
Abstract
Fish pond systems are managed with different practices. Among them, a dry period with one year without water is applied in some cases to promote mineralization of the sediments and control the development of pathogenic bacteria. This dry period induces a drastic disturbance on the plant communities. The objective of this work was to study the influence of a one-year dry period applied every five to seven years on aquatic plant diversity and abundance. For this, we studied the aquatic plant community of 149 fish ponds during the first year after a dried period (Y1), and ponds with a dried period dating back two years (Y2), three years (Y3), four years (Y4) and five to seven years (Ysup5). According to Jackknife index, mean species richness was highest for Y1, with 29 species compared to the other years (24 species for Y2; 19 for Y3; 15 for Y4 and 17 for Ysup5). A total of 15 species were identified as species unique to Y1 and were competitive, fast colonizer and disturbance-tolerant species. Most of these Y1 species developed during the dry year and remain only one year after refilling. After Y1, the evolution of communities was linked to the phenomenon of nestedness based on a loss of several species but not on a complete turnover, with most of species present independently of time. We conclude that a periodic dry period maintains a cycle in plant succession and accommodates highest species richness at the beginning of the cycle.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Ponds are considered isolated systems with connections to major streams, ditches or other waterbodies (Oertli and Frossard 2013). The temporary isolation from the hydrological current system induces stagnant water. The limited average water depth categorizes them as shallow waters. Besides these characteristics, ponds are usually subject to anthropogenic influences, making them unique aquatic environments (Sayer and Greaves 2019).
Fish ponds have been managed for centuries with the economic purpose to provide inland produced fish (Hancz et al. 2015). Fish ponds are aquatic, human-made ecosystem, where several possible farming practices are applied, such as fish stocking, liming, fertilization, feeding and dry-out of the pond. All these actions affect the natural balance of a pond and its trophic web. Therefore, fish ponds are usually described as eutrophicated shallow waterbodies (Robin et al. 2014). Even if fish ponds are artificial waterbodies, they contribute significantly in some regions to the regional biodiversity (Magnus & Rannap 2019; Zamora et al. 2021).
Variations of hydroperiod are common in fish ponds (Šumberová et al. 2021). Sometimes ponds can be naturally affected by severe drought (Collinson et al. 1995). Also, some management practices such as the draining of the fish ponds for fish harvesting induce a short dry period before ponds are refilled. In some cases in Europe, this dry period is prolonged for a complete year, with a crop production in the pond during this period. This one-year drought leads to a complete destruction of the upper parts of the submerged plant communities. During this long disturbance, the sediment surface comes in contact with air and a shallow tillage is applied on the eight to ten centimetres of the bottom before cropping. As a result, oxygen and terrestrial bacteria change the biological processes with a direct effect on the decomposition of organic matter. Sediments tend to be remineralized after each drought (Collinson et al. 1995). When the pond is refilled with water, the nutrient concentrations shift from organic to mineral forms of nitrogen and phosphorus (O’Farrell et al. 2011). Thus, a dry period creates a strong disturbance for the biocenosis after refilling, with new patterns of recovery for chemical and physical processes (Lake 2003). Consequently, the dry period causes important changes in the trophic network by altering the interactions between species groups (Lake 2003; Humphries and Baldwin 2003).
Among the different communities living in such ecosystems, phytoplankton and aquatic plant communities play the roles of primary producers in the trophic network and consumers of mineralized nutrients. Both types of species compete for the same resources, mainly nutrients and light. According to the theory of alternative stable states (Scheffer et al. 1993), one type of species can become dominant in the ecosystem, depending on environmental conditions. In different situations of nutrient concentrations, a clear water state appears and can promote macrophyte dominance (Beklioglu and Moss 1996; Matsuzaki et al. 2020). Within the macrophyte community, a species’ ability to competitively access to limited resources creates different gradients of dominance (Mcceary 1991).
Abundance and diversity of aquatic plants are thus generally closely related to nutrient concentrations (Sarkar et al. 2020). Their diversity is usually lower in ponds with a high concentration of phosphorus due to higher competition with phytoplankton (Korner and Nicklisch 2002). In addition, their functional richness declines when phosphorus concentrations increase (Arthaud et al. 2012b). When nutrient concentrations are moderate, aquatic plants colonize from the shore to the middle of the pond (Oertli and Frossard 2013). In the case of low nutrient concentrations, species with adapted morphologies are able to gather nutrients in sediment or water. Rooted plants are able to access the nutrients from sediment. Submerged plants meet their needs by collecting nutrients from both water and sediment. Floating leaf plants with roots take advantage of free-floating plants, which are only able to get resources from the water column (Bini et al. 1999). These free-floating macrophytes are found in deeper locations with less competition, more nutrient resources and better access to light. Thus, low nutrient conditions often allow a higher biomass and diversity of aquatic plant in ponds.
As a limiting factor, light availability modifies the composition of the aquatic plant community (Toivonen and Huttunen 1995) with an organization following a vertical gradient along the water column (Oertli and Frossard 2013). Even if competition exists among submerged macrophytes (Van Donk et al. 1993) due to light catchment, the main factor limiting aquatic plant development is turbidity. It increases in correlation with phytoplankton growth. Submerged plants shift from the stage of dominance to disappearance when high turbidity makes access to light impossible (Scheffer and Van Nes 2007). More competitive species need optimal access to solar radiation (Netten et al. 2011). Morphologies of leaves can evolve so as to allow the plant to maximize radiation catchment. For every five centimetres underwater, the average radiation changes significantly as well as tissue development for adaptation (Asaeda and Van Bon 1997).
Aquatic plant communities use strategies at different scales to adapt to a disturbance such as drying out or a variation of the water level (Zhao et al. 2021). The recovery promotes a particular species richness which tends to be higher the first year after the dry period than the following years (Arthaud et al. 2013). The highest species richness after a drought is associated with the occurrence of terrestrial species (Sandi et al. 2020), but also small species with sexual reproduction and without storage organs (Arthaud et al. 2012b). The group of submerged aquatic plants are considered the pioneers in such colonization (Qiu et al. 2001; Zhang et al. 2019). They are able to photosynthesize underwater, allowing them to colonize the water after a severe drought disturbance first (Van Den Berg et al. 2001;). They are less affected by the necessity to reach the surface. The division of Charophyta is the dominant group during the first year after the dry period (Hilt and Gross 2008; Zhang et al. 2019). Over a longer timeframe, the colonization of Charophyta occurs first, followed by submerged angiosperms, other green algae and then, cyanobacteria (Scheffer and Van Nes 2007).
The recovery after a dry period also allows new populations to settle (Scheffer and Van Nes 2007). Among the plants studied, some species show a terrestrial life-form capacity. They grow during the dry period but are also able to remain during the following years with water. Seed banks and undestroyed plants during the dry period can be important factors for generating successful recovery (Arthaud et al. 2012a). Furthermore, some observations show that dry periods help the ecosystem to host rare species after refilling (Collinson et al. 1995). Competition and dominance pressure influence which particular scheme of species settlement occurs. Consequently, the first year after the dry period is distinctive for development of aquatic plant communities and species diversity (Kelleway et al. 2020; Caria et al. 2021).
Resilience theory is used to understand the capability of a system to recover after a disturbance (Schulze 1996; Holling 1987). The resilience of an ecosystem is based on its capability to self-organize and adapt to new conditions (Sarremejane et al. 2020). According to resilience theory, a higher diversity of species is expected to provide a larger range of performances and responses to changes. A managed connectivity across generations increases the memory of responses. Seed banks are an example of connectivity through time (Holling 1987). A disturbance can bring drastic changes or shifts from one state to another as a more continuous process with the succession of several communities. This uninterrupted chain of changes assures the ability of an ecosystem to “memorize” previous events (Scheffer and Van Nes 2007). From one season to another, the communities are subject to different conditions. The heterogeneity among the species, species richness, but also diversity in functional and morphological traits, also provide a larger variety of adaptation strategies. Therefore, ecosystems seem to be capable to re-organize themselves in reaction to different disturbances according to adaptive cycles influenced by biological and environmental parameters (Fath et al. 2015).
The specific aims of this study are (a) to study the variations in the aquatic plant species richness and diversity in fish ponds during the years following a one-year dry period, (b) to identify specific species that are adapted to recovery after a dry period, (c) to investigate how functional traits vary depending on the number of years since pond refilling, and (d) to evaluate the resilience of ponds in relation to this disturbance regime.
Material and methods
Study area
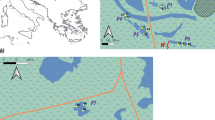
The study was carried out in the Dombes region in southeastern France which is characterized by about 1100 man-made fish ponds and 11,200 ha of water surface organized in connected networks. In this region, ponds have an average surface of 10 ha and a mean depth of 0.8 m. The maximum depth is about 2.5 m but the ponds have a specific topography and the deepest zone represents less than 10% of the surface.
In these ponds, fish were harvested once every year in autumn or winter after draining. The ponds were refilled rapidly with water from either upstream ponds or from rainfall coming from the pond catchment. Fish were stocked in spring after water refilling. After four years of this alternation of fish production and fish harvest in November or December, the ponds were left to dry up for one year. During the dry phase and from Mid-April to early May depending on the weather conditions, a slight tillage was performed on pond bottom to a maximum depth of 10 cm before cropping. The crops consisted of oats, maize, buckwheat or sorghum. At the end of the dry year, water refilling was performed from October after crop harvest. The primary fish species raised in fish ponds were common carp, with more than 60% on the total fish yield, followed by roach and rudd (30%), and a lower quantity of tench, pike or pikeperch (10%) (Wezel et al. 2013). Total fish stocking was between 40 and 60 g m−3.
In total, 149 fish ponds were sampled during the 2008–2020 period, with an average of 12 ponds monitored each year. Among the studied ponds, 33 were sampled the first year after the dry year (Y1), 36 ponds two years after the dry period (Y2), 34 ponds three years after the dry period (Y3), 24 ponds four years after the dry period (Y4), and 22 ponds five to seven years since the last dry period (Ysup5). Three ponds were sampled during the dry period. All ponds were sampled using the following methodology.
All the ponds of the dataset were selected according to the homogeneity of application of the same practices by fish farmers in order to have small range of values for physico-chemical parameters. The transparency and nutrient concentrations were calculated on the basis of the median of six values measured in May end and June during the development phase of aquatic plants and before aquatic plant sampling. The transparency varied between 72 and 93 cm according to the ponds. Total nitrogen and phosphorus concentration in water were between 1.2 and 1.6 mg/L and 0.21 and 0.27 mg/L, respectively.
Aquatic plant sampling
Submerged and floating aquatic plants were sampled in July in a water depth ranging between 60 and 130 cm. A quadrat sampling method was used. The pond was divided into transects and for each transect quadrats of 4 m2 were selected each 50 m (Fig. 1). The total number of quadrats was based on the pond surface (Arthaud et al. 2012b) in order to estimate the observed richness and the percentage of cover (abundance). The percentage of cover for each species was calculated according to the Braun-Blanquet cover-abundance method. We used a scale from 1 to 5 for the ranges of cover: 1 for < 5%, 2 for 5–25%, 3 for 25–50%, 4 for 50–75% and 5 for 75–100% (Wikum and Shanholtzer 1978). The Braun-Blanquet scores for each species were then converted to mean values of percentage cover (2.5; 15; 37.5; 62.5 and 87.5%) to allow statistical analysis (Van der Maarel 2007). The abundance was calculated for each species observed and takes into account the overlapping of plants through the water column. For the purpose of statistical analysis, we have chosen to calculate the means of percentage cover of total number of quadrats per species per site.

Illustration of the quadrat method sampling in a pond
Statistical analysis
All statistical analyses were conducted with the software R version 3.2.4 (R Development Core Team 2010) and its packages Vegan, Indicspecies and Ggplot2.
In accordance with previous studies on fish ponds (Vanacker 2015), the Jackknife index was used to estimate aquatic plant species richness. We used first-order Jackknife richness estimator (package ‘vegan: ecological diversity’ in R), calculated with the following formula:
\({S}_{obs}\): total number of species observed in the sample\({f}_{1}\): number of singleton species (species occurring only once in the data set)
The nonparametric tests of Kruskal–Wallis were conducted on richness and means of abundances per year after the last dry period to compare one year to another (Y1 to Ysup5).
To measure representativeness of each plant species for the different years, the association strength was calculated. This index represents the correlation between the target site community observed each year. The values (positive or negative) reveal the correlation between the observed abundance and the expected abundance under the null hypothesis ‘no relation’. A negative correlation means that the species is excluded from the target group of the year. The formula corresponds to the calculation of a r2 coefficient:
The number of groups is defined by the index K. Np is the expected number of species per group. The parameter ap is the expected sum of the abundances per group. N represents the number of observed species. The indexes can be detailed with Np g = N/K, for the index agp = Npg(ap/Np) and for the index ag = Npg *\(\sum_{\mathrm{k}=1}^{\mathrm{k}}(\mathrm{ak}/\mathrm{Nk})\) (Source: De Caceres and Legendre, 2009).
In our study, we selected, as representative plants for one year, the species for which the correlation index was greater than 0.30 for one year, and negative for all other years. The individual-based index relates species to a target group represented by a year. Indeed, we wanted to look at the particularities of the target group defined in Y1. The closer the value is to 1, the more representative the species is of the group.
Results
Species richness and percentage of cover of aquatic plants
The total number of aquatic plants species observed from 2008 to 2020 in all fish ponds samples was 119 species. The aquatic plant species richness in Y1 was significantly higher than species richness of the other groups, with a mean of 30 species (Fig. 2). Mean species richness declined progressively over the years until Y4, with 16 species. For Ysup5, a slight increase was identified, but one that was not significantly different from the species richness of Y4.

Species richness and standard deviation of aquatic plants of fish ponds according to the year since the last dry year, based on Jackknife index (left figure). Percentage of cover and standard deviation of aquatic plants according to the year since the last dry year (right figure). Y1 means first year after a dry year. Y2, Y3 and Y4 are, respectively, the second to fourth years after a dry year. Ysup5 corresponds to the fifth to seventh year after a dry year. The different letters discriminate the level of significance
The percentage of cover of aquatic plants was also the highest for Y1, with a mean of 65% (Fig. 2). It declined progressively until Y4 (33%) and remained stable afterwards.
Evolution of the plant community over the years
Different analyses were carried out to define a group of species representative of Y1. Among the 119 species observed, 15 were found as specific species for Y1 (Table 1). Among the 15 significantly representative species of Y1, Lemna minor, Oenanthe aquatica, Lycopus europaeus, Alisma plantago-aquatica, Ludwigia palustris, Lythrum salicaria, Alopecurus geniculatus and Rorippa amphibia are characterized by high index value close to 1 (> 0.6). Both components of specificity (A) and fidelity (B) allow these species to occur widely and regularly in Y1 sites. Permutation tests revealed significant p values that confirm the specificity of these plants to Y1 without possible bias (p < 0.01).
Lemma minor, Oenanthe aquatica, Lycopus europaeus, Alopecurus geniculatus and Rumex conglomeratus stand for five species largely restricted to Y1 in the open water area (with A values > 0.92). They are almost exclusively present only during Y1, with R. conglomeratus exclusively found in samples from sampled from this year. However, this species appears in a relative small proportion of sites belonging to Y1 (B = 0.133). Among the indicator species, Riccia fluitans, Rorippa amphibia, Sparganium erectum and Juncus articulatus also were significantly more likely to be found in sites belonging to Y1 (respectively A values between 0.988 and 0.747) but not exclusively (respectively B = 0.267; 0.467; 0.467; 0.333). Mentha aquatica and Persicaria hydropiper were less likely to be found in all Y1 ponds (respectively B = 0.200 and 0.267). L. minor reveals a high degree of fidelity to the group Y1 (B = 0.933) as well as O. aquatica (B = 0.800), Alisma plantago-aquatica (B = 0.800), Ludwigia palustris (B = 0.733), Lythrum salicaria and Ranunculus peltatus (B = 0.667). Most of the sites (> 66%) where they were recorded correspond to Y1.
Three to six species were also associated with a year from Y2 to YSup5. For example, Utricularia ochroleuca, Elatine hydropiper, Lemna gibba, Luronium natans, Hydrocharis morsus and ranae were found as specific for Y2 (Table 2). But according to our methodology, no species appeared statistically representative of a particular year, because of correlation index lower than 0.3.
In the three ponds sampled during the dry year, 33 species were observed in total. Among these species, 10 of the 15 representative species of Y1 were observed. A. geniculatus, L. europaeus, O. aquatica, P. hydropiper, R. peltatus, R. amphibia, R. conglomeratus occurred in all three dried ponds while A. plantago-aquatica, J. articulatus, L. salicaria occurred in only one pond.
We have studied the species occurring in more than 5% of the ponds independently of the time after dry year (Table 3). According to the total dataset, the representative species of Y1 are not considered as the most common species found in ponds, except Lemna minor, Alisma plantago-aquatica and Ranunculus peltatus.
We used statistical analyses based on the occurrence of plant species for the different years and using for Y1 individual-based index A and B components. According to the bibliographical knowledge on the specific traits of each species, we have determined four successive stages in fish pond evolution that occurs after a one-year dry period. These states may be highlighted as steps of pond evolution (Fig. 3). The first state is the resurgence of aquatic state after the dry period (state A) with species observed during the other years but also several specific species not present after Y1. The second state (state B), which was observed in ponds from sampling groups Y2 to Y4, is characterized by high competition for resources with presence of competitive species. State C (Ysup5) is distinguished as having established conditions with also presence of competitors. Finally, state D can be described as the dry year when amphibious species establish from the propagule bank and supply in their turn the propagule bank.

Alternative states cycle adapted from resilience theory illustrating the four states in a fish pond evolution facing a one-year dry period. Inspired by the representation of an adaptive system facing a disturbance (Gunderson 2001)
Discussion
Among the results found, the result of central interest is that highest plant species richness was observed the first year after dry year. We also observed a strong decrease in species richness and a change of species composition from Y1 to Y2. Further, 15 species among the 119 species were identified as representative for Y1, but none of the species was specific to the other years. The decrease in species richness from Y1 to Y2 was linked to a loss of several species but not to a complete species turnover. This last result can be explained as the phenomenon of nestedness, indicated by the pattern characterized by the poorest communities (Y2 to Ysup5) composed of a strict subset of the species found in the richest communities (Y1) (Baselga 2010).
Although the occurrence of most species does not vary much over time from Y2 to Ysup5, we observed that the abundance of each species can differ more significantly. Arthaud et al. (2012b) shows that the duration in water after a dry year does not influence the functional richness but changes the abundance of life-history traits corresponding to morphology, fecundity and longevity of aquatic plants.
Representative species of Y1 and connections to the dry period
Among the 15 representative species of Y1, 12 species were emergent macrophytes whose vegetative parts can appear out of the water. Their ability to find nutrients in the sediment and photosynthesize above water offers them the possibility to be the most productive. These representative emergent species of Y1 are amphibious plants that have a high level of tolerance to floods or drought periods (Willby et al. 2000). These plants are species typically found in seasonally or temporarily inundated environments (Crawford 1977; Willby et al. 2000; Greet et al. 2013).
Ten of the 15 Y1 species were also observed in the ponds sampled during the dry year. This finding indicates that representative species of Y1 develop during the dry period and are able to maintain their population during Y1. Among these species, Mentha aquatica, Ludwigia palustris and Sparganium erectum are known to have the capacity to grow in waterlogged ground in the pond border area.
Among the representative Y1 species, only three are not considered amphibious: Ranunculus peltatus, Lemna minor and Riccia fluitans. The presence of Ranunculus peltatus, as a medium caulescent plant, can be attributed to its ability to produce seeds before the dry period. R. peltatus is known to develop a very important plasticity conferring competitive advantages, which could explain its ability to spread (Garbey et al. 2004). We can suppose that low turbidity facilitated a high ability of seed germination, thereby supporting the development of R. peltatus during Y1. Lemna minor and Riccia fluitans are free-floating plants considered opportunistic with a high growth rate.
Some species observed during the dry year were also abundantly present from Y2 to Y5 as Scirpus martimus, Glyceria fluitans and Persicaria amphibia. These are perennial and competitive species with strong root systems, able to survive in relatively deep and turbid water. Thus, they were adapted to colonize the major surface of our shallow fish ponds characterized by a mean depth of 80 cm.
Other emergent species were observed in the ponds sampled during the dry year which were not found in any pond with water. These species were Bidens tripartita, Echinochloa crus-galli and Persicaria lapathifolia. They are specific to wetlands with waterlogged ground but survive badly in shallow water.
Adaptation of species of Y1 with different strategies
Representative species of Y1 were competitive, fast colonizing, and disturbance-tolerant species. The strategy of competitive colonization is characterized by an important seed dispersion and focus on clonal growth (Wildova et al. 2007). Two abilities are highlighted: colonization with extensive spread development and competition trade-off to face pressure in the free areas and to reach resources. The species indicators of Y1 mainly focus on colonizing in both terrestrial and aquatic conditions in order to be prepared for periods of drought as well as flood.
Some species representative of Y1 are disturbance-tolerant plants whose both vegetative reproduction and extensive seed bank facilitate their presence during the following years in the area (Murphy et al. 1990). We found here Lemna minor, Ranunculus peltatus, Rorippa amphibia, Sparganium erectum, Juncus articulatus, Riccia fluitans. However, some Y1-representative species have developed another strategy regarding reproductive aspects. They have the ability to produce a high density of persistent seeds during a dry period (Arthaud et al. 2012b). This is the case for Alisma plantago-aquatica, Ludwigia palustris and Rumex conglomeratus, whose reproduction is mainly based on seed production and dispersion. Long-resistant seeds and germination on dried sediments are also a way to survive during dry periods and to maintain after water refilling.
Some other Y1 species are very competitive and fast colonizing. This type of Y1 species is, in many cases, deeply and extensively rooted into the sediments, allowing resistance to disturbance (Mari et al. 2010; Zealand and Jeffries 2009). Juncus articulatus, Ludwigia palustris and Alopecurus geniculatus are examples of far-creeping rhizomes (Greet et al. 2013). This root system also can protect sediment from resuspension and thus maintain a relative clear stable state during the first year (Barko et al. 1991).
With regards to the free-floating species in Y1, Lemna minor and Riccia fluitans are not able to develop during the dry year. But their small size, high growth rates, and their dispersion ability by wind or waterbirds and mammals from adjacent filled ponds help them to colonize the pond quickly after water refilling. L. minor and R. fluitans produce more and bigger propagules (Willby 2000) than other free-floating species found in our complete dataset like Spirodela polyrhiza, Lemna gibba and Azolla sp., leading to better resistance to a dry year. This can explain their quicker establishment in Y1.
Resilience of the plant community to severe drought
Based on the alternative states cycle found in our results and hypothesized in resilience theory (Fig. 3), state A appears as a renewal environment with high species richness. It mainly supports ruderal or pioneers species with a high abundance. These species enhance functional diversity of tolerant species that take advantage of the newly opened area, as explained by Šumberová et al. (2021). This high species richness offers a diversity that can react differently to a disturbance (Schulze 1996) and provides a panel of reactions for adaptation. We have discussed the different adaptation strategies of Y1 species facing drought disturbance. The community of representative plants of Y1 appears to be a functional group adapted to facing strong changes and its high resilience is likely to preserve chances of reactions (Holling 1987). This resilience also assures perennial continuity of the communities through time by succession of plants with similar roles (Pelletier et al. 2020).
From a resilience point of view, the dry period can be understood as a period of creative destruction (Gunderson 2001; Holling 1987; Scheffer and Carpenter 2003). It regulates the evolution of the ecosystem by breaking the climax state and bringing renewal (Y1). The phenomenon generates positive benefits in relation to species richness by causing enough pressure on the system to disrupt dominances, loss of resources availability and low diversity, which appears after Y3. The results from analysis of indicator species in plants communities show functional richness and more uncommon species present after the dry period, thus indicating a positive effect of the dry period on the pond ecosystem. This finding is corroborated by other studies which found rare species the following years after a drought (Collinson et al. 1995; Engelhardt 2006) As an example, this pond bed air exposure facilitates Charophytes development as Nitella sp. with some species at risk of extinction in Europe (Auderset and Boissezon 2018). The influence of regular dry periods on a pond’s ecosystem can thus be seen as a necessary and beneficial pressure for the development of aquatic plant community. A regular drying as a human management practice maintains the ecosystem in a long-term functional equilibrium. More generally, as demonstrated by Vanacker et al. (2015) or Phillips et al. (2019), the use of agro-ecological practices to manage fish pond landscapes in Europe, which are also often Natura 2000 zones, should be seriously considered for biodiversity conservation.
Conclusion
The present study investigated the influence of dry periods on aquatic plant community structure in fish ponds. The results show a drastic change in the plant communities during the first year after a dry period. Species richness is highest in this year and composed of many stress-resistant species, then declining with consecutive years. In consecutive years, species richness declines, although the dynamics of plant communities is linked to the phenomenon of nestedness based on a loss of several species but not on a turnover and most of the first-year species are only present in this year. The present study concludes that dry periods can be beneficial for a new dynamic to fish pond plant communities, with year one species characterized by a strategy of competitive, fast colonizing and disturbance-tolerant traits.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Change history
15 July 2023
A Correction to this paper has been published: https://doi.org/10.1007/s10452-023-10047-5
References
Arthaud F, Mousset M, Vallod D, Robin J, Wezel A, Bornette G (2012a) Effect of light stress from phytoplankton on the relationship between aquatic vegetation and the propagule bank in shallow lakes: effect of light stress on propagule bank. Fresh Biol 57:666–675. https://doi.org/10.1111/J1365-2427.2011.02730
Arthaud F, Vallod D, Robin J, Bornette G (2012b) Eutrophication and drought disturbance shape functional diversity and life-history traits of aquatic plants in shallow lakes. Aqua Sci 74:471–481. https://doi.org/10.1007/S00027-011-0241-4
Arthaud F, Vallod D, Robin J, Wezel A, Bornette G (2013) Short-term succession of aquatic plant species richness along ecosystem productivity and dispersal gradients in shallow lakes. J Veg Sci 24:148–156. https://doi.org/10.1111/j.1654-1103.2012.01436.x
Asaeda T, Van Bon T (1997) Modelling the effects of macrophytes on algal blooming in eutrophic shallow lakes. Eco Mod 104:261–287. https://doi.org/10.1016/S0304-3800(97)00129-4
Auderset Joye D, Boissezon A (2018) New insights into the ecology and phenology of two Characeae: N. opaca (Bruzelius) C. Agardh and N. gracilis (Sm.) C. Agardh Bot Letters 165:91–102. https://doi.org/10.1080/23818107.2017.1365259
Barko JW, Gunnison D, Carpenter SR (1991) Sediment interactions with submersed macrophyte growth and community dynamics. Aquat Bot 41:41–65. https://doi.org/10.1016/0304-3770(91)90038-7
Baselga A (2010) Partitioning the turnover and nestedness components of beta diversity. Glob Ecol Biog 19:134–143
Beklioglu M, Moss B (1996) Existence of a macrophyte-dominated clear water state over a very wide range of nutrient concentrations in a small shallow lake. Hydrobiol 337:93–106. https://doi.org/10.1007/BF00028510
Bini LM, Thomaz SM, Murphy KJ, Camargo AFM (1999) Aquatic macrophyte distribution in relation to water and sediment conditions in the Itaipu Reservoir, Brazil. Biology, ecology and management of aquatic plants. Springer, Netherlands, pp 147–154. https://doi.org/10.1007/978-94-017-0922-4_21
Caria MC, Grillas P, Rivieccio G, Bagella S (2021) Effect of inter-annual meteorological fluctuations and wild boar disturbance on the vegetation of Mediterranean temporary ponds. Appl Veg Sci 24:e12624. https://doi.org/10.1111/avsc.12624
Collinson NH, Biggs J, Corfield A, Hodson MJ, Walker D, Whitfield M, Williams PJ (1995) Temporary and permanent ponds: an assessment of the effects of drying out on the conservation value of aquatic macroinvertebrate communities. Biol Conserv 74:125–133. https://doi.org/10.1016/0006-3207(95)00021
Crawford SA (1977) Chemical, physical and biological changes associated with Chara succession in farm ponds. Hydrobiol 55:209–217. https://doi.org/10.1007/BF00017552
De Cáceres M, Legendre P (2009) Associations between species and groups of sites: indices and statistical inference. Ecol 90:3566–3574. https://doi.org/10.1890/08-1823.1
Engelhardt KAM (2006) Relating effect and response traits in submersed aquatic macrophytes. Ecol Appl 16:1808–1820. https://doi.org/10.1890/1051-0761
Fath BD, Dean CA, Katzmair H (2015) Navigating the adaptive cycle: an approach to managing the resilience of social systems. Ecol Soc 20:24. https://doi.org/10.5751/ES-07467-200224
Garbey C, Thiébaut G, Muller S (2004) Morphological plasticity of a spreading aquatic macrophyte, Ranunculus peltatus, in response to environmental variables. Plant Ecol 173:125–137. https://doi.org/10.1023/B:VEGE.0000026336.44576.ff
Greet J, Cousens RD, Webb JA (2013) Seasonal timing of inundation affects riparian plant growth and flowering: implications for riparian vegetation composition. Plant Ecol 217:87–101. https://doi.org/10.1007/s11258-012-0148-8
Gunderson LHP (2001) Understanding transformations in human and natural systems. Island Press, Washington
Hancz C, Nagy Z, Gal D, Varga D (2015) Issues of ecological and economical sustainability of fish culture in the southern hydrological basin of lake Balaton. Acta Agrar Kaposvár 19:25–29
Hilt S, Gross EM (2008) Can allelopathically active submerged macrophytes stabilise clear-water states in shallow lakes? Bas Appl Ecol 9:422–432. https://doi.org/10.1016/j.baae.2007.04.003
Holling CS (1987) Simplifying the complex: the paradigms of ecological function and structure. Europ J Oper Res 30:139–146. https://doi.org/10.1016/0377-2217(87)90091-9
Humphries P, Baldwin DS (2003) Drought and aquatic ecosystems: an introduction. Fresh Biol 48:1141–1146. https://doi.org/10.1046/j.1365-2427.2003.01092.x
Kelleway JJ, Iles JA, Kobayashi T, Ling JE (2020) Resilience of a native soil seed bank in a floodplain lake subjected to cropping, grazing and extended drought. Marine and Fresh Res 72:787–799. https://doi.org/10.1071/MF19386
Körner S, Nicklisch A (2002) Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes. J Phycol 38:862–887. https://doi.org/10.1046/j.1529-8817.2002.t01-1-02001.x
Lake PS (2003) Ecological effects of perturbation by drought in flowing waters. Fresh Biol 48:1161–1172. https://doi.org/10.1046/j.1365-2427.2003.01086.x
Lewis-Phillip J, Brook S, Sayer CD, McCrea R, Siriwardena G, Axmacher JC (2019) Pond management enhances the local abundance and species richness of farmland bird communities. Agricult Ecos Env 273:130–140. https://doi.org/10.1016/j.agee.2018.12.015
Magnus R, Rannap R (2019) Pond construction for threatened amphibians is an important conservation tool, even in landscapes with extant natural water bodies. Wetlands Ecol Manage 27:323–341. https://doi.org/10.1007/s11273-019-09662-7
Mari E, Santi E, Piazzini S, Renzi M, Maccherini S (2010) Development of biological diversity in farmland ponds. J Fresh Ecol 25:95–105. https://doi.org/10.1080/02705060.2010.9664362
Matsuzaki SS, Lathrop RC, Carpenter SR, Walsh JR, Vander Zanden MJ, Gahler MR, Stanley EH (2020) Climate and food web effects on the spring clear-water phase in two north-temperate eutrophic lakes. Limnol Oceanogr. https://doi.org/10.1002/lno.11584
Mccreary NJ (1991) Competition as a mechanism of submersed macrophyte community structure. Aquat Bot 41:177–193. https://doi.org/10.1016/0304-3770(91)90043-5
Murphy KJ, Rørslett B, Springuel I (1990) Strategy analysis of submerged lake macrophyte communities: an international example. Aquat Bot 36:303–323. https://doi.org/10.1016/0304-3770(90)90048-P
Netten JJC, Van Zuidam J, Kosten S, Peeters ETHM (2011) Differential response to climatic variation of free-floating and submerged macrophytes in ditches: macrophyte response to climate. Fresh Biol 56:1761–1768. https://doi.org/10.1111/j.1365-2427.2011.02611.x
O’farrell I, Izaguirre I, Chaparro G, Unrein F, Sinistro R, Pizarro H, Rodríguez P, De Tezanos PP, Lombardo R, Tell G (2011) Water level as the main driver of the alternation between a free-floating plant and a phytoplankton dominated state: a long-term study in a floodplain lake. Aquat Sci 73:275–287. https://doi.org/10.1007/s00027-010-0175-2
Oertli B, Frossard P A (2013) Mares et étangs : Ecologie, gestion, aménagement et valorisation. PPUR, Lausanne
Pelletier MC, Ebersole J, Mulvaney K et al (2020) Resilience of aquatic systems: review and management implications. Aquat Sci 82:44. https://doi.org/10.1007/s00027-020-00717-z
Qiu D, Wu Z, Liu B, Deng J, Fu G, He F (2001) The restoration of aquatic macrophytes for improving water quality in a hypertrophic shallow lake in Hubei Province, China. Ecol Engin 18:147–156. https://doi.org/10.1016/S0925-8574(01)00074-X
R Development Core Team (2010): R: A language and environment for statistical computing. Computer programme. http://www.R-project.org/
Robin J, Wezel A, Bornette G, Arthaud F, Angélibert S, Rosset V, Oertli B (2014) Biodiversity in eutrophicated shallow lakes: determination of tipping points and tools for monitoring. Hydrobiol 723:63–75. https://doi.org/10.1007/s10750-013-1678-3
Sandi SG, Rodriguez JF, Saintilan N (2020) Resilience to drought of dryland wetlands threatened by climate change. Sci Rep 10:13232. https://doi.org/10.1038/s41598-020-70087-x
Sarkar R, Ghosh AR, Mondal NK (2020) Comparative study on physicochemical status and diversity of macrophytes and zooplanktons of two urban ponds of Chandannagar, WB. India Appl Water Sci 10:63. https://doi.org/10.1007/s13201-020-1146-y
Sarremejane R, England J, Sefton CEM, Parry S, Eastman M, Stubbington R (2020) Local and regional drivers influence how aquatic community diversity, resistance and resilience vary in response to drying. Oikos 129:1877–1890. https://doi.org/10.1111/oik.07645
Sayer CD, Greaves HM (2019) Making an impact on UK farmland pond conservation. Aquat Ecos Marine and Fresh Ecos. https://doi.org/10.1002/aqc.3375
Scheffer M, Carpenter SR (2003) Catastrophic regime shifts in ecosystems: linking theory to observation. Trends Ecol Evol 18:648–656. https://doi.org/10.1016/j.tree.2003.09.002
Scheffer M, Van Nes EH (2007) Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiol 584:455–466. https://doi.org/10.1007/s10750-007-0616-7
Scheffer M, Hosper SH, Meijer ML, Moss B, Jeppesen E (1993) Alternative equilibria in shallow lakes. Trends Ecol Evol 8:175–279
Schulze P (1996) Engineering within ecological constraints. National Academies Press, Washington. https://doi.org/10.17226/4919
Šumberová K, Kučerová A, Šorf M, Grill S, Exler N, Vrba J (2021) Drivers of plant species composition of ecotonal vegetation in two fishpond management types. Wetlands Ecol Manage 29(1):93–110. https://doi.org/10.1007/s11273-020-09770-9
Toivonen H, Huttunen P (1995) Aquatic macrophytes and ecological gradients in 57 small lakes in southern Finland. Aquat Bot 51:197–221. https://doi.org/10.1016/0304-3770(95)00458-C
Van Den Berg MS, Coops H, Simons J (2001) Propagule bank buildup of Chara aspera and its significance for colonization of a shallow lake. Hydrobiol 462:9–17. https://doi.org/10.1023/A:1013125603555
Van der Maarel E (2007) Transformation of cover-abundance values for appropriate numerical treatment-Alternatives to the proposals by Podani. J Veg Sci 18:767–770. https://doi.org/10.1111/j.1654-1103.2007.tb02592.x
Van Donk E, Gulati RD, Iedema A, Meulemans JT (1993) Macrophyte-related shifts in the nitrogen and phosphorus contents of the different trophic levels in a biomanipulated shallow lake. Hydrobiol 251:19–26. https://doi.org/10.1007/BF00007160
Vanacker M, Wezel A, Payet V, Robin J (2015) Determining tipping points in aquatic ecosystems: the case of biodiversity and chlorophyll-a relations in fish pond systems. Ecol Indic 52:184–193. https://doi.org/10.1016/j.ecolind.2014.12.011
Wezel A, Robin J, Guerin M, Arthaud F, Vallod D (2013) Management effects on water quality, sediments and fish production in extensive fish ponds in the Dombes region, France. Limnol Ecol Manag Inland Wat 43:210–218. https://doi.org/10.1016/j.limno.2012.11.003
Wikum DA, Shanholtzer GF (1978) Application of the Braun-Blanquet cover-abundance scale for vegetation analysis in land development studies. Environ Manag 2:323–329. https://doi.org/10.1007/BF01866672
Wildová R, Wild J, Herben T (2007) Fine-scale dynamics of rhizomes in a grassland community. Ecogr 30:264–276. https://doi.org/10.1111/j.0906-7590.2007.04756.x
Willby NJ, Abernethy VJ, Demars BOL (2000) Attribute-based classification of European hydrophytes and its relationship to habitat utilization. Fresh Biol 43:43–74. https://doi.org/10.1046/j.1365-2427.2000.00523.x
Zamora-Marín JM, Ilg C, Demierre E, Bonnet N, Wezel A, Robin J, Vallod D, Calvo JF, Oliva-Paterna FJ, Oertli B (2021) Contribution of artificial waterbodies to biodiversity: a glass half empty or half full? Sci Tot Env 753:141987. https://doi.org/10.1016/j.scitotenv.2020.141987
Zealand AM, Jeffries MJ (2009) The distribution of pond snail communities across a landscape: separating out the influence of spatial position from local habitat quality for ponds in south-east Northumberland, UK. Hydrobiol 632:177–187. https://doi.org/10.1007/s10750-009-9837-2
Zhang XK, Liu XQ, Yang DI, Wang ZI (2019) Restoration of aquatic plants after extreme flooding and drought: a case study from Poyang Lake National Nature Reserve. Appl Ecol Env Res 17:15657–15668. https://doi.org/10.15666/aeer/1706_1565715668
Zhao F, Fang X, Zhao Z, Chai X (2021) Effects of water level fluctuations on the growth characteristics and community succession of submerged macrophytes: a case study of Yilong lake. China Water 13:2900. https://doi.org/10.3390/w13202900
Acknowledgements
The project has been partially funded by Auvergne Rhône Alpes Region, France. The authors wish to thank Thomas Lhuillery, Julie Pedrono and Sylvie Prestoz for their assistance on the field and in the laboratory. This publication is the result of many years of field works supported by ISARA team. A special thanks to Fondation Vérots for giving their consent for access to their ponds. Appreciation is also expressed to the fish farmers who made possible the present study
Funding
Partial financial support was received from projects funded by Auvergne-Rhône-Alpes Region (public institution).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Communicated by Télesphore Sime-Ngando.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Fontanilles, A., Wezel, A., Rouifed, S. et al. Disturbance and resilience of aquatic plant communities in fish ponds after temporary dry periods. Aquat Ecol 57, 597–609 (2023). https://doi.org/10.1007/s10452-023-10032-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-023-10032-y