
Abstract
Intercropping Persian walnut (Juglans regia L.) and wheat (Triticum aestivum Linn.) in northwest China is receiving increasing attention because it offers potential advantages for improving land-use efficiency, increasing economic returns to farmers and increased sustainability in crop production. We carried out a two-year field intercropping experiment, using a system composed of young Persian walnut trees with wheat to determine the root competitive interactions between the two species, aboveground biomass and the economic yield of each component of the intercropping system at a spacing of 4 × 6 m in Hetian old oasis, south Xinjiang province, northwest China. Roots were sampled by auger in the Persian walnut tree/wheat intercropping system and in the sole wheat and sole walnut systems, down to a depth of 1 m in the soil profile. The results show that the roots of the trees spread under the crop and that the roots of intercropped wheat tended to have a shallower distribution in the soil profile. The roots of both intercropped walnut and wheat had a lower root length density and root diameter and a higher specific root length at all soil depths than those of sole walnut trees and wheat in 2012. Although root competition belowground led to decreases in yield and biomass of both crops, intercropping of Persian walnut and wheat improved the utilization of plant growth resources (LER > 1) as compared with sole crops. Tree height, diameter at breast height and mean radius of crown of intercropped trees in both intercropping years had lower values when compared with the monoculture. We conclude that interspecific interactions occurred largely as below-ground competition between the species investigated. Competition for soil resources was the primary limiting factor for wheat productivity. Further research is needed to examine the dynamic processes of species interactions in intercropping systems with trees of increasing age.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Agroforestry with fruit trees/food crops which combine an economically valuable tree species with an agricultural crop on the same land, is commonly practiced in many parts of the world such as India (Jamaludheen et al. 1997), southeast Asia (Nissen et al. 1999), north America (Fletcher III et al. 2012), and Africa (Imo 2009; van Asten et al. 2011). Maximizing the use of available resources (water, land, nutrients, light and temperature) (positive interactions) while minimizing resource competition (negative interactions) between trees and crops is the key to increasing yields and overall productivity in agroforestry systems (Ong et al. 1996; Cannell et al. 1996; Zamora et al. 2007). Many studies have shown that productivity of many tree-crop systems is limited by interspecific underground competition for resources (Singh et al. 1989; Schroth 1999; Jose et al. 2000a, 2001; Zamora et al. 2007; Zhang et al. 2013). This has made research on root interaction a major component of alley cropping studies in recent decades. Interest in diversifying farm income and reducing environmental impacts of agriculture has led to the expansion of alley cropping systems (Jose and Gillespie 1998a). Jose et al. (2004, 2006) reported that root spatial distribution of the component species plays an important role in belowground resource competition. Thus, knowledge of the spatial distribution of agroforestry root systems is necessary to gauge the degree of competitive resource sharing among system components (Gregory 1996; Van Noordwijk and Purnomosidhi 1995).
Persian walnut tree (Juglans regia L.)/wheat (Triticum aestivum L.) mixtures play an important role in agricultural production in Hetian old oasis, south Xinjiang Uygur Autonomous Region, northwest China (Zhang et al. 2010) because of the good quality of walnut wood for furniture (Ozkan et al. 2005) and also because of its ecological value as a windbreak in windy and sandy areas (Greb et al. 1961). The nuts of the tree are an edible and nutritious human food (Ozkan 2009). Specifically, the Persian walnut tree is the main cash crop and wheat is a primary food crop and both are harvested once a year in south Xinjiang. By 2011 there were 280 thousand hectares of Persian walnut in Xinjiang Uygur Autonomous Region which accounted for 14.2 % of total production of walnut in China (Li et al. 2014). Persian walnut/crop intercropping has already become one of the main agroforestry systems for walnut production, especially in Hetian old oasis (Qiao et al. 2012). Although young Persian walnut trees produce no nuts at early life stages, certain returns, i.e., the yield and biomass of intercropped wheat derived from intercropping systems can also cover a number of losses for local farmers to some degree. In the prevalent intercropping system Persian walnut trees are grown simultaneously with wheat for about 270 days. When the two plant species grow together, interspecific interactions between them inevitably occur. Although Persian walnut trees are known to produce allelochemicals that inhibit the growth of other plants (Willis 2000), the trees we studied do not cause a problem because allelopathy of walnut only manifests itself about 10 years after establishment (Scott and Sullivan 2007). Therefore, only the degree to which interspecific competition influences root development and distribution is of interest in this study.
Previous studies have shown that interspecific competition in agroforestry systems may reduce root length density and aboveground biomass. For example, Zhang et al. (2013) observed that both tree and crop root length was reduced in three jujube tree (Zizyphus jujuba Mill.)/wheat agroforestry systems in this region. Similarly, Livesley et al. (2000) found that the root length of plants-both trees (grevillea (Grevillea robusta) and senna (Senna spectabilis)) and an arable crop (maize) decreased to some extent. Jose et al. (2000b) found that maize yields were reduced by 35 and 33 % when intercropped with black walnut and red oak (Quercus spp.), respectively. Rao et al. (1991) reported that roots in the upper soil profile caused intense belowground competition for resources and resulted in decreased crop productivity. In a Leucaena/maize intercropping system, the mean height and mean biomass of maize were reduced by 43 and 75 %, respectively (Chamshama et al. 1998). In studies of walnut/crop root systems, Pham et al. (1977) suggested that intermediate-aged black walnut is deep-rooted; almost 80 % of its root system occupies the upper 61 cm of the soil. In a review on extension of tree roots, Stone and Kalisz (1991) reported that vertical and radial root extension of 15-year-old black walnut trees were 3.3 and 4.5 m, respectively.
Although many of the competitive species in tree/crop mixtures have been identified, not all have been adequately quantified and the root distribution in the walnut-wheat system remains to be studied. In the present study we experimentally compared walnut trees grown with wheat with sole walnut in terms of above-ground biomass and grain yield and below-ground root distributions to examine the competitive interactions between the two crop species and the likely response mechanisms in order to provide optimum management strategies and technologies for managing the intercropping system with high yield and high resource use efficiency. Walnut trees and wheat were selected for the research because of their importance as the main economic and food crops, respectively, in Hetian old oasis.
The main objectives of the present study were therefore to examine the advantage of intercropping and effects of mixed species systems on the yield and biomass of the components species, to explore the relative competitive ability of the crop components in the system, and to determine the root distribution and competitive interactions between walnut trees and associated wheat.
Materials and methods
Experimental site
The study was conducted in 2011 and 2012 at Hetian Institute of Agricultural Sciences, Agricultural Technology Extension and Service Center of Hetian Prefecture, Xinjiang Uygur Autonomous Region, and the experimental site (73°37′N, 34°20′E) is located 6 km north of Hetian City at an elevation of 1380 m above sea level. Annual mean temperature is 13.7 °C. Cumulative temperatures above 0 and 10 °C are 4,646 and 4,064 °C, respectively. The frost-free period is 200–220 days. Total solar radiation is 6,627 MJ m−2 year−1. Annual precipitation is 37.1 mm, potential evaporation 2,636 mm, and the region has a typical arid climate. In 2011 and 2012 the annual rainfall was 34.2 and 44.8 mm, respectively. The soil at the site is classified as an Arenosol in the Food and Agriculture Organization (FAO) soil classification system. Some physical and chemical properties of the soil have been published previously (Zhang et al. 2013). The walnut trees (Juglans regia L.) were one year old in 2011 and two years old in 2012.
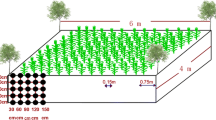
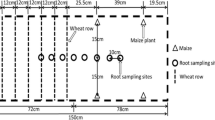
The field experiment was a single-factor design with three replicates in which the treatments were sole wheat, sole Persian walnut trees, and wheat/walnut tree intercropping. The intercropping was designed as an additional series. Wheat and walnut tree intercropping involved planting in 6-m-wide strips (wheat plus two rows of walnut trees), which included a 4.50-m-wide wheat strip (30 rows of wheat with 0.15 m inter-row distance) and 4-m distance between walnut trees within rows. The distance between walnut trees and the nearest wheat row was 0.75 m. Walnut trees thus occupied 40 % of the intercropped area and wheat 60 % (Fig. 1).

Diagrammatic representation of the planting of strips of walnut trees/wheat in intercropped plots in a young walnut/wheat alley cropping system in Xinjiang, northwest China. Distances between rows and gap sizes (cm) are indicated. There was a distance of 30 cm between adjacent root sampling sites
One strip comprising two rows of walnut trees and thirty rows of wheat constituted an intercropping plot, and three rows of walnut trees and sixty rows of wheat comprised a sole walnut tree and wheat plot, respectively. The individual plot area was 12 × 6 m2 for sole walnut trees, sole wheat and wheat/walnut tree intercropping.
All plots were given identical applications of N at 450 kg ha−1 as urea and diammonium phosphate (N), and of phosphorus (P) at 30 kg ha−1 as diammonium phosphate. All the P fertilizer and half of the N were broadcast evenly and incorporated into the top 20 cm of the soil prior to sowing. The remaining half of the N fertilizer was applied at the elongation stage of wheat in the intercropping system, monoculture walnut trees and monoculture wheat.
Winter wheat was sown on October 25 in 2010 and October 16 in 2011. Walnut seedlings were transplanted from a tree nursery into 20-centimeter holes in the experimental field on May 30 in 2011. The stem diameter of the walnut seedlings averaged 2.4 cm, the height averaged 1.42 m and the seedling survival rate was 92.6 % in 2011. Dates of harvesting were June 25 in 2011 and June 15 in 2012 for wheat and October 5 in 2011 and October 2 in 2012 for walnut. Irrigation was carried out on six occasions on March 25, April 14, May 2, May 23, July 25 and September 5 in 2011 and March 20, April 9, April 28, May 20, July 20 and August 30 in 2012. Each irrigation application consisted of 90 mm (900 m3 ha−1). The irrigation practice followed that recommended to farmers by local agronomists. All crops received the same agricultural management practices. The aboveground biomass of walnut trees and grain and straw yields of wheat were determined at maturity of the wheat crop.
The study is part of a larger experiment which includes a randomized complete block design with 24 plots for walnut-wheat alley cropping and 27 plots for jujube tree-wheat alley cropping (Zhang et al. 2013). The walnut tree/wheat intercropping system was established in 2011.
Plant sampling
Grain, biomass and straw yields of wheat were measured at maturity. Grain yield and biomass were determined by harvesting 5 m of each strip in intercropped wheat at maturity. Mortality and stunting were recorded. Stem and canopy dimensions of all experimental trees were measured on October 5, 2011 and October 2, 2012. Trees were then cut at the base to measure biomass. After harvest, plant samples were air-dried and the grain was threshed by hand. Nitrogen, P, and K concentrations in the grain and straw were determined on ground subsamples of oven-dried plant material after digestion in a mixture of concentrated H2SO4 and H2O2. Nitrogen was measured by the micro-Kjeldahl procedure, P by the vanadomolybdate method, and K by flame photometry.
Root sampling
On 12 June in 2011 and 7 June in 2012, when wheat was at its mature stage, an auger sampling method (Böhm 1979) was employed in order to minimize damage to the plots and allow detailed examination of root distribution at later growth stages. Soil cores (5.5 cm diameter × 5.0 cm depth) were collected at 20-cm intervals to a maximum depth of 100 cm to determine the vertical root distribution. To determine the horizontal distribution of walnut tree roots, soil cores were collected from under the center of the walnut tree, and 30 cm from the row for monoculture walnut trees under the center of wheat rows, and 30 cm from the row for monoculture wheat. In the intercropping systems five sampling sites were used as shown in Fig. 1. Soil cores were stored in plastic bags until roots were washed out. Soil moisture contents at different soil depths were determined using auger samples. Plant height, diameter at breast height (DBH), mean radius of crown of walnut trees, wheat yield and total aboveground biomass of all plants were measured and recorded in the field or laboratory. Yield values are reported on a per hectare basis.
Root length density, root diameter and specific root length measurement
All the auger samples (collected on 12 June in 2011 and 7 June in 2012) were weighed. They were immersed in water for at least 1 h. The samples were stirred vigorously and poured through a sieve (0.2 mm2 mesh, 20 cm diameter and 4 cm height). The sieves were suspended in a large water bath and shaken continuously by hand until the roots were washed free of soil. Soil materials remaining on the sieves were removed manually. In Persian walnut tree/wheat intercropping, some auger samples contained the roots of both crop species; therefore, one crop had to be distinguished from the other based on their visual appearance (Zhang et al. 2013). The roots of wheat and Persian walnut tree were distinguished by their different colors, textures and rooting patterns. In the wheat and walnut tree intercropping, for example, the roots of wheat were yellowish and fibrous compared to those of walnut which had smooth surfaces and a brown color and a larger diameter and had a harder texture than the wheat roots. Roots were analyzed for morphological parameters using a WinRHIZO™ (Regent Instruments Inc., Quebec, Canada) image analysis system. Individual root samples were spread out on a clear tray (30 × 48 cm glass frame) that was filled with water and placed on a flatbed scanner. Analysis of the scanned images provided size–class distributions and quantification of the parameters root length, surface area and average root diameter (Zhang et al. 2013). Scanned roots were then oven-dried for 72 h at 70 °C for root biomass and specific root length assessment (Zamora et al. 2007). The separated root fractions were weighed. The data from the auger samples represent the whole population of walnut tree and wheat roots in each soil profile. Results are presented as contour diagrams made by WIN SURFER (v5.01, SURFER MAPPING SYSTEM). Root distribution maps were created where rooting depth and lateral growth of roots could be determined by locating the given contour value in the soil profile.
Land equivalent ratio (LER)
The LER is the ratio of the area under pure stand to the area under intercropping needed to produce an equal amount of yield at the same management level. LER is the most widely accepted index for evaluating the effectiveness of all forms of mixed cropping and has been extended to agroforestry by some researchers (Vandermeer 1989; Rao et al. 1990, 1991; Cao et al. 2012). Moreover, LER indicates the efficiency of intercropping for using the resources of the environment compared with sole cropping (Mead and Willey 1980; Dhima et al. 2007; Zhang et al. 2013). A minimum LER for the alley cropping system should not be below one, which would reflect a positive synergistic or neutral mutual relationship between trees and crops (Gruenewalda et al. 2007). LER is calculated according to:
where Yt and Yw are the biomass values of Persian walnut tree and wheat in pure culture, respectively, and Ytw and Ywt are the biomass values of walnut tree and wheat, respectively, as mixtures. If the ratio is greater than 1.0, intercropping is advantageous, while a value less than 1.0 indicates a disadvantage.
Statistical analysis
All data were submitted to analysis of variance (ANOVA) using SPSS Version 19.0 for Windows and mean values (n = 3) were compared by least significant difference (LSD) at the 5 % level.
Results
Land equivalent ratio (LER), plant growth and aboveground biomass (yields)
The grain yield and aboveground biomass of intercropped wheat were not significantly (P = 0.084 and P = 0.085, respectively) reduced in 2011, and were reduced significantly (P = 0.006 and P < 0.001, respectively) by 7 % in 2012 at wheat maturity compared with monocropped wheat (Table 1). Aboveground biomass of intercropped walnut trees was not altered significantly (P = 0.106) in 2011 and but declined significantly (P = 0.018) by 15 % in 2012 at maturity of walnut tree compared to corresponding sole walnut trees. The results in 2012 show interspecific competition between intercropped wheat and walnut trees in which both crops showed declining aboveground biomass when intercropped and with increasing of tree age, the biomass of both crops and the yield of wheat were reduced significantly (in 2012).
Tree height, diameter at breast height (DBH) and mean radius of the crown of intercropped trees in both intercropping years had lower values when compared with the monoculture (Table 2). There was no significant (P = 0.414, P = 0.070 and P = 0.355, respectively) reduction in intercropped trees in 2011 but they declined significantly (P = 0.031, P = 0.008 and P = 0.03, respectively) by 6, 16 and 13 % in 2012 at maturity (walnut trees) compared with sole cropped trees (Table 2).
Land equivalent ratios (LERs) of Persian walnut/wheat intercropping, based on biomass, were 1.44 in 2011 and 1.45 in 2012 (Table 1).
Spatial distribution of root length density (RLD) of sole-cropped and intercropped wheat
Intercropped wheat had a lower RLD than monocropped wheat in both years (Figs. 2a, b, 3a, b). Specifically, in the wheat/walnut intercropping system in 2012, the RLD of wheat at distances of 30, 60, 90, 120 and 150 cm from the trees were 83, 83, 84, 86 and 87 % of the corresponding RLDs in the sole-cropped wheat in 2012 (Fig. 2a, b), and the differences were significant (P < 0.05). Although the RLDs of wheat in the wheat/walnut intercropping system at the five positions mentioned above from the trees showed a similar downtrend in 2011 (Fig. 2a, b), the differences were not significant (P < 0.05). Although absolute RLDs decreased to some extent, the pattern of horizontal root distribution changed little between the two intercropping systems (Figs. 2b, 3b). The RLD of intercropped wheat was decreased by intercropped walnut at the majority of soil depths, especially in root diameter and specific root length in 2012 (Fig. 3b). For intercropping in 2011, the maximum reduction in RLD of intercropped wheat affected by intercropped walnut was at 0–60 cm from the tree row (Fig. 2b). One step further, the roots of wheat intercropped with the trees tended to have a more shallow distribution compared to sole-cropped wheat, especially in 2012.

Spatial distribution of root length density (cm 125 cm−3) for wheat and one-year-old walnut tree roots in an intercropping system in 2011 in a young walnut/wheat alley cropping system in Xinjiang, northwest China: a Wheat grown alone, b Wheat intercropped with one-year-old walnut trees, c One-year-old walnut trees grown alone, and d One-year-old walnut trees intercropped with wheat

Spatial distribution of root length density (cm 125 cm−3) for wheat and two-year-old walnut trees roots in an intercropping system in 2012 in a young walnut/wheat alley cropping system in Xinjiang, northwest China: a Wheat grown alone, b Wheat intercropped with two-year-old walnut trees, c Two-year-old walnut trees grown alone, and d Two-year-old walnut trees intercropped with wheat
Spatial distribution of root length density (RLD) of sole-cropped and intercropped walnut
The data for walnut tree root distribution in the wheat/walnut tree intercropping system in both years are presented as contour diagrams (Figs. 2c, d and 3c, d) to demonstrate the general performance of the root distribution. The RLD of walnut was lowered by intercropping with wheat compared to sole-cropped walnut inboth years (Figs. 2c, d and 3c, d). For example, the RLDs of intercropped walnut were reduced compared to sole walnut trees (Fig. 3a, b) at 0–80 cm depth in 2011 and at 0–100 cm depth in 2012. In the tree-crop intercropping in both years a low density of the tree roots extended up to and under the wheat row (Figs. 2d, 3d). There were certain changes in the general patterns of distribution of roots of walnut trees with increasing depth either grown alone or intercropped with wheat in 2012 compared with intercropped walnut in 2011 (Fig. 3c, d). The reduction by intercropping was usually greater deeper in the soil and under the crop row positions. The RLDs of Persian walnut in the wheat/walnut intercropping system in 2011 and 2012 were decreased by 9 and 21 %, respectively, compared to the monocropped trees. In the horizontal direction the roots of intercropped walnut trees spread under the wheat plants and maintained a higher RLD, up to 1.5 m distance from the walnut trees, especially in 2012 (Figs. 2d, 3d).
Rooting depths of intercropped walnut trees were decreased by associated wheat. The RLDs of intercropped walnut decreased especially below 80 cm soil depth compared to the corresponding monocropped walnut trees in 2012 (Fig. 3d). The 4 cm/125 cm3 RLD contour was located below 80 cm soil depth in sole-cropped walnut trees, but the same contour was located above 80 cm soil depth in tree-crop intercropping in 2012 (Fig. 3c, d). However, the difference in rooting depth was not apparent in 2011. For example, the 0.5 cm/125 cm3 RLD contour was distributed below 75 cm in the soil profile when walnut grew alone but was distributed in almost the same position in intercropping in 2011 (Fig. 2c, d). The same phenomenonin walnut was also observed almost throughout the soil profile irrespective of sole cropping or intercropping in 2011 (Fig. 2c, d).
Spatial distribution of root diameter (RD) of sole-cropped and intercropped wheat
The distribution of the RD of wheat is presented in Figs. 4a, b and 5a, b. The average RDs were 0.33 and 0.34 mm for the whole soil profile in sole–cropped wheat in 2011 and 2012, respectively. The average RDs were 0.30 and 0.26 mm for the whole soil profile in intercropped wheat in 2011 and 2012. Intercropped wheat had a lower RD than monocropped wheat. Intercropping led to thinner wheat roots. The average RDs of intercropped wheat decreased by 10 (P > 0.05) and 31 % (P < 0.05) (Figs. 4a, b, 5a, b) compared to sole wheat in both years. The average RDs were 0.45 and 0.46 mm in the top 60 cm of the soil profile of sole wheat in the 2 years but the average RD values were 0.40 and 0.35 mm for intercropped wheat in the same soil depth range. The RDs of intercropped wheat were less than that of sole wheat at the majority of soil depths and especially in the top 60 cm of the soil profile (Figs. 4a, b, 5a, b). The phenomenon was more evident in the intercropped wheat in 2012.

Spatial distribution of root diameter (cm 125 cm−3) for wheat and one-year-old walnut trees roots in an intercropping system in 2011 in a young walnut/wheat alley cropping system in Xinjiang, northwest China: a Wheat grown alone, b Wheat intercropped with one-year-old walnut trees, c One-year-old walnut trees grown alone, and d One-year-old walnut trees intercropped with wheat

Spatial distribution of root diameter (cm 125 cm−3) for wheat and two-year-old walnut trees roots in an intercropping system in 2012 in a young walnut/wheat alley cropping system in Xinjiang, northwest China: a wheat grown alone, b Wheat intercropped with two-year-old walnut trees, c Two-year-old walnut trees grown alone, and d Two-year-old walnut trees intercropped with wheat
Spatial distribution of RD of sole-cropped and intercropped walnut
The distribution of the RD of Persian walnut is presented in Figs. 4c, d and 5c, d. The average RD values were 0.13 and 0.21 mm for the whole soil profile in monocropped walnut trees, but the average values were 0.11 and 0.16 mm in intercropped trees in 2011 and 2012, respectively. In the walnut-wheat alley cropping the average RD of walnut trees was lower than that of wheat (Figs. 4, 5) at all depths. Intercropped Persian walnut trees had lower RD values than sole cropped trees at the associated sampling positions.
The average RDs were 0.19 and 0.30 mm in the top 60 cm of the soil profile in monocropped walnut but the average RDs were 0.17 and 0.24 mm in the corresponding soil profile in intercropped trees in 2011 and 2012, respectively. The RD of intercropped walnut was less than that of sole-cropped trees at the majority of soil depths, especially at 0–60 cm soil depth (Fig. 4c, d, 5c, d). Vertical differences in RD between sole and intercropped trees were apparent. For example, the 0.08-mm RD contour was located below 40 cm soil depth in sole trees but was located above 40 cm soil depth in intercropping in 2011 (Fig. 4c, d). In 2012, the 1.4-mm RD contour was below 40 cm in the soil profile when trees grew alone but was above 40 cm in intercropped plots (Fig. 5c, d).
Spatial distribution of SRL of sole-cropped and intercropped wheat
Figure 6 shows that the SRL of wheat was affected significantly (P < 0.05) by intercropping in which it was greater than the corresponding SRL of sole wheat in 2012 (Fig. 6a–e). However, there was no significant difference in SRL between intercropped wheat and monoculture wheat (P > 0.05) in 2011 at all soil depths. We did not observe any significant difference (P > 0.05) in the effect on the SRL of intercropped wheat of distance from the trees in either year. However, there was a general trend in the intercropping system, i.e. as the distance from the walnut tree increased, the SRL values of intercropped wheat increased gradually in both years. The results indicate that the greater the distance from the walnut tree, the higher the SRL of intercropped wheat.

Specific root length (SRL, cm mg−1) of sole and intercropped wheat at a 0–20 cm soil depth, b 20–40 cm soil depth, c 40–60 cm soil depth, d 60–80 cm soil depth and e 80–100 cm soil depth in a young walnut/wheat alley cropping system in Xinjiang, northwest China. Bars with different small and capital letters denote significant (P < 0.05) difference in the SRL between sole wheat and wheat intercropped with walnut in 2011 and 2012, respectively
Spatial distribution of SRL of sole-cropped and intercropped walnut
The data for walnut SRL in the 2 years are presented in Fig. 7 to demonstrate the general performance of the root spatial distribution. Although there were no significant differences in the SRL of trees in the 2 years at all soil depths, a general increasing trend of the SRL values of Persian walnut as the distance from the walnut trees increased was observed in the sole and intercropping systems. The trend indicates a thinning of the roots with increasing distance from the base of the walnut trees. The fact that older walnut trees had smaller SRL than 1-year-old trees suggests that younger trees had thinner roots. Intercropping increased walnut SRL, with almost all the SRL of intercropped walnut greater than the associated SRL of monocropped trees in the 2 syears. Similar to wheat, we did not observe any significant difference in either year in the effect of distance from the trees on the SRL of intercropped walnut.

Specific root length (SRL, cm mg−1) of sole and intercropped walnut trees at a 0–20 cm soil depth, b 20–40 cm soil depth, c 40–60 cm soil depth, d 60–80 cm soil depth and e 80–100 cm soil depth in a young walnut/wheat alley cropping system in Xinjiang, northwest China. Bars with different small and capital letters denote significant (P < 0.05) difference in the SRL between sole walnut trees and walnut trees intercropped with wheat in 2011 and 2012, respectively
Discussion
Land equivalent ratio (LER), growth, biomass and yield
Walnut/wheat intercropping showed significant yield advantages of intercropping with all LERs of aboveground biomass above one in both years. Droppelmann et al. (2000) reported that the LERs for Acacia saligna (Labill.) H. Wendl/sorghum (Sorghum bicolor (L.) Moench.) intercropping at high planting density were clearly higher than one. Jama et al. (1995) observed that the total LERs were 1.09 for leucaena (Leucaena leucocephala)/maize systems and 1.28 for senna (Senna siamea syn. Cassia siamea)/maize systems in semiarid Machakos, Kenya. In another parallel study Rivest et al. (2010) calculated an LER of 2.4 in a hybrid poplar (Populus nigra L. x P. maximowiczii A. Henry)/soybean association in southwest Quebec, Canada. Mead and Willey (1980) also considered that the yield of the intercropping combination is indeed better than sole cropping in many intercropping systems. Previous work in the same region as the present study involved intercropping of jujube trees (Zizyphus jujuba Mill.) and wheat and Zhang et al. (2013) found that the LER values of three ages of jujube tree-wheat systems were all greater than one. The LERs mentioned above in these agroforestry systems indicate that there is complementarity in resource use between the different species. However, in our study, it is very likely to decrease with time as trees gradually interfere competitively for the underground resources and light; at later stages, allelochemicals released from walnut tree roots (Jose and Gillespie 1998b) may negatively impact wheat growth.
The results show that there was a significant decrease in intercropped wheat yield in 2012 and the aboveground biomass of the intercropped trees also decreased in the same year compared to sole-cropped wheat and walnut trees, respectively. Similarly, tree height, DBH and mean radius of the crown of intercropped trees decreased significantly compared to monocropped trees in 2012. A similar phenomenon was observed in two apple-crop systems in which apple and soybean yields declined by 22 and 12 %, respectively, and the yields of apple and peanut were reduced by 13 and 11 %, respectively (Gao et al. 2013). In another study, Wanvestraut et al. (2004) reported that a barrier treatment had higher cotton lint yield (677 kg ha−1) than the corresponding non-barrier treatment (502 kg ha−1) in a pecan (Carya illinoensis) and cotton (Gossypium hirsutum) intercropping system because of interspecific competition for soil resources in the non-barrier treatment, and the average height of cotton in the barrier treatment was 112 cm while that in the non-barrier treatment was only 82 cm. A similar conclusion was reached by Singh et al. (1989) when they examined a four-year-old Leucaena leucocephala Lam hedgerow/crop intercropping system in which the total dry matter production of both cowpea and sorghum in the alleys was reduced by 70–80 % compared to sole stands. These studies, including our walnut/wheat system, suggest that there is interspecific below-ground competition in the tree/crop mixtures. However, biomass and yield in the tree-crop system did not decrease significantly in 2011, and neither did the tree height, DBH and mean radius of the crown. The results in both years answer our first question and indicate that plant growth, biomass and yield will decrease with belowground interspecific competition in the tree/crop mixtures.
Interspecific competition and root spatial distribution
Root morphology and root distribution are believed to be critical factors in determining the magnitude of interspecific competition underground in intercropping systems (George et al. 1996; Hauggaard-Nielsen et al. 2001). The outcomes of the experiment also answer our second and third questions on root length density, root diameter and specific root length being adversely affected by belowground competition and there were significant differences in RLD, RD and SRL between wheat and walnut trees, and reduced root growth in wheat between two-year-old and one-year-old Persian walnut. Belowground interspecific interactions in the walnut/wheat intercropping appeared in response to competition. The RLDs and RDs of both intercropped species decreased compared to the monocrops. Intercropped wheat had a lower RLD and RD than monocropped wheat in the 2 years (Figs. 2a, b, 3a, b, 4a, b, 5a, b). This indicates that the older the walnut roots the greater the inhibition of the wheat roots. Similar results have been observed in other tree/crop systems. For instance, Wanvestraut et al. (2004) reported that pecan root biomass deeper in the soil (30–90 cm) of a barrier treatment was significantly greater than that found in the non-barrier treatment at the same soil depth in a pecan (Carya illinoensis K. Koch)/cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Wanvestraut et al. (2004) suggested that this rooting pattern of pecan posed intense competition for water with the cotton plants in the non-barrier treatment. Teklehaimanot and Quedraogo (2004) noted that the RLD of sorghum planted with néré (Parkia biglobosa Benth) was reduced due to competition for water between these two species. Other authors have also suggested that the interactions between trees and wheat species may lead to an increased capture of a limiting growth resource (i.e. nutrients, water, light) (Ong et al. 1996; Ashton 2000; García-Barrios and Ong 2004) and may lead to lower RLD of crops. These findings are also in agreement with those of Bolte and Villanueva (2006) who compared pure and mixed stands of European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) in which the sum of species fine root attributes in the mixed stand plots was less than in both pure beech and pure spruce stand plots. Root length: weight ratio (SRL) is a plant parameter that shows the relationship between root penetration intensity and belowground biomass allocation (Zamora et al. 2007). Fitter (1991) and Eissenstat and Yanai (1997) pointed out that the SRL can be seen as an indicator of the reaction of roots to inputs and outputs, and a larger SRL indicates a thinner root. In our study the SRLs of intercropped trees and wheat were higher than those of sole cropped trees and wheat (Figs. 6, 7). This result is in agreement with Zamora et al. (2007) who reported that specific root length was highest in the monoculture (179 cm g−1) and lowest in the non-barrier treatment (146 cm g−1) with the barrier treatment being intermediate (165 cm g−1) in a pecan/cotton intercropping system. The authors considered sole-cropped plants to have greater advantage in allocating carbon in the root system, resulting in higher specific root length. They also found that belowground competition for resources between pecan and cotton in the non-barrier treatment resulted in 25 and 33 % reductions in total root length (359 cm) when compared to that of the barrier (477 cm) and monoculture (539 cm) treatments. Jose et al. (2000a) also found that the fine root biomass in both black walnut and red oak decreased as distance and depth increased in non-barrier treatments compared to barrier treatments in a tree-crop interaction experiment, and the fine root biomass increased at a distance of 1.1 m within the 0–30 cm soil layer for black walnut in the barrier treatment. Other studies have also found that the root density of Douglas fir decreased more gradually with depth in mixtures than in the pure stands in a Douglas fir mixed forestry system (Hendriks and Bianchi 1993). However, this shows that resource competition underground is not always the case in mixed tree–crop intercropping compared to monoculture (Grams et al. 2002). We also found that the roots of intercropped walnut trees spread under wheat (Figs. 2b, d, 3b, d) and tend to have a more shallow distribution in the soil profile compared to sole-cropped walnut trees. Considering the fact that winter wheat germinates in early March, a high demand for nutrients can be expected during that stage. Walnut trees are usually growing in May and the date of spring budbreak for 2011 and 2012 was April 25 and April 23, respectively. It is possible that by the time the walnut tree bloomed the winter wheat had already taken up considerable soil nutrients and water thus forcing the walnut trees to depend more on deeper soil nutrients for growth and development when interplanted in an alley cropping system. Given this, we consider that giving more water or fertilizer may help intercropped wheat/trees to grow more. The shallower fine root systems of mixed trees compared to pure trees may increase drought vulnerability (Schmid 2002).
Allelopathy
Allelochemicals are present in many plant species and are released into the soil around the roots by a variety of mechanisms including decomposition of residues and root exudation. Allelochemicals such as juglone are known to affect the germination, growth and root distribution of a number of plant species (Jose et al. 2004). Walnut is a notorious allelopathic tree species (Jose and Gillespie 1998b). There have been many studies in recent years on walnut multi-cropping systems and one of the dominant narratives about walnut has remained approximately the same: allelopathic in general (Ares and Brauer 2004; Bertin et al. 2003; Bohanek and Groninger 2005; Jose and Gillespie 1998a, b). However, it has been emphasized that juglone would be unlikely to affect sensitive species after the initial planting of walnut. Hence, many plants are potential candidates for interplanting with black walnut in the first few years (Scott and Sullivan 2007). Based on this view and considering that the Persian walnut trees used in our study were still very young the role of allelopathy as a factor affecting the root distribution of both crops can be ruled out.
Shade effects
Shading by associated trees is a factor reducing crop yields in agroforestry systems (Jose et al. 2004). Competition for light between perennial woody trees and annual crops will lead to yield or biomass decline (Gillespie et al. 2000; Lin et al. 1999; Nissen et al. 1999). However, the competitive intensity for light is influenced by many factors such as canopy structure, alley width, plant age and tree height (Jose et al. 2004), and especially alley width and mean radius of the tree crown. Burner and Brauer (2003) showed that herbage yield was unaffected at alley widths of 4.9 m and above in a loblolly pine-mixed grass/forb silvopasture. Light transmittance was up to 90 % at this spacing. Garrett and Jones (1998) suggested that shade will not be a consideration during the early years of in a walnut/wheat system in Missouri in the USA. In our study the mean radius of the walnut tree crown was only 17.5 and 19.5 cm in intercropping systems (2011 and 2012, respectively), the tree ages were 1 and 2 years, and the intercropping fields had a larger alley width (6 m), so that canopy width and the corresponding shading could not change the amount of light reaching the understory canopy. Similarly, the tree crown width and tree spacing had little effect on the microclimate of the intercropping system. Thus, the shade effects from Persian walnut trees to wheat were considered to be negligible and ignored in the present study.
Conclusions
Tree-based intercropping systems are very important in Hetian old oasis and are an effective practice for increasing the efficiency of arable land use. We conclude that competition between young Persian walnut trees and wheat was mainly belowground. The roots of the trees spread under the cereal crop and there were consequently relatively low RLDs, RDs and higher SRLs at all soil depths compared with sole-cropped walnut trees, especially in the second year (2012). The roots of wheat intercropped with Persian walnut tended to have a more shallow distribution in the soil profile and had smaller RLD, RD and higher SRL than sole-cropped wheat. The closer to the tree row the more intense was the interspecific competition. Apparent root competition decreased the yield and biomass of both tree and crop species. This provides direct evidence for our hypothesis that the lower growth in the tree-crop association resulted from interspecific competition belowground and less RLD and RD and more SRL in the soil profile. Furthermore, there were significant differences in RLD, RD and SRL of wheat, and in the inhibitory effects on wheat root growth between two-year-old and one-year-old Persian walnut. Overall, a long-term study should be established to examine the rooting patterns in Hetian old oasis over longer periods of many years to determine the dynamics of interspecific competition and root growth in the tree-crop intercropping system.
References
Ares A, Brauer D (2004) Growth and nut production of black walnut in relation to site, tree type and stand conditions in south-central United States. Agrofor Syst 63:83–90
Ashton PS (2000) Ecological theory of diversity and its applications to mixed species plantation systems. In: Ashton MS, Montagnini F (eds) The silvicultural basis for agroforestry systems. CRC Press, Boca Raton, pp 61–77
Bertin C, Yang X, Weston LA (2003) The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 256:67–83
Bohanek JR, Groninger JW (2005) Productivity of European black alder (Alnus glutinosa) interplanted with black walnut (Juglans nigra) in Illinois, USA. Agrofor Syst 64:99–106
Böhm W (1979) Methods of studying root systems. Springer, Berlin
Bolte A, Villanueva I (2006) Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur J For Res 125:15–26
Burner DM, Brauer DK (2003) Herbage response to spacing of loblolly pine trees in a minimal management silvopasture in southeastern USA. Agrofor Syst 57:69–77
Cannell MGR, Van Noordwijk M, Ong CK (1996) The central agroforestry hypothesis: the tree must acquire resources that the crop would not otherwise acquire. Agofor Syst 34:27–31
Cao FL, Kimmins JP, Wang JR (2012) Competitive interactions in Ginkgo and crop species mixed agroforestry systems in Jiangsu, China. Agrofor Syst 84:401–415
Chamshama SAO, Mugasha AG, Kløvstad A, Haveraaen O, Maliondo SMS (1998) Growth and yield of maize alley cropped with Leucaena leucocephala and Faidherbia albida in Morogoro, Tanzan. Agrofor Syst 40:215–225
Dhima KV, Lithourgidis AS, Vasilakoglou LB, Dordsas CA (2007) Competition indices of common vetch and cereal intercrops in two seeding ratio. Field Crop Res 100:249–256
Droppelmann KJ, Ephrath JE, Berliner PR (2000) Tree/crop complementarity in an arid zone runoff agroforestry system in northern Kenya. Agrofor Syst 50:1–16
Eissenstat DM, Yanai RD (1997) The ecology of root life span. Adv Ecol Res 27:1–6
Fitter AH (1991) Characteristics and functions of root systems. In: Waisel Y, Eshel A, Kafkafi U (eds) Plant roots: the hidden half. Marcel Dekker, New York, pp 3–25
Fletcher EH III, Thetford M, Sharma J, Jose S (2012) Effect of root competition and shade on survival and growth of nine woody plant taxa within a pecan [Carya illinoinensis (Wangenh.) C. Koch] alley cropping system. Agrofor Syst 86:49–60
Gao LB, Xu HS, Bi HX, Xi WM, Bao B, Wang XY, Bi C, Chang YF (2013) Intercropping competition between apple trees and crops in agroforestry systems on the Loess Plateau of China. Plos One 8:1–8. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0070739
García-Barrios L, Ong CK (2004) Ecological interactions in simultaneous agroforestry systems in the tropics: management lessons. Agrofor Syst 61:221–236
Garrett HE, Jones JE (1998) Companion crop recommendations for planting with black walnuts. In: Jones JE, Mueller R, Van Sambeek JW (eds). Nut production handbook for eastern black walnut. Southwest Missouri Resources, Conservation & Development (RC & D) Inc. Columbia, pp 102–104
George SJ, Kumar BM, Wahid PA, Kamalam NV (1996) Root competition for phosphorus between the tree and herbaceous components of silvopastoral systems in Kerala, India. Plant Soil 179:189–196
Gillespie AR, Jose S, Mengel DB, Hoover WL, Pope PE, Seifert JR, Biehle DJ, Stall T, Benjamin TJ (2000) Defining competition vectors in a temperate alley cropping system in the midwestern USA. 1 Production physiology. Agrofor Syst 48:25–40
Grams TEE, Kozovits AR, Winkler JB, Sommerkorn M, Blaschke H, Häberle KH, Matyssek R (2002) Quantifying competitiveness in woody plants. Plant Biol 4:153–158
Greb BW, Black AL (1961) Effects of windbreak plantings on adjacent crops. J Soil Water Conserv 16:223–227
Gregory PJ (1996) Approaches to modeling the uptake of water and nutrients in agroforestry systems. Agrofor Syst 34:51–65
Gruenewalda H, Brandt BKV, Schneider BU, Bens O, Kendziab G, Hüttl RF (2007) Agroforestry systems for the production of woody biomass for energy transformation purposes. Ecol Eng 29:319–328
Hauggaard-Nielsen H, Ambus P, Jensen ES (2001) Temporal and spatial distribution of roots and competition for nitrogen in pea-barley intercrops—a field study employing 32P technique. Plant Soil 236:63–74
Hendriks CMA, Bianchi FJJA (1993) Root density and root biomass in pure and mixed forests of Douglas fir and beech. Neth J Agric Sci 43:321–331
Imo M (2009) Interactions amongst trees and crops in taungya systems of western Kenya. Agrofor Syst 76:265–273
Jama BA, Nair PKR, Rao MR (1995) Productivity of hedgerow shrubs and maize under alley cropping and block planting systems in semiarid Kenya. Agrofor Syst 31:257–274
Jamaludheen V, Kumar BM, Wahid PA, Kamalam NV (1997) Root distribution pattern of the wild jack tree (Artocarpus hirsutus Lamk.) as studied by 32P soil injection method. Agrofor Syst 35:293–336
Jose S, Gillespie AR (1998a) Allelopathy in black walnut (Juglans nigra L.) alley cropping. I. Spatio-temporal variation in soil juglone in a black walnut-corn (Zea mays L.) alley cropping system in the midwestern USA. Plant Soil 203:191–197
Jose S, Gillespie AR (1998b) Allelopathy in black walnut (Juglans nigra L.) alley cropping. II. Effects of juglone on hydroponically grown corn (Zea mays L.) and soybean (Glycine max L. Merr.) growth and physiology. Plant Soil 203:199–205
Jose S, Gillespie AR, Seifert JR (2000a) Defining competition vectors in a temperate alley cropping system in the Midwestern USA. 2. Competition for water. Agrofor Syst 48:41–49
Jose S, Gillespie AR, Seifert JR, Mengel DB, Pope PE (2000b) Defining competition vectors in a temperate alley cropping system in the mid-western USA. 3. Competition for nitrogen and litter decomposition dynamics. Agrofor Syst 48:61–77
Jose S, Gillespie AR, Seifert JR, Pope PE (2001) Comparison of minirhizotron and soil core methods for quantifying root biomass in a temperate alley cropping system. Agrofor Syst 52:161–181
Jose S, Gillespie AR, Pallardy SG (2004) Interspecific interactions in temperate agroforestry. Agrofor Syst 61:237–255
Jose S, Williams RA, Zamora DS (2006) Belowground ecological interactions in mixed-species forest plantations. For Ecol Manag 233:231–239
Li Z-X, Yang L-L, Abulizi B, Yan S-K, Liu K, Zhu Z-J, Yang Z-Q, Liu J, Sheng X-H, Maihemujiang, Ma Z-J (2014) The development of walnut mechanization of production equipment in Xinjiang. Xinjiang Agric Sci 51:973–980
Lin CH, McGraw RL, George MF, Garrett HE (1999) Shade effects on forage crops with potential in temperate agroforestry practices. Agrofor Syst 44:109–119
Livesley SJ, Gregory PJ, Buresh RJ (2000) Competition in tree-row agroforestry systems. 1. Distribution and dynamics of fine roots length and biomass. Plant Soil 227:149–161
Mead R, Willey RW (1980) The concept of a land equivalent ratio and advantages in yields from intercropping. Exp Agric 16:217–228
Nissen TM, Midmore DJ, Cabrera ML (1999) Aboveground and belowground competition between intercropped cabbage and young Eucalyptus torelliana. Agrofor Syst 46:83–93
Ong CK, Black CR, Marshall FM, Corlett JE (1996) Principles of resource capture and utilization of light and water. In: Ong CK, Huxley P (eds) Tree-crop interactions: a physiological approach. CAB International, Wallingford, pp 73–158
Ozkan MM (2009) Some nutritional characteristics of fruit and oil of walnut (Juglans regia L.) growing in Turkey. Iran J Chem Chem Eng 28:57–62
Ozkan G, Koyuncu MA (2005) Physical and chemical comparison of some walnut (Juglans regia L.) genotypes grown in Turkey. Grasas Aceites 56:141–146
Pham CH, Yen CP, Cox GS, Garrett HE (1977) Slope position, soil water storage capacity, and black walnut root development. In: Balmer WE (ed) Soil moisture-site productivity management symposium. USDA Forest Service, Southern Forest Experiment Station, Myrtle Beach, pp 326–335
Qiao X, Lei JJ, Chen XW, Zhao Q, Zhang HZ, Huang TR, Li GK (2012) Study on growth, development and yield formation of wheat under walnut-wheat intercropping system. Chin J Agrometeorol 33:540–544
Rao MR, Sharma MM, Ong CK (1990) A study of the potential of hedge row intercropping in semi-arid India using a 2-way systematic design. Agrofor Syst 11:243–258
Rao MR, Sharma MM, Ong CK (1991) A tree/crop interface design and its use for evaluating the potential of hedge row intercropping. Agrofor Syst 13:143–158
Rivest D, Cogliastro A, Bradley RL, Olivier A (2010) Intercropping hybrid poplar with soybean increases soil microbial biomass, mineral N supply and tree growth. Agrofor Syst 80:33–40
Schmid I (2002) The influence of soil type and interspecific competition on the fine root system of Norway spruce and European beech. Basic Appl Ecol 3(4):339–355
Schroth G (1999) A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agrofor Syst 43:5–34
Scott R, Sullivan WC (2007) A review of suitable companion crops for black walnut. Agrofor Syst 71:185–193
Singh RP, Ong CK, Saharan N (1989) Above and below ground interactions in alleycropping in semi-arid India. Agrofor Syst 9:259–274
Stone EL, Kalisz PJ (1991) On the maximum extent of tree roots. For Ecol Manag 46:59–102
Teklehaimanot JB, Quedraogo SJ (2004) Fine root distribution of pruned trees and associated crops in parkland system in Burkina Faso. Agrofor Syst 60:13–26
van Asten PJA, Wairegi LWI, Mukasa D, Uringi NO (2011) Agronomic and economic benefits of coffee–banana intercropping in Uganda’s smallholder farming systems. Agric Syst 104:326–334
Van Noordwijk M, Purnomosidhi MP (1995) Root architecture in relation to tree–soil–crop interactions and shoot pruning in agroforestry. Agrofor Syst 30:161–173
Vandermeer JH (1989) The ecology of intercropping. Cambridge University Press, New York
Wanvestraut RH, Jose S, Nair PKR, Brecke BJ (2004) Competition for water in a pecan (Carya illinoensis K. Koch)—cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Agrofor Syst 60:167–179
Willis RJ (2000) Juglans spp., juglone and allelopathy. Allelopath J 7:1–55
Zamora DS, Jose S, Nair PKR (2007) Morphological plasticity of cotton roots in response to interspecific competition with pecan in an alley cropping system in the southern United States. Agrofor Syst 69:107–116
Zhang BJ, Gao YH (2010) Overview of walnut culture in China. Acta Hortic (ISHS) 861:39–44
Zhang W, Ahanbieke P, Wang BJ, Xu WL, Li LH, Christie P, Li L (2013) Root distribution and interactions in jujube tree/wheat agroforestry system. Agrofor Syst 87:929–939
Acknowledgments
This work was financially supported by the Chinese Ministry of Agriculture (Project No. 201003043-01) and the National Natural Science Foundation of China (Project No. 31460335). We also thank Dr. Hans Lambers and two anonymous reviewers for their valuable suggestions and comments toward improving an early version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Zhang, W., Ahanbieke, P., Wang, B.J. et al. Temporal and spatial distribution of roots as affected by interspecific interactions in a young walnut/wheat alley cropping system in northwest China. Agroforest Syst 89, 327–343 (2015). https://doi.org/10.1007/s10457-014-9770-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-014-9770-x