxonomic groups (Tews et al. 2004; Crawley and Harral 2001; Turtureanu et al. 2014). In our study, species richness and abundance differed significantly across defined habitat types, and the richest habitat depended on the biological group and scale. Plant richness was highest in wood pastures and shrubs, both at plot and landscape scales, reflecting the benefit of the fine-scale gradients created by woody plants for understory pasture (Marañon et al. 2009; Moreno et al. 2013), but also the high β-diversity (low Sorensen index) for most of the GHC studied. Although, high spatial heterogeneity is a common feature of many other European semi-natural dry grasslands (Turtureanu et al. 2014), in our study area open pastures showed a lower spatial heterogeneity (higher similarity among plots) than other GHCs.
Earthworm diversity did not differ among GHC at the plot scale, but were more abundant in open pastures and near water bodies than in woody habitats, a fact probably explained by the lower moisture content usually found underneath trees and shrubs in Iberian dehesas (Cubera and Moreno 2007a, b; Rolo et al. 2013). By contrast, at the landscape scale, earthworm diversity was higher in woody habitats and agricultural crops, reflecting the higher spatial heterogeneity of earthworm community in woody habitats compared to open pastures and wet areas, as reported for cold-humid mixed deciduous forest in Canada (Whalen 2004) and for sub-humid tropical gallery forests in Colombia (Jiménez et al. 2011). Earthworms were more diverse and abundant beneath tree/shrub canopies than in open areas in dry-warm rangelands of Israel, but abundance and diversity did not differ between shady and sunny microhabitats in cooler sites (Pavlíček et al. 1996). Maestre and Cortina (2002) found similar canopy effects of Stipa tenacissima tussocks in the rangelands of semiarid Southeastern Spain. Hence, it seems that the role of scattered trees or other plants would depend on soil-climate conditions, although more studies are needed to unravel the processes affecting the specific distribution of earthworms in dry wood pastures.
Spiders were also more abundant in herbaceous lines and open pastures. However, at plot scale the richest GHC were shrubs and pastures dominated by perennial species, with woody habitats showing again the lowest similarity among plots and the richest GHCs at landscape scale. Also Miyashita et al. (2012) reported that richness and abundances of major constituent spider species were highest in intermediate mixtures of forests and paddy fields, and that this effect derives from multi-scale landscape heterogeneity.
Bees were more abundant and diverse in annual pastures and wood lines when counted at plot scale. At a landscape scale the most diverse GHCs were again wood lines with wood pastures and shrubs. Although bees, especially domestic bees, were very abundant in annual pastures in early spring, in summer as pastures dried, bees become dependent on flowering shrubs (e.g. Rubus spp., Thymus spp., and Lavandula spp.) and honeydew-secreting oaks. While in early spring domestic bees are the most abundant, by late spring and summer the less abundant solitary bees became the dominants (data not shown). Our results reflect the importance of habitat mosaic and landscape heterogeneity for bees, as 39 of a total of 60 species were found only in one habitat type. Pollinators are considered as a crucial component in ecosystems and the importance of loss of diversity of this functional group, with negative ecological and economical effects, is widely documented (Potts et al. 2010 for a review).
Iberian dehesas have been created and maintained by a combination of different land uses commonly practiced within ranches (Moreno and Pulido 2009). This combination usually entails a rotational management scheme of crops, fallows, pastures and shrublands leading to a mosaic of habitats that favors the diversity of plant and animal species (Benton et al. 2003; Concepción et al. 2012). The complementary use of different dehesa habitats has been reported for different vertebrate species, such as red deer (Cervus elaphus L.), small and medium mammals (e.g. rabbits, hares) and birds (e.g. passerines, woodpigeon, cranes, vultures) (see Díaz et al. 2013 for a review). In the dehesas studied here, spatial heterogeneity is revealed by (i) the differences among neighbor farms (significant Farm factor in GLMMs; Table 1), (ii) the low species affinity among plots within GHCs (Table 3), and (iii) the low species affinity among GHCs (Table 4). The high proportion of species unique to GHCs type in this study reinforces the importance of the spatial heterogeneity of dehesa system for species richness of the four biological groups studied. The ‘habitat heterogeneity hypothesis’ assumes that structurally complex habitats increase species diversity as they provide more niches and diverse ways of exploiting environmental resources (Tews et al. 2004). In most habitats, plant communities determine the physical structure of the environment, and therefore, have a considerable influence on the distributions and interactions of animal species. In the dehesas studied, woody pastures and other woody habitats harbored the highest plant richness and this was associated to higher richness of the three animal groups studied at landscape scale.
Over the last decades most Mediterranean silvopastoral systems have been following two divergent trends: intensification of land use in some areas, and land abandonment and shrub encroaching in others (Pinto-Correia 2000; Papanastasis 2004). In both cases the diversity of land uses and associated habitats are notably reduced (Plieninger and Wilbrand 2001). Our results suggest that these trends could have important detrimental effects for regional biodiversity as the loss of certain habitats would result in the loss of species. Successive measures granted by the European Common Agricultural Policy (CAP) did not halt the progressive reduction of the spatial heterogeneity of most of the European pastoral woodlands and wooded pastures (Bergmeier et al. 2010). According to Concepción et al. (2012), landscape-scale management options should take priority over local agri-environmental measures for biodiversity conservation, and they should include the maintenance of a diversity of farming activities in extensive rangelands so as to guarantee the conservation of habitat diversity. Specifically, the mix of wood pastures with open pastures at different spatial scales seems essential for the conservation of dehesa biodiversity.
The disproportionate importance of marginal habitats
Besides the diversity of main (productive/managed) fields, dehesas commonly contain different landscape elements, such as hedges, woodlots, artificial ponds, roadsides or boundary strips interspersed among the main paddocks. Tews et al. (2004) denoted these distinct spatial elements, which provide resources and/or shelter for animal and plant species, as keystone structures (distinct spatial structures providing resources, shelter or ‘goods and services’ crucial for other species). The importance of these keystone structures for farm biodiversity has been highlighted in the last decade. In our study, although both wood pastures and open pastures accounted for a high number of species, the presence of some marginal habitats (unmanaged habitats that occupy a low proportion of farm surface), including linear features, played a very significant role in terms of species richness at the farm and landscape level. These keystone structures or ecological infrastructures (sensu Boller et al. 2004) occupied a very small area of the farms studied (<10 %) and harbored a high proportion of the species found (Fig. 3). The proportion of species that were found only in those keystone elements reached up to ca. 30 % for spider species, 40 % for bee species and 45 % for earthworm species (Fig. 3).
Marginal habitats are expected to increase species richness by supporting species not found in open and wood pastures (Díaz et al. 2103), although positive effects would be somewhat weaker than in intensive agricultural landscapes devoid of natural vegetation (Batáry et al. 2011). Tscharntke et al. (2005) stated that, in simple landscapes under more intensive agriculture, the conservation of marginal habitats is more important than in complex landscapes. However, we found that even in extensive, seminatural agroecosystems, such as Iberian dehesas, the presence of different habitats, and more notably the presence of marginal habitats, also play an important role for landscape biodiversity.
In many regions, keystone structures are being seriously affected by changes in farming practices. Concentrating conservation efforts on these non-productive elements interspaced in intensively used fields could stabilize species diversity in these ecosystems at a high level while having a minimum impact on agricultural land use (Berger et al. 2003; Tews et al. 2004). Our results support this principle and the recent European Common Agricultural Policy of promoting Ecological Focus Areas that aims to devote 7 % of the land within each farm to nature conservation (EU Regulation 1311/2013). Some of these marginal, habitats, such as small woodlots for livestock sheltering and shrub patches, can play a positive role on the management and persistence of extensive silvopastoral systems due to its significance for tree regeneration (Ramírez and Díaz 2008; Pulido et al. 2010; Díaz et al. 2013).
Extensive low-input farming, biodiversity and food production
The ecosystem services provided by biodiversity in relation to human activity is a central issue in conservation science (Winfree and Kremen 2009; Petz and van Oudenhoven 2012).The flow of these services depends on the management of the agricultural ecosystem and on the species that provide more positive effects for crop production (e.g., soil fertility, pollination) and those that reduce negative ones (e.g., crops pest) (Zhang et al. 2007). Farm types and farm landscapes valuable for biodiversity are more likely among extensive low-input farming systems with the presence of non-productive or marginal habitats that mimic natural habitats, as it is the case of the Iberian dehesas here studied. However, as world food demand is expected to more than double by 2050, the low productivity of extensive farming systems is a major threat to their existence. The continuous loss of bees in many regions, presumably caused by land intensification and the use of agrochemicals, is a good example (Potts et al. 2010). Decisions about how to meet the challenge of world food demand will have profound effects on wild species and habitats. Two competing solutions have been proposed: wildlife-friendly farming or land-sharing, which boosts densities of wild populations on farmland but may decrease agricultural yields, and land-sparing, which minimizes demand for farmland by increasing yield (Green et al. 2005). While ecological intensification, sensu Doré et al. (2011), of extensive low-productive farming could become a need to increase food production while maintaining ecological services of farms, according to our results the maintenance of a land use mosaic and diverse interspaced non-productive keystone-structures would probably be still needed to ensure biodiversity conservation.
Conclusions
While pastoral landscapes have been mostly deforested over the centuries (Bergmeier et al. 2010) the conservation of trees in a pasture matrix is still common in some Mediterranean silvopastoral systems such as in Iberian dehesas. Trees provide multiple woody and non-woody plant products, high-quality food, livestock and game products, recreational or cultural services through multiple activities conducted with a comparatively low environmental impact (Moreno and Pulido 2009). Trees also provide important ecosystem services such as carbon sequestration, soil fertilization and control against erosion, microclimate amelioration, and shelter for livestock. Here we have shown that trees also contribute positively to the diversity of four taxonomic groups within an agricultural landscape (see also Díaz et al. 2013 for a revision).
Although high biodiversity values found in Iberian dehesas can be partly explained by the existence of a habitat dominated by scattered trees, the intimate mix of tree and treeless pastures has also a significant role. While at landscape scale the diversity of the four biological groups studied was higher in wood pastures and other woody habitats, at plot scale they were more abundant and/or biodiverse at open pastures. The low proportion of shared species among habitats and among plots within each habitat type, and the high proportion of species found in unique plots or habitats indicated that every habitat contributes to farm biodiversity. Marginal land uses and linear features, which occupy a low proportion of the farm area, harbored a good number of species that were not found in the main field of dehesas studied. These results support policy measures implemented in many European countries for the maintenance of farm keystone structures and reveal that these measures should not be applied exclusively in more intensive farming systems. Nevertheless, more studies are still needed to clarify how important keystone structures are for biodiversity under different farming intensities, from intensive monocultures to extensive silvopastoral systems. More studies are also needed to unveil how the spatial distribution of those keystone structures affects the biodiversity of main fields and that of the whole farm and landscape.
References
Azcárate FM, Sánchez AM, Arqueros L, Peco B (2002) Abundance and habitat segregation in Mediterranean grassland species: the importance of seed weight. J Veg Sci 13:159–166
Batáry P, Báldi A, Kleijn D, Tscharntke T (2011) Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis. Proc R Soc B Biol Sci 278:1894–1902
Bengtsson J, Ahnström J, Weibull A-C (2005) The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol 42:261–269
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Berger G, Pfeffer H, Kächele H, Andreas S, Hoffmann J (2003) Nature protection in agricultural landscapes by setting aside unproductive areas and ecotones within arable fields (‘Infield Nature Protection Spots’). J Nat Conserv 11:221–233
Bergmeier E, Petermann J, Schröder E (2010) Geobotanical survey of wood-pasture habitats in Europe: diversity, threats and conservation. Biodivers Conserv 19:2995–3014
Blondel J, Aronson J, Boudiou JY, Bœuf G (2010) The Mediterranean Basin—biological diversity in space and time. Oxford University Press, Oxford
Boller EF, Häni F, Poehling H-M (2004) Ecological infrastructures: ideabook on functional biodiversity at the farm level. Agridea, Lindau
Bugalho MN, Caldeira MC, Pereira JS, Aronson JA, Pausas J (2011) Mediterranean oak savannas requirehuman use to sustain biodiversity and ecosystem services. Front Ecol Environ 5:278–286
Bunce RGH, Bogers MMB, Roche P, Walczak M, Geijzendorffer IR, Jongman RHG (2011) Manual for habitat and vegetation surveillance and monitoring. Alterra report 2154. Alterra, Wageningen
Burgess PJ (1999) Effects of agroforestry on farm biodiversity in the UK. Scott For 53:24–27
Calama R, Tomé M, Sánchez-González M, Miina J, Spanos K, Palahi M (2010) Modelling non-wood forest products in Europe: a review. For Syst 19:69–85
Campos P, Huntsinger L, Oviedo JL, Starrs PF, Díaz M, Standiford RB, Montero G (eds) (2013) Mediterranean Oak Woodland Working Landscapes. Dehesas of Spain and Ranchlands of California, vol 16., Landscape SeriesSpringer, New York
Carrete M, Donázar JA (2005) Application of central-place foraging theory shows the importance of Mediterranean dehesas for the conservation of the cinereous vulture, Aegypius monachus. Biol Conserv 126:582–590
Chao A (2005) Species richness estimation. In: Balakrishnan N, Read CB, Vidakovic B (eds) Encyclopedia of statistical sciences. Wiley, New York, pp 7909–7916
Chao A, Chazdon RL, Colwell RK, Shen T-J (2005) A new statistical approach for assessing compositional similarity based on incidence and abundance data. Ecol Lett 8:148–159
Coleman BD, Mares MA, Willig MR, Hsieh Y-H (1982) Randomness, area, and species richness. Ecology 63:1121–1133
Colwell RK (2013) EstimateS: statistical estimation of species richness and shared species from samples. Version 9. User’s Guide and application published. http://www.purl.oclc.org/estimates. Accessed 2 Jan 2014
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philos Trans R Soc (Ser B) 345:101–118
Colwell RK, Chao A, Gotelli NJ, Lin S-Y, Mao CX, Chazdon RL, Longino JT (2012) Models and estimators linking individual-based and sample-based rarefaction, extrapolation, and comparison of assemblages. J Plant Ecol 5:3–21
Concepción ED, Díaz M, Baquero RA (2008) Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landscape Ecol 23:135–148
Concepción ED, Díaz M, Kleijn D, Báldi A, Batáry P, Clough Y, Gabriel D, Herzog F, Holzschuh A, Knop E, Marshall JP, Tscharntke T, Verhulst J (2012) Interactive effects of landscape context constrain the effectiveness of local agri-environmental management. J Appl Ecol 49:695–705
Costa D, Timmermans MJTN, Sousa JP, Ribeiro R, Roelofs D, Van Straalen NM (2013) Genetic structure of soil invertebrate populations: collembolans, earthworms and isopods. Appl Soil Ecol 68:61–66
Crawley MJ, Harral JE (2001) Scale dependence in plant biodiversity. Science 291:864–868
Cubera E, Moreno G (2007a) Effect of single Quercus ilex trees upon spatial and seasonal changes in soil water content in Dehesas of central western Spain. Ann For Sci 64:355–364
Cubera E, Moreno G (2007b) Effect of Land use on soil water dynamic in dehesas of central-western Spain. Catena 71:298–308
Cuthbertson A, McAdam J (1996) The effect of tree density and species on carabid beetles in a range of pasture-tree agroforestry systems on a lowland site. Agrofor Forum 7:17–20
Dennis P, Shellard LJF, Agnew RDM (1996) Shifts in arthropod species assemblages in relation to silvopastoral establishment in upland pastures. Agrofor Forum 7:14–17
Dennis P, Bogers MMB, Bunce RGH, Herzog F, Jeanneret P (2012) Biodiversity in organic and low-input farming systems. Handbook for recording key indicators. Alterra report 2308. Alterra, Wageningen
Díaz M, Campos P, Pulido FJ (1997) The Spanish dehesas: a diversity of land use and wildlife. In: Pain D, Pienkowski M (eds) Farming and birds in Europe: the common agricultural policy and its implications for bird conservation. Academic Press, London
Díaz M, Tietje MD, Barrett RH (2013) Effects of management on biological diversity and endangered species. In: Campos P, Huntsinger L, Oviedo JL, Starrs PF, Díaz M, Standiford RB, Montero G (eds) Mediterranean oak woodland working landscapes. Dehesas of Spain and ranchlands of California. series: landscape series, vol 16. Springer, New York
Doré T, Makowski D, Malézieux E, Munier-Jolain N, Tchamitchian M, Tittonell P (2011) Facing up to the paradigm of ecological intensification in agronomy: revisiting methods, concepts and knowledge. Eur J Agron 34:197–210
EC (2005) Agri-environment Measures. Overview on general principles, types of measures, and application. European Commission, Directorate General for Agriculture and Rural Development. Unit G-4—evaluation of measures applied to agriculture, studies http://www.ec.europa.eu/agriculture/publi/reports/agrienv/rep_en.pdf. Accessed 01 Nov 2014
EEA (2010) Corine land cover 2006 inventory. On-line dataset, version 13 (02/2010)—coordination of information on the environment, European Environment Agency. http://www.eea.europa.eu/data-and-maps/data/corine-land-cover-2006-raster. Accessed March 2010
EEC (1992) Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. http://www.ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm. Accessed 02 Nov 2014
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin J-L (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14:101–112
Fischer J, Stott J, Law BS (2010) The disproportionate value of paddock trees. Biol Conserv 143:1564–1567
Flohre A, Fischer C, Aavik T, Bengtsson J, Berendse F, Bommarco R, Ceryngier P, Clement LW, Dennis C, Eggers S, Emmerson M, Geiger F, Guerrero I, Hawro V, Inchausti P, Liira J, Morales MB, Oñate JJ, Pärt T, Weisser WW, Winqvist C, Thies C, Tscharntke T (2011) Agricultural intensification and biodiversity partitioning in European landscapes comparing plants, carabids, and birds. Ecol Appl 21:1772–1781
Gabriel D, Sait SM, Kunin WE, Benton TG (2013) Food production vs. biodiversity: comparing organic and conventional agriculture. J Appl Ecol 50:355–364
Gallardo A, Rodríguez-Saucedo JJ, Covelo F, Fernández-Alés R (2000) Soil nitrogen heterogeneity in a dehesa ecosystem. Plant Soil 222:71–82
Gaston KJ (2000) Global patterns in biodiversity. Science 405:220–227
Gibbons P, Lindenmayer DB, Fisher J, Manning AD, Weinberg A, Seddon J, Ryan P, Barrett G (2008) The future of scattered trees in agricultural landscapes. Conserv Biol 22:1309–1319
Gillet F, Murisier B, Buttler A, Gallandat J-D, Gobat J-M (1999) Influence of tree cover on the diversity of herbaceous communities in subalpine wooded pastures. Appl Veg Sci 2:47–54
Green RE, Cornell SJ, Scharlemann JPW, Balmford A (2005) Farming and the fate of wild nature. Science 307:550–555
Herzog F, Jeanneret P, Ammari Y, Angelova S, Arndorfer M, Bailey D, Balázs K, Báldi A, Bogers M, Bunce RGH, Choisis J-P, Cuming D, Dennis P, Dyman T, Eiter S, Elek Z, Falusi E, Fjellstad W, Frank T, Friedel JK, Garchi S, Geijzendorffer IR, Gomiero T, Jerkovich G, Jongman RHG, Kainz M, Kakudidi E, Kelemen E, Kölliker R, Kwikiriza N, Kovács-Hostyánszki A, Last L, Lüscher G, Moreno G, Nkwiine C, Opio J, Oschatz M-L, Paoletti MG, Penksza K, Pointereau P, Riedel S, Sarthou J-P, Schneider MK, Siebrecht N, Sommaggio D, Stoyanova S, Szerencsits E, Szalkovski O, Targetti S, Viaggi D, Wilkes-Allemann J, Wolfrum S, Yashchenko S, Zanetti T (2013) Measuring farmland biodiversity. Solutions 4:52–58
Hole DG, Perkins AJ, Wilson JD, Alexander IH, Grice PV, Evans AD (2005) Does organic farming benefit biodiversity? Biol Conserv 122:113–130
Jiménez JJ, Decaëns T, Amezquita E, Rao I, Thomas RJ, Lavelle P (2011) Short range spatial variability of soil physico-chemical variables related to earthworm clustering in a neotropical gallery forest. Soil Biol Biochem 43:1071–1080
Jose S (2012) Agroforestry for conserving and enhancing biodiversity. Agrofor Syst 85:1–8
Kleijn D, Baldi A (2005) Effects of set-aside land on farmland biodiversity: comments on Van Buskirk and Willi. Conserv Biol 19:963–966
Kleijn D, Sutherland WJ (2003) How effective are European agri-environment schemes in conserving and promoting biodiversity? J Appl Ecol 40:947–969
Kleijn D, Kohler F, Báldi A, Batáry P, Concepción ED, Clough Y, Díaz M, Gabriel D, Holzschuh A, Knop E, Kovács A, Marshall EJP, Tscharntke T, Verhulst J (2008) On the relationship between farmland biodiversity and land-use intensity in Europe. Proc R Soc B 276:903–909
Kleijn D, Rundlöf M, Scheper J, Smith HG, Tscharntke T (2011) Does Conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol 26:474–481
Kristensen P (2003) EEA core set of indicator. Revised version April, 2003. European Environment Agency, Copenhagen
Maestre F, Cortina J (2002) Spatial patterns of surface soil properties and vegetation in a Mediterranean semi-arid steppe. Plant Soil 241:279–291
Manning AD, Fischer J, Lindenmayer DB (2006) Scattered trees are keystone structures—implications for conservation. Biol Conserv 132:311–321
Marañón T (1986) Plant species richness and canopy effect in the savanna-like “dehesa” of S.W. Spain. Ecol Mediterr 12:131–141
Marañón T, Pugnaire FI, Callaway RM (2009) Mediterranean-climate oak savannas: the interplay between abiotic environment and species interactions. Web Ecol 9:30–43
Martín J, López P (2002) The effect of Mediterranean dehesa management on lizard distribution and conservation. Biol Conserv 108:213–219
Mills LS, Soule ME, Doak DF (1993) The keystone species concept in ecology and conservation. Bioscience 43:219–224
Miyashita T, Chishiki Y, Takagi SR (2012) Landscape heterogeneity at multiple spatial scales enhances spider species richness in an agricultural landscape. Popul Ecol 54:573–581
Moreno G, Obrador-Olán JJ (2007) Effects of trees and understorey management on soil fertility and nutritional status of holm oaks in Spanish dehesas. Nutr Cycl Agroecosyst 78:253–264
Moreno G, Pulido FJ (2009) The functioning, management, and persistente of dehesas. In: Riguero-Rodriguez A, Mosquera-Losada MR, McAdam J (eds) Agroforestry systems in Europe. Current status and future prospects. Advances in agroforestry series. Springer, New York, pp 127–161
Moreno G, Bartolome JW, Gea-Izquierdo G, Cañellas I (2013) Overstory-understory relationships. In: Campos P, Huntsinger L, Oviedo JL, Starrs PF, Díaz M, Standiford RB, Montero G (eds) Mediterranean oak woodland working landscapes. Dehesas of Spain and ranchlands of California, vol 16., Series: landscape seriesSpringer, New York, pp 145–180
Myers et al (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Opermmann R, Beaufoy G, Jones G (2012) High nature value farming in Europe. 35 European countries—experiences and perspectives. Verlag regionanlkultur, Germany
Papanastasis VP (2004) Vegetation degradation and land use changes in agrosilvopastoral systems. In: Schnabel S, Ferreira A (eds) Sustainability of agrosilvopastoral systems. Advances in GeoEcology, vol 37. Catena Verlag, Reiskirchen, pp 1–12
Paracchini ML, Petersen J-E, Hoogeveen Y, Bamps C, Burfield I, van Swaay C (2008) High nature value farmland in Europe—an estimate of the distribution patterns on the basis of land cover and biodiversity data. Report EUR 23480, European Environmental Agency, ISPRA, Italy
Pavlíček T, Csuzdi C, Smooha G, Beiles A, Nevo E (1996) Biodiversity and microhabitat distribution of earthworms at “Evolution Canyon”, a Mediterranean microsite, Mount Carmel, Israel. Isr J Zool 42:449–454
Petz K, van Oudenhoven APE (2012) Modelling land management effect on ecosystem functions and services: a study in the Netherlands. Int J Biodivers Sci Ecosyst Serv Manag 8:135–155
Pinto-Correia T (2000) Future Development in Portuguese Rural Areas: how to manage agricultural support for landscape conservation? Landsc Urb Plan 50:95–106
Plieninger T, Wilbrand C (2001) Land use, biodiversity conservation, and rural development in the dehesas of Cuatro Lugares, Spain. Agrofor Syst 51:23–34
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25:345–353
Primdahl J, Peco B, Schramek J, Andersen E, Onate JJ (2003) Environmental effects of agri-environmental schemes in Western Europe. J Environ Manag 67:129–138
Pulido F, García E, Obrador JJ, Moreno G (2010) Multiple pathways for tree regeneration in anthropogenic savannas: incorporating biotic and abiotic drivers into management schemes. J Appl Ecol 47:1272–1281
Ramírez JA, Díaz M (2008) The role of temporal shrub encroachment for the maintenance of Spanish holm oak Quercus ilex dehesas. For Ecol Manage 255:1976–1983
Rolo V, Plieninger T, Moreno G (2013) Facilitation of holm oak recruitment through two contrasted shrubs species in Mediterranean grazed woodlands: patterns and processes. J Veg Sci 24:344–355
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79–92
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol Lett 8:857–874
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist TO, Dormann CF, Ewers RW, Fründ J, Holt RD, Holzschuh A, Klein AM, Kleijn D, Kremen C, Landis DA, Laurance W, Lindenmayer D, Scherber C, Sodhi N, Steffan-Dewenter I, Thies C, van der Putten WH, Westphal C (2012) Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biol Rev 87:661–685
Turtureanu PD, Palpurina S, Becker T, Dolnik C, Ruprecht E, Sutcliffe LME, Szabó A, Dengler J (2014) Scale- and taxon-dependent biodiversity patterns of dry grassland vegetation in Transylvania agriculture. Ecosyst Environ 182:15–51
Whalen JK (2004) Spatial and temporal distribution of earthworm patches in corn field, hayfield and forest systems of southwestern Quebec, Canada. Appl Soil Ecol 27:143–151
Whittingham MJ (2007) Will agri-environment schemes deliver substantial biodiversity gain, and if not why not? J Appl Ecol 44:1–5
Winfree R, Kremen C (2009) Are ecosystem services stabilized by differences among species? A test using crop pollination Proc. R Soc B 276:229–237
Zhang W, Ricketts TH, Kremen C, Carney K, Swinton SM (2007) Ecosystem services and dis-services to agriculture. Ecol Econ 64:253–260
Zomer RJ, Trabucco A, Coe R, Place F (2009) Trees on farm: analysis of global extent and geographical patterns of agroforestry. ICRAF Working Paper no. 89. Nairobi, Kenya
Acknowledgments
We are grateful to all farmers who allowed access to their fields and provided information on land use and management. This work was funded by the European Union through the FP7 project BioBio (Indicators for biodiversity in organic and low-input farming systems; www.biobio-indicators.org), and is a contribution by MD to the projects Consolider Montes (CSD2008-00040), VULGLO (CGL2010-C03-03), REMEDINAL3-CM (S2013/MAE-2719) and BACCARA (CE: FP7-226299).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Moreno, G., Gonzalez-Bornay, G., Pulido, F. et al. Exploring the causes of high biodiversity of Iberian dehesas: the importance of wood pastures and marginal habitats. Agroforest Syst 90, 87–105 (2016). https://doi.org/10.1007/s10457-015-9817-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-015-9817-7