
Abstract
A great number of farmland wildlife species has shown a sharp population decline in European countries, mainly caused by changes in agricultural practices leading to habitat alterations. Within this scenario, the identification of agricultural practices providing economic benefits to farmers and, at the same time, favoring higher biodiversity levels is a key challenge. This is especially interesting in Mediterranean livestock farming environments where bird numbers decrease due to intensification. Aiming at assessing the benefit to biological diversity of certain types of management practices in grasslands, we experimentally planted enriched pastures. Through a monitoring program of the bird and arthropod communities performed in Extremadura (W Spain) during 2 years (2016–2018), we compared relative richness of birds, abundance, and density values of birds and arthropods between treatment (three plots with enriched pastures) and control areas (six plots of natural grasslands). We found that enriched pastures had higher levels of relative abundance and density of wildlife with respect to natural grasslands and provided a greater variability and availability of trophic resources to birds. Habitat use by birds and arthropods not only depended on the type of pastures but also on the season, the study area, the presence of livestock and the vegetation height. Sowing enriched pastures constitutes a proactive measure stimulating biodiversity in Mediterranean extensive livestock farms that is also beneficial at the socioeconomic level. Therefore, co-financing this measure within the framework of subsidies from the Common Agricultural Policy of the European Union, as well as its inclusion in the future operational programs approved by the authorities, are essential.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Biodiversity protection entails the need of applying the best techniques of habitat, wildlife, and plant management, within an adaptive framework integrated in the conservation biology discipline (McCarthy and Possingham 2007; Allan and Stankey 2009; Wilmer et al. 2018). In the case of agricultural environments and others assimilated to natural pseudo-steppe lands, the decline in biodiversity indexes during the last 20 years are of great concern in Europe (Inger et al. 2015; Emmerson et al. 2016; Traba and Morales 2019).
A large number of agrosystem-dwelling wildlife species have suffered population declines due to shifts in habitat quality of the landscapes they inhabit. Specifically, changes in human management practices have led to the abandonment of agricultural lands with the subsequent development of forests and shrubby vegetation (Sokos et al. 2013; Zakkak et al. 2015) and, on the other hand, the intensification of agricultural and livestock uses has reduced the availability of feeding, refuge, and breeding areas for wildlife (Benton et al. 2003; Tscharntke et al. 2005; Emmerson et al. 2016). In the case of the European Union, one of the factor triggering changes in agricultural uses comes from the incentives of the Common Agricultural Policy (CAP) with which farming activities and products are subsidized based on regulated principles and priorities (European Parliament and Council of the European Union 2013; Barnes et al. 2016). Although biodiversity conservation has been a priority during the most recent cycles of the CAP planning, almost none of the applied indexes and proxies have shown a positive relationship between CAP subsidies and the improvement of the conservation status of species inhabiting these farmlands (Pe’er et al. 2014; Alons 2017; Navarro and López-Bao 2018). Population declines in vertebrates (mainly birds) and invertebrates arise as a result of the mechanization of labors, the earlier harvesting of crops, the generalized use of agrochemicals, or the lack of areas (i.e., copses and fallows) free of agricultural use around crop plots or pastures (Dudley et al. 2017; Palacín and Alonso 2018; Traba and Morales 2019).
Birds and invertebrates are taxa strongly affected by changes in agricultural uses in Europe (Benton et al. 2002; Attwood et al. 2008; Navarro and López-Bao 2018). Certain species are even threatened as is the case of the little bustard Tetrax tetrax, the Dupont’s lark Chersophilus duponti, the lesser kestrel Falco naumanni, or the black-bellied sandgrouse Pterocles orientalis, so there are specific conservation programs ongoing (Iñigo and Barov 2010a, 2010b; BirdLife International 2015). In the same way, emblematic species such as the great bustard Otis tarda are subject to coordinated international initiatives for their protection (Raab et al. 2009). These steppe-land birds are highly sensitive selecting specific areas with enough availability of food, refuge, and quietness according to their ecological requirements (Traba et al. 2015; Robleño et al. 2017). Nevertheless, it is important to take into account that nowadays, they depend on the sustainable human activities practiced at farmlands that benefit their ecological requirements.
With the purpose of favoring biodiversity at agricultural environments, it is necessary to apply management measures reconciling the presence of wildlife with socioeconomic uses, in a win-win framework. Farming makes up the livelihoods of many people and must provide sufficient incomes to maintain the socioeconomic structure in rural areas. Therefore, maintaining agricultural and livestock activities should be sustainable over time, including the possibility of subsidizing these actions in compliance with environmentally sustainable criteria (European Parliament and Council of the European Union 2013). In this scenario, it is very useful to know what agricultural practices can provide economic benefits to the farmers and, at the same time, enhance wildlife and plant diversity. This is especially interesting in Mediterranean grazing environments, where the decline of the bird communities has been remarkable due to management intensification (Palacín and Alonso 2018) and where the need for nutrient-rich natural pastures for livestock prevails (Bernués et al. 2011).
We experimentally sow enriched pastures in areas of extensive livestock use in Mediterranean environments of the Iberian Peninsula with the purpose of assessing the differences it could generate on several biodiversity indexes in relation to surrounding natural grasslands. These planted pasturelands met two main expectations: they are beneficial in relation to agronomic interests and there are references in previous studies of positive effects on biodiversity (Potts et al. 2009; Teixeira et al. 2014; Walden and Lindborg 2016; Hernández-Esteban et al. 2019). The objectives were: (1) to know the relative abundance of wildlife species (invertebrates and birds) in pasture plots of different types, comparing natural grasslands with enriched pastures. (2) To determine the spatial and temporal patterns of use by farmland birds, recognizing which groups or species respond more positively to the improved pastures. (3) To assess the type of management—sowing time, intensity and timing of grazing, perimeter protection, mowing, etc.—needed in the enriched pastures for increasing biodiversity. In particular, our hypotheses were the following: (1) planting enriched pastures provides a greater variability and availability of trophic resources, as well as a taller vegetation height (Potts et al. 2009; Walden and Lindborg 2016; Hernández-Esteban et al. 2019). (2) There are higher values of species richness, relative abundance, and density of farmland birds and arthropods in the improved pastures with respect to the surrounding natural pastures which would entail a positive measure to integrate into official programs of environmental subsidies (Navarro and López-Bao 2018). (3) There is variation in the habitat use depending not only on the type of pasture but also on the season, the location, the presence of livestock grazing, and the vegetation height.
Methods
Studied area and species
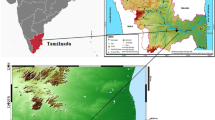
We carried out the study in Extremadura (W Spain, Fig. 1) on pseudo-steppe ecosystems dominated by grasslands. The potential vegetation of the study area corresponds to the Luso-extremadurense Mesomediterranean series of holm oak Quercus rotundifolia (Pyrobourgaeanae-querceto rotundifoliae sigmentum; Rivas-Martínez 1987). According to the Habitats Directive (Annex I; Council of the European Union 1992), the existing plant communities are included as “Semi-natural dry grasslands and scrubland facies,” the most relevant belonging to 34.5: “Pseudo-steppe with grasses and annuals (Thero-Brachypodietea)” which is listed as a priority habitat. The secondary stages of these grasslands consist of Spanish lavenders Lavandula stoechas and yellow Mediterranean brooms Retama sphaerocarpa.

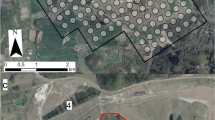
Study area in western Spain (Extremadura region). The three working zones are marked with red circles and the network of Special Protection Areas for Birds (SPA, Directive 2009/147/CE) is shown as pink shading. Aerial photographs of the treatment plots where enriched pastures were planted (blue contour) and the control plots (red contour) with the performed transect for bird and arthropod monitoring (black lines) in each of the three-labelled working zones are also shown. The black spots indicate the location of the observation points of great bustards Otis tarda in each zone
In study area, the woody vegetation is limited to the surroundings of streams and boundaries between livestock farms, having been generally removed during the nineteenth and twentieth centuries for their conversion into pasturelands (Vicente and Alés 2006). In particular, the area is managed with the purpose of providing food for livestock, traditionally ovine Ovies aries but more recently and increasingly bovine Bos taurus.
The natural dynamics of the plant communities are dependent on the climate of Mediterranean environments (Allué 1990). Rains usually take place from October to May, being minimal between June and September. The average annual rainfall is 489 mm. The mean monthly temperatures vary between 7.7 °C in January and 26.0 °C in July (data from the National Meteorological Institute. Cáceres Observatory).
Within the aforementioned area, we selected three working zones for assessing the effect of different types of grasslands (treatment and control, see below) on population parameters of birds and arthropods, which met the following requirements. (1) The zones were in Special Protected Areas for Birds (SPA) of the Natura 2000 network characterized by the presence of priority bird species linked to farmlands (MITECO-Junta de Extremadura 2019). (2) The zones belonged to private farms willing to collaborate that accepted the management conditions of our field test—see below—and had a minimum area of 200 ha. (3) There was an extensive or semi-extensive livestock exploitation regime, that is, sheep and/or cows feeding on the grasslands throughout the year—although alternating between different plots of the farm—without developing transhumance (Ruiz and Ruiz 1986) and with food supplementation during the periods of greater shortage of natural pastures.
The bird species targeted in our study (see list of species detected in Supplementary Material) are typical of Mediterranean pseudo-steppe environments and are present in the areas where the field work was developed. They show a decreasing population trend in Extremadura and globally in both Spain and the rest of Europe (EEA 2019). The whole bird community was subsequently grouped into various taxonomic and/or behavioral categories as described in Table 1 to inform their particular traits in habitat selection. Special effort was devoted to the great bustard due to its representative and emblematic role in agrosystems and its gregarious behavior (Magaña et al. 2011; Casas et al. 2019), so specific efforts were focused to monitor them with different methodological approaches. In relation to arthropods, we did not propose their study at the specific level, considering only their abundance and relative biomass as a whole. The study began in October 2015 with the preparation and planting of enriched pastures (see below). Subsequently, we performed field sampling during 2 years from March 2016 to February 2018.
Characterization of the study plots: implemented management measure
In each of the three working areas, three 30 ha plots were selected as experimental units (in total nine experimental units monitored). These three plots per area were geographically marked, geo-referenced, and contiguously located (Fig. 1). One of the three plots was randomly selected as treatment (where the enriched pasture was sown with the help of a tractor and the corresponding gears to carry out the work, see below) and the other two acted as control, maintaining their previous vegetation and regular dynamics of livestock use. The shape of each plot was approximately rectangular trying to ease the establishment of sampling transects (see below).
It would have been more appropriate to have an equal and higher number of treatment and control plots for increasing the robustness and representativeness of our results, but it was not possible to expand a greater number of treatment sites due to lack of funding for its implementation. Therefore, we preferred to increase the number of control plots to improve the accuracy of the results in these areas which did not bias the comparison with the treatment areas, but rather implied a better fit of data for the control plots.
The so-called enriched pasture was characterized by dryland herbaceous species adapted to the geological and weather features of the area. The enriched pasture does not depend on additional irrigation to the natural rainfall, has high nutritional and palatable value for livestock, is favored in terms of conservation and regeneration by a sustainable and non-intensive level of grazing, and has an approximate length of about 10 years (Loi et al. 2005; Lüscher et al. 2014). The species composition of these pasturelands was made up of seeds (82 kg/ha planted) of the following groups: four varieties of underground clovers—Trifolium subterraneum (12.2%), Persian clover Trifolium resupinatum (2.4%), Balansa clover Trifolium michelianum (1.2%), lucerne Medicago sativa (2.4%)—yellow lupin Lupinus luteus (18.3%), common vetch Vicia sativa (6.1%), triticale (wheat × rye) Triticum aestivum × Secale cereale (30.5%), raygrass Lolium sp. (8.5%), inoculizers, and seed pelletization (18.3%). Sowing the enriched pasture consisted of a deep labor (25 to 35 cm), turning soil horizons with plows with the objective of eliminating other vegetal remains to reduce competition with the planted vegetation. Then, we made a shallow labor (15 to 20 cm) with a harrow of discs for the preparation of the sowing substrate leaving the ground as smooth and fluffy as possible. It was supplemented with phosphorus (NPK 5-10-10) and carbonates Ca3 (PO4)2 fertilization. The sowing was carried out in lines with a front cereal planter, or with a precision planter, and a roller curl to bury the seeds superficially (1–2 cm). The total cost of establishing this enriched pasture, including seeds, fertilizers, and personnel, was 597 €/ha.
The control plots were natural grasslands, with typical species of the habitat type “Pseudo-steppe with grasses and annuals (Thero-Brachypodietea)”: Agrostis castellana, Brachypodium distachyon, Festuca elegans, Poa bulbosa, and Rumex bucephalophorus Trifolium subterraneum (Ríos and Salvador 2009). The dynamics of these pastures depends on the rainfall as well as on the livestock numbers, so that the vegetative period generally occurs during winter and spring. Each of these plots also accounted for 30 ha approximately.
Cattle and sheep were the only livestock present in the study areas (cattle in two of the three sampling areas and sheep only in the area called Mingalozana). They grazed the monitored plots throughout the year in an extensive way (without solid food supplementation except in summer) and with densities per unit area between 0.2 and 0.5 UGM/ha. UGM—bigger livestock unit—is equivalent to an animal with energy needs of 3000 forage units. A cow of about 550 kg corresponds to one UGM and an adult sheep to 0.15 UGJM approximately (MARM 2010). In the case of enriched pastures, with the purpose of their proper implantation, their consumption by livestock was avoided until the first natural seed production in order to ensure its natural reseeding. Therefore, these plots of pastures lacked grazing for at least 9 months after their sowing.
Fieldwork
After the selection of the experimental units (plots) and sowing of the enriched pasture, we started a monitoring program especially focused on birds and invertebrates, consisting of three sampling methods.
First, with the purpose of assessing the habitat use by the great bustard, we selected one elevated observation point in each of the three working zones. These were away from the study plots to avoid disturbances to the great bustards (min. 1 km of distance) and with good visibility of the entire zone. From these points, on a weekly basis, we visually counted the individuals present distinguishing between those within the treatment plot and in the rest of the observable territory that performed as control plots. The census lasted for 1 h during the maximum foraging activity period, avoiding the central hours. We recorded in each count: time, maximum simultaneous number of great bustards in each plot, maximum total number of different individuals observed in each plot, activity carried out (feeding, resting, courtship), and number of livestock heads present in each plot.
Second, within the three plots—one treatment and two controls—of each of the three working zones, linear census transects of 1 km length were established. Two field technicians previously standardized for the identification of targeted wildlife and in the methods to collect data were the only that sampled the transects on foot with the purpose of recording all birds detected (heard or observed) in two lateral bands: up to 50 m and more than 50 m from the observer up to the boundaries of the study plot. The birds were assigned to their species and to their presence within or outside these lateral bands aiming at subsequently calculate relative abundances (number of birds observed in the plot divided by the length in km of the transect) and relative densities (number of birds observed in the plot within the area in ha delimited by the two 50 m lateral bands on each side of the observer for the entire length of the transect). Census was performed once a month, during the maximum activity time of the birds (from sunrise to 3 h later).
Thirdly, during the return along transects described above, arthropods were sampled to check if a greater abundance would be related to the grassland type (enriched pastures or natural grasslands). For this, we carried out standardized captures through two methods: the first one with butterfly net (Joern 2005; Pocco et al. 2010) so that every 200 m of advance, we stopped and collected during 30 s the arthropods perched on the vegetation around that point. Thus, in each itinerary, there were five different collection events of arthropods with butterfly net. After each collection event, the net was covered to avoid captured arthropods from escaping so individuals were finally counted and weighed. The second method consisted of collecting the arthropods located on the ground and vegetation included within a 40 × 40 cm open quadrat (Gardiner and Hill 2006) that was randomly thrown every 200 m of transect. We put the collected arthropods in a pot for later counting and weighing without drying them. Both methods alternated sequentially every 100 m along the 1000 m transect. The targeted arthropods were > 1 mm in length according to the mesh size of the butterfly net used and the visual capability for its collection through the open quadrat.
Coinciding with arthropod sampling, we measured the height of the vegetation with the help of a 50 cm long ruler at a minimum of four points randomly distributed along the transect, subsequently recording the resulting average in centimeters.
Variables considered and statistics
Following the fieldwork, we considered different response and explanatory variables which were subsequently subject of statistical analyses. In total, we assessed eight response and six explanatory variables (Table 1).
For studying the habitat use by the great bustard, the number of observed individuals was assigned to treatment plot (30 ha) or, on the contrary, to the rest of observable area from the observation point. This latter varied according to the working zone (“1” = 930 ha; “2” = 685 ha; “3” = 738 ha; Fig. 1) and acted as control plots. In relation to the bird community (see Supplementary Material), the analysis of the relative abundance (birds/km) and relative density (birds/ha) showed a high correlation between each other (Spearman r = 0.85), so we chose the variable relative density in the subsequent analyses for simplification. For arthropods, the correlation values between the proposed response variables (biomass and relative abundance in the two sampling methods) were always less than 0.62, so both were included in further analyses.
The response variables (Table 1) were analyzed with respect to all explanatory variables, except in the case of the great bustard relative density that did not include the presence/absence of livestock signs in their analyses. Owing to the observations of this species were made at > 1 km, it was not possible to detect the presence of tracks in the studied plots, so the number of livestock heads (sheep and cattle) observed in each plot was used as alternative index. The relative abundance of arthropods was also considered explanatory only analyzing the bird abundance and richness patterns. For studies of the bird and arthropod communities, we neither included the number of livestock heads in the plots as explanatory variable because this was only proposed for the distant observations of great bustards (Table 1). We nested the explanatory variables zone and season in the variable type of area to know their influence in relation to the type of pasture management. On the other hand, since there was a high number of null observations during the sampling of great bustards, the variable great bustard relative density was log10-transformed to include it in the multivariate analyses.
First, aiming at determining a greater predictive power of the different potential subsets of explanatory variables in the results, we calculated the most parsimonious model using the Akaike’s information criteria (AIC, Burnham and Anderson 2002). We subsequently selected those combinations with the lowest AIC value and evaluated the estimates (β ± SD) for each level of the variables to know their relationship (+/−) with the response variable, as well as the Type 1 LR test (χ2) to know the significance value (p) of the covariates jointly included in the most parsimonious model.
Next, we performed GLMM for each of the response variables which showed a Poisson distribution and a log-function, in order to know what explanatory variables, as well as their interactions, influenced the results. The results (statistics, degrees of freedom, and value of statistical significance) of the analyses resulting in a confidence level greater than 95% (p < 0.05) are shown. All analyses were conducted with the Statistica 7.0 software (StatSoft, Tulsa, USA).
Results
We performed 300 different observation days of great bustards in the three working areas, with 27 different individuals counted in the treatment plots and 356 in the control areas. Regarding the counts of the bird community, in the 207 transects, we obtained a relative density of 5.83 birds/ha ± 8.41, for a total of 29,248 individuals corresponding to 43 different species. The mean species richness was 7.08 species/transect ± 2.19. The most abundant species registered were de calandra lark Melanocorypha calandra, the corn bunting Emberiza calandra, the Eurasian skylark Alauda arvensis, and the meadow pipit Anthus pratensis (Table 2). In the 207 samplings of arthropods with the butterfly net, we captured 2933 different individuals with a total body mass of 148.7 g, while with the open quadrat methodology, we counted 3881 specimens with a total body mass of 148.1 g.
Great bustard
The relative density of the great bustard varied according to environmental characteristics, being the type of area the variable with a greater explanatory power (Table 3). In the treatment plots, great bustards were observed more abundantly (mean 0.0059 birds/ha ± 0.0205) than in the control plots (mean 0.0030 birds/ha ± 0.0204). Additionally to the type of area (F1,56 = 75.08; p < 0.0001), the GLMM analyses showed a significant influence of the vegetation height (F1 = 4.11; p = 0.047) so that bustards were more frequent in areas with higher vegetation (Fig. 2).

a Mean ± CI (95%) of the relative density of great bustards Otis tarda expressed as the number of individuals/ha—log10-transformed—depending on the type of area studied (T = treatment, with enriched pastures, and C = control, with natural not improved grasslands). b Linear regression (black line) of the relationship between the relative density of great bustards expressed as number of individuals/ha—log10-transformed—and the height of natural vegetation (grasslands) in centimeters in both T and C zones
Bird community
The sampling of farmland birds showed that species richness followed a model in which the season was the more parsimonious variable (Table 3), resulting in higher values during spring and summer. GLMM analyses supported this pattern and only offered significant results for the interaction between type of area*season (F6,191 = 2.08; p = 0.050) and livestock signs presence (F1,191 = 4.32; p = 0.038).
Meanwhile, numbers of counted birds depended on the environmental traits. The model with a greater explanatory power (Table 3) ratified how the areas with enriched pastures and higher vegetation height hosted higher relative densities of birds, which also increased during the autumn (the type of area*season interaction offered significant values in the multivariate analyses: F6,191 = 3.03; p = 0.007; Fig. 3). In addition, a greater number of livestock signs was negatively related to the relative density of birds (F1,191 = 4.39; p = 0.037).

Mean ± CI (95%) of the relative density values of steppe and agrosystem-dwelling birds (number of individuals/ha) observed per sampling transect between treatment areas (with enriched pastures) and control areas (with natural not improved grasslands) between the different seasons of the year
By groups of species as described in Table 1, the granivorous passerines also responded positively to a combined effect of the treatment plots and a higher vegetation height, the season being also significant with greater abundances during autumn (type of area*season F6,191 = 2.29; p = 0.036; Table 3). Regarding the two bustard species jointly, none variable significantly determined a greater or lesser relative abundance. However, the model that best explained the relative abundance included the vegetation height and the type of area, as well as the respective interactions between these two variables and the season (Table 3). Finally, the higher abundances of sandgrouses better adjusted to the model composed of the greater arthropod abundance, the type of area, with increased numbers in areas with enriched pastures (F1,191 = 4.80; p = 0.029), a greater vegetation height (F1,191 = 5.58; p = 0.019), and the interactions among type of area, area, and season (Table 3).
Arthropods
The collected arthropods mainly corresponded to insects (Diptera, Orthoptera, Lepidoptera, and Coleoptera) in both the imago and larval stages, as well as arachnids. The differences in arthropod abundance were mostly based on the season, the zone, and the type of area (Table 3). In general, we collected more individuals during spring and summer, and in plots with enriched pastures. These general patterns were observed both for the relative abundance and for the total biomass of arthropods, through both sampling methods (butterfly nets and open quadrats), and were supported by the GLMM (Table 4; Fig. 4).

Mean ± CI (95%) of the relative abundance (number of individuals collected per sampling transect) of arthropods between treatment areas (with enriched pastures) and control areas (with natural not improved grasslands) between the different seasons, using the butterfly net collection method
Discussion
Effects on wildlife
Our results showed a positive effect of enriched pastures on the richness of birds and the relative abundance of birds and arthropods. Although it would have been preferable to expand the number and area of treatment and control sites (90 ha treatment vs. 180 ha control), the sample size under study (a mean of 23 transects per plot with a monthly basis), its geographic distribution among different areas, and the sampling duration (2 consecutive years), our study provides interesting conclusions of a favorable effect of the enriched pastures on the levels of biodiversity proxies considered. Nevertheless, more studies are needed to support these conclusions and to enhance the implications on the management of these pasturelands. In this sense, it would also have been desirable to assess the previous situation in the sampling plots to improve the accuracy of the conclusions with a before-after control-impact approach (BACI, Chevalier et al. 2019). Additionally to enriched pastures, other variables also influenced the bird and arthropod abundances, such as the relative abundance of invertebrate food resources, the variation of conditions at the local scale between the different working zones, the height of the vegetation, and the season, this latter due to the dynamics associated to the different life-cycle periods of the target species (Benton et al. 2003; Ivits et al. 2011).
The great bustard exhibited a greater preference for the treatment plots, mainly for feeding purposes, even though the small scale of the sampled areas (30 ha with respect to the rest of the observable territory from the observation points) and considering its large spatial requirements with respect to other steppe-land birds (Alonso et al. 1995; Suárez-Seoane et al. 2002; Concepción and Díaz 2011; Ponjoan et al. 2012). For other bird species, the higher densities recorded in the treatment areas modulated according to the season so the study areas hosted greater bird numbers during the autumn. It is worth noting that several threatened species such as the pin-tailed and black-bellied sandgrouses showed larger concentrations in these improved grasslands, following their habitat selection patterns towards areas of greater abundance of food resources after the breeding season (Martín et al. 2010), including the presence of more arthropods comparatively. The species that most clearly exhibited preference for enriched pastures were those smaller and granivorous. Larks such as the calandra lark, the Eurasian skylark, and the greater short-toed lark Calandrella brachydactyla, as well as the corn bunting, were more abundant in these improved grasslands, particularly during the months of October to December probably due to a greater availability of seeds and vegetable sprouts (Suárez-Seoane et al. 2002; Robleño et al. 2017).
The presence of enriched pastures and the season were the two variables that jointly explained a greater arthropod abundance and biomass. These pastures present a greater diversity of nutritional plants in relation to the surrounding environment due to the implantation of different types of vegetal species with high nutritional properties (especially with more protein content, see “Characterization of the study plots: implemented management measure” section). Spring was the season with a greater abundance, overlapping with the life-cycle phase of invertebrates in which they are more active and detectable (Curry 1993).
Our results are consistent with the previous studies that comparatively assessed the richness of birds, as well as the abundance of birds and arthropods between different types of grasslands, mainly in temperate climates. In general, higher biodiversity values are reached in lands with a greater plant heterogeneity and more resources available (Benton et al. 2003; Attwood et al. 2008; Sokos et al. 2013), which are offered by the planted enriched pastures. Likewise, higher abundances of farmland birds are recorded in areas where herbaceous vegetation shows a higher height and diversity.
Management of natural and enriched pastures
In relation to the management of the enriched pasture, and following the methodology applied in our study, we recommend the provision of additional resources to the sown plants through inorganic fertilizers during planting. On the other hand, it is necessary to avoid grazing by domestic and wild ungulates on the improved pasture during the first year after its sowing with the help of a perimeter fence. So the growth of the seeds, the proper development of the vegetation, and its subsequent natural seeding, which will provide natural regeneration of these annual herbaceous plants, would be guaranteed.
The geographic expansion of enriched pastures to a greater scale would increase the heterogeneity of landscapes dominated by Mediterranean natural grasslands often overexploited, providing patches of singular within a relatively monotonous environment. It also favors pollinating insects of great importance in promoting the productivity of these habitats (Potts et al. 2009). Anyway, the settlement of improved pasture plots should be posed in a complementary and non-generalized manner so that the natural and priority grassland habitats would not be depleted in their features nor extension, based on the need for their conservation. On the other hand, the difficulty of implanting key elements for landscape diversification on these grazing areas such as field borders, stonewalls, and strips of vegetation without grazing makes this alternative for biodiversity promotion even more important (Fahrig et al. 2011; Šálek et al. 2018).
Future challenges
In the face of the biodiversity crisis of European agrosystems, it is necessary to provide concrete and active measures favoring both the incomes of farmers and the abundance and richness of wild species. The latter, in addition, provide ecosystem services in these environments (Benayas and Bullock 2012; Garfinkel and Johnson 2015) so they must be taken into consideration for their intrinsic value. In this sense, there are plenty efforts made by different institutions encouraging good environmental practices for the management of agricultural lands, enabling their financing through European policies (i.e., Henle et al. 2008). In this sense, the CAP has a key role as an economic engine in rural areas, through the promotion of environmentally sustainable practices. There are not many specific measures in areas devoted to livestock allowing the amelioration of biodiversity levels through actions potentially subsidized by the CAP, by complying eco-conditionality (Pillar 1) or through voluntary measures (Pillar 2), in accordance with the financing lines of the most recently approved programs (Pe’er et al. 2014). For this reason, the implementation of enriched pastures would act as a co-fundable measure within the CAP scope, with the purpose of favoring biodiversity in Mediterranean livestock landscapes. Negotiations on a new operational program should consider this action as a priority either as a precondition for receiving other types of subsidies about productivity or maintenance of herds, or as a voluntary measure aimed at improving the overall quality of the grazed grasslands (Alons 2017; Navarro and López-Bao 2018).
References
Allan C, Stankey GH (2009) Adaptive environmental management, vol 351. Springer, New York
Allué JL (1990) Atlas fitoclimático de España. Taxonomías. Madrid: Ministerio de Agricultura, Pesca y Alimentación. Instituto Nacional de Investigaciones Agrarias
Alons G (2017) Environmental policy integration in the EU’s common agricultural policy: greening or greenwashing? J Eur Public Policy 24:1604–1622
Alonso JC, Alonso JA, Martín E, Morales MB (1995) Range and patterns of great bustard movements at Villafáfila, NW Spain. Ardeola 42:69–76
Attwood SJ, Maron M, House APN, Zammit C (2008) Do arthropod assemblages display globally consistent responses to intensified agricultural land use and management? Glob Ecol Biogeogr 17:585–599
Barnes A, Sutherland LA, Toma L, Matthews K, Thomson S (2016) The effect of the Common Agricultural Policy reforms on intentions towards food production: evidence from livestock farmers. Land Use Policy 50:548–558
Benayas JMR, Bullock JM (2012) Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 15:883–899
Benton TG, Bryant DM, Cole L, Crick HQ (2002) Linking agricultural practice to insect and bird populations: a historical study over three decades. J Appl Ecol 39:673–687
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Bernués A, Ruiz R, Olaizola A, Villalba D, Casasús I (2011) Sustainability of pasture-based livestock farming systems in the European Mediterranean context: synergies and trade-offs. Livest Sci 139:44–57
BirdLife International (2015) European red list of birds. Official publications of the European Communities, Luxembourg
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag, New York
Casas F, Mougeot F, Arroyo B, Morales MB, Hervas I, García De la Morena EL, Fagan WF, Viñuela J (2019) Opposing population trajectories in two Bustard species: a long-term study in a protected area in Central Spain. Bird Conserv Int 29:308–320
Chevalier M, Russell JC, Knape J (2019) New measures for evaluation of environmental perturbations using before-after-control-impact analyses. Ecol Appl 29:e01838
Concepción ED, Díaz M (2011) Field, landscape and regional effects of farmland management on specialist open-land birds: does body size matter? Agric Ecosyst Environ 142:303–310
Council of the European Union (1992) Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Official J Eur Comm 206:7–50
Curry JP (1993) Grassland invertebrates: ecology, influence on soil fertility and effects on plant growth. Chapman & Hall, London
Dudley N, Attwood SJ, Goulson D, Jarvis D, Bharucha ZP, Pretty J (2017) How should conservationists respond to pesticides as a driver of biodiversity loss in agroecosystems? Biol Conserv 209:449–453
EEA (European Environment Agency) (2019) The European Environment. State and Outlook 2020. Publications Office of the European Union, Luxembourg
Emmerson M, Morales MB, Oñate JJ, Batáry P, Berendse F, Liira J, Pärt T (2016) How agricultural intensification affects biodiversity and ecosystem services. Adv Ecol Res 55:43–97
European Parliament & Council of the European Union (2013) Regulation (EU) No 1306/2013 of the European Parliament and of the Council of 17 December 2013 on the financing, management and monitoring of the common agricultural policy and repealing Council Regulations (EEC) (…). Off J Eur Union 347:549–607
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Martin JL (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14:101–112
Gardiner T, Hill J (2006) A comparison of three sampling techniques used to estimate the population density and assemblage diversity of Orthoptera. J Orthop Res 15:45–51
Garfinkel M, Johnson M (2015) Pest-removal services provided by birds on small organic farms in northern California. Agric Ecosyst Environ 211:24–31
Henle K, Alard D, Clitherow J, Cobb P, Firbank L, Kull T, …Wascher D (2008). Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe. A review. Agric Ecosyst Environ 124:60-71.
Hernández-Esteban A, López-Díaz ML, CáceresY MG (2019) Are sown legume-rich pastures effective allies for the profitability and sustainability of Mediterranean dehesas? Agrofor Syst 93:2047–2065
Inger R, Gregory R, Duffy JP, Stott I, Voříšek P, Gaston KJ, Hill J (2015) Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecol Lett 18(1):28–36
Iñigo A, Barov B (2010a) Action plan for the lesser kestrel Falco naumanni in the European Union. SEO/BirdLife and BirdLife International for the European Commission, Brussels
Iñigo A, Barov B (2010b) Action plan for the little bustard Tetrax tetrax in the European Union. Brussels: SEO|BirdLife and BirdLife International for the European Commission
Ivits E, Buchanan G, Olsvig-Whittaker L, Cherlet M (2011) European farmland bird distribution explained by remotely sensed phenological indices. Environ Model Assess 16:385–399
Joern A (2005) Disturbance by fire frequency and bison grazing modulate grasshopper assemblages in tallgrass prairie. Ecology 86:861–873
Loi A, Howieson JG, Nutt BJ, Carr SJ (2005) A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems. Australian J Exp Agric 45:289–299
Lüscher A, Mueller-Harvey I, Soussana JF, Rees RM, Peyraud JL (2014) Potential of legume-based grassland–livestock systems in Europe: a review. Grass Forage Sci 69:206–228
Magaña M, Alonso JC, Palacín C (2011) Age-related dominance helps reduce male aggressiveness in great bustard leks. Anim Behav 82:203–211
MARM-Ministerio de Medio Ambiente y Medio Rural y Marino (2010) Real Decreto 1131/2010, de 10 de septiembre, por el que se establecen los criterios para (…) zonas remotas a efectos de eliminación de ciertos subproductos animales no destinados a consumo humano generados en las explotaciones ganaderas. Boletín Oficial del Estado 239:83996–83999
Martín CA, Casas F, Mougeot F, García JT, Viñuela J (2010) Seasonal variations in habitat preferences of the pin-tailed sandgrouse in agrarian pseudo-steppes. Ardeola 57:191–198
McCarthy MA, Possingham HP (2007) Active adaptive management for conservation. Conserv Biol 21:956–963
MITECO (Ministry for the Ecological Transition) & Junta de Extremadura (2019) Special Protected Areas for Birds. Extremadura. Summarized information. https://www.miteco.gob.es/es/biodiversidad/temas/espacios-protegidos/red-natura-2000/zepa_extremadura.aspx
Navarro A, López-Bao JV (2018) Towards a greener common agricultural policy. Nature Ecol Evol 2:1830–1833
Palacín C, Alonso JC (2018) Failure of EU Biodiversity Strategy in Mediterranean farmland protected areas. J Nat Conserv 42:62–66
Pe’er G, Dicks LV, Visconti P, Arlettaz R, Báldi A, Benton TG et al (2014) EU agricultural reform fails on biodiversity. Science 344:1090–1092
Pocco ME, Damborsky MP, Cigliano MM (2010) Comunidades de ortópteros (Insecta, Orthoptera) en pastizales del Chaco Oriental Húmedo, Argentina. Anim Biodivers Conserv 33:119–129
Ponjoan A, Bota G, Mañosa S (2012) Ranging behaviour of little bustard males, Tetrax tetrax, in the lekking grounds. Behav Process 91:35–40
Potts SG, Woodcock BA, Roberts SPM, Tscheulin T, Pilgrim ES, Brown VK, Tallowin JR (2009) Enhancing pollinator biodiversity in intensive grasslands. J Appl Ecol 46:369–379
Raab R, Julius E, Spakovszky P, Nagy S (2009) Guidelines for monitoring of population parameters of great bustard and of the effects of management measures. CMS Memorandum of Understanding on the conservation and management of the Middle-European population of the great bustard. Brussels, BirdLife International
Ríos S, Salvador F (2009) 6220 Pastizales xerofíticos mediterráneos de vivaces y anuales (*). In VV.AA. Bases ecológicas preliminares para la conservación de los tipos de hábitat de interés comunitario en España. Madrid: Ministerio de Medio Ambiente, y Medio Rural y Marino
Rivas-Martínez S (1987) Memoria del Mapa de Series de Vegetación de España. Hojas 17 y 22. ICONA, Madrid
Robleño I, Bota G, Giralt D, Recasens J (2017) Fallow management for steppe bird conservation: the impact of cultural practices on vegetation structure and food resources. Biodivers Conserv 26:133–150
Ruiz M, Ruiz JP (1986) Ecological history of transhumance in Spain. Biol Conserv 37:73–86
Šálek M, Hula V, Kipson M, Daňková R, Niedobová J, Gamero A (2018) Bringing diversity back to agriculture: smaller fields and non-crop elements enhance biodiversity in intensively managed arable farmlands. Ecol Indic 90:65–73
Sokos CK, Mamolos AP, Kalburtji KL, Birtsas PK (2013) Farming and wildlife in Mediterranean agroecosystems. J Nat Conserv 21:81–92
Suárez-Seoane S, Osborne PE, Alonso JC (2002) Large-scale habitat selection by agricultural steppe birds in Spain: identifying species–habitat responses using generalized additive models. J Appl Ecol 39:755–771
Teixeira RFM, Proença V, Valada T, Crespo D, Domingos T (2014) Sown biodiverse pastures as a win-win approach to reverse the degradation of Mediterranean ecosystems. In: Hopkins A et al (eds) EGF at 50: the future of European grasslands. Proceedings of the 25th General Meeting of the European Grassland Federation. IBERS, Aberystwyth University, Aberystwyth, pp 258–260
Traba J, Morales MB (2019) The decline of farmland birds in Spain is strongly associated to the loss of fallowland. Sci Report 9:1–6
Traba J, Morales MB, Carmona CP, Delgado MP (2015) Resource partitioning and niche segregation in a steppe bird assemblage. Comm Ecol 16:178–188
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol Lett 8:857–874
Vicente ÁM, Alés RF (2006) Long term persistence of dehesas. Evidences from history. Agrofor Syst 67:19–28
Walden E, Lindborg R (2016) Long term positive effect of grassland restoration on plant diversity-success or not? PLoS One 11:e0155836
Wilmer H, Derner JD, Fernández-Giménez ME, Briske DD, Augustine DJ, Porensky LM (2018) Collaborative adaptive rangeland management fosters management-science partnerships. Rangel Ecol Manag 71:646–657
Zakkak S, Radovic A, Nikolov SC, Shumka S, Kakalis L, Kati V (2015) Assessing the effect of agricultural land abandonment on bird communities in southern-eastern Europe. J Environ Manag 164:171–179
Acknowledgements
R. Higuero, M. Abascal, J. Caldera, and L. M. González helped in different phases of the study. The Junta de Extremadura provided essential help and authorized the implementation of the project. The managers of the farms in which the fieldwork was performed (Casatejada, Mingalozana y Martinagómez) collaborated fruitfully so we are obliged to all of them. This work was carried out within the framework of the project Developing management guidelines in areas of ecological interest on agricultural lands, commissioned by the Spanish Ministry for the Ecological Transition and developed by TRAGSATEC. Two anonymous reviewers improved the first versions of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 16 kb)
Rights and permissions
About this article

Cite this article
Moreno-Opo, R., Pina, M. & Mogena, A. Sowing enriched pastures for extensive livestock enhances the abundance of birds and arthropods in Mediterranean grasslands. Eur J Wildl Res 67, 40 (2021). https://doi.org/10.1007/s10344-021-01486-2
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-021-01486-2