
Abstract
Purpose
To determine whether P7C3-A20, a proneurogenic neuroprotective agent, can protect the retinal ganglion cells (RGCs) of rats from optic nerve crushing.
Methods
The left optic nerve of 67 rats was crushed, and 5.0 mg/kg/day of P7C3-A20 (crush-P7C3) or its vehicle (crush-placebo) was injected intraperitoneally for 3 days from one day prior to the crushing. The protective effects were determined by the number of Tuj-1-stained RGCs and by the ratio of the mRNA levels of BAX/Bcl-2 on day 7. The levels of NAD and NAD-related genes were also determined.
Results
The density of RGCs was 2009.4 ± 57.7 cells/mm2 in the sham controls; it was significantly lower in the crush-placebo group at 979.7 ± 144.3 cells/mm2 (P < 0.0001). The neuroprotective effects of P7C3-A20 was demonstrated by the significantly higher density of 1266.0 ± 193.1 cells/mm2 than in the crush-placebo group (P = 0.01, Scheffe). After crushing the optic nerve the BAX/Bcl-2 ratio was higher in the optic nerves and retina, application of P7C3-A20 significantly reduced this ratio. P7C3-A20 significantly increased the NAD level in the untouched optic nerves from 1.36 ± 0.05 to 1.59 ± 0.10 nmol/mg protein (P = 0.02, t test). Crushing the optic nerve decreased the level to 1.27 ± 0.21 nmol/mg protein and P7C3-A20 preserved the level at 1.43 ± 0.10 nmol/mg protein. Crushing the optic nerve decreased the mRNA levels of Nampt and Sirt-1 in the optic nerves, while P7C3-A20 significantly restored the levels.
Conclusions
P7C3-A20 can protect RGCs from optic nerve crushing possibly through preserving the NAD levels in the optic nerves.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The optic nerve degenerates following the infliction of various types of insults including inflammation, trauma, and ischemia. Under these conditions, the axons degenerate primarily distally, i.e., away from the cell bodies, from the injured site, a process known as Wallerian degeneration. In the optic nerve, the degeneration also spreads to the retinal ganglion cells (RGCs) through a condition known as retrograde degeneration. The degenerative process, especially Wallerian degeneration, is believed to be caused mainly by the disruption of axonal transport from the cell bodies and is thus considered to be a passive and progressive process.
The results of studies on Wallerian degeneration slow (Wlds) mice suggest that this process is not necessarily passive [1], and some molecular mechanisms may be actively involved in the degeneration [2]. If this is correct then it may be possible to target the mechanisms by medications.
The wlds gene encodes nicotinamide mononucleotide adenylyltransferase type 1 (NMNAT-1) which is involved in the formation of the nicotinamide adenine dinucleotide (NAD) salvage pathway [3, 4]. DBA/2J mice have progressive eye abnormalities that closely mimic human hereditary glaucoma, and when they are backcrossed with Wlds mice, the offspring are resistant to glaucomatous optic neuropathy [5]. These results combined with other findings [6, 7] suggest that preserving the NAD levels should delay axonal degeneration and prevent optic nerve atrophy from various insults.
P7C3 belongs to a class of aminopropyl carbazole chemicals discovered through comprehensive in vivo screening of neurogenesis [8]. It was established that this chemical binds and activates nicotinamide phosphoribosyltransferase (NAMPT) [9], a self-limiting enzyme involved in the NAD salvage pathway. This binding may enhance the biogenesis of NAD. P7C3 has been shown to be neuroprotective against traumatic brain injuries [10, 11] and neurodegeneration in experimental animal models [12, 13].
The purpose of this study was to determine whether P7C3-A20, an active variant of P7C3 [9], can protect the RGCs from retrograde degeneration after the optic nerve of rats is crushed. Either P7C3-A20 or its vehicle (DMSO) was injected intraperitoneally for 3 days from one day prior to the crushing, and their effects were determined by counting the immunohistochemically stained RGCs on day 7 post-crushing. We also determined the BAX/Bcl-2 ratio in the retina and the optic nerve by real-time PCR on day 7. In addition, we determined the NAD levels in the retina and the optic nerve using a NAD/NADH kit (Abcam, Cambridge, UK).
Methods
Animals
Nine-week-old, male Wistar rats were purchased from Japan SLC (Shizuoka, Japan) and housed in an air-conditioned room with a temperature of approximately 23 °C and humidity of 60%. The room lights were on a 12:12 light:dark cycle. All animals were handled in accordance with the ARVO Resolution for the Use of Animals in Ophthalmic and Vision Research. The experimental protocol was approved by the Committee of Animal Use and Care of the Osaka Medical College (No. 28024). A total of 67 rats was used.
Chemicals
Unless noted, all chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA). P7C3-A20, an active variant of P7C3 [9], was purchased from AdooQ Bioscience (Irvine, CA, USA). P7C3 at ≥5 mg/kg has proneurogenic activity in rats [8]. We chose a concentration of 5 mg/kg of P7C3-A20 for these experiments. It has been shown that P7C3 can cross the blood–brain barrier [8].
Anesthesia and euthanasia
All surgery was performed under general anesthesia by an intraperitoneal injection of a mixture of medetomidine (0.75 mg), midazolam hydrochloride (4.0 mg), and butorphanol tartrate (5.0 mg/kg body wt). Rats were euthanized by exposure to CO2 at a rate of 6 L/min in a cage (13.8 L) with wood-shaving bedding.
Optic nerve crushing
Animals were anesthetized as described, and an incision was made along the midline of the skull to expose the superior surface of the left eye. The superior rectus muscle was incised to expose the left optic nerve, and the left optic nerve 2 mm behind the eye was crushed with forceps for 10 s [14]. Care was taken not to occlude the blood vessels and cause retinal ischemia. We confirmed by indirect ophthalmoscopy that the retinal circulation was not blocked and verified this by real-time PCR, demonstrating that the HIF-1α gene was not up-regulated [15]. Specifically, the mean ± SD level of the mRNA of the retinal HIF-1α gene determined on day 7 after crushing the optic nerve was 1.04 ± 0.9 fold (range 0.95–1.23) of that of the control.
Animals received intraperitoneal injections of 5.0 mg/kg/day of P7C3-A20 or its vehicle (100% DMSO, 100 µl) for 3 days from one day prior to surgery. As control, a sham operation was performed on the left eyes of other animals by exposing the optic nerve in the same way but the nerve was not crushed. The right eyes were untouched in either type of rats.
Labeling retinal ganglion cells
Following the crushing of the optic nerve the loss of RGCs occurs in a delayed fashion [16, 17]. The number of RGCs remains unchanged for 5 days and then abruptly decreases by 50% on day 7 and to less than 10% on day 14 [16]. The death of the RGCs is mainly due to apoptosis [17].
To study the effects of crushing the optic nerves, rats were killed on day 7 and the retinas were carefully removed from the eyes as described [18]. In brief, rats were euthanized by CO2, and the globe was proptosed by placing forceps around the optic nerve just behind the eyeball. The globe was transected along the equator and the cornea and lens were removed. The retina was detached from the pigment epithelium by pressing upward with forceps and removed by cutting its attachment to the optic nerve head. The isolated retina was immediately placed in phosphate buffered saline (PBS) solution, and any vitreous remaining on the isolated retina was carefully removed.
The isolated retinas were then sandwiched between nylon mesh sheets (N0255HD, NBC Meshtec, Tokyo, Japan) and fixed in 4% PFA in PBS overnight at 4 °C. After washing in PBS and blocking in PBS containing 1.0% BS and 0.3% triton X-100, the retinas were incubated with Alexa 488-conjugated mouse monoclonal neuron-specific class III beta-tubulin antibody (Tuj-1, 1:500; Covance, Princeton, New Jersey, USA). Tuj-1 is a specific marker for RGCs [19]. The retinas were placed in the same medium overnight at 4 °C, washed with PBS, and cover slipped the next morning.
To determine the number of RGCs, the stained flat mounts were photographed with a fluorescent microscope (BZ X700, Keyence, Osaka, Japan). Eight areas (0.48 × 0.48 mm) from the 4 quadrants of the retina at a distance of 1.0 and 1.5 mm from the margin of the optic disc were photographed. All of the Tuj-1-positive cells in an area of 0.2 × 0.2 mm at the center of each image were counted using the NIH ImageJ program.
The mean density of the RGC/mm2 was calculated, and the loss of RGCs was determined by comparing the density in the retinas of animals receiving P7C3-A20 or its vehicle after crushing the optic nerve to that of retinas from sham operated optic nerves (n = 5–7 for each group). The number of RGCs was counted by one observer (SM) who was masked as to whether it was from an experimental or a sham animal.
Changes in expression of Bax, Bcl-2, and NAD-related genes in retina and optic nerve
The ratio of Bax/Bcl-2 is an indicator of neuronal death by apoptosis [20]. We determined the changes in the expression of the Bax and Bcl-2 genes in the retina and the optic nerve by real-time PCR (RT-PCR) on day 7 after the optic nerve was crushed. In addition, we determined the expression of the NAD-related genes, Nampt, Nmnat-1, and Sirt-1, by RT-PCR. For this, retinas were obtained as described, and a 4 mm length of the optic nerve with the crushed site at the center was excised. The tissues were homogenized in lysis buffer, and the total RNA was extracted using a NucleoSpin RNA kit (TakaRa, Ohtsu, Shiga, Japan). The quality and quantity of the RNAs were assessed with a BioSpectrometer (Eppendorf, Hamburg, Germany).
The purified RNA was then reverse transcribed with the PrimeScript RT reagent and gDNA Eraser kits according to the manufacturer’s instructions (TaKaRa). Quantitative real-time PCR analysis was performed with the Thermal Cycler Dice Real Time System, TP870 (TaKaRa) Premix Ex Taq (Probe qPCR; TaKaRa), and the TaqMan Gene Expression Assays for the targeted genes (Applied Biosystems, Foster City, CA, USA). The rat TaqMan Gene Assays for BAX Rn99999125_m1, Bcl2 Rn02532682_g1, NAMPT Rn00822043_m1, NMNAT1 Rn01516826_m1, and SIRT1 Rn01428096_m1 were used. The amplicons were detected with the relevant probes tagged with a MGB quencher and a FAM dye. All reactions were run with the following cycling parameters; 30 s at 95 °C followed by 40 cycles at 95 °C for 5 s and 60 °C for 30 s. A standard curve of the cycle thresholds was constructed using serial dilutions of the cDNA samples. The relative quantities of mRNAs were calculated with 18 s (Hs99999901_s1) serving as an internal control.
Concentration of NAD in optic nerve and retina
Because it has been shown that the neuroprotective effects of P7C3 are mediated in part through the activation of NAD synthesis [9], the NAD level was measured in the optic nerves and retinas on day 7 with the NAD/NADH kit (Abcam) according to the manufacturer’s instruction. Briefly, excised tissues were homogenized in 400 μL of NADH/NAD extraction buffer supplied with the kit. After centrifugation (15,000 g), the supernatant was divided into 2 sets. One-half of the supernatant was incubated at 60 °C for 30 min to decompose the NAD (containing only NADH). The other half was kept on ice, and the sample was composed of both NAD and NADH (total NAD). Then, 50 μL of each sample was placed in 96-well culture plates and 100 μL of reaction mix (supplied in the kit) was added to each well. After converting NAD to NADH with the developing agent, the absorbance at 450 nm was measured with a microplate reader (SH-100 Lab, Corona, Ibaraki, Japan). The value of NAD was calculated by subtracting NADH from the total NAD, and the levels were normalized by the protein concentration determined by Bio-Rad protein assay (Bio-Rad, Hercules, CA, USA).
Statistical analyses
The data are presented as the means ± standard deviations (SDs). Statistical analyses were done by one-way analysis of variance (ANOVA), and if significant changes were detected, an appropriate post hoc test was performed for statistical comparisons among groups. Student t tests were used to determine the significance of differences between the 2 groups. The level of significance was set at P < 0.05.
Results
Decrease in number of RGCs after optic nerve crush
Representative photomicrographs of flat-mounted retinas taken approximately 1.0 mm from the optic disc margin are shown in Fig. 1a and at higher magnification in Fig. 1b. The RGCs are stained green with Alexa 488-conjugated Tuj-1, and it can clearly be seen that the number of RGCs is reduced after the optic nerve was crushed (crush-placebo). There appears to be a greater number of surviving RGCs after crushing the optic nerve in animals that received intraperitoneal injections of P7C3-A20 (5.0 mg/kg/day × 3 days; crush-P7C3).

Representative photomicrographs of flat mounted retinas stained with Alexa 488-conjugated Tuj-1 antibody demonstrating the retinal damages on day 7 after crushing the optic nerve. a Retina from sham control after crushing the optic nerve (left) and injected intraperitoneally with vehicle (middle), and retina after crushing the optic nerve and injecting P7C3-A20 intraperitoneally (right panel). Photographs were taken 1.0 mm from the optic disc margin. Bar = 100 µm. b Images shown in Fig. 1a are presented at higher magnification. Bar = 100 µm. c Density of retinal ganglion cells (RGCs/mm2). There is a significant reduction in the number of RGCs after crushing the optic nerve (crush-placebo). An injection of P7C3-A20 (crush-P7C3) depressed the reduction. Data are shown as the means ± standard deviations (SDs). (control n = 5, crush-placebo: n = 7, crush-P7C3: n = 5). (# P < 0.05, ## P < 0.01; Scheffe)
The mean ± SD of the number of RGCs stained by Tuj-1 antibody was 2009.4 ± 57.7 cells/mm2 in the sham operated rats (sham control, n = 5), and the mean number was decreased significantly to 979.7 ± 144.3 cells/mm2 (n = 7) on day 7 in the crush-placebo group (P < 0.0001; Scheffe; Fig. 1c). The reduction was significantly depressed to 1266.0 ± 193.1 cells/mm2 (n = 5) in the crush-P7C3 group (P = 0.01, Scheffe test; Fig. 1c).
Bax/Bcl-2 ratio
The effects of crushing of the optic nerve on the retina were also determined by the expressions of the Bax and Bcl-2 genes on day 7 by real-time PCR (Fig. 2). The mRNA level of Bax in the optic nerve was increased by 7.8 ± 1.8 fold from the sham group and the crush-placebo group (P < 0.0001, Scheffe test; n = 8), while the increase was depressed significantly by 4.05 ± 1.05 fold in animals treated with P7C3-A20 (P < 0.0001, Scheffe; n = 8; Fig. 2a). On the other hand, the mRNA level of Bcl-2 was decreased by 0.38 ± 0.13 fold in the crush-placebo group, while the level was maintained at significantly higher level of 0.72 ± 0.15 fold in the crush-P7C3 group (P = 0.0008, Scheffe). Thus, the ratio of Bax/Bcl-2 was 24.1 ± 11.9 fold in the crush-placebo group, while the level was significantly depressed by 6.06 ± 2.53 fold in the crush-P7C3 group (P = 0.0002, Scheffe).

Changes in the expression of Bax and Bcl-2 in the optic nerve (a) and retina (b). a The level of Bax is increased and Bcl-2 decreased in the optic nerve after the nerve is crushed (crush-placebo). These changes are significantly lower in animals treated with P7C3-A20 (crush-P7C3). Thus, P7C3-A20 significantly depressed the ratio of Bax/Bcl-2. Data are the means ± standard deviations (SDs). (control: n = 6, crush-placebo: n = 8, crush-P7C3: n = 8). (## P < 0.01, ### P < 0.001; Scheffe). b The BAX levels are similar among groups but P7C3-A20 (crush-P7C3) significantly increases the Bcl-2 levels, and significantly decreases the BAX/Bcl-2 ratio compared to that of the crush-induced level (crush-placebo). Data are the means ± standard deviations (SDs). (control: n = 6, crush-placebo: n = 8, crush-P7C3: n = 8). (# P < 0.05, ## P < 0.01, ### P < 0.001; Scheffe)
The Bax and Bc-l2 levels in the retinas are shown in Fig. 2b. In the retina, the mRNA levels of Bax in the experimental groups were not significantly different from those of the sham controls (P = 0.45, ANOVA). In contrast, the level of Bcl-2 was increased by 1.68 ± 0.34 fold from the sham control in animals treated with P7C3-A20 (crush-P7C3, P = 0.0005, Scheffe). Thus, the ratio of Bax/Bcl-2 was 1.29 ± 0.6 fold in the crush-placebo group, while it was significantly depressed at 0.72 ± 0.27 fold in the crush-P7C3 group (P = 0.03, Scheffe).
NAD concentration in retina and optic nerve
A plot of the NAD concentrations in the optic nerve and the retina on day 7 after the crushing is shown in Fig. 3. In the untouched optic nerve, the level of NAD was 1.36 ± 0.05 nmol/mg/protein (n = 4). An injection of P7C3-A20 significantly increased the level to 1.59 ± 0.10 nmol/mg/protein (P = 0.02, t test; n = 4; Fig. 3a). The NAD concentrations in the optic nerve which had been crushed was 1.27 ± 0.21 nmol/mg/protein (crush placebo, n = 4; Fig. 3a). This reduction from the untouched optic nerve (1.36 ± 0.05) was not significant (P = 0.44, t test; Fig. 3a). On the other hand, the NAD level was 1.43 ± 0.10 nmol/mg/protein in animals with their optic nerves crushed and injected with P7C3-A20 (crush-P7C3, n = 4; Fig. 3a). Thus, P7C3-A20 seemed to preserve the NAD levels in the crushed optic nerves, but the difference from the crush-placebo group was not significant (P = 0.14, t test; Fig. 3a).

Levels of nicotinamide adenine dinucleotide (NAD) in the optic nerve (a) and retina (b). a P7C3-A20 significantly increases the NAD concentration in the untouched optic nerve, and preserves the NAD level in the crushed optic nerve. Data are shown as the means ± standard deviations (SDs). (n = 4, each group; *P < 0.05, t test). b The NAD level in the retina is not significantly different in the different groups (P > 0.05, t test). Data are shown as the means ± standard deviations (SDs). (n = 4, each group)
In the retinas of untouched optic nerves, the level of NAD was 2.39 ± 0.30 nmol/mg/protein (n = 4), and in animals receiving P7C3-A20 the level was 2.46 ± 0.31 nmol/mg/protein (n = 4) (n = 4, Fig. 3b). These differences were not significant (P = 0.75, t test). The NAD concentration in the retinas of animals with their optic nerves crushed was 2.21 ± 0.32 nmol/mg/protein (crush-placebo, n = 4; Fig. 3b). The difference from the retinas with untouched optic nerves was not significant (P = 0.44, t test). The mean retinal NAD level was 2.36 ± 0.39 nmol/mg/protein in animals with their optic nerves crushed and treated by P7C3-A20 (crush-P7C3, n = 4; Fig. 3b). The difference in the NAD levels between animals treated with or without P7C3-A20 was not significant (P = 0.57, t test; Fig. 3b).
Changes in expression of Nampt, Nmnat-1, and Sirt-1 genes
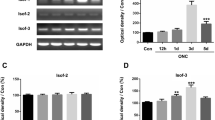
Because P7C3 is known to activate the NAMPT gene, the self-limiting enzyme of the NAD salvage pathway, we examined whether an injection of P7C3-A20 will change the mRNA levels of Nampt, Nmnat-1, and Sirt-1 genes by real-time PCR. The results of the optic nerves are shown in Fig. 4a, and those of the retinas are shown in Fig. 4b.

Changes in the expression of NAD-related genes in the optic nerve (a) and retina (b) determined by RT-PCR. a Crushing the optic nerve decreases the mRNA levels of nicotinamide phosphoribosyltransferase (Nampt) and Sirt-1, which is restored by P7C3-A20. The nicotinamide mononucleotide adenylyltransferase type 1 (Nmnat-1) levels are not significantly different in the different groups. Data are shown as the means ± standard deviations (SDs). (n = 4, each group). b Crushing the optic nerve decreases the mRNA levels of retinal Nampt. The retinal Sirt-1 level is significantly higher in the crush P7C3 group than in the crush placebo group. Data are shown as the means ± standard deviations (SDs). (*P < 0.05, **P < 0.01, ANOVA followed by Fisher; n = 4, each group)
In the optic nerve, the changes in the Nampt mRNA levels were significant (P = 0.0003, ANOVA; Fig. 4a) in the sham controls, crush-placebo, and crush optic nerve groups that received P7C3-A20. Crushing the optic nerve (crush-placebo, n = 4) significantly decreased the Nampt mRNA levels in the optic nerve by 0.60 ± 0.12 fold from the sham control (P = 0.001, Fisher; n = 4, Fig. 4a). The level was preserved by 0.83 ± 0.06 fold in the crush-P7C3 group (n = 4). The difference between the crush-placebo and crush-P7C3 groups was significant (P = 0.004, Fisher; Fig. 4a).
Changes in the Nampt levels in the retina were also significant (P = 0.04, ANOVA) among these groups (Fig. 4b). The level in the crush-placebo group was significantly lower by 0.76 ± 0.06 fold from the sham control (P = 0.04, Fisher), while that of the crush-P7C3 group was 1.0 ± 0.13 fold of the sham control. However, the difference between the crush-placebo and crush-P7C3 groups was not significant (P = 0.07, Fisher; Fig. 4b).
Changes in the Nmnat-1 levels among the groups were not significant either in the optic nerves (P = 0.8, Fig. 4a) or in the retinas (P = 0.08, ANOVA; Fig. 4b).
The mRNA levels of Sirt-1 were also determined because SIRT-1 is a NAD-dependent histone deacetylase [21]. Changes of the Sirt-1 levels were significant among the groups both in the optic nerves (P = 0.02, Fig. 4a) and the retinas (P = 0.02, ANOVA; Fig. 4b). Crushing the optic nerve (crush-placebo, n = 4) significantly decreased the mRNA level of Sirt-1 by 0.58 ± 0.09 fold from the sham control (Fig. 4a). The level was preserved by 0.84 ± 0.11 fold in the crush-P7C3 group (n = 4). The difference was also significant (P = 0.04, Fisher; Fig. 4a).
In the retina, crushing the optic nerve (crush-placebo, n = 4) decreased the retinal Sirt-1 level by 0.68 ± 0.15 fold from the sham control (Fig. 4b). The level was increased by 1.29 ± 0.41 fold from the sham control in animals with their optic nerves crushed and treated by P7C3 (n = 4). The difference between the sham control and the crush-placebo was not significant (P = 0.2), while the difference between the crush-placebo and the crush-P7C3 groups was significant (P = 0.04, Fisher; Fig. 4b).
Discussion
Our results show that 5 mg/kg/day of P7C3-A20 for 3 days had a neuroprotective effect on the RGCs after the optic nerve was crushed in rats. These effects were determined by counting the number of surviving RGCs, and determining the level of the BAX and Bcl-2 genes. In addition, P7C3-A20 maintained the NAD levels in the crushed optic nerves and restored the reduction of the Nampt and Sirt-1 genes after the optic nerve was crushed.
It has been shown that P7C3 activates NAMPT and increases the formation of nicotinamide mononucleotide (NMN) [9]. NAMPT has been shown to be a neuroprotective agent against neuronal ischemic injury [22]. NMNAT-1, which synthesizes NAD from NMN, has also been shown to be neuroprotective against excitotoxic injuries [23]. Thus, our finding of the preserved NAD levels in the optic nerve may be related to the neuroprotective properties of P7C3.
The higher mRNA levels of Nampt in animals in the crush-P7C3 group than in the crush-placebo group support the idea that the action of P7C3-A20 was directed at the NAMPT gene. Because the Nmnat-1 levels were not changed significantly, it is reasonable to consider that the increase in NMN resulted in an increase of NAD and the induction of the SIRT-1 gene. The absence of a reduction in the Nmnat-1 level in the crushed optic nerve is probably because NMNAT-1 is a nuclear protein [24], and thus a decline was seen in the retinal Nmnat-1 level.
NAD is known to play a crucial role in mitochondrial function [25], and its level is known to be decreased in degenerated axons [6]. While the subset of mitochondrial proteins is supplied by the cell body through axonal transport, mitochondrial function, including ATP synthesis, and their membrane potentials are maintained by locally synthesized proteins in the axons [26]. Although the mechanisms are still controversial [7], an overexpression of Wlds or NMNAT-1 can preserve the NAD levels and delay the axonal degeneration [6]. In addition, exogenously applied NAD can maintain the ATP levels and prevent axonal degeneration [7]. Thus, the neuroprotective effects of P7C3-A20 may be linked to local energy production by the mitochondria in the axons through the formation of NAD.
Crushed optic nerves generate the entry of extracellular Ca2+ leading to a rapid rise in intracellular Ca2+, which can then trigger the activation of calpain [27] and autophagy-mediated axonal degeneration [28]. These cascades triggered by the Ca2+ entry are the main molecular mechanisms that cause axonal degeneration [13]. Wang et al. [2] hypothesize that NAD is neuroprotective in these processes because it can control the opening of ion channels and enhance the ability of Ca2+ buffering by mitochondria [29].
It has been shown that an overexpression or pharmacological activation of the SIRT-1 gene is neuroprotective for RGCs from various injuries including optic nerve crush [30, 31] and optic neuritis [32]. SIRT-1 is known to be expressed in the RGCs [33], and also in astrocytes of the CNS [34]. SIRT-1 is also known to inhibit the action of pro-apoptotic and pro-inflammatory genes such as the p53 and NFκB genes through deacetylation [35]. In addition, SIRT-1 induces the expression of BDNF, a neurotrophic factor [36]. Thus, it is highly likely that SIRT-1 is involved in the neuroprotection by P7C3-A20 through the suppression of apoptosis of RGCs as well as neuro-inflammation and astrogliosis in the optic nerve. We suggest that P7C3-A20 may work against both the RGCs and their axons in different ways. A different expression pattern of NAD and NAD-related genes as well as the BAX and Bcl-2 genes between the optic nerve and the retina may support this suggestion.
Our findings indicate that P7C3-A20 probably has protective effects on the RGCs from optic nerve injury, but there are limitations to this study. One limitation is that we did not determine whether P7C3-A20 increased the protein levels and activities of NAMPT, NMNAT-1and SIRT-1. In addition, we did not determine the degree of axonal degeneration, or whether the inhibition of SIRT-1 abolished the effects of P7C3-A20. These issues need to be determined in future studies. However, it is reasonable to consider that the survival of the RGCs is highly likely to reflect the degree of axonal degeneration.
In conclusion, P7C3-A20 neuroprotects RGCs from traumatic optic nerve injuries. This chemical may be practically promising because it may be directed to both the RGCs and their axons.
References
Lunn ER, Perry VH, Brown MC, Rosen H, Gordon S. Absence of wallerian degeneration does not hinder regeneration in peripheral nerve. Eur J Neurosci. 1989;1:27–33.
Wang JT, Medress ZA, Barres BA. Axon degeneration: molecular mechanisms of a self-destruction pathway. J Cell Biol. 2012;196:7–18.
Mack TG, Reiner M, Beirowski B, Mi W, Emanuelli M, Wagner D, et al. Wallerian degeneration of injured axons and synapses is delayed by a Ube4b/Nmnat chimeric gene. Nat Neurosci. 2001;4:1199–206.
Conforti L, Wilbrey A, Morreale G, Janeckova L, Beirowski B, Adalbert R, et al. WldS protein requires Nmnat activity and a short N-terminal sequence to protect axons in mice. J Cell Biol. 2009;184:491–500.
Howell GR, Libby RT, Jakobs TC, Smith RS, Phalan FC, Barter JW, et al. Axons of retinal ganglion cells are insulted in the optic nerve early in DBA/2J glaucoma. J Cell Biol. 2007;179:1523–37.
Wang J, Zhai Q, Chen Y, Lin E, Gu W, McBurney MW, et al. A local mechanism mediates NAD-dependent protection of axon degeneration. J Cell Biol. 2005;170:349–55.
Sasaki Y, Vohra BP, Lund FE, Milbrandt J. Nicotinamide mononucleotide adenylyl transferase-mediated axonal protection requires enzymatic activity but not increased levels of neuronal nicotinamide adenine dinucleotide. J Neurosci. 2009;29:5525–35.
Pieper AA, Xie S, Capota E, Estill SJ, Zhong J, Long JM, et al. Discovery of a proneurogenic, neuroprotective chemical. Cell. 2010;142:39–51.
Wang G, Han T, Nijhawan D, Theodoropoulos P, Naidoo J, Yadavalli S, et al. P7C3 neuroprotective chemicals function by activating the rate-limiting enzyme in NAD salvage. Cell. 2014;158:1324–34.
Yin TC, Britt JK, De Jesus-Cortes H, Lu Y, Genova RM, Khan MZ, et al. P7C3 neuroprotective chemicals block axonal degeneration and preserve function after traumatic brain injury. Cell Rep. 2014;8:1731–40.
Dutca LM, Stasheff SF, Hedberg-Buenz A, Rudd DS, Batra N, Blodi FR, et al. Early detection of subclinical visual damage after blast-mediated TBI enables prevention of chronic visual deficit by treatment with P7C3-S243. Invest Ophthalmol Vis Sci. 2014;55:8330–41.
Tesla R, Wolf HP, Xu P, Drawbridge J, Estill SJ, Huntington P, et al. Neuroprotective efficacy of aminopropyl carbazoles in a mouse model of amyotrophic lateral sclerosis. Proc Natl Acad Sci U S A. 2012;109:17016–21.
De Jesus-Cortes H, Xu P, Drawbridge J, Estill SJ, Huntington P, Tran S, et al. Neuroprotective efficacy of aminopropyl carbazoles in a mouse model of parkinson disease. Proc Natl Acad Sci U S A. 2012;109:17010–5.
Kurimoto T, Ishii M, Tagami Y, Nishimura M, Miyoshi T, Tsukamoto Y, et al. Xylazine promotes axonal regeneration in the crushed optic nerve of adult rats. Neuroreport. 2006;17:1525–9.
Tonari M, Kurimoto T, Horie T, Sugiyama T, Ikeda T, Oku H. Blocking endothelin-B receptors rescues retinal ganglion cells from optic nerve injury through suppression of neuroinflammation. Invest Ophthalmol Vis Sci. 2012;53:3490–500.
Berkelaar M, Clarke DB, Wang YC, Bray GM, Aguayo AJ. Axotomy results in delayed death and apoptosis of retinal ganglion cells in adult rats. J Neurosci. 1994;14:4368–74.
Isenmann S, Wahl C, Krajewski S, Reed JC, Bahr M. Up-regulation of bax protein in degenerating retinal ganglion cells precedes apoptotic cell death after optic nerve lesion in the rat. Eur J Neurosci. 1997;9:1763–72.
Winkler BS. The electroretinogram of the isolated rat retina. Vis Res. 1972;12:1183–98.
Snow RL, Robson JA. Migration and differentiation of neurons in the retina and optic tectum of the chick. Exp Neurol. 1995;134:13–24.
Schelman WR, Andres RD, Sipe KJ, Kang E, Weyhenmeyer JA. Glutamate mediates cell death and increases the bax to bcl-2 ratio in a differentiated neuronal cell line. Brain Res Mol Brain Res. 2004;128:160–9.
Dvir-Ginzberg M, Gagarina V, Lee EJ, Hall DJ. Regulation of cartilage-specific gene expression in human chondrocytes by SIRT1 and nicotinamide phosphoribosyltransferase. J Biol Chem. 2008;283:36300–10.
Zhao Y, Liu XZ, Tian WW, Guan YF, Wang P, Miao CY. Extracellular visfatin has nicotinamide phosphoribosyltransferase enzymatic activity and is neuroprotective against ischemic injury. CNS Neurosci Ther. 2014;20:539–47.
Verghese PB, Sasaki Y, Yang D, Stewart F, Sabar F, Finn MB, et al. Nicotinamide mononucleotide adenylyl transferase 1 protects against acute neurodegeneration in developing CNS by inhibiting excitotoxic-necrotic cell death. Proc Natl Acad Sci U S A. 2011;108:19054–9.
Berger F, Lau C, Dahlmann M, Ziegler M. Subcellular compartmentation and differential catalytic properties of the three human nicotinamide mononucleotide adenylyltransferase isoforms. J Biol Chem. 2005;280:36334–41.
Stein LR, Imai S. The dynamic regulation of nad metabolism in mitochondria. Trends Endocrinol Metab. 2012;23:420–8.
Kaplan BB, Gioio AE, Hillefors M, Aschrafi A. Axonal protein synthesis and the regulation of local mitochondrial function. Results Probl Cell Differ. 2009;48:225–42.
Saatman KE, Abai B, Grosvenor A, Vorwerk CK, Smith DH, Meaney DF. Traumatic axonal injury results in biphasic calpain activation and retrograde transport impairment in mice. J Cereb Blood Flow Metab. 2003;23:34–42.
Knoferle J, Koch JC, Ostendorf T, Michel U, Planchamp V, Vutova P, et al. Mechanisms of acute axonal degeneration in the optic nerve in vivo. Proc Natl Acad Sci USA. 2010;107:6064–9.
Tamsett TJ, Picchione KE, Bhattacharjee A. NAD+ activates KNa channels in dorsal root ganglion neurons. J Neurosci. 2009;29:5127–34.
Zuo L, Khan RS, Lee V, Dine K, Wu W, Shindler KS. SIRT1 promotes RGC survival and delays loss of function following optic nerve crush. Invest Ophthalmol Vis Sci. 2013;54:5097–102.
Kim SH, Park JH, Kim YJ, Park KH. The neuroprotective effect of resveratrol on retinal ganglion cells after optic nerve transection. Mol Vis. 2013;19:1667–76.
Shindler KS, Ventura E, Rex TS, Elliott P, Rostami A. SIRT1 activation confers neuroprotection in experimental optic neuritis. Invest Ophthalmol Vis Sci. 2007;48:3602–9.
Ozawa Y, Kubota S, Narimatsu T, Yuki K, Koto T, Sasaki M, et al. Retinal aging and sirtuins. Ophthalmic Res. 2010;44:199–203.
Scuderi C, Stecca C, Bronzuoli MR, Rotili D, Valente S, Mai A, et al. Sirtuin modulators control reactive gliosis in an in vitro model of Alzheimer’s disease. Front Pharmacol. 2015;5:89. doi:10.3389/fphar.2014.00089.
Hernandez-Jimenez M, Hurtado O, Cuartero MI, Ballesteros I, Moraga A, Pradillo JM, et al. Silent information regulator 1 protects the brain against cerebral ischemic damage. Stroke. 2013;44:2333–7.
Jeong H, Cohen DE, Cui L, Supinski A, Savas JN, Mazzulli JR, et al. Sirt1 mediates neuroprotection from mutant huntingtin by activation of the TORC1 and CREB transcriptional pathway. Nat Med. 2011;18:159–65.
Acknowledgements
The authors thank Professor Duco Hamasaki, Bascom Palmer Eye Institute, University of Miami School of Medicine, for discussions and editing this manuscript.
Funding was provided by Grant-in-Aid for Scientific Research (KAKENHI) (Grant No. 16K11336).
Conflicts of interest
H. Oku, None; S. Morishita, None; T. Horie, None; Y. Nishikawa, None; T. Kida, None; M. Mimura, None; S. Kojima, None; T. Ikeda, None.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Oku, H., Morishita, S., Horie, T. et al. Protective effect of P7C3 on retinal ganglion cells from optic nerve injury. Jpn J Ophthalmol 61, 195–203 (2017). https://doi.org/10.1007/s10384-016-0493-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10384-016-0493-6