
Abstract
The lowermost carbonate beds of the Cerro San Pedro (San Pedro de la Cueva, Sonora State, Mexico) previously assigned to the Mississippian, belong in fact to the lower Ordovician. The limestone of this succession is often dolomitized and chertified, and displays gastropods, pelmatozoans, sponges, and trilobites as major bioclastic components. The different microfacies show that high-energy grainstones, proximal tempestites, and distal tempestites dominated the sedimentation. The paleoenvironments of deposition correspond to an inner ramp, a mid-ramp, and perhaps the upper part of an outer ramp. The strata are characterized by the incertae sedis cyanobacteria Nuia sibirica. A taxonomic revision and discussion of these cyanobacteria, often confused with ooidic grains, is emphasized. Some data are presented on other microfossils, such as primitive, monothalamous foraminifers: Rauserina sp., Vicinesphaera sp., and Neoarchaesphaera sp., leperditicopida and their endolithic microperforations, and primitive chaetetids. Compared to the contemporaneous deposits of the USA, some paleobiological components, e.g., the lithistid siliceous sponges Archaeoscyphia, stromatoporoids? Pulchrilamina, and receptaculacean algae Calathium, are quite rare in the studied section of Sonora, but the predominance of tempestites in the carbonate succession shows that boundstones formed before were systematically eroded and resedimented. The paleogeographic implications are the following: (1) a lower Ordovician intertropical Nuia Province is newly defined in the western part of the lower Ordovician intertropical belt; (2) San Pedro de la Cueva constitutes one of the westernmost outcrops of this new Nuia Province; (3) from Sonora, Nuia extends eastward as far as South China; (4) due to its westernmost paleoposition, San Pedro de la Cueva was frequently affected by tropical storms; and (5) northern lower Ordovician terranes of Mexico (Sonora, Chihuahua, and Baja California) are entirely distinct from the southern ones (Oaxaca). The paleopositions and paleogeographic connections of these northern Mexican terranes with Laurentia, Avalonia, and peri-Gondwanan parts of South America, through the Iapetus and Rheic oceans, still remain disputable or unknown.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The geological record in Sonora State (Mexico) comprises a several-thousand-meters-thick Paleozoic series, which our team has investigated for a long time from both the biostratigraphic and paleoenvironmental points of view (Vachard et al. 2000a, b, c; El Albani et al. 2005; Derycke-Khatir et al. 2005; Palafox 2011; Buitrón-Sánchez et al. 2012; Palafox et al. 2013). We have concluded that during the late Paleozoic, and especially during the Carboniferous (Vachard et al. 2000b, c; Gómez-Espinosa et al. 2008), the future area of Mexico was composed of terranes dispersed in the Rheic Ocean. The closing of this ocean amalgated these Precambrian and Paleozoic terranes from various origins, to constitute the basement of Mexico (Nance et al. 2010). Another part of this basement, in northern Mexico, is composed of the southwestern extremity of the North American Craton along the northern margin of the Iapetus and Rheic Oceans. In its southern states, Mexico includes deposits of (1) the deep bottom of the Rheic Ocean under the form of a thick flysch series named the Acatlán Complex, (2) shallow siliciclastic deposits from several microplates, including the large tectonic blocks of Oaxaquia, Chortis and Maya, and (3) some outcrops generally limited to isolated hills (San Salvador Patlanoaya, Olinalá, San Juan Ixtaltepec, Los Hornos; see Vachard et al. 2000a, b, 2004) where some fragmented, shallow, carbonate shelves were present. No part of Mexico corresponds to the southern margin (i.e., the peri-Gondwanan border) of the Rheic Ocean; these latter crop out in Colombia and Venezuela (i.e., in northern South America). The oceanic history of Mexican terranes within the Iapetus Ocean, precursor of the Rheic, is less clear during the lower Paleozoic, and especially during the transition period from Iapetus to the Rheic opening. In spite of their quite poor knowledge, some preliminary studies have suggested the crucial and crustal significance of a number of terranes in Mexico (e.g., Robison and Pantoja-Alor 1968; Pantoja-Alor 1970; Buitrón and Rivera-Carranco 1984; Sour and Buitrón 1987; Keppie et al. 1996, 2003, 2007, 2008a, b; Landing et al. 2007).
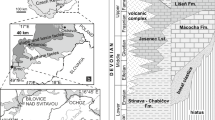
The lower Ordovician carbonate series of northern Mexico, known in the states of Baja California, Sonora and Chihuahua (Fig. 1; Table 1), have been traditionally compared to the El Paso Group of Texas and Oklahoma (USA) (Fig. 1c). Due to its academic, industrial, and economic interests, the El Paso Group, named by Richardson (1904), was accurately studied in the southeastern USA (e.g., Cloud and Barnes 1948; Hayes 1972; Wilson 1975; Toomey and Nitecki 1979; Roux 1985; Taylor et al. 2004, 2012; Pearce 2012; Lucia 2012; Sternbach 2012; Gregg and Shelton 2012; Miller et al. 2012; Fritz et al. 2012) (Figs. 1c, 2). In Baja California, Sonora and Chihuahua states, i.e., the Mexican parts of the North American Craton, there are sediments that correspond to the transition from a continental ramp (with shallow-sea carbonates) to an oceanic basin (with deposits bearing graptolites and radiolaria). The Furongian (upper Cambrian) to lower Ordovician sequences of southern Mexico in Tiñu (Fig. 1a) differ completely, indicating that as early as the early Paleozoic, the paleogeography and geodynamics of Mexico were compartmentalized in various independent terranes.

Geographical maps. a Map of Mexico and adjacent countries. B Belize, G Republic of Guatemala, RH Republic of Honduras, RS Republic of El Salvador, USA United States of America, USM United States of Mexico. The box corresponds to Sonora and adjacent states. b Map of Sonora State in Mexico, with the main roads, main cities, and location of the studied area. The box corresponds to the area detailed in Fig. 3. c Northern states of Mexico (USM) and southern states of the USA with reference sections of the El Paso Formation (1–5) in Texas and Oklahoma (according to Toomey and Nitecki 1979, slightly modified). Baja Cal. N. Baja California Norte, Baja Cal. S. Baja California Sur, Coah. Coahuila, N.L. Nuevo León, Tamps. Tamaulipas, USA United States of America, USM United States of Mexico. 1 Wichita Mountains, 2 Arbuckle Mountains, 3 Franklin Mountains (i.e., the El Paso Fm stratotypic area), 4 Marathon area, 5 Llano region

Correlation scheme of uppermost Cambrian to lower Ordovician, Laurentian, and global chronostratigraphic units with conodont-based zones in North America (according to Taylor et al. 2012). The studied section, dated as lower Ordovician by Nuia sibirica, can be tentatively assigned to the Macerodus dianae-Reutterodus andinus interval (see explanations in the text)
The aims of this paper are to: (1) describe the microfacies of the poorly known Cerro San Pedro section (San Pedro de la Cueva, Sonora State, Mexico); (2) give a description of the microfaunal and algal assemblages; (3) reconstruct the depositional paleoenvironments; (4) provide new taxonomic data on the constituents discussed, especially Nuia and some primitive foraminifers, Neoarchaesphaera, Vicinesphaera, and Rauserina; (5) provide data on the lower Ordovician paleogeography of Sonora, and (6) define a tropical Nuia Province within the lower Ordovician Low Latitude Belt.
Ordovician successions in Sonora
The Ordovician outcrops in Sonora were mainly studied or compiled by Peiffer-Rangin (1979, 1987); Peiffer-Rangin et al. (1980), Eguizábal-Martínez (1988); Poole and Madrid (1988); Poole et al. (1995a); Stewart et al. (1990, 1999); Pérez-Ramos (1996); Page et al. (2012) and Stevens et al. (2014). They are doubtfully present in the northwestern part of the state (in the El Bisani hills; Brunner 1975, 1984; spelled Rancho Bízani by Poole et al. 1995a); relatively well known in the central part: Rancho Las Norias, Sierra San Juan de Dios, Cerro Sonora, Cerro Cobachi, Rancho La Casita, Sierra Martínez, Los Chinos, Sierra López, and the Barita de Sonora mine area (Fig. 3; Table 1), and relatively poorly known in the eastern part (Sierra Agua Verde, Cerro San Pedro, and Arivechi).

Geographical map of the central part of Sonora State (Mexico) with the Paleozoic and Triassic outcrops (according to Stewart et al. 1990; slightly modified). Toponymy: Rancho ranch, Cerro hill, Mina mine, Río river, Sierra range. G Graptolites (i.e., deep oceanic deposits), Q quartzites (i.e., epicontinental deposits). Boxed area San Pedro de la Cueva area (see Fig. 4)
Except for a recent study in Cerro Salazar (central Sonora; Beresi et al. 2012), the lower Ordovician succession of Sonora is poorly known. This paper on the San Pedro de la Cueva area allows us to describe an equivalent of the central part of the El Paso Formation in Texas, Oklahoma, New Mexico, and Arizona (e.g., Le Mone 1969; Ethington and Clark 1964, 1971; Wilson 1975; Toomey and Nitecki 1979; Taylor et al. 2012; Pearce 2012). Other already-known equivalents are, in the USA, the Manitou Formation in Colorado (Swett 1964; Gerhard 1972), Pogonip Group in Utah and Nevada (Wilson 1975; Carozzi 1989), and Garden City Formation in Utah (Pearce 2012). In Mexico, the El Paso Formation has also been identified in boreholes of Chihuahua State (López-Ramos 1969; Vachard and Téllez-Girón 1986; Haenggi 2001 with references therein; and this study: Table 1).
Geological setting of the studied area
The Cerro San Pedro is situated near the village of San Pedro de la Cueva (Sonora) in the northeastern border of the Novillo Reservoir (Figs. 3, 4), which impounds fresh water as far as Hermosillo, the capital of the state. It is a hill of 676 m, formed by several still poorly dated Paleozoic series (Fig. 4). Its Paleozoic sequence was initially studied by Peiffer-Rangin (1987), who assigned to it a late Tournaisian to early Viséan age. The hill was designated as Cerro El Azogue, but this name does not appear on the official maps. During a re-study of Cerro San Pedro, we were surprised to see that a part of the supposed Mississippian carbonates located at the northern edge of the hill (Peiffer-Rangin 1987, p. 52) is a lower Ordovician microfacies.

Cerro San Pedro, San Pedro de la Cueva, and Novillo Reservoir. Geographical and schematic geological map (based on Garcia-Cortez and Hernandez-Morales 2010); the red box indicates the location of the section
The lower Ordovician section described here is located in an arroyo (dry creek) at the foot of the Cerro San Pedro (Fig. 4). Coordinates of its base are 29°17′11′′N and 109°43′41′′W. The field section, oriented in a NE–SW direction with beds dipping at 48°S, shows approximately 50 m of pinkish to grayish limestone, occasionally affected by some normal faults (Fig. 5a) and/or differences of paleobathymetry (Fig. 5b). The limestone is thick-bedded (up to 2-m-thick beds), bioturbated, often wavy-bedded, chertified, or dolomitized.

Bioturbation is common in the lower part of the section (Fig. 6a, b, f), where lenses of black chert are aligned in the bedding planes (Fig. 6b, c). Obviously, the chert lenses succeeded the bioturbation (Fig. 6b). Various tempestites are recorded in this lower part of the sequence (Fig. 6c–f), whereas in the upper part of the series, biotubation, chert and tempestite are less common (Fig. 6g, h).

Field photos. (Scale bars: pencil for a, e = 14 cm; hammer for b–d, h = 33 cm; white segment for f = 10 cm; measuring tape of Miss Dulce Reyes for g = 2 m long). a Numerous bioturbations; sample Az 2/1. b Bioturbation passing to chert; sample Az 2/4. c Stratified chert and a chanellized unit; sample Az2-4. d Erosive contact (underlined) between two tempestites; sample Az 2/5. e Tempestite passing to a laminite, with a strongly erosive contact (underlined); sample Az 2/7. f Tempestite rich in transported grains passing to an autochthonous wackestone; sample Az 2/9. g Upper part of the series with chert; sample Az 2/10d. h Uppermost part of the series without chert and with Archaeoscyphia (A); sample Az 2/10 g
Compared to the lower Ordovician Boquinete Formation of the nearby Sierra Agua Verde (Ochoa-Granillo and Sosa-León 1993; Stewart et al. 1990, 1999), the studied series lithostratigraphically corresponds to Lower Carbonate Unit 2 (upper unit), dated from the upper middle Tremadocian to Floian, i.e., most probably to the M. dianae to R. andinus conodont zones (Fig. 2). Attempts at conodont extraction during this study were unsuccessful.
Microfacies and paleoenvironments
The predominant microfacies are wackestone to floatstone; they are associated with subordinate packstone and grainstone. All these microfacies contain a varied fauna, mainly composed of stenohaline forms such as gastropods, echinoderms, brachiopods, trilobites, ostracods, and siliceous sponges (Figs. 7, 8, 9, 10, 11, 12). The entire paleontological assemblage is composed of sporadic cyanobacteria Girvanella problematica Nicholson & Etheridge, 1878 and Nuia sibirica Maslov, 1954; receptaculitales Calathium? sp.; foraminifers Neoarchaesphaera sp., Vicinesphaera sp., and Rauserina sp., and various fragments of metazoans: sponges (monaxon, tetraxon, and polyaxon spicules, and entire skeletons of Archaeoscyphia sp.), gastropods (relatively common macluritids, bellerophontids and loxonemids), chaetetids (very rare), brachiopods (scarce), trilobites (relatively common), ostracods, leperditicopids, and echinoderms (common; mostly if not all pelmatozoans).

mf1 microfacies (Scale bars 500 μm). a Bioclastic wackestone with fenestrae (f), as an extraclast included in a bioclastic packstone with crinoids; sample Az2-5. b Bioclastic grainstone with dissociated, thin valves of ostracods and rare micropellets; spots of silicification are widespread (arrows); sample Az2-7. c Planar laminated grainstone (tidalite?) with couplets of micropellets and fragments of ostracods, sample Az2-7. d Detail of the laminae; sample Az2-7

mf1 and mf2 microfacies [Scale bars 1 mm for (a) 5 mm for (b); and 500 μm for c, d]. a Extraclastic grainstone where the micritic or biomicritic extraclasts are entirely surrounded by a layer generated by microperforations of type 1 (cortoid type 1 according to Flügel 2004; see Figs. 14d, 15), sample Az 3. b Bioclastic wackestone with a curved extraclast (chip) of microbialites (see “curled mud chips” of Wilson 1975, p. 83), sample Az 11. c Bioclastic and intraclastic grainstone with ostracods, echinodermal fragments, and Vicinesphaera (v), sample Az 2/5. d Bioclastic wackestone with Nuia sibirica (n), sample Az 6b

mf1, mf3, and mf5 microfacies (scale bars 1 mm). a Bioclastic and intraclastic grainstone, with some small dark intraclasts widespread in the microfacies and concentrated in its lower part, sample Az2-2-1. b Sequence boundary (SB) between a wackestone with spicules (bottom) and a grainstone with pellets and quartz grains, sample Az 2/4. c Typical distal tempestite: bioclastic grainstone with burrows (arrows; see the circular swirls of skeletal debris in Flügel 2004, pl. 19, Fig. 1) passing to a bioclastic wackestone (arrows), sample Az 2/3. d Tempestite: bioclastic grainstone and bioclastic wackestone with numerous burrows (arrow), sample Az 2/9.1

mf3 microfacies (scale bars = 5 mm for Fig. 10a, b; 1 mm for Fig. 10c; and 2.5 mm for Fig. 10d). a Recrystallized, poorly bioclastic wackestone with brachiopod (top left) and leperditicopid (top, centre; magnified Fig. 16g), echinoderm ossicles and trilobites (centre, right), sample Az 12a. b Bioclastic wackestone with gastropods (g), other molluscs (m), and Archaeoscyphia (see detail Fig. 16p), sample Az 18. c Bioclastic wackestone with echinoderm (e), gastropod (g) and trilobite (t) remains, along with leperditicopid valves (l) with microperforations of type 2 (see Figs. 15, 18b), sample Az 8b. d Bioclastic grainstone (bottom) with a lepertidicopid showing connected valves poorly infected by microperforations of type 2 (see details Fig. 16i) passing to mudstone (top), sample Az 8 k

mf3A and mf3B microfacies (scale bars 5 mm). a Bioclastic packstone (left) passing to a silty micrite (right), sample Az 15d. b Silty micrite with curved lamination (hummocky cross stratification?) passing to a bioclastic packstone/wackestone, by intermediary of a gutter-cast (?) with lag deposit, sample Az 9a. c Bioclastic wackestone with vertical burrow (b1) passing to a silty micrite with horizontal burrow (b2), sample Az 10a. d Tempestite: bioclastic wackestone overlain by a silty mudstone, sample Az 5

mf4 and mf5A-B microfacies (scale bars = 5 mm for Fig. 12a, b; 1 mm for Fig. 12c; and 2.5 mm for Fig. 12d; and 500 μm for Fig. 12e). a Tempestite: bioclastic packstone (bottom) with irregular surface (arrow) passing to a silty mudstone (top), sample Az 16b. b Echinoderm grainstone (bottom) passing to an echinoderm wackestone and a silty mudstone (top), sample Az 21. c Wackestone with abundant monaxon sponge spicules, sample Az 2-/2b. d A poorly bioclastic wackestone overlain by a wackestone with spicules, sample Az 2c. e Wackestone poor in bioclasts (ostracod-rich dark areas probably represent bioturbation), sample Az 2/8
The microfacies and biota are characteristic of open-marine, well-oxygenated, and well-circulated conditions with normal marine salinity, deposited below the fair weather wave base (FWWB) in a deeper, open-marine shelf environment with water depths of at least some tens of meters (Wilson 1975; Flügel 2004, 2010). High-energy deposits generated by waves (grainstone) and storm (packstone to floatstone) are obvious. Silica of the chert nodules and chert bands is probably derived from numerous siliceous sponges, the presence of which is indicated by abundant sponge spicules.
Five principal types of microfacies (mf1 to mf5; Figs. 5b, 13; Table 2) were observed, which have been correlated to the standard microfacies types (SMF) and ramp microfacies (RMF) of Flügel (2004) (Table 2). First, we note that the SMF and RMF of Flügel (2004) are relatively poorly applicable to the lower Ordovician. Nevertheless: (1) due to the rarity of Flügel’s microfacies SMF12 to SMF26, the innermost ramp and the intertidal and supratidal environments are almost totally absent from our material, except for an extraclast with fenestrae (Fig. 7a); (2) the different microfacies mf1A–D (Fig. 13) are located in the inner ramp, above the FWWB; under the form of bioclastic and/or micropelloidal grainstones; (3) the microfacies mf2 probably correspond to proximal tempestites because they contain predominantly photozoan bioclasts (e.g., Nuia, Girvanella, Calathium?), and the microfacies mf3 and mf4 probably correspond to distal tempestites because they are devoid of photozoan elements and composed of heterozoan elements (e.g., leperditicopids, gastropods, trilobites); and (4) because the deepest microfacies of Flügel (i.e., SMF1 or SMF18) are rarely observed in our material, the deposits of middle and outer ramps seem to be only represented by wackestone with monaxon spicules (microfacies mf5A), or poorly fossiliferous wackestone (mf5B).

Composition and interpretation of microfacies and reconstructed transect of Cerro San Pedro. Az samples from El Azogue, FWWB fair-weather wave base, mf microfacies studied in this paper, SWB storm wave base. 1–5 (left column) refer to regional microfacies types (mf) described in the text
The microfacies mf1 (Figs. 7a–d, 8a–c) are represented by various grainstones; i.e., they were deposited in agitated environments with swell, wind waves, storm waves, tidal currents and bottom currents. The shallowest environments exhibit fenestrae (only recorded as intraclasts; Fig. 7a), grainstones with ostracods (Fig. 7b), and laminites (tidalites?) with couplets of grainstone with micropellets of microbialites and grainstone with thin valves of ostracods (Fig. 7c, d), extraclastic grainstones (Fig. 7a, b), and typical bioclastic grainstones (Fig. 7c).
Under the FWWB, there are various bioclastic microfacies, but all seem to correspond to several types of tempestites. The microfacies mf2 corresponds to proximal tempestites, according to the criteria of Flügel (2004), which contain the cyanobacteria Nuia and Girvanella (Fig. 8d). As these tempestites contain photozoan bioclasts, and frequently present mixed grainstone and packstone microfacies, they probably accumulated near the FWWB (Burchette and Wright 1992) (at a depth of approximately 10–20 m or even shallower). The paleoecology of Nuia, reconstructed for instance by Roux (1985), permits us to indicate that the biotopes with Nuia were located in shallower environments (depth 0–5 m).
The microfacies mf3 shows a heterozoan assemblage with echinoderm ossicles, ostracods, leperditicopids, gastropods and sporadic brachiopods (Figs. 10a–d, 11a–d); where pelmatozoan accumulations are relatively common, but not in the echinoderm limestones of mf4 (Fig. 12a, b). Several deposits are interpreted as tempestites because of the abnormal concentration in bioclasts (especially, pelmatozoan ossicles) and some sedimentary features: e.g., graded bedding, bioturbation on the top of beds; erosive surfaces; mixing of packstone and grainstone (Figs. 10d, 11a–d, 12a, b); however, no hummocky stratifications were observed in the field, because the deposits consist of carbonates and not siliciclastics or mixed sediments as well as relatively atypical lower Paleozoic tempestites (Sepkoski et al. 1991). Nevertheless, hummocky beds and gutter casts are possibly present in a single microfacies (Fig. 11b). Some contacts between distinct lithologies are transitional (Fig. 11c); others are well marked (Fig. 11a, b, d); the latter are secondarily affected by pressure solution, and show stylolitic joints (Fig. 11d). Their deposits occur probably in the lower part of an inner ramp and upper part of a mid-ramp. The traditional bioconstructions with Archaeoscyphia of the El Paso Group were not found in situ in this study, but they contributed probably to the reworked material of a part of these tempestites (Fig. 10b), possibly located in the upper part of this paleobathymetric zone (see Wilson 1975, text-fig. IV-3 p. 99).
The microfacies mf3 and mf4 were supposedly deposited in the outer part of a mid-ramp, as far as the SWB, whereas the microfacies mf5, deposited near the SWB with wackestone with spicules (mf5A; Fig. 12c, d) and poorly fossiliferous wackestone (mf5B; Fig. 12d, e), correspond to the deepest environments of an outer mid-ramp, or was already located on an outer ramp. However, they can rapidly emerge and be reworked, (1) in agitated environments, as ellipsoidal extraclasts, which are microperforated by probable cyanobacteria (microperforations of type 1) (Fig. 8a), and (2) in tempestites, as ellipsoidal fragments of biomicrite with reworked spicules (Fig. 14d, m). Several sequence boundaries (SB) between bioclastic grainstones of mf1 and wackestones with spicules of mf5A have been observed (Fig. 9b, c). Tectonically controlled uplift may have influenced this observed stratigraphic pattern. However, a close scrutiny of the regional stratigraphic pattern would be required in order to evaluate the tectonic influence on local to regional basin architecture.

Microfossils (scale bars: a–d, g, h = 0.100 mm; e, f, i–k = 0.500 mm). a Reticulate ostracod, tangential section, sample Az 2b. b, c, e, f, h–l Nuia sibirica. b Transverse section with two rings, sample Az 6. c Transverse section with one ring, sample Az 6a. e Two sections (n) in a bioclastic wackestone with echinodermal plates (e), sample Az 2/6. f One section (n) in a bioclastic grainstone, sample Az 2/2. h Oblique section with two rings and a possible bifurcation in transverse section, sample Az 6c. i Transverse section, sample Az2-6. j Oblique section with broad dark center, sample Az 6d. k Transverse section, sample Az2-6. l Longitudinal section of a not bifurcated specimen, sample Az 6e. d Biomicritic extraclast (i) with microperforations of type 1 (arrows) at the external margin with the matrix of dolomitized micrite, sample Az 2/3. g, n Girvanella problematica Nicholson and Etheridge (1878). g A set of trichomes with pellets of microbialites, sample Az 4. n Girvanella problematica, some trichomes arrowed, sample Az 10b. m Ellipsoidal extraclast of wackestone with short monaxon spicules, sample Az 9 k. o Hexaster spicule with a Nuia (top, right), sample Az 10Ba. p Polyaxon spicule with Nuia (n), sample Az 2/6; q, r Vicinesphaera sp. q Transverse section, sample Az 2/5. r Subtransverse section, sample Az 2/6. s, t Rauserina sp. s sample Az 14. t sample Az 13b
The reconstructed paleoenvironments (Fig. 13, Table 2) are consistent with the regional paleogeographic data in central Sonora (Fig. 3), and allow to suggest that (1) the limits between the early Ordovician shallow and deep seas were located south of San Pedro, probably in Cerro Cobachi, Barita de Sonora, Sierra El Aliso, and Sierrra El Encinal, and (2) the innermost ramp and the intertidal zone are situated in the north and west of San Pedro, and could correspond to the stromatolite development known for example in the Rancho Las Norias area (Fig. 3).
Systematic paleontology
Cyanobacteria
The lower Ordovician cyanobacteria, found in the photic zone of San Pedro de la Cueva, are Girvanella problematica Nicholson & Etheridge, 1878, Nuia sibirica Maslov, 1954, and microperforations of type 1.
The tubes of Girvanella problematica (Fig. 14g, n) are generally numerous in a nodular thallus; they are closely packed but twisted or contorted. Their external diameter is 15–20 μm with a thin wall of 3–5 μm. Although the morphologies of the filaments of Girvanella are very simple and their sizes homogeneous, many species and groups of species have been proposed (Wood 1963; Perret and Vachard 1977; Mamet and Roux 1977; Héroux et al. 1977). The genus appears to have diversified into different species during the Mississippian (Tournaisian–Viséan), even when its species are under discussion (compare Perret and Vachard 1977 and Mamet and Roux 1977). In the early and middle Paleozoic, the dominant species is Girvanella problematica, the type species (erroneously synonymized with G. ducii by Mamet and Roux 1977). The range of G. problematica seems to be Cambrian–Devonian.
Some minute microperforations of type 1 (Figs. 8a, 14d, m, 15) are obvious around biomicritic extraclasts, which are generally sponge-spicule bearing. They are constituted of minute tubules finely distributed at their periphery, forming cortoids (see Flügel 2004). They seem similar to Bevocastria hubbardi Mamet & Roux, 1975 (as revised by Vachard et al. 2014) and/or the probably misinterpreted Ordovician “perforating Girvanella” (Klement and Toomey 1967; Riding 1975; Toomey and LeMone 1977).

Different types of microperforations. a Perforations of type 1; b perforations of type 2; c perforations of type 3; d perforations of type 4 (explanations in the text; schemes without scale to compare with microfacies figures)
Genus Nuia Maslov, 1954.
Type species Nuia sibirica Maslov, 1954.
Diagnosis Thallus, cylindrical to botryoid in shape, with a central, dark, cylindrical part (i.e., round in transverse section) corresponding to the emplacement of possible trichomes, and a “wall” with one layer of fine, acicular, radiate, crystals of clear calcite. So-called multilayered walls are in reality successive attached, monolayered thalli. Rare bifurcations can occur (Mamet and Roux 1982; Ross et al. 1988). Productivity can be strong to weak according to the bioaccumulations or scarce occurrences observed in microfacies.
Remarks Nuia is a taphotaxon at the sense of Lucas (2001); i.e., it is not the exact mirror of a living morphology, but a morphospecies appearing during the taphonomic process, as an early diagenetic object derived from an ontogenetic object. It is only in this sense, that the comparison of Nuia to Microcodium, ooids or microstromatolites, seems to be understandable; noting however that such interpretations are sometimes too conjectural because, for example, Paleozoic classical oolites were described as the algal genus Bobolites Voronova in Voronova and Radionova (1976). The observations summarized herein confirm the reconstruction sketch of Nuia by Vachard and Téllez-Girón (1986) as the most likely interpretation; it is re-illustrated herein (Fig. 17d). Vachard and Téllez-Girón (1986) concluded with an assignment to the cyanobacteria. Contrary to the assertion of Riding and Fan (2001); Perret and Vachard (1977) never assigned Nuia to the red algae. This puzzling proposal was made by Huang and Bian (1983) due to a disputable interpretation of the taphonomic transformation of the trichome mucilage. The so-called Nuia in the Devonian of the Urals (Shuysky 1973b) or in Belgium (Mamet and Boulvain 1992; Pratt 1995) more likely correspond to other eodiagenetic taphotaxa such as Palaeomicrocodium or Tuborecta Saltovskaya, 1981. Putative comparisons, notably with atypical, upper Cambrian ooidal grains, have also lead to misinterpretations (e.g., Ross et al. 1988, fig. 1.1–4; Spincer 1998, figs. 6.1–3, 7.1; Lothringer 1993, fig. 8).
The American outcrops with Nuia are well known, based on the work of Toomey and Klement (1966); Johnson (1966); Toomey (1967); Ahr (1971); Wilson (1975); Guilbault et al. (1976); Toomey and LeMone (1977); Gnoli and Serpagli (1980); Mamet and Roux (1982); Roux (1985, 1991b); Vachard and Téllez-Girón (1986); Peiffer-Rangin (1987); Ross et al. (1988); Carozzi (1989); Albertstadt and Repetski (1989); Clemons (1991); Mamet and Shalaby (1995); especially in the El Paso Group of Texas and New Mexico (USA) and of Chihuahua (Mexico), the Hitt Canyon Formation (also called Sierrite Limestone) of the Mud Springs Mountains (New Mexico) (Karl Krainer, pers. comm., February 2015), the Pogonip Group (Nevada), the Kinblade Formation (Colorado), the Bromide Fm (Oklahoma), the Black Rock Fm (Arkansas), the subsurface rocks of Arkansas, Mississippi, Alabama and Tennessee (Alberstadt and Repetski 1989), the Saint-Lawrence Platform (New York, Vermont, U.S.A; Québec, Canada), the Sarbach beds of southern Alberta (Canada), and the San Juan Cordillera (Argentina). Additionally, Nuia was mentioned in Kazakhstan, Siberia, Tarim in NW China, Jiangsu and Guizhou in South China, as well as in Antarctica (Flügel 2004). By contrast, the Nuia mentioned in other areas have neither been illustrated nor confirmed (e.g., middle Ordovician of Scotland only mentioned but not illustrated by Ince 1984, p. 230) or obviously belong to other taxa (e.g., late Cambrian of Newfoundland in Conoglio and James (1985) or central Texas (Spincer 1998)). Consequently, they are excluded by us from the genus Nuia. It is noteworthy that such false Nuia are generally late Cambrian (Conoglio and James 1985; Ross et al. 1988; Spincer 1998), Ordovician sensu lato (Lothringer 1993), or middle Ordovician (Ince 1984; Pratt 1995) in age. As mentioned by Ross et al. (1988), no middle Ordovician Nuia was found in Kazakhstan by Gnilovskaya (1972) in her accurate study of the algae of this area. This confirms that Nuia sensu stricto is only early Ordovician in age.
Occurrence Eastern Siberia (Angara, Nyuya, Zherba, Biryuk and Lena rivers) (e.g., Maslov 1954, 1956, 1973; Reitlinger 1959; Dubatolov et al. 1985); Canada (Québec); U.S.A. (Nevada, Texas, New Mexico, Oklahoma, Utah, Colorado, Arkansas, New York, Vermont; Roux 1985 and additional references); Mexico (Chihuahua and Sonora: Vachard and Téllez-Girón 1986; Peiffer-Rangin 1987; Eguizábal-Martínez 1988); Argentina (Gnoli and Serpagli 1980; Keller 1999, 2012; Astini 2001); Kazakhstan (Chu-Illi Mountains; Reitlinger 1959); China: Tarim (Riding and Fan 2001), Jiangsu (Huang and Bian 1983), Guizhou (Li et al. 2014), and Hubei (Li et al. 2015); and Antarctica (Flügel 2004).
Nuia sibirica Maslov, 1954.
Figures 8d, 14b, c, e, f, h–l, o, 16j, s, 17a–d.

Microfossils (Scale bars a–d, r, s = 0.250 mm; e–g, i, k, l = 0.500 mm; h, j = 0.100 mm; m–o, q = 0.05 mm, p = 1 mm). a–d, n, o Neoarchaesphaera sp. (n, arrowed) a sample Az 15. b Sample Az 15a. c Sample Az 17. d sample Az 2. n Sample Az 2. o Sample Az 2. e Echinoderm ossicle (thecal plate?) with microperforations of type 4, sample Az 16b. f, g, i, k, l Leperditicopida with microperforations of type 2. f Sample Az 16. g Sample Az 12. i Sample Az 8i. k Microperforations of type 2 (2) and type 3 (3), sample Az 13. l Sample Az 13. j, s Nuia sibirica, j Typical transverse section sample Az 6. s Subtransverse section with two rings (see also Fig. 12d), sample Az 20a. m Vicinesphaera sp., sample Az 3. p Archaeoscyphia sp., oblique section (see Fig. 14b), sample Az 18a. q, r Rauserina sp. q Sample Az 19c. r Associated with echinoderm and brachiopod, sample Az 19b

Reconstructions of Nuia. a–c Stages of calcification. a In the living cyanobacteria, the heterocysts are aligned in the median plane, or at the base when a second thallus is attached on the first one. The trichomes are directed perpendicularly to the thallus axis, or arranged in fans. Their individual calcification leads to the “fibers” of calcite. b After disappearance of the trichomes, their richness in organic matter gives rise to a dark space, central or basal. The fibers less rich in organic matter appear in clear calcite. c When a second attached thallus entirely surrounds the first thallus, a complete dark space or a dark line appears. d Complete reconstruction of the taphotaxon (according to Vachard and Téllez-Girón 1986). Accordingly, Nuia is relatively similar to some Nostocales Rivulariaceae, even if all the representatives of this family are only known in fresh waters
1954 Nuia sibirica Maslov, p. 526, pl. 1, fig. 2.
1986 Nuia sibirica Maslov - Vachard and Téllez-Girón, p. 15–16, pl. 1, figs. 1–11 (with synonymy).
This previous synonymy list of Vachard and Téllez-Girón (1986) is completed here by the following references:
1964 algal (?) allochem-Swett, figs. 2d–f, 3a.
1966a Nuia texana Johnson, p. 433, pl. 57, figs. 1–7.
1970 Nuia texana Johnson - Young, p. 2305 (no illustration).
?1972 Nuea (sic) - Gerhard, p. 8 (no illustration).
1977 the possible alga, Nuia - Cooks and Taylor, p. 56, text fig. 10 p. 57, text fig. 40 p. 74, text fig. 44 p. 76.
non 1981 Nuia sibirica Maslov-Saltovskaya, p. 114, pl. 1, figs. 20, 21 (=?Tuborecta).
1983 Nuia siberica [sic] Maslov - Huang and Bian, pl. 1, figs. 1–10.
1984 Nuia - Beresi, text fig. p. 79, pl. 1 p. 80.
1985 Nuia sibirica Maslov - Dubatolov et al., p. 5 (no illustration).
1985 Nuia sibirica Maslov - Roux, pl. 6, fig. 8.
1988 Nuia - Ross et al., figs 4.1–4, 5.1–4.
1989 Nuia sibirica Maslov - Carozzi, p. 209, 210, 212, text-fig. 11–1 p. 210, text-11.2F p. 211.
1989 Nuia - Albertstadt and Repetski, p. 225, 227, 228, 231, 237, 238, 240, fig. 4J.
1991 Nuia - Clemons, p. 28–30, figs. 14, 32a–d, 46.
1991b Nuia - Roux, p. 351 (no illustration).
non 1992 Nuia sibirica Maslov - Mamet and Boulvain, p. 288, pl. 1, figs. 11–13, pl. 2, figs. 15,16 (more similar to microcodiaceans; probably Palaeomicrocodium).
1995 Nuia sibirica Maslov - Mamet and Shalaby, pl. 6, figs. 5–14.
?1998 Nuia siberica [sic] Maslov - Spincer, p. 577, 578, 580 (no illustration).
p. 1998 Nuia - Spincer, p. 577, 578, 580, 581, 582, 583 (non fig. 3.1–3, 6.2?, 3, 7.1–2).
1999 Nuia - Keller, p. 38, 40, fig. 24. 1, 5.
2001 Nuia sibirica Maslov - Riding and Fan, p. 803, pl. 3, figs. 4–5.
2004 Nuia - Flügel, pl. 98, fig. 1.
2012 Nuia - Pearce, p. 13, 14, 31, 32, 43, 44, 45 (no illustration).
2012 Nuia siberica [sic] Maslov - Pearce, p. 16 (no illustration).
2012 Nuia - Keller, p. 995 (no illustration).
2012 Nuia – Beresi et al., p. 334, 338, 339, Fig. 4d, n.
2013 Nuia - Ketner, text-fig. 15 p. 10.
non 2013 Nuia sibirica - Sinha and Trampisch, p. 346, pl. 2, Fig. 4 (an alga, dasycladalean or issinellacean).
2015 Nuia fragment - Li et al. fig 7b.
Description The morphotypes of this study are identical to those of Toomey and Klement (1966) and Johnson (Johnson 1966) from western Texas. Moreover, as suggested by Guilbault et al. (1976; figs. 1, 2), the different lower Ordovician species from the USA and Russia are probably synonymous. Moreover, all Ordovician morphospecies compiled by Shuysky (1973a, p. 68, b, p. 94–95) might be synonyms of the type species Nuia sibirica, due to its extreme morphological variability.
Dimensions: Maximal length (L) = 400–2000 μm; external diameter (D) = 150–250 μm; diameter of internal black part (=“central cavity”) (d) = 50–90 μm; thickness of calcified part (“wall”) (s) = 25–50 μm; width of clear crystals (c) = 10–20 μm.
Occurrence As for the genus.
Other Algae?
The chlorophyte algae remain questionable during this period; the true Dasycladales, Chaetoporales, and Bryopsidales appear in the middle-late Ordovician (Gnilovskaya 1972; Elliott 1972; Bourque et al. 1981; Roux 1985; Poncet 1986; Mamet et al. 1992; Mamet and Shalaby 1995), and, in our opinion, neither the receptaculitales Calathium nor the Cambrian seletonellaceans of Kordé (1973) belong to chlorophyte algae. On the other hand, only one elianellacean (i.e., solenoporacean auctorum) red alga was observed (Fig. 16h).
Some outer plates of receptaculitales have been observed (Fig. 18h), similar to those illustrated by Mamet et al. (1992, pl. 11, figs. 10–13, pl. 12, figs. 1–3).

Microfossils (scale bars 0.500 mm; except for d = 0.100 mm; and f, k = 1.000 mm). a–c, f Leperditicopida with spherical microperforations of type 2 (some arrowed; see Fig. 18 b, f). a sample AZ 7c (see also Fig. 11a). b sample Az 8c. c sample Az 8e. f sample Az 9d. d Pelmatozoan (echinoderm) columnal (stem ossicle) with pentaradiate lumen, sample Az 7g. e Pelmatozoan ossicle from a free feeding appendage (arm sensu lato), sample Az 7h. g Echinoderm plate, bright spots are secondary silicifications, sample Az 2/2b. h Outer plate of receptaculitale Calathium? sp., sample Az 2/6. i Pelmatozoan ossicle, sample Az2-5. j Leperditicopida with geopetal filling, sample Az 2/15. k Two trilobite fragments (top and bottom left), sample Az 2/15. l, m Chaetetid indet., two fragments of colonies partly silicified, sample Az 2/14
Algal and/or fungal endolithic borings are common in our material. Modern endolithic microperforations, which have diameters of less than 100 μm, are produced by phototrophic (cyanobacteria, chlorophytes, and rhodophytes) and heterotrophic (bacteria and fungi) groups (Perry and MacDonald 2002; Gektidis et al. 2007). Due to light penetration as a limiting factor, many bathymetric distributions were proposed for the microendoliths (Golubic et al. 1975, 2005; Kobluk and Kahle 1977; Tavernier and Golubic 1993; Perry and MacDonald 2002; Gektidis et al. 2007; with references therein). In general, the cyanobacteria inhabit the supra- and intertidal areas, a mixed community of cyanobacteria and chlorophyta colonize shallow waters, and heterotrophic fungi and bacteria are dominantly located in deep aphotic waters.
In this study, the most abundant microperforations are spherical or ampulliform cavities, bored in the wall of a majority of the leperditicopida: type 2 microperforations (Figs. 10a–d, 15, 16f, g, i–l, 18a–c, f).
They affect always the outer surface of the wall and, apparently, do not cross through the wall. They are separated by large imperforate intervals. These microperforations affect almost all of the leperditicopid valves, but are apparently less common in our Devonian collections or in the literature (see e.g., Préat and Kasimi 1995, pl. 12, figs. 3–6; Mamet and Préat 2005, pl. 3, fig. 6, and 2007, fig. 46). On the other hand, microperforations of type 2 seem to be the consequence of an active boring in the wall of living leperditicopida, because 99% of these microperforations only affect the upper part of one or both valves. In contrast, the other types of microperforations (1, 3 and 4) affect dead, and often fragmented skeletons. These microperforations differ from those affecting the ostracod valves of the middle Ordovician of Poland (Olempska 1986), but are comparable with the Cretaceous borings described by Asgaard and Bromley (1991), which were produced while the affected rhynchonellid brachiopods were alive.
Type 3 microperforations (Figs. 15, 16k) are thin and short, and are organized in parallel series, also parallel to the surfaces of the bioclasts; they principally affect trilobite and brachiopod shell fragments.
Type 4 microperforations (Figs. 15, 16e) are represented by only one perforation, similar in dimensions to type 2 but more ampulliform than subspherical, and which affects an echinoderm ossicle.
Phylum Rhizaria Cavalier-Smith 2002.
Subphylum Foraminifera d’Orbigny 1826 nom. translat. Cavalier-Smith 2002.
Remarks The oldest unquestionable foraminifers are known in the African Taoudeni Basin (Algeria-Mali) and adjacent areas (Culver 1991, 1994; Culver et al. 1996). They were found in shallow marine siltstones, which are dated from the end of the early Cambrian (e.g., Bubik 2001). Rare Cambrian foraminifers were also mentioned in Kazakhstan (Reitlinger 1959). After that, the foraminifers really begin their history during the Ordovician.
All Ordovician foraminifers are unilocular or bilocular. The most primitive forms, Archaeochitosa, Chitinolagena, Labyrinthochitinia, and Maylisoria have membranous or tectinous skeletons (Loeblich and Tappan 1987; Vdovenko et al. 1993 with references therein), and probably belong to the Allogromiata (sensu Vachard 2016). On the other hand, more evolved genera were described with an agglutinated wall and probably belong to the Astrorhizata (sensu Vachard 2016). They are, in alphabetical order, Amphitremoida, Bathysiphon, Blastammina, Hemisphaerammina, Hyperammina, Lagenammina, Lakites, Lavella, Marsipella, Ordovicina, Psammosphaera, Pelosina, Rhabdammina, Raibosammina, Sorosphaera, Stegnammina, Tholosina, Tolypammina, Thurammina, and Thuramminoides (e.g., Moore et al. 1952; Rauzer-Chernousova and Reitlinger 1957; Loeblich and Tappan 1964, 1987; Langer 1969; Conkin and Conkin 1979; Poyarkov 1979; Lipps 1992; Sabirov and Gushchin 2006). Paradoxically, a majority of these genera of Ordovician Astrorhizata have a type species which was described in the Holocene seas.
Recently, the importance of the genus Amphitremoida was highlighted by Nestell and Tolmacheva (2004) in the lower Ordovician of NW Russia, and in the lower-middle Ordovician (Floian-Darriwillian) of Argentina by Nestell et al. (2009, 2011). According to these latter authors, the middle Ordovician foraminiferal assemblage of Argentina includes the genera or morphogenera Amphitremoida, Lakites, Lavella, Ordovicina and Pelosina. In contrast, some other early Ordovician foraminifers mentioned in Russia are still questionable and probably confused with Devonian forms (see Sabirov and Gushchin 2006; compare with Poyarkov 1969, 1979; and see discussions in Vachard et al. 2010, p. 220 and Nestell et al. 2011, p. 606).
Three early Ordovician genera are present in our material of Sonora, Neoarchaesphaera sp., Vicinesphaera sp. and Rauserina sp. All these genera have a true microgranular wall and not an agglutinated wall. No bilocular or plurilocular foraminifers have been found during our study. Some authors consider nevertheless that plurilocular, coiled and uniseriate genera are present, such as “Nodosaria” and “Aristorespira” (Sabirov and Gushchin 2006 with references therein) and, since the middle Ordovician, Reophax blackriveranus Gutschick, 1986, or, in the Silurian Reophax sp. (described by Ireland 1939). We cannot confirm these reports, and, in our opinion, plurilocular forms only appear in the upper lower to lower middle Devonian (Emsian and/or Eifelian) and first become common in Givetian time (see also Poyarkov 1979; text-figs. 20–23). Moreover, “Reophax” blackriveranus possesses an organic basal layer in its wall, whereas this organic layer is unknown within the Holocene type species. It is also worth noting that the distribution of Reophax Montfort, indicated as Cambrian to Holocene (e.g., in Cushman and Waters 1928, p. 37), or “middle Ordovician (rare)-upper Devonian–lower Carboniferous, Permian–Holocene” by Vdovenko et al. (1993, p. 25), corresponds to the longest range registered for a plurilocular foraminifer. Moreover, the upper Devonian “Reophax” belong most probably to Oxinoxis Gutschick; the lower Carboniferous “Reophax” are tests of Darjella Malakhova and/or Lugtonia Cummings; and the Permian “Reophax” forms are tests and/or internal molds of possible Endothyranella, Spireitlina or Nodosariata. Consequently, the genus Reophax sensu stricto probably only appears in the Triassic.
?Class Fusulinata Fursenko 1958 emend. Vachard 2016.
?Subclass Afusulinana Vachard et al. 2010.
Order Parathuramminida Mikhalevich 1980.
Description See Vachard (2016).
Remarks About this order and its disputed assignment, see discussions in Kaczmierczak (1976); Mikhalevich (1980); Vachard (1994), (2016); Kazmierczak and Kremer (2005); Mamet (2006); Versteegh et al. (2009); Schlagwinteit et al. (2013); and herein, the paragraph devoted to Rauserina.
Family Parathuramminidae Bykova in Bykova and Polenova, 1955.
Genus Neoarchaesphaera Miklukho-Maklay, 1963.
Type species Neoarchaesphaera bykovae Miklukho-Maklay, 1963 (=Archaesphaera magna sensu Bykova in Bykova and Polenova 1955 non Suleimanov 1945).
Synonyms Parathurammina (part.); Archaesphaera (part.) auctorum.
Description Small-sized Parathuramminidae with an irregular angular-rounded to spherical profile, fairly abundant papilliform to longer protuberances apparently not perforated, broad central chamber, thin dark-microgranular wall, and inconspicuous apertures.
Remarks See discussions in Loeblich and Tappan (1987) and Vdovenko et al. (1993).
Occurrence Discovered in the lower Ordovician of Sonora. Silurian of the Urals (Pronina 1963). Relatively frequent in the Devonian with e.g., Parathurammina sensu Malakhova (1969, pl. 48, figs. 330–331, pl. 49, fig. 337), Parathurammina? sensu Racki and Sobón-Podgorska (1993, text-fig. 9a–c), and “Thurammina without marked projections” sensu Holcová and Slavík (2013, text-fig. p. 215).
Neoarchaesphaera sp.
Figure 16a, b, d, n, o.
Description Small parathuramminid characterized by a relatively broad chamber, 1–7 radiate protuberances in random section, in prolongation of the smooth angular corners of the chambers, and a unilayered, dark-microgranular wall. The protuberances are very short. The wall is thin, dark and microgranular. Dimensions: Outer diameter = 40–70 μm; inner diameter = 30–50 μm; wall thickness = 5–10 μm; length of protuberances = 5–10 μm; basal diameter of protuberances = 3–5 μm.
Occurrence Lower Ordovician of Sonora (very rare).
Genus Vicinesphaera Antropov, 1950.
Type species Vicinesphaera squalida Antropov, 1950.
Description Small parathuramminid characterized by a relatively broad spherical chamber. The wall is thick, conical to subquadratic, and with inconspicuous perforations. It is thin, dark and microgranular.
Remark Although Vicinesphaera was questionably reported from the Cambrian (Reitlinger 1959; Chuvashov 1965; Armstrong and Mamet 1977), its presence is indisputable in our lower Ordovician collections.
Occurrence ?Cambrian–Ordovician–Viséan; cosmopolitan.
Vicinesphaera sp.
Description The test is conical. The inner chamber is medium-sized and perfectly spherical. The wall is particularly thick. Dimensions: minimal diameter = 225 μm; maximal diameter = 280 μm; chamber diameter = 80 μm; wall thickness = 150 μm.
Occurrence lower Ordovician of NW Mexico (very rare).
Genus Rauserina Antropov, 1950.
Type species Rauserina notata Antropov, 1950.
Diagnosis Test free, monothalamous but with two or several chambers, connected by an areal, intercameral opening. External apertures are inconspicuous. Wall dark-microgranular.
Remark Rauserina is a very questionable foraminifer because: (1) its type of wall exists in many groups; e.g., in cyanobacteria such as Girvanella, Halysis Høeg, 1932 and Tuxekanella Riding & Soja, 1993 (see Skompski 2010), calcitarcha, tuberitinoids, parathuramminoids and irregularinoids (see discussion about all these latter taxa in Vachard 2016); (2) many similar groups of connected, minute, spherical bodies exist in Devonian Ningbingellina Mamet, 1998, Viséan “Ningbingellina” (sensu Devuyst 2006), Permian Floritheca Gaillot & Vachard, 2007, generally interpreted as algal spores or microproblematica; as well as some transverse sections of groups of tubular, cyanobacterial trichomes: Halysis and Tuxekanella; for instance, the Rauserina of Brazhnikova and Rostovceva (1966; pl. 1, Figs. 21, 22, 27–29, 32); (3) they are apparently devoid of functional apertures as well as wall microforamina allowing the passage of the pseudopodia; and (4) when they re-used “Rauserina”, Mamet et al. (1992) assigned its to the calcisphaeridae; consequently, they have made an important emendation (not mentioned in their description). Furthermore, we note that all the so-called multilocular foraminifers in the lower Paleozoic more resemble these groups of spheres than an undisputable foraminifer (Gutschick 1962, 1986; Scott et al. 2003). On the other hand, it is probable that many Paleozoic “Sorosphaera” do not correspond to the Holocene genus created by Brady (1879), and are most probably secondarily silicified Rauserina. There are Sorosphaera bicella Dunn, 1942; S. bicelloidea Stewart and Lampe, 1947; S. colombiensis Stewart and Lampe, 1947; S.? cooperensis Conkin et al., 1968; S. multicella Dunn, 1942; S. osgoodensis Stewart and Priddy, 1941; S. papilla Gutschick and Treckman, 1959; S. subconfusa Dunn, 1942, S. tricella Moreman, 1930; see also S.? sp. of Culver, 1994 and Psammosphaera bipartita Ireland, 1939.
Occurrence Ordovician of Québec (Canada) and the USA (including the Sorosphaera spp. mentioned above). Discovered in the Ordovician of Sonora (Mexico). Silurian–lower Tournaisian (and even upper Tournaisian, according to Kulagina 2013) of Russian Platform and South Urals, Western Siberia, Donbass, Tien-Shan, and Italy.
Rauserina sp.
? 1973 Rauserina-Browne and Pohl, p. 195, pl. 5, figs. 4–6.
?1985 Rauserina notata-Dubatolov et al., p. 40 (no illustration).
?1992 Rauserina notata Antropov-Mamet et al., pl. 11, figs. 1–8 (with synonymy).
?1995 Rauserina notata Antropov-Mamet and Shalaby, pl. 5, Figs. 11, 12, 13.
Description These groups of spherical chambers have a dark microgranular wall but the median apertures were not observed. They correspond to Rauserina sensu Mamet et al. (1992) but also to Webbinelloidea tholus (Moreman, 1933) sensu Amsden et al. (1980). Length = 4400 μm; height = 200 μm; diameter of chambers = 70–100 µm; wall thickness = 7–15 µm.
Occurrence ?Ordovician–?Devonian. North America and Siberia.
Other Microfossils.
Chaetetid indet.
Figure 18l, m.
Rare specimens of small cerioid colonies with slender corallites, and perforate walls, aseptate and rare tabulae, might correspond to Lichenariidae. It is noteworthy that the primitive chaetetetids share morphological similarities with calcisponges, bryozoans and paleofavositids (tabulate corals). In contrast to the San Pedro de la Cueva section, bryozoans were relatively common in the Ojinaga-1 borehole studied by Vachard and Téllez-Girón (1986).
Ostracods and leperditicopida
Figures 7b–d, 8c, 10a, c–d, 11a–d, 12e, 14a, f, 16f, g, i, k, l, 18a–c, f, j.
Many true ostracods were observed, as well as many leperditicopids conventionally treated as ostracods. Although leperditicopids first become common and widespread in the middle Ordovician of North America (Berdan 1984; Williams and Siveter 2008), the group is known to appear in the lower Ordovician in Baltica (Williams and Siveter 2008). It is also abundant in the lower Ordovician of San Pedro. Similarly, true ostracods are mentioned in the lower Ordovician of Argentina (Salas et al. 2007). Therefore, the areas where appear both groups, leperditicopids and ostracods, are still discussed, but Sonora seems to be a good candidate.
Echinoderm ossicles
Figures 7b–d, 8c, 9d, 10a, 11d, 12a, b, 14e, f, 154, 16e, r, 18d, e, g–i.
Echinoderm remains are locally abundant in deposits from the middle inner ramp to deeper inner ramp environments. They are mostly recrystallized and fragmented (particularly in tempestites) (Fig. 12a, b), but some retain their original shape and even some stereom details (e.g., Figure 16r). Among them, some unidentified thecal ossicles are observed (e.g., Figure 18e), but most of recognizable elements are disarticulated from pelmatozoan (stemmed) echinoderms. Various free feeding-appendage ossicles include blastozoan-like, “brachiolar” elements (simple oral groove with a low depth–width ratio, e.g., Figure 18d), together with uniserial, more typical crinoid-like “brachials”. These ossicle types (Fig. 18e, i) have a deeper and narrower tripartite food groove and interestingly lack an axial canal, a diagnostic feature seen only in derived crinoids. It is worth noting that according to the diagnostic-features of blastozoans, crinoids and other echinoderms (e.g., stylophorans) advanced by Guensburg et al. (2010), it is very speculative to suggest an assignment of disarticulated elements from feeding appendages, moreover based on thin-sections, except from the depth–width ratio of oral groove, which is a subjective character. Indeed, even if indisputable crinoids have been defined from the Tremadocian (lower Ordovician) upward, Clausen et al. (2009) and Kouchinsky et al. (2010, 2015) have illustrated and discussed etched crinoid-like uniserial ossicles from lower to middle Cambrian limestones that would appear very similar to the ones discussed herein in thin section, but the affinity of which remains highly debated.
Other pelmatozoan ossicles from the Ordovician deposits of Cerro San Pedro include columnals with pentagonal lumen and outline (although slightly rounded). Lumen and outline have opposed angles. Due to partial to complete recrystallization, it is impossible to decipher the holomeric or polymeric nature of the columnals. As for brachials (sensu lato), it is impossible to definitely conclude the crinoid affinity of disarticulated columnals, as noted by Donovan (1986) (see also Clausen and Smith 2008). Only five crinoid genera are known from the early Tremadocian and two additional from the late Tremadocian, however, they already illustrate the predominant subclasses (Ausich et al. 2015). Therefore, if the crinoid affinity of some elements described herein is confirmed, they would represent very early representatives of this group. The presence of blastozoans is also testified by a thecal plate bearing a pectinirhomb-like structure (rhombiferan, Fig. 18g).
Paleobiogeography and Mexican terranology
Brief history of Mexican terrane studies
The tectonostratigraphic terranes of Mexico were accurately defined from Campa and Coney (1983) to Keppie et al. (2003) and Keppie (2004). As indicated by Keppie (2004, p. 769), “individual Mexican terranes have received different names (Campa and Coney 1983; Sedlock et al. 1993; Dickinson and Lawson 2001), leading to confusion and/or double-barreled names”; for example, Mixteco was proposed to replace Mixteca.
The terrane research (e.g., Coney et al. 1980; Howell 1989; Sengör and Dewey 1991; Dewey et al. 1991; Vaughan et al. 2005) has been peculiarly fruitful for the geological knowledge of Mexico. However, some weaknesses in the interpretations result from (1) an exaggeration of subdivisions of each tectonic unit interpreted as distinct terranes; (2) the history and geography, i.e., the delimitations of the terranes in space and times, are very poorly controlled; and (3) local reconstructions and palinspatic maps are largely derivative of the global reconstructions (like those, for example, of Scotese and McKerrow 1990; Dalziel 1997 and Blakey 2008). In this paper, we try to provide revised plate-tectonic reconstructions based on the paleobiogeography of Nuia and primitive foraminifers.
Iapetus Ocean, Rheic Ocean, and Mexican terranes
Acatlán complex and its equivalents
The Acatlán Complex (Rodríguez-Torres 1970; Ortega-Gutiérrez 1981) is unanimously interpreted as the vestige of a Paleozoic ocean, but the identity of this ocean, Iapetus, Rheic or another, and its exact location are debated (Ortega-Gutiérrez et al. 1999; Keppie and Ramos 1999; Nance et al. 2007a, b; Bozkurt et al. 2008; Morales-Gámez et al. 2008; Keppie et al. 2007, Keppie et al. 2008a, b). Long regarded as deposited up to the early Permian, and then structured by the Ouachitan orogenesis (Handschy et al. 1987; Hale-Ehrlich and Coleman 1993; Haenggi 2001; Ross 1991), the Acatlán Complex, and especially its Tecomate Fm, locally includes allochthonous elements of middle Permian age, from middle Wordian to late Capitanian (Vachard et al. 1993, 1997, 2004; Keppie et al. 2004; Sánchez-Zavala et al. 2004). Consequently, their tectonosedimentary history continues at least during the late Permian and/or early Triassic. On the other hand, as no fusulinids of middle Permian age are known in South- and Central America, the carbonate series within the Acatlán Complex correspond to olistolites coming from the North American Craton or to seamounts, insular arcs, or islands situated in the south of this craton but without connections with Perigondwana. The foraminiferal assemblages of the Oaxaquia olistolites are especially similar to those of Texas; not only with regard to the fusulinids but also the smaller foraminifers Abadehella, Neoendothyra and Globivalvulina ex gr. vonderschmittti Reichel, first found in North America in Olinalá (Vachard 1993; Vachard et al. 1993), and then re-found in Texas (Nestell and Nestell 2006; Nestell et al. 2006).
Guayacán group in Sonora
This Group (290 m thick), recorded in Central Sonora (Table 1), corresponds in our opinion to an oceanic series, probably uninterrupted, from Ordovician to uppermost Devonian. The Ordovician graptolites of this ocean in Sonora were discovered by Peiffer-Rangin et al. (1980). The uppermost Devonian was dated with brachiopods by Noll et al. (1984), and in some calciturbidites with kamaenaceans and parathuramminids by our team (unpublished data). Furthermore, many examined microfacies of Guayacán Group are rich in internal molds of radiolarians and confirmed deep-sea and/or oceanic deposits. Distally, the Guayacán Group has probably for equivalent the Marathon Formation in Texas (Young 1970) and the ophiolithic Granjeno Schist of the Sierra Madre Terrane, which overlies a basement of ca. 1 Ga, the Novillo Gneiss (Nance et al. 2007b); therefore contemporaneous of the Grenvillian basement of Laurentia and the Oaxacan Complex of the microcontinent of Oaxaquia.
Oaxaquia microcontinent
Oaxaquia was speculated with the intent to reconstruct a terrane located between Oaxaca and Tamaulipas with a Mesoproterozoic basement of ca. 1 Ga age, the Oaxacan Complex (Ortega-Gutiérrez et al. 1995). It is clear that this terrane did no more exist during the Carboniferous–Permian, because the paleogeography of these systems appears almost similar to the modern geography of Mexico (Vachard et al. 2000a, b), with the Acatlán Complex corresponding to the oceanic bottom, and with narrow carbonate platforms, currently reworked large olistoliths or more or less preserved in situ: San Salvador Patlanoaya (Vachard et al. 2000a, b), Matzizi (Vachard et al. 2000b), Olinalá (Vachard et al. 1993; Vachard 1993; Nestell 1999), San Juan Ixtaltepec (Vachard et al. 1997), and Los Hornos (Vachard et al. 2004; Keppie et al. 2004). It must be pointed out that the Oaxaca terrane of Keppie et al. (1996) and Keppie (2004) differs from Oaxaquia, and this complicates the discussion.
The sedimentary Ordovician series of Oaxaquia (Tiñu Group or Tiñu Formation from the Oaxaca State) completely differs, in its assemblages and geological characters, from that of the northern Mexican states summarized before (Table 1). As indicated by Landing et al. (2007, p. 909): “The Tiñu Formation in Oaxaca State, southern Mexico, provides the only record of fossiliferous Paleozoic rocks south of the Laurentian successions in Sonora and Chihuahua states and Texas and north of the Gondwanan sequences in Andean Colombia and Venezuela”. The Tiñu Formation is subdivided into a lower member, the Yucachica Member (Furongian, upper Cambrian in age), and the Río Salinas Member (lower Ordovician, Tremadocian). Landing et al. (2007) have interpreted the deposits as cold-sea carbonates (due to the presence of a heterozoan association and absence of ooids and evaporites). We agree with the interpretation of a heterozoan association, but not with that of a shallow cold sea. However, Landing et al. (2007, p. 914) highlighted that the accumulation is a “trilobite and echinoderm hash”. This accumulation corresponds more, in our opinion, to a middle ramp deposit, and is therefore independent of the criteria of temperature and depth only applicable to inner ramp deposits (Flores de Dios et al. 1998). This assemblage may perfectly belong to tropical or subtropical shelves, but in deeper paleoenvironments, independent of the possibilities of cool-water carbonates in tropical environments (see James 1997; Samankassou 2002; Soreghan et al. 2008).
On the other hand, the upper part of the Río Salinas “Member”, although of the same marly lithology and color, displays different tectonic structures from the lower part (personal observation near the Santiago Ixtaltepec village, Guerrero, Mexico), which may correspond either to a slumping or to two different tectonic slices of different ages. Its boundary with the overlying Tournaisian Santiago Formation is also unclear. Consequently, our goal consists in the discovery of the age of the upper part of the Río Salinas “Member” and its exact limit with the overlying formation which was dated as late Tournaisian by our team. Therefore, the age of the upper part of the Río Salinas “Member” should be reassessed and the formation renamed as the current name is pre-occupied in Mexico. However, the most important problem to solve remains the detailed paleogeography of this sector of Oaxaca, because very different paleogeographic affinities were proposed by previous authors (see below).
Prior to the Carboniferous, Oaxaquia might have existed, but it is currently impossible to demonstrate if (1) Oaxaquia was the oceanic bottom of the Iapetus’ northern margin and became parautochthonous; or (2) Oaxaquia was the oceanic bottom of the Iapetus’ southern margin, initially connected with Gondwana, and then displaced. It is possible that the oceanic bottoms of Iapetus and Rheic occupy the same areas in Mexico and were closed and opened exactly in the same areas, because the Pennsylvanian-early Permian Rheic paleogeography of Sonora (e.g., Bozkurt et al. 2008; Nance et al. 2010; Palafox 2011) existed already during the early Ordovician; and despite of the wandering of the North American craton (Cocks and Torsvik 2011, Fig. 3a–c), Sonora remained always at the extremity of this continent, and always remained the limitroph of an oceanic area. Therefore, in Sonora, the limits of the carbonate ramps and the oceanic deposits are the same during the early Ordovician (Fig. 19) as they were during the Carboniferous–Permian (see Palafox 2011), and the Iapetus and Rheic oceans had the same limits and the same extent across Sonora during the Paleozoic. This renewed paleogeography probably explains why the Acatlán Complex was first considered as having been deposited on the Iapetus and then on the Rheic oceanic bottom (Ortega-Gutiérrez et al. 1999; Nance et al. 2007a, b).

Proposed interpretations and reconstructions
Paleobiogeographical value of Nuia
The photophile and warm stenotherm characteristics of the cyanobacteria Nuia, and its frequent association with the Ordovician reefs permits us to indicate a tropical distribution for Nuia, even if the reefal ecosystems are very rarely represented during the Ordovician and only encountered in Mexico, USA, Canada and Siberia. In Mexico, the reefs built by corals, bryozoans and/or microbialites, form bars that protected lagoons where the first calcareous chlorophytes like bryopsidales and dasycladales developed, in a photozoan association with gastropods, bivalves, and the first orthoceratids (all these groups possess an aragonitic mineralization). The accumulations located in the reef-front are dominated by pelmatozoan remains (Echinodermata). In this early Ordovician tropical zone, the Nuia Province as defined here, was located on each part of the Equator [see the reconstructions of Roux (1991a); Poole et al. (1995a); Matte (2001) and von Raumer et al. (2003)]. The principal distribution areas of Nuia encompass two sectors, one from Sonora (Mexico) to Quebec (Canada) as well the western side of the North American craton; another one from Kazakhstan up to Siberia (Russia) (Gnoli and Serpagli 1980; Vachard and Téllez-Girón 1986) (Fig. 20a). The first sector corresponds exactly to the margins of Laurentia, as reconstructed for a long time (Cook et al. 1975; Petersen et al. 1976; Fig. 19). The specimens of Nuia from Greenland (Arctida) connect these two subprovinces or continental blocks and confirm that the three paleocontinents, Laurentia, Arctida and Siberia, were located in the early Ordovician tropical zones (Fig. 20b), with narrow connections, probably via the terranes Barentsia, Verkhoyansk, Alaska-Chukotka and Farewell (Golonka 2002; Torsvik and Cocks 2013a).

Paleoprovince with Nuia. a Distribution in a modern geography (modified after Vachard and Téllez-Girón 1986). b The same distribution in a paleomap after Golonka (2002), with distance between the South China-Indochina and Gondwana modified (see text) and illustration of the Nuia Province. The stars in a and b indicate the new report from the studied area
By contrast, (1) the distribution of coeval taxa of Nuia, Archaeoscyphia and Calathium for example, seems to be broader and includes North America, Argentina, northern Europe, Australia and China (Finks and Rigby 2003; with references therein for Archaeoscyphia; and Nitecki et al. 1999, for Calathium); this broader distribution is probably related to larval stages which did not exist in Nuia; and (2) the distribution of the first leperditicopids might be restricted to the western border of Laurentia (in its lower Ordovician location), in Sonora, and the southern border with the San Juan, Newfoundland and Svarlbard localities indicated by Williams and Silvester (2008), but re-positioned in the paleomaps of Cocks and Torsvik (2011) and Torsvik and Cocks (2013b).
Therefore, several paleomaps of Dalziel et al. (1994), Keppie and Ortega-Gutiérrez (1995), Astini et al. (1995), Dalziel (1997), Golonka (2002), von Raumer et al. (2003), Cocks and Torsvik (2011), and Torsvik and Cocks (2013b), reflect the biogeographic distribution of Nuia and, among these paleomaps, the palinspatic Earth reconstruction of Golonka (2002) is the most adequate (Fig. 20b). The maps of Bozkurt et al. (2008), Sengör et al. (2014) or Golonka (2002) indicates that the Kipchak arc is not yet connected to the Russian Craton (i.e., western Laurentia) during the early Ordovician, while it will be connected during the middle Ordovician (Sengör et al. 2014, fig. 8a, b), permitting possible connections between Siberia and Laurentia. Tarim is probably connected with the South China terranes, where Nuia is relatively poorly known; nevertheless, it was occasionally mentioned, whereas Archaeoscyphia, Calathium and Pulchrilammina are more commonly described (e.g., Adachi et al. 2011a, b; Li et al. 2014, 2015). Furthermore, Tarim and the South China terranes can be connected via the Anamia (=Indosinia/Indochina terrane) and the Mongolian (=Kazakhstanian) terranes of Torsvik and Cocks (2013a). The Chu-Ilii terrane is located on one of the Kazakhstania terranes sometimes placed near the Equator during the early Ordovician (Popov et al. 2013, fig. 10.1). Moreover, the distance between the South China-Indochina (= Indosinia = Anamia) and Gondwana paleocontinent is probably broader than indicated by Golonka (2002) or Torsvik and Cocks (2013b). In contrast, Baltica and Avalonia are related to cold temperate to subpolar waters and are separated from the Nuia Province by a broad branch of the Iapetus.
Harper et al. (2013) have defined a low-latitude brachiopod Province. This is larger than the Nuia Province defined here, because it includes the northeastern part of Gondwana (Sibumasu, Australia, eastern part of India, autochthonous Argentina, Chile, etc.), where Nuia is absent. Although autochthonous Argentina and Chile are effectively located in the tropical zone, they are devoid of Nuia, whereas the Precordillera-Argentina San Juan terrane of these authors (with Nuia) is located near the North American Craton. We agree with this map, nevertheless, due to the probable continuity of Laurentia and Siberia (as in the paleomap of Golonka 2002, fig. 5, for example), we bring together these two continents and we take away South Chile- autochthonous Argentina from Australia-Sibumasu.
For the Precordillera Terrane and autochthonous Argentina, the reconstructions of Astini et al. (1995), Dalziel (1997), Keller (1999, 2012), and Harper and Servais (2013) are probably the best for understanding (a) the paleolocation of its source in Laurentia; (b) the migration way of the terrane; and then (c) the location of the transform faults affecting this terrane.
Due to the paleobiogeographical comparisons suggested by Robison and Pantoja-Alor (1968) with the Argentina Precordillera, a similar Laurentian origin might be suggested for the Tiñu Terrane, preferentially to other hypotheses like: (1) a migration from Argentina to Mexico (as initially admitted by Robison and Pantoja-Alor 1968); (2) an origin from the Sierra Garzón in Colombia (as proposed in numerous papers by Keppie); and (3) the connection between Tiñu and the paleocontinent of Baltica, recently proposed by Landing et al. (2007). The first hypothesis seems to be the more convincing, because other terranes have migrated from South America to North America (see Keppie et al. 2003), displaced across South America such as the Madre de Dios terrane (southern Chile; see e.g., Ramos 1988; Hervé 1993), the fusulinids of which are related with those of Titica Lake (Peru-Bolivia border) or those of North America. We can suggest that, if Sonora shares the paleobiogeography of the North American Craton, other areas of Mexico belong clearly to another province (supercontinent or microcontinent): i.e., the peri-Gondwanan border of the Rheic ocean with affinities to Argentina or to the Oaxaquia, a block isolated within the Rheic between Laurentia and Gondwana (Dalziel 1997; Murphy et al. 2006; Landing et al. 2007; Keppie et al. 2008a, b). Oaxaquia and the Chortis and Maya blocks were interpreted as parts of the southern margin of the Rheic Ocean and show a paleobiogeographic affinity to the paleocontinent of Gondwana, or more probably a location between the two Rheic margins. Moreover, the initial location and its definitive paleogeographic emplacement remain discussed because the geological architecture of Mexico is poorly known and very complex.
Conclusions
-
1.
The Cerro San Pedro succession (Sonora, NW Mexico) exhibits the classical microfacies in the El Paso Formation of the southern North American Craton. The northernmost outcrops of Mexico (Sonora and Chihuahua) expose some of the oldest ecosystems with cnidarian (corals), bryozoan, recptaculitales and leperditicopids from the Earth history.
-
2.
The reefal ecosystems, rarely represented during the Ordovician, and mainly known from the USA, Canada and Siberia are in Mexico only known as allochthonous clasts in bioclastic tempestites.
-
3.
In the San Pedro section, the marine environments appear shallow and subtropical; and the assemblage is photozoan to heterozoan. Numerous tempestites are indicated by the accumulation of microperforated extraclasts or pelmatozoan ossicles.
-
4.
An early Ordovician age is based on the occurrence of the cyanobacteria Nuia sibirica. The chlorophyte algae remain questionable during this period; the true Dasycladales, Chaetoporales and Bryopsidales appeared in the middle–late Ordovician. Consequently, the receptaculitales Calathium, as well as the so-called Cambrian dasycladales, most probably belong to other algal groups. On the other hand, the Vermiporellaceae, often interpreted as the first dasycladales, are more probably chaetophorales. Only one ellianacean (red? algae) was observed.
-
5.
The oldest foraminifers of North America have been re-found during this study; moreover, they are the first Ordovician foraminifers mentioned in Mexico, with the genera Neoarchaesphaera, Vicinesphaera and Rauserina.
-
6.
All these taxa have a dark-microgranular wall and not an agglutinated wall.
-
7.
Macrofaunal fragments are classical in the El Paso Formation, especially of the Archaeoscyphia sponges, the gastropods and the sponge spicules. The leperditicopids are abundant and perhaps have their FAD in Sonora.
-
8.
The endolithic perforations of the leperditicopids of Sonora need further detailed studies.
-
9.
The cyanobacteria Nuia is considered here as a paleogeographic marker of the western province in the tropical belt during the early Ordovician. The studied area in Sonora is considered as the westernmost part of the Nuia Province.
-
10.
The Sonora shares the paleobiogeography of the North American Craton. Other areas of Mexico belong clearly to another province. As probable parts of the northern margin of the Iapetus ocean, they have then migrated toward the Perigondwanan border; i.e., the southern margin of the Rheic ocean, and shared affinities with Argentina or blocks isolated within the Rheic between Laurentia and Gondwana. The subcontinent Oaxaquia and the Chortis and Maya blocks, which were interpreted as parts of the southern margin of Rheic ocean, probably possess paleobiogeographic affinities with the paleocontinent of Gondwana or more probably a location between the two Rheic margins. Moreover, the initial location and its definitive paleogeographic emplacement remain discussed because of the very complex geological architecture of Mexico.
References
Adachi N, Ezaki Y, Liu Jian-Bo (2011a) Early Ordovician stromatoporoid Pulchrilamina spinosa from South China: Significance and implications for reef development. In: Aretz M., Delculée S., Denayer J., Poty E. (eds) In: Abstracts, 11th Symposium on Fossil Cnidaria and Sponges, Liège, August 19–29, 2011 Kölner Forum Geol Paläont 19:6–7
Adachi N, Jian-Bo Liu, Ezaki Y (2011b) Early Ordovician shift in reef construction from microbial to metazoan reefs. Palaios 26:106–114
Ahr WM (1971) Paleoenvironment algal structures and fossil algae in the Upper Cambrian of central Texas. J Sediment Petrol 41(1):205–216
Albertstadt L, Repetski JE (1989) A Lower Ordovician Sponge/Algal Facies in the Southern United States and its Counterparts Elsewhere in North America. Palaios 4:225–242
Almazán-Vázquez E (1989) El Cámbrico-Ordovícico de Arivechi, en la región centrooriental del estado de Sonora. Rev Inst Geol Univ Nal Autón Méx 8(1):58–66
Almazán-Vázquez E, Fernández-Aguirre MA 1987. Los terrenos paleozoicos de la región serrana de Sonora. In: Paleozoico de Chihuahua. Sociedad Geológica Mexicana, La Gaceta Geológica 1(1):97–104
Almazán-Vázquez E, Buitrón-Sánchez B, Franco-Vega O (2006) Formación Pozo Nuevo: una secuencia litoestratigráfica de plataforma del Ordovícico Temprano de la región central de Sonora, Mexico. Rev Mex Cienc Geol 23(1):23–38
Amsden TW, Toomey DF, Barrick JE (1980) Paleoenvironment of Fitzhugh Member of Clarita Formation (Silurian, Wenlockian) Southern Oklahoma. Okla Geol Surv Circ 83:1–54
Antropov IA (1950) New Late Devonian foraminiferal species from some regions of the eastern Russian Platform. Akad nauk SSSR, Izvest Kazan Fil, Geol Inst 1, ser geol, p 21–33 (in Russian)
Armstrong AK, Mamet BL (1977) Carboniferous microfacies, microfossils, and corals, Lisburne Group, Arctic Alaska. Geol Surv Prof Paper 849:1–144
Astini RA (2001) Nuia y Girvanella a través de la transición cambro-ordovícica (Formación Volcancito) en el Famatina: significado paleoambiental y paleogeográfico. Ameghiniana 38(3):243–255
Astini RA, Benedetto JL, Vaccari NE (1995) The early Paleozoic evolution of the Argentine Precordillera as a Laurentian rifted, drifted and collided terrane: a geodynamic model. Geol Soc Am Bull 107(3):253–273
Ausich WI, Kammer TW, Rhenberg EC, Wright DF, Smith A (2015) Early phylogeny of crinoids within the pelmatozoan clade. Palaeontology 58(6):937–952
Berdan JM (1984) Leperditicopid ostracodes from Ordovician rocks of Kentucky and nearby states and characteristic features of the order Leperditicopida. Geol Surv Prof Paper 1066-J:J1–J40
Beresi MS (1984) Dispersión geográfica y estratigráfica de Nuia (microorganismo algal) en la Precordillera de San Juan. Memoria III Congreso Latinoamericano de Paleontología, 14–18 Octubre, Oaxtepec, México, p 73–83
Beresi MS, Cabaleri NG, Buitrón-Sánchez BE, Rodríguez MC, Heredia SE, Franco M, Tortello MF (2012) Microfacies, biota y paleoambientes sedimentarios del Ordovícico temprano-medio del Cerro Salazar, Sonora central, Mexico. Rev Mex Cienc Geol 29(2):330–345
Blakey RC (2008) Gondwana palaeogeography from assembly to break-up—A 500 m.y. odyssey. In: Fielding CR, Frank TD, Isbell JL (eds) Resolving the Late Paleozoic Ice Age in Time and Space. Geol S Am S 441:1–28
Bourque PA, Mamet B, Roux A (1981) Algues siluriennes du synclinorium de la Baie des Chaleurs, Québec, Canada. Rev Micropal 24(2):83–126
Bozkurt E, Pereira MF, Strachan R, Quesada C (2008) Evolution of the Rheic Ocean. Tectonophysics 461:1–8
Brady HB (1879) Notes on some reticularian Rhizopoda of the Challenger Expedition. Part III. 1. Classification. 2. Further notes on new species. 3. Note on Biloculina mud. Quart J Micro Sci N Ser 21:31–71
Brazhnikova NE, Rostovceva LP (1966) Foraminifera. In: Aizenverg DE (ed) Fauna of the lowest Tournaisian of C t1 a the Donetz basin. Akad nauk Ukr SSR, Inst Geol Nauk, pp 9–42 (in Russian)
Bridges LW (1964) Stratigraphy of Mina Plomosas-Placer de Guadalupe area. In: Geology of Mina Plomosas-Placer de Guadalupe area, Chihuahua, Mexico. West Texas Geological Society, Field Trip Guidebook Publication 64–50, p 50–59
Aasgard U, Bromley, RG (1991) Population dynamics and autecology of “Rhynchonella triangularis”, a Late Cretaceous rocky coast brachiopod. In: MacKinnon DI, Lee DE, Campbell JD (eds) Brachiopods through time. In: Proceedings of the 2nd International Brachiopod Congress, University of Otago, Dunedin, New Zealand, 5–9 February 1990, Balkema Rotterdam, p 247–252
Browne RG, Pohl ER (1973) Stratigraphy and genera of calcareous Foraminifera of the Fraileys facies (Mississippian) of central Kentucky. Bull Am Paleontol 64(280):173–243
Brunner P (1975) Estudio estratigráfico del Devónico en el área de El Bisani, Caborca, Sonora. Rev Inst Mex Petról 7(1):16–45
Brunner P (1984) Los conodontos de México. Memoria del Tercer Congreso Latinoamericano de Paleontología, Oaxtepec, México, p 84–91
Bubik M (2001) Foraminifera in the Middle Cambrian of the Barrandian area (Czech Republic). J Czech Geol Soc 46(3–4):195–198
Buitrón BE, Rivera-Carranco E (1984) Lingúlidos (Brachiopoda-Inarticulata) del Ordovícico de Oaxaca, México. Memoria, III Congreso Latinoamericano de Paleontología, Oaxtepec, México, p 54–61
Buitrón-Sánchez BE, Vachard D, Almazán-Vázquez E, Palafox JJ (2012) Una secuencia cratónica completa del Carbonífero al Pérmico inferior expuesta en los cerros El Tule, noreste de Sonora, México. Rev Mex Cienc Geol 29(1):39–62
Burchette TP, Wright VP (1992) Carbonate ramp depositional systems. Sediment Geol 79:3–57
Bykova EV, Polenova EN (1955) Devonian foraminifers and radiolarians from the Volgo-Urals and of the Central Devonian field and their stratigraphical importance. Trudy VNIGRI 87:1–141 (in Russian)
Campa MF, Coney PJ (1983) Tectonostratigraphic terranes and mineral resource distributions in Mexico. Can J Earth Sci 20:1040–1051
Carozzi AV (1989) Carbonate rock depositional models; a microfacies approach. Advanced reference Series, Physical and Life Sciences, Prentice Hall, Upper Saddle River, New Jersey, p 604
Cavalier-Smith T (2002) The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int J Syst Evol Micr 52:297–354
Chuvashov BI (1965) Upper Devonian foraminifers and algae from the western slope of the Central and Southern Urals. Akad Nauk SSSR, Ural Filial, Trudy Inst Geol 74:1–93 (in Russian)
Clausen S, Smith AB (2008) Stem structure and evolution in the earliest pelmatozoan echinoderms. J Paleontol 82:737–748
Clausen S, Jell PA, Legrain X, Smith AB (2009) Pelmatozoan arms from the Middle Cambrian of Australia: bridging the gap between brachioles and brachials? Lethaia 42:283–296
Clemons RE (1991) Petrography and depositional environments of the Lower Ordovician El Paso Formation. N M Bur Mines Miner Resour 125:5–66
Cloud PE Jr, Barnes VE (1948) The Ellenburger Group of central Texas. Univ Tex Publ 4621:1–473
Cocks LRM, Torsvik TH (2011) The Palaeozoic geography of Laurentia and western Laurussia: a stable craton with mobile margins. Earth Sci Rev 106:1–51
Coney PJ, Jones DL, Monger JWH (1980) Cordilleran suspect terranes. Nature 288:329–333
Conkin JE, Conkin BM (1979) North American Ordovician agglutinate foraminifera. University of Louisville, Studies in Paleontology and Stratigraphy 8:1–24
Conkin JE, Conkin BM, Canis WF (1968) Mississippian foraminifera of the United States. Part 3. The limestones of the Chouteau Group in Missouri and Illinois. Micropaleontology 14:133–178
Conoglio M, James N (1985) Calcified algae as sediment contributors to early Paleozoic limestones: evidence for deep-water sediments of the Cow Head Group, western Newfoundland. J Sediment Petrol 55(5):754–764
Cook TD, Bally AW (with the collaboration of Milner S, Buffler RT, Farmer RE, Clark DK, Heuer WC, Loofs JW, Laverde JF, Villarreal HG Jr.) (1975) Stratigraphic Atlas of North and Central America. Princeton University Press, Princeton New Jersey, p 1–272
Cooks HE, Taylor ME (1977) Comparison of continental slope and shelf environments in the Upper Cambrian and lowest Ordovician of Nevada. SEPM Sp P 25:51–81
Culver SJ (1991) Early Cambrian Foraminifera from West Africa. Science 254:689–691
Culver SJ (1994) Early Cambrian Foraminifera from the southwestern Taoudeni Basin, West Africa. J Foramin Res 24:191–202
Culver SJ, Repetski JE, Pojeta J Jr, Hunt D (1996) Early and Middle (?) Cambrian metazoan and protistan fossils from West Africa. J Paleontol 70(1):1–6
Cushman JA, Waters JA (1928) Some Foraminifera from the Pennsylvanian and Permian of Texas. Contrib Cushman Lab Foram Res 4(2):31–55
D’Orbigny A (1826) Tableau méthodique de la classe des Céphalopodes. Ann Sci Nat 7(2):245–314
Dalziel IWD (1997) Neoproterozoic–Paleozoic geography and tectonics: review, hypothesis, environmental speculation. Geol Soc Am Bull 109(1):16–42
Dalziel IWD, Dalla Salda LH, Gahagan LM (1994) Paleozoic Laurentia–Gondwana interaction and the origin of the Appalachian–Andean mountain system. Geol Soc Am Bull 106:243–252
Derby JR., Raine RJ, Runkel AC, Smith MP (2012) Paleogeography of the great American carbonate bank of Laurentia in the earliest Ordovician (early Tremadocian): The Stonehenge transgression. In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:5–13
Derycke-Khatir C, Vachard D, Degardin JM, Flores de Dios A, Buitrón B, Hansen M (2005) Late Pennsylvanian and Early Permian chondrichthyan microremains from San Salvador Patlanoaya (Puebla, Mexico). Geobios 38:43–55
Devuyst FX (2006) The Tournaisian–Viséan boundary in Eurasia. Definition, biostratigraphy, sedimentology and early evolution of the genus Eoparastaffella (foraminifer). Thèse Univ Catho Louvain, p 1–430
Dewey JF, Gass IG, Curry GB, Harris NBW, Sengör AMC (1991) Allochthonous terranes. Cambridge Univ Press, Cambridge, pp 1–199
Dickinson WR, Lawson TF (2001) Carboniferous to Cretaceous assembly and fragmentation of Mexico. Geol Soc Am Bull 1113(9):1142–1160
Donovan SK (1986) Pelmatozoan columnals from the Ordovician of the British Isles, Part 1. MonogrPalaeontogr Soc 568:1–68
Dubatolov VN, Krasnov VI, Bogush OI, Zadorozhnyi VM, Stepanov CA, Ratanov LS, Bidzhakov VI, Zapivalov NP, Serdyul ZYa, Mukhina IP (1985) Stratigraphy of the Palaeozoic of the southeastern part of the West-Siberian Plain. In: Dubatolov VN, Kanygin AV (eds) Biostratigraphy of the Palaeozoic of the western Siberia. Akad nauk SSSR, Sibirsk Otdel, Tr Inst Geol Geofiz 619:4–49 (in Russian)
Dunn PH (1942) Silurian Foraminifera of the Mississippi Basin. J Paleontol 16(3):317–342
Eguizábal-Martínez FJ (1988) Estudio estratigráfico-sedimentológico de las rocas del Paleozoico en el prospecto Agua Prieta, estados de Sonora y Chihuahua. Inst Mex Petrol, Subdir Tecno Explor, Proyecto C-3020:1–152
El Albani A, Vachard D, Fürsich F, Buitrón B, Flores de Dios A (2005) Depositional environment and biofacies characterization of the Upper Pennsylvanian–Lower Permian deposits of the San Salvador Patlanoaya section (Puebla, Mexico). Facies 50:629–645
Elliott GF (1972) Lower Palaeozoic green algae from Southern Scotland and their evolutionary significance. Bull Br Mus (Nat Hist) Geology 22(4):357–377
Taylor JF, Myrow PM, Ripperdan RL, Loch JD, Ethington RL (2004) Paleoceanographic events and faunal crises recorded in the Upper Cambrian and Lower Ordovician of west Texas and southern New Mexico. In: Nelson EP, Erslev EA (eds) Field Trips in the Southern Rocky Mountains, USA. GSA Field Guide 5:169–185
Ethington RL, Clark DL (1964) Conodonts from the El Paso Formation (Ordovician) of Texas and Arizona. J Paleontol 8(4):685–704
Ethington RL, Clark DL (1971) Lower Ordovician conodonts in North America. In Sweet WC, Bergström SM (eds), Symposium on conodont biostratigraphy. Geol Soc Am Mem 127:63–82
Finks RM, Rigby JK (2003) Geographic and stratigraphic distribution. In Rigby JK (ed) Treatise on invertebrate paleontology, Part E, Porifera Revised, vol 2, Introduction to the Porifera. The Geological Society of America and the University of Kansas Press, pp 275–296
Flores de Dios LA, Vachard D, Buitrón-Sánchez BE (1998) The Tiñu, Santiago-Ixtaltepec and Yododeñe Fms, Oaxaca State: sedimentological, stratigraphic and paleogeographic reinterpretations. IIn: GCP Project 376, Laurentia–Gondwana connections before Pangea, Program and Abstracts, Oaxaca City, Oaxaca, Mexico, p 16
Flügel E (2004) Microfacies of carbonate rocks, analysis, interpretation and application. Springer Publisher, Berlin, pp 1–976
Flügel E (2010) Microfacies of carbonate rocks, analysis, interpretation and application. Springer Publisher, Berlin, pp 1–984
Fritz RD, Medlock P, Kuykendall MJ, Wilson JL (2012) The geology of the Arbuckle Group in the midcontinent: Sequence stratigraphy, reservoir development, and the potential for hydrocarbon exploration. In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:203–273
Fursenko AV (1958) Fundamental steps of development of foraminiferal fauna in the geological past. Akad nauk Beloruskoi SSR, Instit Geol nauk, Trudy 1:10–29 (in Russian)
Gaillot J, Vachard D (2007) The Khuff Formation (Middle East) and time-equivalents in Turkey and South China: biostratigraphy from Capitanian to Changhsingian times (Permian), new foraminiferal taxa, and palaeogeographical implications. Coloquios de Paleontología 57:37–223
Garcia-Cortez JA, Hernandez-Morales P (2010) Carta geológico-minera, San Pedro de la Cueva (H12-D34). Servicio Geológico Mexicano, Sonora
Gektidis M, Dubinsky Z, Goffredo S (2007) Microendoliths of the shallow euphotic zone and shaded habitats at 30°N - Eilat, Israel—palaeoecological implications. Facies 53:43–55
Gerhard LC (1972) Canadian depositional environments and paleotectonics, central Colorado. In De Voto H (ed) Paleozoic stratigraphy and structural evolution of Colorado. Quart Colorado Sch Mines 67(4):1–36
Gnilovskaya MB (1972) Calcareous algae from Middle/Late Ordovician of eastern Kazakhstan. Akad nauk SSSR, Inst Geol Geokhron Dokembriya, Izdatelstvo “Nauka”, p 1–196 (in Russian)
Gnoli M, Serpagli E (1980) The problematical microorganism Nuia in the Lower Ordovician of Precordilleran Argentina and its paleogeographic significance. J Paleontol 5(6):1245–1251
Golonka, J (2002) Plate-tectonic maps of the Phanerozoic. In: Kiessling W, Flügel E, Golonka, J (eds) Phanerozoic reef patterns. SEPM Spec P 72:21–75
Golubic S, Perkins RD, Lukas KJ (1975) Boring microorganisms and microborings in carbonate substrates. In: Frey RW (ed) The study of trace fossils. Springer Verlag, Berlin, pp 229–259
Golubic S, Radtke G, Le Campion-Almusard T (2005) Endolithic fungi in marine ecosystems. Trends Microbiol 13:229–235
Gómez-Espinosa C, Vachard D, Buitrón-Sánchez B, Almazán-Vázquez E, Mendoza-Madera C (2008) Pennsylvanian fusulinids and calcareous algae from Sonora (Northwestern Mexico), and their biostratigraphic and paleobiogeographic implications. C R Palevol 7(5):259–268
Gregg JM, Shelton KL (2012) Mississippi Valley-type mineralization and ore deposits in the Cambrian–Ordovician great American carbonate bank. In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:161–185
Guensburg TE, Mooi R, Sprinkle J, David B, Lefebvre B (2010) Pelmatozoan arms from the mid-Cambrian of Australia: bridging the gap between brachioles and brachials? Comment: there is no bridge. Lethaia 43:432–440
Guilbault JP, Hubert C, Mamet B (1976) Nuia et Halysis, deux algues ordoviciennes énigmatiques des Basses-Terres du Saint-Laurent. Nat Can 103(2):119–132
Gutschick RC (1962) Arenaceous foraminifera from oncolites in the Mississippian Sappington Formation. J Paleontol 36:1291–1304
Gutschick RC (1986) Middle Ordovician agglutinated foraminifera including Reophax from the Mifflin Formation, Platteville Group of Illinois. J Palaeont 60:233–248
Gutschick RC, Treckman JF (1959) Arenaceous foraminifera from the Rockford Limestone of northern Indiana. J Paleontol 23(2):229–250
Haenggi WT (2001) Tectonic history of the Chihuahua through, Mexico and adjacent USA, part I: the pre-Mesozoic setting. Bol Soc Geol Mex 54:28–66
Hale-Ehrlich WS, Coleman JL Jr (1993) Ouachita–Appalachian juncture: a Paleozoic transpressional zone in the Southeastern USA. AAPG Bull 77(4):552–568
Handschy JW, Keller GR, Smith KJ (1987) The Ouachita system in Northern Mexico. Tectonics 6(3):323–330
Harper DAT, Servais T (2013) Early Palaeozoic Biogeography and Palaeogeography. Geol Soc Mem 38:1–490
Harper DAT, Rasmussen CMØ, Liljeroth, M, Blodgett RB, Candela Y, Jin J, Percival IG, Roug J-Y, Villas E, Zhan RB (2013) Biodiversity, biogeography and philogeography of Ordovician rhynchonelliform brachiopods. In: Harper DAT, Servais T (eds) Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London Memoir 38:127–144
Hayes PT (1972) Stratigraphic nomenclature of Cambrian and Lower Ordovician rocks of easternmost southern Arizona and adjacent westernmost New Mexico. Geol Surv Bull 1372-B:B1–B21
Héroux Y, Hubert C, Mamet B, Roux A (1977) Algues siluriennes de la Formation de Sayabec (Lac Matapédia, Québec). Can J Earth Sci 14(12):2865–2908
Hervé F (1993) Paleozoic metamorphic complexes in the Andes of Aysén, southern Chile (west of?Occidentalia). In: Proceedings of the first circum-pacific and circum-Atlantic Terrane Conference, 5–22 November 1993, Guanajuato (Mexico), p 64–65
Høeg OA (1932) Ordovician algae from the Trondheim area. In: Kiaer J (ed) The Hovin Group in the Trondheim area, II. Paleontological part. Skrifterutgitt av det Norske Videnskaps-Akademi i Oslo 1, Mat.-naturv. Klasse 1927(4):63–96
Holcová K, Slavík L (2013) The morphogroups of small agglutinated foraminifera from the Devonian carbonate complex of the Prague Synform (Barrandian area, Czech Republic). Palaeogeogr Palaeocl 386:210–224
Howell DG (1989) Tectonics of suspect terranes: mountain building and continental growth. Springer, Netherlands, 232p
Huang ZC, Bian LZ (1983) Discovery of the Ordovician alga Nuia siberica [sic] in China. Acta Paleontol Sin 22(1):61–64
Ince D (1984) Sedimentation and tectonism in the Middle Ordovician of the Girvan district, SW Scotland. Trans R Soc Edinb Earth Sci 75:225–237
Ireland HA (1939) Devonian and Silurian foraminifera from Oklahoma. J Paleontol 13(2):190–202
James NP (1997) The cool-water carbonate depositional realm. In: James NP, Clark JAD (eds) Cool-water carbonates. SEPM Spec P 56:1–20
Johnson JH (1966) The Late Cambrian algal genus Nuia from Brewser County, Texas. J Paleontol 40(2):433–434
Kazmierczak J (1976) Volvocacean nature of Paleozoic non-radiosphaerid calcispheres and parathuramminid “foraminifera”. Acta Paleontol Pol 21(3):245–258
Kazmierczak J, Kremer B (2005) Early post-mortem calcified Devonian acritarchs as a source of calcispheric structures. Facies 51:573–584
Keller M (1999) Argentine Precordillera: sedimentary and Plate Tectonic History of Laurentian Crustal Fragment in South America. Geol Soc Am Spec Papers 341:1–131
Keller M (2012) The Argentine Precordillera: A Little American Carbonate Bank. In Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:985–1000
Keppie JD (2004) Terranes of Mexico revisited: a 1.3 billion year odyssey. Int Geol Rev 46:765–794
Keppie JD, Ortega-Gutiérrez F (1995) Provenance of Mexican Terranes: isotopic constraints. Int Geol Rev 37:813–824
Keppie JD, Ramos VA (1999) Odyssey of terranes in the Iapetus and Rheic Oceans during the Paleozoic. In: Keppie JD, Ramos VA (eds) Laurentia–Gondwana Connections Before Pangea. Geol S Am S 336:267–276
Keppie JD, Dostal J, Murphy JD, Nance RD (1996) Terrane transfer between eastern Laurentia and western Gondwana in the early Paleozoic. Constraints and global reconstructions. In: Nance RD, Thompson, MD (eds) Avalonian and related Peri-Gondwanan Terranes of the Circum-North Atlantic. Geol S Am S 304:369–380
Keppie JD, Nance RD, Murphy JB, Dostal J (2003) Tethyan, Mediterranean, and Pacific analogues for the Neoproterozoic–Paleozoic birth and development of peri-Gondwanan terranes and their transfer to Laurentia and Laurussia. Tectonophysics 365:195–219
Keppie JD, Sandberg CA, Miller BV, Sánchez-Zavala JL, Nance RD, Poole FG (2004) Implications of Latest Pennsylvanian to Middle Permian Paleontological and U-Pb SHRIMP Data from the Tecomate Formation to Re-dating Tectonothermal Events in the Acatlán Complex, Southern Mexico. Int Geol Rev 46:745–753
Keppie JD, Dostal J, Elías-Herrera M (2007) Ordovician-Devonian oceanic basalts in the Cosoltepec Formation, Acatlán Complex, southern México: Vestiges of the Rheic Ocean. In: Linnemann U, Nance RD, Kraft P, Zulauf G (eds) The evolution of the Rheic Ocean in North America: From Avalonian-Cadomian active margin to Alleghanian-Variscan collision. Geological Society of America Special Paper 423, pp 477–487
Keppie JD, Dostal J, Miller BV, Ramos-Arias MA, Morales-Gámez M, Nance RD, Murphy JB, Ortega-Rivera A, Lee JWK, Housh T, Cooper P (2008a) Ordovician–earliest Silurian rift tholeiites in the Acatlán Complex, southern Mexico: evidence of rifting on the southern margin of the Rheic Ocean. Tectonophysics 461:130–156
Keppie JD, Dostal J, Murphy JD, Nance RD (2008b) Synthesis and tectonic interpretation of the westernmost Paleozoic Variscan orogen in southern Mexico: from rifted Rheic margin to active Pacific margin. Tectonophysics 461(1–4):277–290
Ketner KB (2013) Stratigraphy of Lower to Middle Paleozoic rocks of Northern Nevada and the Antler. US Geol Surv Prof Paper 1799:1–18
Klement KW, Toomey DF (1967) Role of the blue-green alga Girvanella in skeletal grain destruction and lime-mud formation in the Lower Ordovician of west Texas. J Sediment Petrol 37:1045–1051
Kobluk DR, Kahle CF (1977) Bibliography of the endolithic (boring) algae and fungi and related geologic processes. B Can Petrol Geol 25(1):220–223
Kordé KB (1973) Vodorosli Kembriya (Cambrian algae). Akad nauk SSSR, Trudy Paleontol Instit 139:1–34
Kouchinsky A, Bengtson S, Clausen S, Gubanov A, Malinky JM, Peel JS (2010) A Middle Cambrian fauna of skeletal fossils from the Kuonamka Formation, northern Siberia. Alcheringa, p 1–67
Kouchinsky A, Bengtson S, Clausen S, Vendrasco MJ (2015) An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontol Pol 60(2):421–512
Kulagina E (2013) Taxonomic diversity of foraminifers of the Devonian–Carboniferous boundary interval in the South Urals. Bull Geosc 88(2):265–282
Landing E, Westrop SR, Keppie JD (2007) Terminal Cambrian and lowest Ordovician succession of Mexican West Gondwana: biotas and sequence stratigraphy of the Tiñu Formation. Geol Mag 144(6):909–936
Langer W (1969) Foraminifera aus dem Alt-Paläozoikum der Karnischen Alpen. Sonderdrück Carinthia II 79(159):34–60
LeMone DV (1969) Lower Paleozoic rocks in the El Paso area. In: Cordoba DA, Wengerd SA, Shomaker JW (eds.) In: The Border Region (Chihuahua, Mexico, & USA), New Mexico Geological Society 20th Annual Fall Field Conference Guidebook, p 68–79
Li QJ, Li Y, Kiessling W (2014) Early Ordovician sponge-Calathium-microbial reefs on the Yangtze Platform margin of the South China Block. GFF Upps 136(1):157–161
Li QJ, Li Y, Wang JP (2015) Kiessling W (2015), Early Ordovician lithistid sponge-Calathium reefs on the Yangtze Platform and their paleoceanographic implications. Palaeogeogr Palaeoclimatol Palaeoecol. doi:10.1016/j.palaeo.2015.02.034
Lipps JH (1992) Origin and early evolution of foraminifera. Studies in Benthic Foraminifera, Benthos’90, Sendai, 1990, Tokai university Press, p 3–9
Loeblich AR, Tappan H (1964) Sarcodina, chiefly “Thecamoebians” and Foraminiferida. In: Moore RC (ed) Treatise of Invertebrate Paleontology, Part C, Protista 2. The Geological Society of America and the University of Kansas Press, 2 volumes, p C1–C900
Loeblich AR, Tappan H (1987) Foraminiferal genera and their classification. Van Nostrand Reinhold Company Publisher, 2 volumes: 1 vol. text: X + 970 p., 1 vol. plates: VIII + 212 p. + 847 pl
Lopez-Ramos E (1969) Marine Paleozoic rocks of Mexico. Am Assoc Petr Geol B 53(12):2399–2417
Lothringer CJ (1993) Allochthonous Ordovician strata of Rancho San Marcos, Baja California Norte, Mexico, In: Gastil, RG, Miller, RH (eds) The Prebatholithic Stratigraphy of Peninsular California. Geol Soc Amer, Sp Paper 279:11–22
Lucas SG (2001) Taphotaxon. Lethaia 34:30
Lucia FJ (2012) The great Lower Ordovician cavern system, In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:81–109
Malakhova NP (1969) Fauna of the metalliferous series of the pyrit outcrops of the Southern Urals. Akad nauk SSSR, Ural Filial, Trudy Inst Geol Geokhim 81:1–84 (in Russian)
Mamet B (1998) A Late Devonian microfossil with dasyclad algae affinities from northwestern Australia. Alcheringa 22:21–28
Mamet B (2006) Taxonomy of Viséan marine calcareous algae, Fernie, British Columbia (Canada). Riv Ital Paleont S 112(3):323–357
Mamet B, Boulvain F (1992) Microflore des monticules micritiques frasniens “F2j” de Belgique. Rev Micropal 35(4):283–302
Mamet B, Préat A (2005) Microfaciès d’une lentille biohermale à la limite Eifélien/Givétien (Wellin, bord sud du Synclinorium de Dinant). Geol Belg 8(5):85–111
Mamet B., Préat A (2007) Eifelian–Givetian stromatoporoid-coral reefs, Belgium. In: Vennin E, Aretz M, Boulvain F, Munnecke A (eds) Facies from Palaeozoic reefs and bioaccumulations. Publ Scient Muséum, Mém Muséum nat Hist nat 195:191–193
Mamet B, Roux A (1975) Algues dévoniennes et carbonifères de la Téthys occidentale, Troisième partie. Rev Micropal 18(3):134–187
Mamet B, Roux A (1977) Algues rouges dévoniennes et carbonifères de la Téthys occidentale. 4e partie. Rev Micropal 1(4):215–266
Mamet B, Roux A (1982) Sur le mode de croissance de Nuia, algue incertae sedis. Geobios 15(6):959–965
Mamet B, Shalaby H (1995) Algues benthiques ordoviciennes de la Plate-forme du Saint-Laurent. Rev Micropal 38(3):229–244
Mamet B, Roux A, Lapointe M, Gauthier L (1992) Algues ordoviciennes et siluriennes de l’île d’Anticosti (Québec, Canada). Rev Micropal 35(3):211–248
Maslov VP (1954) On the early Silurian of eastern Siberia. In: Voprosy Geologii Azii. Akad nauk SSSR 1:495–529 (in Russian)
Maslov VP (1956) Fossil calcareous algae from SSSR. Akad nauk SSSR, Trudy Instit Geol Nauk, 160:1–301 (French translation BRGM n 3517) (in Russian)
Maslov VP (1973) Atlas of rock-building organisms (calcareous and siliceous organisms). Izdat “Nauka”, p 1–111 (in Russian)
Matte P (2001) The Variscan collage and orogeny (480–290 Ma) and the tectonic definition of the Armorica microplate: a review. Terra Nova 13:122–128
Mikhailevich VI (1980) Systematics and evolution of foraminifera in the light of new data on their cytology and ultrastructure. Akad nauk SSSR Trudy Zool Inst 94:42–61 (in Russian)
Miklukho-Maklay AD (1963) Late Palaeozoic of Central Asia. Izdat Leningrad Univ, p 1–328 (in Russian)
Miller JF, Loch JD, Taylor JF (2012) Biostratigraphy of Cambrian and Lower Ordovician strata in the Llano uplift, central Texas. In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:187–202
Montijo-González A, Teran-Ortega LA (1988) Geología del área de Rebeico con enfasis en el Paleozoico. Tesis Universidad de Sonora, Departamento de Geología, Sonora, pp 1–95
Moore RC, Lalicker CL, Fischer AG (1952) Invertebrate fossils. McGraw-Hill, New York, pp 1–766 (v–xiii)
Morales-Gámez M, Keppie JD, Norman M (2008) Ordovician–Silurian rift-passive margin on the Mexican margin of the Rheic Ocean overlain by Carboniferous–Permian periarc rocks: evidence from the eastern Acatlán Complex, southern Mexico. Tectonophysics 461(1–4):291–310
Moreman WL (1930) Arenaceous Foraminifera from Ordovician and Silurian limestones of Oklahoma. J Paleontol 4:42–59
Moreman WL (1933) Arenaceous Foraminifera from the lower Paleozoic rocks of Oklahoma. J Paleontol 7:393–397
Murphy JB, Gutiérrez-Alonso G, Nance RD, Fernandez-Suarez J, Keppie JD, Quesada C, Strachan RA, Dostal J (2006) Origin of the Rheic Ocean: rifting along a Neoproterozoic suture? Geology 34(5):325–328
Nance RD, Miller BV, Keppie JD, Murphy JB, Dostal J (2007a) Vestige of the Rheic Ocean in North America: The Acatlán Complex of southern Mexico. In: Linnemann U, Nance RD, Kraft P, Zulauf, G (eds) The evolution of the Rheic Ocean in North America: From Avalonian–Cadomian active margin to Alleghanian–Variscan collision. Geol S Am S 423:437–452
Nance RD, Fernández-Suárez J, Keppie JD, Storey C, Jeffries TE (2007b) Provenance of the Granjeno Schist, Ciudad Victoria, México: Detrital zircon U-Pb age constraints and implications for the Paleozoic paleogeography of the Rheic Ocean. In: Linnemann U, Nance RD, Kraft P, Zulauf G (eds) The evolution of the Rheic Ocean in North America: From Avalonian–Cadomian active margin to Alleghanian–Variscan collision. Geol S Am S 423:453–464
Nance RD, Gutiérrez-Alonso G, Keppie JD, Linnemann U, Murphy JB, Quesada C, Strachan RA, Nigel H, Woodcock NH (2010) Evolution of the Rheic Ocean. Gondwana Res 17:194–222
Nestell MK (1999) Correlation of Permian Tethys Neoschwagerinida (fusulinaceans) from the United States. In: Proceeding international symposium “Upper Permian stratotypes of Volga region”. Moscow Geos, p 161–171
Nestell GP, Nestell MK (2006) Middle Permian (Late Guadalupian) foraminifers from Dark Canyon, Guadalupe Mountains, New Mexico. Micropaleontology 52(1):1–50
Nestell GP, Tolmacheva TYu (2004) Early Ordovician foraminifers from the Lava River Section, northwestern Russia. Micropaleontology 50(3):253–280
Nestell MK, Nestell GP, Wardlaw BR, Sweatt MJ (2006) Integrated biostratigraphy of foraminifers, radiolarians and conodonts in shallow and deep water Middle Permian (Capitanian) deposits of the “Rader silde”, Guadalupe Mountains, West Texas. Stratigraphy 3(3):161–194
Nestell GP, Mestre A, Heredia S (2009) First Ordovician Foraminifera from South America: a Darriwilian (Middle Ordovician) fauna from the San Juan Formation, Argentina. Micropaleontology 55(4):329–344
Nestell G, Heredia S, Mestre A, Beresi M, González M (2011) The oldest Ordovician foraminifers (Oepikodus evae conodont Zone, Floian) from South America [Les plus vieux foraminifères (Zone à conodontes Oepikodus evae, Floien) d’Amérique du Sud]. Geobios 44:601–608
Nicholson HA, Etheridge R Jr (1878) A monograph of the Silurian Fossils of the Girvan District of Ayrshire with special reference to those contained in the “Gray Collection”. Scotl Geol Surv Mem 1(1):1–135
Nitecki MH, Mutvei H, Nitecki DV (1999) A phylogenetic debate on a problematic fossil taxon. Springer Science + Business Media, Berlin, pp 1–239
Noll JH, Dutro JT Jr, Beus SS (1984) A new species of the Late Devonian (Famennian) brachiopod Dzieduszyckia from Sonora, Mexico. J Paleontol 58:1412–1421
Ochoa-Granillo JA, Sosa-León JP (1993) Geología y estratigrafía de la Sierra Agua Verde con enfasis en el Paleozoico. Tesis, Universidad de Sonora, p 1–59
Olempska E (1986) Endolithic microorganisms in Ordovician ostracod valves. Acta Palaeontol Pol 31(3–4):229–236
Ortega-Gutiérrez F (1981) Metamorphic belts of southern Mexico and their tectonic significance. Geofís Int 20–3:177–202
Ortega-Gutiérrez F, Ruiz J, Centeno-García E (1995) Oaxaquia, a Proterozoic microcontinent accreted to North America during the late Paleozoic. Geology 23(12):1127–1130
Ortega-Gutiérrez F, Elías-Herrera M, Reyes-Salas M, Macías-Romo C, Lopez R (1999) Late Ordovician–Early Silurian continental collision orogeny in southern Mexico and its bearing on Gondwana–Laurentia connections. Geology 27:719–722
Page W, Harris AG, Repetski JE (2012) The Cambrian–Ordovician rocks of Sonora, Mexico, and Southern Arizona, Southwestern margin of North America (Laurentia). In: Derby R, Fritz RD, Longacre SA, Morgan WA, Stembach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:897–908
Palafox JJ (2011) Du Craton au Rhéïque : le nord et le centre de l’Etat du Sonora (Mexique) au Paléozoïque supérieur (Dévonien–Pennsylvanien), lithostratigraphie, biostratigraphie, approche géochimique et implications géologiques. Thèse Université Lille 1:1–315
Palafox JJ, Vachard D, Clausen S, Buitrón B, Devaere L (2013) The late Devonian and Carboniferous sequences in Sonora (Mexico). N M Mus Nat Hist Sci 60:322–323
Pantoja-Alor J (1970) Rocas sedimentarias paleozoicas de la régión centro-septentrional de Oaxaca. Sociedad Geológica Mexicana-Libro-Guía de la Excursión México-Oaxaca, p 67–84
Pearce H (2012) A study of the bioherms of the Early Ordovician Garden City Formation and a literature review of Early Ordovician organic buildups. Utah State University, All Graduate Plan B and other Reports. Paper 1931–67. http://digitalcommons.usu.edu/gradreports/193
Peiffer-Rangin F (1979) Les zones isopiques du Paléozoïque supérieur du nord-ouest mexicain témoins du relais entre les Appalaches et la cordillère ouest-américaine. C r Acad Sci Paris, série D 288:1517–1519
Peiffer-Rangin F (1987) Biostratigraphic study of Paleozoic rocks of northeastern and central Sonora. Thèse, Université de Paris, p 1–109
Peiffer-Rangin F, Echavarri Perez A, Salas-Piza G, Rangin C (1980) Sur la présence d’Ordovicien supérieur à Graptolites dans le nord-ouest du Mexique. C r Acad Sci Paris série D 290:13–16
Perez-Ramos O (1996) Investigación bibliográfica de la tectónica y sedimentación del Paleozoico y Mesozoico en Sonora. Posgrado Ciencias Tierra, p 1–69
Perret MF, Vachard D (1977) Algues et pseudo-algues des calcaires serpoukhoviens d’Ardengost (Hautes-Pyrénées). Ann Paléontol (Invertébrés) 63(2):85–156
Perry CT, MacDonald IA (2002) Impacts of light penetration on the bathymetry of reef microboring communities: implications for the development of microendolithic trace assemblages. Palaeogeogr Palaeoclimatol 186:101–113
Petersen MS, Rigby JK, Hintze LF (1976) Historical geology of North America. WMC Brown Company Publishers, Dubuque, pp 1–193
Poncet J (1986) Les algues calcaires du Paléozoïque inférieur de la baie d’Hudson et de l’Archipel Arctique Canadien. Bull Cent Rech Explor Prod Elf Aquitaine 10(2):259–282
Poole FG, Madrid RJ (1988) Allochthonous Paleozoic eugeoclinal rocks of the Barita de Sonora mine area, central Sonora, Mexico. In: Rodríguez-Torres R (ed) El Paleozoico de la Región Central del Estado de Sonora. Excursiones de Campo, Instituto de Geología, Universidad Nacional Autónoma de Mexico, Hermosillo, pp 32–41
Poole FG, Stewart JH, Repetski JE, Harris AG, Ross RJ Jr, Ketner KB, Amaya-Martínez R, Morales-Ramirez JM (1995a) Ordovician carbonate-shelf rocks of Sonora, Mexico. In: Ordovician odyssey: short papers for the seventh international symposium on the Ordovician System, 1995 Pacific Section, SEPM, p 267–275
Poole FG, Stewart JH, Berry WBN, Harris AG, Repetski JE, Madrid RJ, Ketner KB, Carter C, Morales-Ramirez JM (1995b) Ordovician ocean-basin rocks of Sonora, Mexico. In: Ordovician Odyssey: short papers for the seventh international symposium on the Ordovician System, 1995 Pacific Section, SEPM, p 277–284
Popov LE, Holmer LE, Bassett MG, Ghobadi-Pour M, Percival IG (2013) Biogeography of Ordovician linguliiform and craniiform brachiopods. In: Harper DAT, Servais T (eds) Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London Memoir 38:117–126
Poyarkov BV (1969) Stratigraphy and foraminifera from the Devonian deposits of Tian-Shan. Akad nauk Kirgiz SSR, Instit Geol, Uprav Geol Kirgiz SSR, Izdat “Ilim”, p 1–186 (in Russian)
Poyarkov BV (1979) Development and diffusion of the Devonian foraminifera. Akad nauk SSSR, Dalnevost Nauch Tsentr, Dalnevost Geol Insitut, Izdat “Nauka”, p 1–172 (in Russian)
Pratt BR (1995) The origin, biota and evolution of deep-water mud-mounds. In: Monty CLV, Bosence DWJ, Bridges PH, Pratt BR (eds) Carbonate mud-mounds, their origin and evolution. International Association Sedimentologists 23:11–48
Préat A, Kasimi R (1995) Sédimentation de rampe mixte silico-carbonatée des couches de transition eiféliennes-givétiennes franco-belges. Première partie: microfaciès et modèle sédimentaire. Bull Cent Rech Explor Prod Elf Aquitaine 19(2):329–375
Pronina TV (1963) Silurian foraminifers and some associated microorganisms from Ufa Amphitheatre. Palaeontologicheskii Zhurnal 1963(4):3–13 (in Russian)
Racki G, Sobón-Podgorska J (1993) Givetian and Frasnian calcareous microbiotas of the Holy Cross Mountains. Acta Palaeontol Pol 37(2–4):255–289
Ramos VA (1988) Late Proterozoic–Early Palaeozoic of South America—a collisional history. Episodes 11:168–174
Rauzer-Chernousova D, Reitlinger E (1957) Development of Foraminifera during the Paleozoic and their stratigraphic importance. Akad nauk SSSR, Izvestya, ser geol, p 103–128 (in Russian French translation by Sigal S, Sigal J, 1818:1–31)
Reitlinger EA (1959) Atlas of microfossils and problematica in the old deposits of Siberia. Akad nauk SSSR Trudy Inst Geol 25:1–62 (in Russian)
Richardson GB (1904) Report of a geologic reconnaissance in trans-Pecos Texas, north of the Texas and Pacific Railway. Univ Tex Miner Surv Bull 9:1–119
Riding R (1975) Girvanella and other algae as depth indicators. Lethaia 8:173–179
Riding R, Fan JS (2001) Ordovician calcified algae and cyanobacteria, northern Tarim basin subsurface, China. Palaeontology 44(4):783–810
Riding R, Soja CM (1993) Silurian calcareous algae, cyanobacteria, and microproblematica from the Alexander Terrane, Alaska. J Paleontol 67:710–728
Robison RA, Pantoja-Alor J (1968) Tremadocian trilobites from the Nochixtlán region, Oaxaca, Mexico. J Paleontol 42(3):767–800
Rodríguez-Torres R (1970) Geología metamórfica del área de Acatlán, Estado de Puebla. Sociedad Geológica Mexicana, Libro-guía de la excursión México-Oaxaca, p 51–54
Ross CA (1991) Pennsylvanian Paleogeography of the Western United States. In: Cooper JD, Stevens CH (eds) Paleozoic Paleogeography of the Western United States. II. Pacific Section, Society of Economic Paleontologists and Mineralogists, Los Angeles 67:137–148
Ross RJ Jr, Valusek JE, James NP (1988) Nuia and its environmental significance. New Mexico Bureau of Mines & Mineral Resources 44:115–121
Roux A (1985) Introduction à l’étude des algues fossiles paléozoïques (de la bactérie à la tectonique des plaques). Bull Cent Rech Explor Prod Elf Aquitaine 9(2):465–699
Roux A (1991a) Chapter 17, Ordovician algae and global tectonics. In: Riding R (ed) Calcareous algae and stromatolites. Springer Verlag, Berlin, pp 335–348
Roux A (1991b) Chapter 18, Ordovician to Devonian marine calcareous algae. In: Riding R (ed) Calcareous algae and stromatolites. Springer Verlag, Berlin, pp 349–369
Sabirov AM, Gushchin BM (2006) New Early Ordovician calcareous foraminifers of the Middle Tien Shan. Paleontol J 40(1):11–19
Salas MJ, Vannier J, Williams M (2007) Early Ordovician ostracods from Argentina: their bearing on the origin of the binodicope and palaeocope clades. J Paleontol 81:1384–1395
Saltovskaya VD (1981) Silurian and Devonian foraminifers from the Zeravshano-Gissarskoi oblast. Voprosy Mikropal 24:105–115 (in Russian)
Samankassou E (2002) Cool-water carbonates in a paleoequatorial shallow-water environment: the paradox of the Auernig cyclic sediments (Upper Pennsylvanian, Carnic Alps, Austria–Italy) and its implications. Geology 30(7):655–658
Sánchez-Zavala JL, Ortega-Gutiérrez F, Keppie JD, Jenner GA, Belousova E, Maciás-Romo C (2004) Ordovician and Mesoproterozoic zircons from the Tecomate Formation and Esperanza granitoids, Acatlán Complex, southern Mexico: local provenance in the Acatlán and Oaxacan Complexes. Int Geol Rev 46:1005–1021
Schlagwinteit F, Hladil J, Nose M, Salerno C (2013) Palaeozoic record of Thaumatoporella Pia, 1927 (incertae sedis)? Geol Croat 66(3):155–182
Scotese CR, McKerrow WS (1990) Revised World maps and introduction. In: Mckerrow WS, Scotese, CR (eds) Palaeozoic palaeogeography and biogeography. Geol Soc Mem 12:1–21
Scott DB, Medioli F, Braund R (2003) Foraminifera from the Cambrian of Nova Scotia: the oldest multichambered foraminifera. Micropaleontology 49(2):109–126
Sedlock RL, Ortega Gutiérrez F, Speed RC (1993) Tectonostratigraphic terranes and tectonic evolution of Mexico. Geol Soc Am Bull 278:1–153
Sengör AMC, Dewey JF (1991) Terranology: vice or virtue? In: Dewey JF, Gass IG, Curry GB, Harris NBW, Sengör AMC (eds) Allochthonous Terranes. Cambridge University Press, Cambridge, pp 1–21
Sengör AMC, Natal’in BA, van der Voo R, Sunal G (2014) A new look at the Altaids: a superorogenic complex in Northern and Central Asia as a factory of continental crust. Part II, Palaeaomagnetic data, reconstructions, crustal growth and global sea-level. Austrian J Earth Sci 107:131–181
Sepkoski JJ, Bambach RK, Droser ML (1991) Secular changes in Phanerozoic event bedding and the biological overprint. In: Einsele G, Ricken W, Seilacher A (eds) Cycles and events in stratigraphy. Springer Verlag, Berlin, pp 298–312 (Chapter 2, Article 2.6)
Sheehan PM (1975) Upper Ordovician and Silurian brachiopods from the Solis Limestone, Chihuahua, Mexico. J Paleontol 49(1):200–211
Shuysky VP (1973a) On discoveries of the algal genus Nuia in the Early Devonian of the Urals. Sverdlovskii Ordena Trudonogo Krasnogo Znameni Gornyi Institut Im. V.V. Vakhrusheva, In: Fauna and biostratigraphy of Middle and Late Palaeozoic from the Urals 93:65–68 (in Russian)
Shuysky VP (1973b) Early Devonian reefal calcareous algae from the Urals. Akad nauk SSSR, Ural Nauch Tsentr, Instit Geol Geokhim, p 1–119 (in Russian)
Sinha HN, Trampisch C (2013) Calcareous algal association from the Lower Paleozoic Tethyan sedimentary sequence of the Shiala Formation, Indian Gondwana. J Geol Soc India 82:339–350
Skompski S (2010) Paleobiogeographical significance of the Late Silurian microproblematicum Tuxekanella Riding and Soja. J Paleontol 84(2):346–351
Soreghan GS, Soreghan MJ, Poulsen CJ, Young RA, Eble CF, Sweet DE, Davogustto OC (2008) Anomalous cold in the Pangean tropics. Geology 36(8):659–662
Sour F, Buitrón BE (1987) Los graptolitos del Tremadociano de Ixtaltepec, Oaxaca, consideraciones sobre el límite Cámbrico–Ordovícico en la región. Rev Soc Mex Paleontol 1(1):380–394
Spincer BR (1998) Oolitized fragments of filamentous calcimicrobes and the pseudofossil affinity of Nuia Maslov from the upper Cambrian rocks of central Texas. J Paleontol 72:577–584
Sternbach CA (2012) Petroleum resources of the great American carbonate bank In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:125–160
Stevens CH, Forrest G, Poole FG, Amaya-Martínez R (2014) Late Paleozoic fusulinids from Sonora, México: importance for interpretation of depositional settings, biogeography, and paleotectonics (Fusulínidos del Paleozoico tardío de Sonora, México: importancia para la interpretación de los ambientes de depósito, la biogeografía y la paleotectónica). Rev Mex Cienc Geol 31(1):14–27
Stewart GA, Lampe L (1947) Foraminifera from the Middle Devonian bone beds of Ohio. J Paleontol 21(6):529–536
Stewart GA, Priddy RR (1941) Arenaceous foraminifera from the Niagaran rocks of Ohio and Indiana. J Paleontol 15(4):366–375
Stewart JH, Poole FG, Ketner KB, Madrid RJ, Roldán-Quintana J, Amaya-Martínez R (1990) Tectonics and stratigraphy of the Paleozoic and Triassic southern margin of North America, Sonora, Mexico. In: Gehrels GE, Spencer JE (eds) Geologic excursions through the Sonoran Desert region, Arizona and Sonora, vol 7. Arizona Geologic Survey, Special Paper, Tucson, Arizona, pp 183–202
Stewart JH, Poole FG, Harris AG, Repetski JE, Wardlaw BR, Mamet BL, Morales-Ramírez JM (1999) Neoproterozoic(?) to Pennsylvanian inner shelf, miogeoclinal strata in Sierra Agua Verde, Sonora, Mexico. Rev Mex Cienc Geol 16(1):35–62
Suleimanov IS (1945) New species of Tournaisian smaller foraminifers from the petroliferous region of Ishimbayev. Dokl Akad Nauk SSSR 48(2):124–127 (in Russian)
Swett K (1964) Petrology and paragenesis of the Ordovician Manitou formation along the Front Range, Colorado. J Sediment Petr 34:615–624
Tavernier A, Golubic S (1993) Pliocene microboring assemblages in shells of the Purisima Formation (Capitola, California). In: Barratolo F, De Castro P, Parente M (eds) Studies on Fossil Benthic Algae. Boll Soc Paleont Ital. Special Volume 1:405–414
Taylor JF, Repetski JE, Loch JD, Leslie SA (2012) Biostratigraphy and chronostratigraphy of the Cambrian–Ordovician great American carbonate bank. In: Derby JR, Fritz RD, Longacre SA, Morgan WA, Sternbach CA (eds) The Great American Carbonate Bank: The geology and economic resources of the Cambrian–Ordovician Sauk megasequence of Laurentia. AAPG Memoir 98:15–35
Toomey DF (1967) Additional occurrences and extension of stratigraphic range of the problematical microorganism Nuia. J Paleontol 41(6):1457–1460
Toomey DF, Klement KW (1966) A problematical microorganism from the El Paso Group (Lower Ordovician) of west Texas. J Paleontol 40(6):1304–1311
Toomey DF, LeMone D (1977) Some Ordovician and Silurian algae from selected areas of southwestern United States. In: Flügel E (ed) Fossil Algae. Springer, Berlin p, pp 351–359
Toomey DF, Nitecki MH (1979) Organic buildups in the lower Ordovician (Canadian) of Texas and Oklahoma. Fieldiana Geol N Ser 2:1–18
Torsvik TH, Cocks LRM (2013a) New global palaeogeographical reconstructions of the Early Palaeozoic and their generation. In: Harper DAT, Servais T (ed) Early Palaeozoic Biogeography and Palaeogeography. GeolSoc Mem 38:5–24
Torsvik TH, Cocks LRM (2013b) Gondwana from top to base in space and time. Gondwana Res 24:999–1030
Vachard D (1993) Fusulínidos del Guadalupiano superior (Pérmico) de Olinalá, Guerrero, Mexico. Rev Inst Mex Petrol 25(2):5–19
Vachard D (1994) Foraminifères et moravamminides du Givétien et du Frasnien du domaine Ligérien (Massif Armoricain, France). Palaeontogr Abt A 231(1–3):1–92
Vachard D (2016) Macroevolution and biostratigraphy of the Paleozoic foraminifers. In: Montenari M (ed) Stratigraphy and timescales 1. Elsevier, Amsterdam, pp 257–323. doi:10.1016/bs.sats.2016.10.005
Vachard D, Téllez-Girón C (1986) El alga Nuia en el Ordovícico de México; hipotesis diversas. Rev Inst Mex Petrol 18(2):12–25
Vachard D, Oviedo A, Flores de Dios A, Malpica R, Brunner P, Guerrero M, Buitrón BE (1993) Barranca d’Olinalá (Guerrero): une coupe de référence pour le Permien du Mexique central; étude préliminaire. Ann Soc Géol Nord 2(2):155–162
Vachard D, Flores de Dios A, Buitrón B (1997) Sur une nouvelle localité à fusulines du Wordien (Permien supérieur) du Mexique : conséquences paléogéographiques. Geobios 30(3):361–370
Vachard D, Flores de Dios A, Buitrón B, Grajales M (2000a) Biostratigraphie et paléogéographie des calcaires carbonifères et permiens de San Salvador Patlanoaya (Puebla, Mexique). Geobios 33(1):5–33
Vachard D, Flores de Dios A, Pantoja J, Buitrón B, Arellano J, Grajales M (2000b) Les fusulines du Mexique, une revue biostratigraphique et paléogéographique. Geobios 33(6):655–679
Vachard D, Vidaurre-Lemus M, Fourcade E, Requena J (2000c) New early Permian fusulinid assemblage from Guatemala. C R Acad Sci 331:789–798
Vachard D, Flores de Dios A, Buitrón B (2004) Guadalupian and Lopingian (Middle and Late Permian) deposits from Mexico and Guatemala, a review with new data. Geobios 37(1):99–115
Vachard D, Pille L, Gaillot J (2010) Palaeozoic Foraminifera: systematics, palaeoecology and responses to the global changes (Les foraminifères paléozoïques: systématique, paléoécologie et réponses aux changements globaux). Rev Micropal 53:209–254
Vachard D, Haig DW, Mory AJ (2014) Lower Carboniferous (middle Visean) foraminifers and algae from an interior sea, Southern Carnarvon Basin, Australia. Geobios 47:57–74
Vaughan APM, Leat PM, Pankhurst RLJ (2005) Terrane processes at the margins of Gondwana. Geol Soc Sp 246:1–21
Vdovenko MV, Rauzer-Chernousova DM, Reitlinger EA, Sabirov AA (1993) Guide for the systematics of Palaeozoic smaller foraminifers. Ross Akad nauk, Komiss Mikropaleont, “Nauka”, p 1–128 (in Russian)
Vega R, Araux E (1987) Estratigrafía del Paleozoico en el área del Rancho Las Norias, Sonora Central. Bol Dpto Geol Uni-Son 4(1–2):41–50
Versteegh GJM, Servais T, Streng M, Munnecke A, Vachard D (2009) A discussion and proposal concerning the use of the term calcispheres. Palaeontology 52(2):343–348
Voronova LG, Radionova EP (1976) Palaeozoic algae and microphytolites. Izdat Nauka 294:1–220 (in Russian)
von Raumer JF, Stampfli GM, Bussy F (2003) Gondwana-derived microcontinents—the constituents of the Variscan and Alpine collisional orogens. Tectonophysics 365:7–22
Williams M, Siveter DJ (2008) The earliest leperditicope arthropod: a new genus from the Ordovician of Spitsbergen. J Micropaleontol 27:97–101
Wilson JL (1975) Carbonate facies in geologic history. Springer, Berlin, pp 1–471
Wood A (1963) The British Carboniferous species of Girvanella (calcareous algae). Palaeontology 6(2):264–273
Young LM (1970) Early Ordovician sedimentary history of Marathon Geosyncline, Trans-Pecos, Texas. AAPG Bull 54(12):2303–2316
Acknowledgements
The study was supported by the projects PAPIIT 03-04-48367 of Conacyt and Ecos M13U01 and the grants of the projects NSh-974.2003.5 and MK-723.2004.4. We appreciated the help of Elisabeth Locatelli, Spencer Lucas, Karl Krainer, George Dix and John Repetski. Thanks to the reviewers of this version, Oliver Lehnert and Bruno Granier, for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Vachard, D., Clausen, S., Palafox, J.J. et al. Lower Ordovician microfacies and microfossils from Cerro San Pedro (San Pedro de la Cueva, Sonora, Mexico), as a westernmost outcrop of the newly defined Nuia Province. Facies 63, 18 (2017). https://doi.org/10.1007/s10347-017-0497-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-017-0497-9