
Abstract
This study examines the habitat selection of grey partridge (Perdix perdix) and the effect of habitat type within partridge home ranges on nest site choice and nest success. Data were collected via radio-tracking in three different areas in the Czech Republic between 2002 and 2010. Compositional analysis was performed on 12 habitat types and uncultivated habitats (such as ruderals, field margins and game refuges) were selected the most during the pre-nesting and nesting periods. Grey partridges tended to nest in uncultivated habitats and avoided nesting in dominant cereal monocultures. We tested six variables that may determine nest site choice and success. Of these, the study area, proportion of selected habitat and habitat diversity (expressed by the Simpson’s diversity index) significantly affected nest placement in uncultivated habitats. Despite this, the lowest nest failure rate was observed in the predominant cereal habitat. Our findings suggest a possible ecological trap for partridges throughout our study areas. Due to the lack of uncultivated habitats, partridges favoured nesting in habitats with a higher predation risk. Conservation managers should increase the proportion of uncultivated areas and promote habitat diversity for partridge populations in central Europe.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
There has been a strong decline in the abundance of many bird species within agricultural landscapes since the latter half of the twentieth century (Siriwardena et al. 1998; Chamberlain et al. 2000; Donald et al. 2001; Aebischer and Ewald 2010; Reif and Vermouzek 2019; Traba and Morales 2019). Agricultural land dominates the European landscape and is an important habitat for many species (Robinson and Sutherland 2002). Chemical usage, agricultural intensification and changes away from more traditional methods are most often cited as causes of the decline in population sizes of farmland species (Newton 2004; Frenzel et al. 2016). One of the most important impacts of agricultural intensification is the loss of habitat heterogeneity, which is key for biodiversity (Benton et al. 2003). This is mainly caused by merging of smaller fields into larger units, accompanied by the removal of field margins and other unmanaged patches within agricultural landscapes (Robinson and Sutherland 2002). However, unmanaged habitats including fallows, ruderals, grass and shrub patches, are essential for most farmland bird species (Traba and Morales 2019). The sowing of crop with minimal variability also contributes to the homogenisation of the landscape. With such practices comes a loss of suitable habitats for foraging, nesting and rearing of young (Benton et al. 2003). The loss of suitable habitat is known to result in increased predation pressure on populations of farmland species (Whittingham and Evans 2004). Working in synergy with habitat loss, this subsequently results in rapid declines of farmland species (Evans 2004). Therefore, the solution lies in habitat heterogeneity restoration through the support of small field units with different crop types and increased field edges with interspersed fallows, ruderals, grass and shrub patches (Sasaki et al. 2020). Knowledge of habitat selection and requirements of threatened farmland species forms the base of recommendations for changes in agricultural landscapes which could slow down or stop population declines.
The grey partridge is a native European species whose occurrence is associated to agricultural landscapes; however, the species has experienced one of the largest population declines out of all farmland birds (Kuijper et al. 2009). Previously, the species occurred in steppe habitats with preferences for continuous grass interspersed with taller and denser cover (e.g. hedgerows and scrubs) providing concealment and areas used during the breeding season (Cramp and Simmons 1982). Additionally, field margins and rough grassy strips are often used for foraging or nesting (Vickery et al. 2004). Novoa et al. (2002) stated that important habitats for partridges during the breeding season are those with shrub canopy cover higher than 40%. Pairs that occurred more often in these habitats in the spring had a higher probability of successfully rearing a brood. Furthermore, pre-nesting habitat use may reflect the nesting habitat quality (Novoa et al. 2002). The importance of dense vegetation is more pronounced in the nesting period. Vegetation cover provides concealment and thus limits the predator’s ability to detect the nest and incubating female (Albrecht and Klvaňa 2004). Partridges have higher nesting success in areas with more abundant permanent vegetation which is selected due to increased nesting cover (Panek 1997). In addition to vegetation density, substrate type is an important factor for partridge nest site selection. Partridge nests are often placed in habitats with a high amount of dead grass and leaf litter (Rands 1988). Partridges use this material to cover their nests during the laying period (Cramp and Simmons 1982) but not during incubation (Jenkins 1961; Černý et al. 2018). Egg covering is generally perceived as an effective antipredator strategy (Opermanis 2004; Kreisinger and Albrecht 2008; Prokop and Trnka 2011) which is important especially during the laying period when the nests are left unattended for longer durations (Lack 1947). During incubation, the eggs are masked by the cryptically coloured hen who incubates very firmly and leaves the nest for only very short time periods (Černý et al. 2018). During this crucial period, any excessive movement around the nest could reveal its position to predators (Skutch 1949; Martin et al. 2000). Considering that permanent vegetation cover with dead grass and dry vegetation increases nest success (Rands 1988; Panek 1997) and dense vegetation provides better concealment for the nest and the incubating female (Albrecht and Klvaňa 2004), it is predicted that nests placed in uncultivated habitats suffer from lower predation rates than in cultivated habitats.
Using telemetry data from three different populations of grey partridges, we describe the habitat composition in partridge pre-nesting and nesting home ranges and analyse habitat selections in the modern agricultural landscapes of central Europe. We hypothesised that (1) grey partridges will select unmanaged habitats rather than cultivated (dominant) areas and (2) nest success will be higher in unmanaged habitats than in other areas. From our results, we will make recommendations for conservation managers focused on halting the decline of this disappearing game species.
Methods
Study area
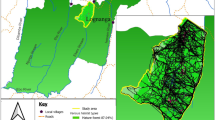
Data were collected from three study areas in the Czech Republic (Fig. 1) during the following years: 2002 (Prague), 2003–2004 (Písek) and 2009–2010 (Milešín). The first study area was located on the southwest edge of the capital city Prague (50° 02′ 25″ N, 14° 18′ 48″ E; 300–370 m a.s.l.; 5 km2). This suburban area consists mainly of fields, unmanaged habitats (ruderals, shrubs, field margins, etc.) and infrastructure with gardens and areas of freshwater (for details see Supplementary Material Table S1). The partridge population was the highest density there with more than 70 pairs/km2 in weedy patches and 5 pairs/km2 in the surrounding farmland (Šálek et al. 2004). The second study area lies in South Bohemia, south of the town Písek (49° 17′ 05″ N, 14° 08′ 43″ E; 360–460 m a.s.l.; 19 km2). This agricultural landscape consists of fields, unmanaged habitats, meadows, forest patches, infrastructure and areas of freshwater. Partridge population density ranged from 2 to 5 pairs/km2 in this area (for details see Šálek and Marhoul 2008). The third study area is located in the region of Vysočina between the villages of Milešín and Nová Ves (49° 22′ 55″ N, 16° 12′ 06″ E; 480–620 m a.s.l.; 17 km2). It is also characterized by agricultural habitats with a high proportion of arable fields. In addition, there are forest fragments, meadows, unmanaged areas and infrastructure, gardens and areas of freshwater. Partridge population density ranged from 2 to 4 pairs/km2 in the third study area (for details, see Rymešová et al. 2013).

Map of the Czech Republic with three study areas and their habitat compositions
Radio tracking
Partridges were caught with mist nets at dusk, with drop-traps or with a handheld net placed on a long pole at night with two spot lights to dazzle them (Benítez-Lopéz et al. 2011). In Milešín, coveys were caught in winter with the help of local gamekeepers using drop-traps at feeding sites and partridges were kept in aviaries until the end of winter according to traditional game management (for details see Rymešová et al. 2013). Some birds were also caught with a handheld net at night in Milešín (especially unmarked individuals from pair). All individuals (Prague: 2002 n = 44; Písek: 2003 n = 21 and 2004 n = 17; Milešín: 2009 n = 58 and 2010 n = 49) were equipped with a necklace radio transmitter (TW-4, Biotrack, UK, weight = 6.5 g). We used AR 8000 broadband receivers (AOR, Japan), IC-R10 (Icom, USA) and SIKA (Biotrack, UK) and Yagi antenna optimized for 151 MHz suitable for radio-tracking. Partridges were tracked between two to five times for a week, and their locations were obtained using triangulation of the transmitted signal and additional direct observations (whenever it was safe without disturbing the individual). Each location was marked whilst in the field on a satellite image of the study area. Nests were searched from late April to July. To ease this process, we tracked signals of females from pairs during the day. If the female was in different place than the male (more than 10 m) and stayed there for at least 15 min, we identified this place by triangulation as a possible nest site. After the female left this location, we carefully checked the site. Once the nest was found, we recorded its location, number of eggs and habitat. We checked each known nest over 2- to 3-day intervals to determine its fate—a successful nest (hatching of chicks) or nest failure (predation of the nest or the nesting female or nest abandonment).
Habitat categories
We defined twelve representative habitat categories in the study areas: cereal, oilseed rape, other crops, fallow fields, ruderals, meadows, field margins, shrubs, game refuges, forests, gardens and infrastructure. These categories were chosen on the basis of their management, proportion in the study area, patch size, vegetation characteristics (height, cover and density) and food supply for grey partridges (Table 1). The cereal category included wheat, oat, barley and rye. Generally, this habitat is medium/high in height (up to 1 m) with dense vegetation usually across large areas. There were no differences in height between cereal fields with spring and winter crops during the main nesting period of partridges. Oilseed rape was placed in a separate category as it creates a habitat with tall stems (> 1 m) which provide sufficient cover for grey partridges across large areas. The category ‘other crops’ included all other non-dominant heterogeneous habitats in the study area. These were different from the ‘cereal’ and ‘oilseed rape’ categories and included beet, maize, strawberries, potatoes, pea and poppy. Fallow fields are fields without any agricultural management in that calendar year. Ruderals included grassy habitats, sometimes with scattered small shrubs, without any human management. In contrast, meadows are grassy habitats that are mowed several times a year. Fields with clover were included in the ‘meadows’ category. The category ‘field margins’ contained grassy habitats of a linear character often between fields, along the edges and also along roads. We also defined wild-flower strips as ‘field margins’. ‘Shrubs’ included dense shrubs with grassy undergrowth. ‘Game refuges’ comprised unmanaged small grassy patches with shrubs and trees up to 1 ha, which are often scattered in fields or lie along their edges. ‘Forests’ are tree areas larger than 1 ha. ‘Gardens’ included orchards and small cultivated patches with vegetables, which are regularly managed by landowners and have a typical varied composition of crops and fruit trees. The ‘infrastructure’ category included all manmade habitats (mainly buildings) and freshwater areas. These habitats are not suitable for partridges and however there is a small possibility of birds occurring in them. Nests of grey partridge were found in eight of twenty two identified habitat types (Supplementary Material Table S2). The eight nesting habitats were merged into two categories ‘uncultivated’ (fallow fields, field margins, game refuges and ruderals) and ‘cultivated’ habitats (cereal, gardens, meadows and oilseed rape) for analysis of nest site choice.
Sample size
We collected data for 97 partridges (Prague: 2002 n = 22; Písek: 2003 n = 11 and 2004 n = 9; Milešín: 2009 n = 25 and 2010 n = 30) and their 66 nests (Prague: 2002 n = 12; Písek: 2003 n = 10 and 2004 n = 5; Milešín: 2009 n = 19 and 2010 n = 20). Only data from individuals with at least 10 locations (set as the lowest threshold) were used for home range estimation. Individuals with less than 10 fixes (n = 2) were excluded from the habitat selection analysis. We calculated home range sizes for both the pre-nesting and nesting periods from individual locations using 75% kernel density estimates (i.e. KDE, h reference method) in the software ArcView 3.3 (including Home Range Analysis extension). Pre-nesting home ranges were calculated from the subset of locations, corresponding to the period from the first day of partridges pair observations (8th February–10th May, median 18th March) to one day prior the date when the first egg was laid. Locations for the computation of nesting home ranges were chosen in the period from the date the first egg was laid (21st April–20th July, median 7th May) to the end of the nesting period (hatching, abandonment or predation of the known nest or female). We used the date May 1st for all nests that were found late; this was done when it was not possible to estimate the exact date when the first egg was laid.
Habitat evaluation
We used R 3.6.1 (R Core Team 2019) for all analyses. First, we created habitat layers in ArcView 3.3 on the basis of underlying satellite images for each area for each year. Actual crop composition was updated with collected data from field surveys. Subsequently, the habitat layers were clipped to the shape of each home range using the XTools extension and area sizes of different habitat categories were obtained (m2). We calculated Jaccard dissimilarity indices to test dissimilarity between study areas and years using the vegdist function from the ‘vegan’ package in R (Oksanen et al. 2019).
Compositional analysis
We used the compana function with a randomisation test and 1000 repetitions from the ‘adehabitatHS’ package (Calenge 2006) for habitat compositional analysis (Aebischer and Robertson 1992; Aebischer et al. 1993; Kenward 2001) for the pre-nesting and nesting home ranges separately. Compositional analysis was performed for each study area separately as habitat availability differed between years in the Písek and Milešín study areas. We used two methods of compositional analysis to identify selected habitats: (1) percentage of each habitat within the home range classed as ‘used sites’ and percentage of habitats for the whole study area as ‘available sites’ (HRxAR) or (2) percentage of locations in the different habitats within the home range as ‘used sites’ and percentage of habitat areas in the home range as ‘available sites’ (LxHR). We replaced all zero value habitat categories in an individual home range with the value 0.001 following recommendations of Aebischer et al. (1993). The habitat categories ‘other crops’ and ‘forests’ were excluded in the Písek study area for the pre-nesting home ranges (LxHR type) due to their absence in both the ‘used’ and ‘available’ sites.
Nest habitat selection
A χ2 test for association was used to test if the number of nest locations (a) in the most selected and other habitats (sum of other habitat types cases), (b) in the dominant and other habitats, and (c) in cultivated and uncultivated habitats differed from the expected frequencies in each of three study areas. The expected frequencies were calculated from areas of known nesting habitats in the whole study area. At first, an area percentage of each habitat was computed from the subset of habitat types used for nesting. The number of identified nests in the study area (N) was then compared with area percentages of known nesting habitats to obtain expected frequencies.
We used the glm function from the ‘stats’ package (R Core Team 2019) to test which predictor variables affect nest site choice. These were coded as nest locations in cultivated or uncultivated habitats. The following predictors were used: (1) proportion of selected habitat (HRxAR method) in the pre-nesting home ranges, (2) proportion of dominant habitat in the pre-nesting home ranges, (3) Simpson’s diversity indices for habitat types in each pre-nesting home range, (4) month of laying of the first egg (April, May, June or July), (5) nest order (the first clutch or second replacement clutch) and (6) study area (Prague, Písek and Milešín). Proportions of selected and dominant habitats in each home range during the pre-nesting period were used as partridges choose the nest site in this period, whereas in the nesting period, the first egg has already been laid. The proportion of selected habitats was logarithmically transformed (ln x + 1) to obtain a normal distribution.
Nesting success
We used the glmer function from the ‘lme4’ package for generalized mixed-effect modelling (Bates et al. 2015) to test our second hypothesis: which studied parameters influence nest success? These were subsequently coded as the successful hatching of at least one chick or alternative nest failure (abandonment, predation or destruction). The following predictors were used: (1) nest location in the selected habitat (resulted from HRxAR method in all following cases), (2) nest location in the dominant habitat, (3) nest location in cultivated habitats, (4) month when the first egg was laid, (5) nest order, (6) proportion of the selected habitat in each nesting home range, (7) proportion of the dominant habitat in the nesting home range and (8) Simpson’s diversity indices for habitats in nesting home ranges. We included study areas as random effects in the model. We calculated daily nest failure rate (dnfr) according to Mayfield (1975) for comparison of nesting success.
Results
The proportion of habitats differed between study areas, home ranges and locations (Fig. 2). Cereal was dominant in all study areas—Prague 38.9%, Písek 38.0% in 2003 and 41.3% in 2004, Milešín 54.1% in 2009 and 54.9% in 2010. Prague had the largest proportion of uncultivated habitats (field margins, ruderals and shrubs; 24.2%) being almost two times more frequent than in Písek (fallow fields, field margins, game refuges and ruderals; 13.6% in 2003 and 12.2% in 2004) and four times more frequent than in Milešín (field margins and game refuges; 6.0% for both years). These differences were confirmed by the Jaccard dissimilarity index values. The suburban study area in Prague is the most different from the other two areas (78.4% dissimilarity from Písek and 85% dissimilarity from Milešín). Conversely, the Jaccard dissimilarity index between Písek and Milešín was 36.5%. Dissimilarity between years in Písek was 7.2% and 9.7% in Milešín. Cereal was also the most commonly represented habitat in partridge home ranges (Fig. 3) and locations (Fig. 4) with an exception in Prague during the pre-nesting period when partridge locations mostly occurred in ruderals.

Mean proportions of habitats in each study area. Legend: whole study area (black bars), pre-nesting home ranges (dark grey full bars), pre-nesting locations (light grey full bars), nesting home ranges (dark grey hatched bars), nesting locations (light grey hatched bars); CE—cereal, FM—field margins, GA—gardens, IN—infrastructure, OR—oilseed rape, OC—other crops, FO—forests, GR—game refuges, ME—meadows, RU—ruderals, FF—fallow fields, SH—shrubs

Proportions of habitats in grey partridge home ranges during the pre-nesting (Prague: n = 21, Písek: n = 14, Milešín: n = 51) and nesting periods (Prague: n = 26, Písek: n = 23, Milešín: n = 59). Legend: CE—cereal, FM—field margins, GA—gardens, IN—infrastructure, OR—oilseed rape, OC—other crops, FO—forests, GR—game refuges, ME—meadows, RU—ruderals, FF—fallow fields, SH—shrubs. Median with 25% and 75% quantiles (boxes) and 1.5 times interquartile range (whiskers) are indicated

Proportions of grey partridge locations in habitats during the pre-nesting (Prague: n = 21, Písek: n = 14, Milešín: n = 51) and nesting periods (Prague: n = 26, Písek: n = 23, Milešín: n = 59). Legend: CE—cereal, FM—field margins, GA—gardens, IN—infrastructure, OR—oilseed rape, OC—other crops, FO—forests, GR—game refuges, ME—meadows, RU—ruderals, FF—fallow fields, SH—shrubs. Median with 25% and 75% quantiles (boxes) and 1.5 times interquartile range (whiskers) are indicated
The results of the compositional analysis showed that grey partridges did not use habitats randomly according to their proportions within study areas or home ranges (Table 2). They mostly selected unmanaged habitats such as field margins, ruderals and game refuges. However, there were exceptions for selections described by LxHR in Prague where the most selected were other crops during the pre-nesting period and gardens during the nesting period (Table 2). Partridges avoided habitats such as infrastructure and forests.
A total of 66 nests were located in eight habitat types—field margins (n = 21), ruderals (n = 15), game refuges (n = 4), fallow fields (n = 2), cereal (n = 8), meadows (n = 7), oilseed rape (n = 5) and gardens (n = 4). We analysed habitat proportions separately for each study area but combined data across years. This was done due to the low dissimilarity in habitat proportions between years for the same areas and no change in proportions of either the most selected nesting habitats or dominant habitat between years. Results of the χ2 test showed that the nests were located more often in selected habitats (field margins, ruderals or game refuges) than in all other habitats combined in all of the study areas. Nest location probability in selected habitats was more than two times higher in Prague (χ2 = 8.631, df = 1, p = 0.003), five times higher in Písek (χ2 = 13.280, df = 1, p < 0.001) and nine times higher in Milešín (χ2 = 124.030, df = 1, p < 0.001) than expected frequencies. However, partridges avoided the dominant habitat when choosing nest sites. Nests were placed in dominant habitats four times less often in Prague (χ2 = 13.813, df = 1, p < 0.001) and Písek (χ2 = 13.335, df = 1, p < 0.001), and more than six times less often in Milešín (χ2 = 58.293, df = 1, p < 0.001) than would be on the basis of expected frequencies. In 63.6% of cases, nests were located in uncultivated areas and 36.4% of cases in cultivated areas. Nests were preferably located in uncultivated habitats (fallow fields, field margins, game refuges and ruderals) in all three study areas (Prague: χ2 = 11.329, df = 1, p < 0.001; Písek: χ2 = 43.406, df = 1, p < 0.001; Milešín: χ2 = 101.360, df = 1, p < 0.001).
We found that the proportion of selected habitats in pre-nesting home ranges, Simpson’s diversity indices for habitats in each pre-nesting home range and the study area were significant predictors of nest site locations within cultivated and uncultivated habitats (Table 3). Nests were more often located in uncultivated habitats when the proportion of selected habitats in the pre-nesting home range was higher (Fig. 5). There was also higher habitat diversity in partridge home ranges (lower value of Simpson’s diversity index) indicating that nests will be more frequently located in uncultivated habitats (Fig. 6). Uncultivated habitats were most often used as nest sites in the Písek study area, whereas cultivated habitats were most often used as nest sites in Milešín (Fig. 7).

Logistic model of relationships between the estimated probability that nests are located in uncultivated habitats (habitat type = 1) and proportion of selected habitats in pre-nesting home ranges (75% KDE) with 95% confidence interval (grey colour)

Logistic model of relationships between the estimated probability that nests are located in uncultivated habitats (habitat type = 1) and Simpson’s diversity index values in pre-nesting home ranges (75% KDE) with 95% confidence interval (grey colour)

Comparison of the proportion of nests located in uncultivated habitats (grey) and cultivated habitats (white) in Prague (n = 12), Písek (n = 15) and Milešín (n = 39)
Nineteen out of 66 nests successfully hatched chicks (i.e. 29%; four nests in Prague, five in Písek and 10 in Milešín). Predation was the cause of nest failure in 37 cases (n = 17 for predation of clutch, n = 13 for predation of female and n = 7 for predation of both clutch and female; for details see Supplementary Material Table S2). Predation mostly occurred in uncultivated habitats (n = 27) with the highest number of cases in field margins (n = 16). From the remaining nests, three were destroyed by mowing, three were abandoned by the female and four nests had an unknown cause of failure. Nest location in the dominant habitat was the only significant predictor of nest success (Table 4). Nests located in the dominant habitat had more than twice lower failure rate than nests located in other remaining habitat categories (Fig. 8).

Daily nest failure rate (±SE) of nests located in the dominant habitat (cereal; n = 8 nests) and other habitats (n = 58 nests)
Discussion
Although the three study areas differed in their habitat composition, they demonstrate the conditions of typical agricultural landscapes in the Czech Republic well, where large cultivated areas dominate and contain a minimum of unmanaged grassy patches which provide perennial cover for grey partridges. Since similar landscape structures are also described from other Central European countries (Panek 1992; Kaiser 1998; Tworek 2002; Wübbenhorst and Leuschner 2006; Ronnenberg et al. 2016; etc.), our findings can be generalized to the agricultural landscape across Central Europe.
When comparing the habitat composition between the three different study areas, we clearly showed the importance of uncultivated habitats for grey partridge occurrence within the modern agricultural landscapes of Central Europe. Uncultivated habitats, especially field margins, ruderals and game refuges, were the most selected habitats. Selections for unmanaged habitats by grey partridge have been described in previous studies (e.g. Meriggi et al. 1991; Šálek et al. 2004; Buner et al. 2005; Rantanen et al. 2010), but there are some exceptions when partridges selected crops rather than unmanaged habitats (Bro et al. 2000a; Rantanen et al. 2010). We report similar results whereby cereal was the second most selected habitat in all study areas. It is possible that taller cereal crops provide better cover during the nesting period, as suggested by Bro et al. (2000a), and seem to provide better protection against predators (Whittingham and Evans 2004). However, our data showed that selections for cereal habitats were found not only for the nesting period but also for the pre-nesting period when vegetation was still low. It can therefore be assumed that the increased selection for cereal will be caused by other factors than solely vegetation cover.
Using both methods of compositional analysis we provide a comprehensive picture of habitat selections in the grey partridge. We analysed the selection of home ranges within the study areas (HRxAR) and also habitat utilisation on the basis of individual locations within home ranges (LxHR; Aebischer and Robertson 1992). Comparing these methods, we found that cereal was the second highest selected habitat in partridge home ranges (HRxAR), but when comparing cereal rank in the second habitat utilisation analysis, using partridge locations, we found selection to be much lower. Similarly, a study on reintroduced partridges in the UK showed differences between HRxAR and LxHR methods, whereby crops were the most selected habitat using HRxAR and margins were the most selected using LxHR methods in spring (Rantanen et al. 2010). Based on this, we suggest that cereal could be selected by chance due to its dominance and wide availability across our study areas. Partridges could be forced to use cereals due to their most selected habitats are surrounded with cereal monocultures.
Comparing selections between the HRxAR and LxHR methods also showed that partridges often occurred near infrastructure especially in Písek and Milešín. Whilst infrastructure was one of the most selected habitat using the HRxAR method, it was strongly avoided using the LxHR method. This could be explained (1) by a dense network of roads and tracks which may interfere with home ranges, (2) as a side effect of high selection for field margins which often lie next to the roads or (3) by the shift of partridges closer to human settlements. Harmange et al. (2019) showed the same trend in habitat selection by partridges, when selection increased for human infrastructure, especially for roads and tracks. Areas close to roads also have a higher probability of nest placement (Reitz et al. 2002). However, taking into account the results from Prague, where the most selected habitat was ruderal and the least selected habitat was infrastructure, it is more probable that selection for infrastructure is a side effect of selection for field margins.
We have shown that the grey partridge tends to place nests in the most selected habitats (field margins, ruderals or game refuges) and avoided nesting in the dominant habitat (cereal). A study from 1960s recorded similar results when 88% of partridge nests were placed in unmanaged or extensively grazed habitats (Bouchner and Fišer 1967). Furthermore, Blank et al. (1967) found the highest nest density in incomplete hedges and grass tracks representing typical uncultivated habitats. Bro et al. (2000a) found that partridges mostly selected cereal and linear habitat features as nest sites. However, the nests located in the cereals were in most cases within 20 m from the field edge (Bro et al. 2000a), thus suggesting partridges still nested near the edge and close to uncultivated linear habitats. Generally, there was an increased tendency of nest placement in cereal and cultivated habitats, likely caused by the lack of more selected (attractive) habitats compared to areas in Germany and Poland (Wübbenhorst and Leuschner 2006). This hypothesis is supported by our findings that the effort to place the nest in uncultivated habitats increases with the increasing area of the selected habitats (field margins, ruderals or game refuges) in pre-nesting home ranges. Also, a study on the red-legged partridge (Alectoris rufa) from Spain showed that cereal was the most common nesting habitat, even though it was not preferred by partridges overall (Casas and Viñuela 2010).
Nest site choice is one of the most important aspects affecting nest success (Ricklefs 1969). Nesting habitats can increase nest success primarily by providing cover from predators (Albrecht and Klvaňa 2004). Predation is the most common cause of partridge nest failure (Bro et al. 2000b). However, predation rates may vary between regions in respect to other factors such as gamekeeping practices (Jenkins 1961). Nesting success may be improved by predator control (Tapper et al. 1996), but there must be some aspects taken into account: (1) costs of predation control; (2) limited area where it is sustainable to carry out regular predator control; (3) short-term efficiency, as free space is quickly occupied by new individuals (Kuijper et al. 2009) and (4) targeted persecution of apex predators often leads to an increase of mesopredators (Evans 2004) which also have a significant negative impact via nest destruction (Bro et al. 2000b). In addition, predation rate is mostly related to habitat composition (Evans 2004). Therefore, recent studies suggest that improving agricultural practices could be more effective than predator control (Casas and Viñuela 2010; Knauer et al. 2010). It is advisable to try to identify the aspects needed for successful nesting and to support them using appropriate management techniques.
Our results do not directly show the effect of nest placement in selected habitats in regard to nest survival as we hypothesised. However, we found significant positive effects between nest placement in the dominant habitat and hatching success of at least one chick from the nest. Nests placed in the dominant habitat (cereal) had lower daily nest failure rates than nests located in other habitats. Bro et al. (2000b) achieved a similar result where nest survival was highest in cereal habitats. However, cereal is simultaneously the most selected nesting habitat in North-Central France, where the study took place (Bro et al. 2000a). On the contrary, partridges avoided nesting in cereal habitats in our study areas. Placing nests in cereal seems to be ‘the best of a bad situation’ within the modern agricultural landscape which lacks sufficient areas of unmanaged habitats. When we consider that there are a low proportion of selected habitats and uncultivated habitats in agricultural landscapes in general, this finding leads us to presume that selected habitats act as an ecological trap (Battin 2004). Rantanen et al. (2010) came to a similar conclusion when studying reintroduced grey partridges that the use of crops had positive effects whereas use of field margins had a negative effect on survival of adult birds. A convincing explanation can be that the selected habitats are often also selected by predators, which find more food here than elsewhere. A low proportion of uncultivated habitats results in a higher density of predators and prey in small isolated areas (Bro et al. 2004). In addition, uncultivated habitats are mostly on the edge of other habitats and the edge effect increases nest predation rates (Evans 2004). It is better for prey to occur in more represented habitats where detectability of prey by predators is lower due to larger and more homogeneous habitats.
However, aside from the lower predation risk, nests in cereal monocultures are also at risk from agricultural machinery (Jenkins 1961; Casas and Viñuela 2010). Incidents of nest failure were also observed in our study areas whereby two incubating hens were killed during the mowing of meadows (Rymešová et al. 2012). With the advancing breeding season, modern agricultural practices are an increasing threat to the success of partridge nests. Replacement clutches later in season suffered more from harvesting during July than the first clutches in France (Bro et al. 2000b). In addition to the direct destruction of nests by machinery, the effect of disturbances caused by humans resulting in nest abandonment is also important (Jenkins 1961). It is clear that a higher frequency of disturbances can be expected in managed areas compared to unmanaged habitats. With these findings we could, despite our results, hypothesise that increasing areas of unmanaged habitats in agricultural landscape could reduce predation risk of nests placed there. Therefore, it would be more convenient for grey partridges to place nests in uncultivated habitats within heterogeneous landscapes.
Heterogeneous landscapes dominated by small-sized fields and higher proportion of uncultivated habitats play an important role for the grey partridge (Šálek et al. 2004; Joannon et al. 2008). These two habitat features improve survival of partridges (Kaiser 1998) and also increase nest success (Casas and Viñuela 2010) which is the key factor for stability of grey partridge populations (Potts 1986; Bro et al. 2000c; Aebischer and Ewald 2004).
Data availability
The datasets used and analysed during the current study is available from the corresponding author on reasonable request.
References
Aebischer NJ, Ewald JA (2004) Managing the UK grey partridge Perdix perdix recovery: population change, reproduction, habitat and shooting. Ibis 146:181–191
Aebischer NJ, Ewald JA (2010) Grey partridge Perdix perdix in the UK: recovery status, set-aside and shooting. Ibis 152:530–542
Aebischer NJ, Robertson PA (1992) Practical aspects of compositional analysis as applied to pheasant habitat utilization. In: Priede IG, Swift SM (eds) Wildlife telemetry: remote monitoring and tracking of animals. Elis Horwood, Chichester, pp 285–293
Aebischer NJ, Robertson PA, Kenward RE (1993) Compositional analysis of habitat use from animal radio-tracking data. Ecology 74:1313–1325
Albrecht T, Klvaňa P (2004) Nest crypsis, reproductive value of a clutch and escape decisions in incubating female mallards Anas platyrhynchos. Ethology 110:603–613
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48
Battin J (2004) When good animals love bad habitats: ecological traps and the conservation of animal populations. Conserv Biol 18:1482–1491
Benítez-Lopéz A, Mougeot F, Martín CA, Casas F, Calero-Riestra M, García JT, Viñuela J (2011) An improved night-lighting technique for the selective capture of sandgrouse and other steppe birds. Eur J Wildl Res 57:389–393
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Blank TH, Southwood TRE, Cross DJ (1967) The ecology of the partridge: I. Outline of population processes with particular reference to chick mortality and nest density. J Anim Ecol 36:549–556
Bouchner M, Fišer Z (1967) Příspěvek ke hnízdní bionomii koroptví a ke ztrátám na hnízdech a vejcích. Lesnický Časopis 13:713–725
Bro E, Reitz F, Clobert J, Mayot P (2000a) Nest-site selection of grey partridge (Perdix perdix) on agricultural lands in north-central France. Game and Wildlife Science 17:1–16
Bro E, Reitz F, Clobert J, Mayot P (2000b) Nesting success of grey partridge (Perdix perdix) on agricultural land in North-Central France, relation to nesting cover and predator abundance. Game and Wildlife Science 17:199–218
Bro E, Sarrazin F, Clobert J, Reitz F (2000c) Demography and the decline of the grey partridge Perdix perdix in France. J Appl Ecol 37:432–448
Bro E, Mayot P, Corda E, Reitz F (2004) Impact of habitat management on grey partridge populations: assessing wildlife cover using a multisite BACI experiment. J Appl Ecol 41:846–857
Buner F, Jenny M, Zbinden N, Naef-Daenzer B (2005) Ecologically enhanced areas – a key habitat structure for re-introduced grey partridges Perdix perdix. Biol Conserv 124:373–381
Calenge C (2006) The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals. Ecol Model 197:516–519
Casas F, Viñuela J (2010) Agricultural practices or game management: which is the key to improve red-legged partridge nesting success in agricultural landscape? Environ Conserv 37:177–186
Černý M, Rymešová D, Šálek M (2018) Switches in covering of eggs in grey partridge Perdix perdix clutches during laying and incubation. Folia Zool 67:154–157
Chamberlain DE, Fuller RJ, Bunce RGH, Duckworth JC, Shrubb M (2000) Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. J Appl Ecol 37:771–788
Cramp S, Simmons KEL (1982) The birds of the Western Palearctic, Vol. 2. Oxford University Press, Oxford
Donald PF, Green RE, Heath MF (2001) Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc R Soc Lond B 268:25–29
Evans KL (2004) The potential for interactions between predation and habitat change to cause population declines of farmland birds. Ibis 146:1–13
Frenzel M, Everaars J, Schweiger O (2016) Bird communities in agricultural landscapes: what are the current drivers of temporal trends? Ecol Indic 65:113–121
Harmange C, Bretagnolle V, Sarasa M, Pays O (2019) Changes in habitat selection patterns of the gray partridge Perdix perdix in relation to agricultural landscape dynamics over the past two decades. Ecol Evol 9:5236–5247. https://doi.org/10.1002/ece3.5114
Jenkins D (1961) Population control in protected partridges (Perdix perdix). J Anim Ecol 30:235–258
Joannon A, Bro E, Thenail C, Baudry J (2008) Crop patterns and habitat preferences of the grey partridge farmland bird. Agron Sustain Dev 28:379–387
Kaiser W (1998) Grey partridge (Perdix perdix) survival in relation to habitat quality. Gibier Faune Sauvage 15:157–162
Kenward RE (2001) A manual for wildlife radio tagging. Academic Press, London
Knauer F, Küchenhoff H, Pilz S (2010) A statistical analysis of the relationship between red fox Vulpes vulpes and its prey species (grey partridge Perdix perdix, brown hare Lepus europaeus and rabbit Oryctolagus cuniculus) in Western Germany from 1958 to 1998. Wildlife Biol 16:56–65
Kreisinger J, Albrecht T (2008) Nest protection in mallards Anas platyrhynchos: untangling the role of crypsis and parental behaviour. Funct Ecol 22:872–879
Kuijper DPJ, Oosterveld E, Wymenga E (2009) Decline and potential recovery of the European grey partridge (Perdix perdix) population - a review. Eur J Wild Res 55:455–463
Lack D (1947) The significance of clutch-size in the partridge (Perdix perdix). J Anim Ecol 16:19–25
Martin TE, Scott J, Menge C (2000) Nest predation increases with parental activity: separating nest site and parental activity effects. Proc R Soc Lond B 267:2287–2293
Mayfield H (1975) Suggestions for calculating nest success. Wilson Bull 87:456–466
Meriggi A, Montagna D, Zacchetti D (1991) Habitat use by partridges (Perdix perdix and Alectoris rufa) in an area of northern Apennines, Italy. Boll Zool 58:85–90
Newton I (2004) The recent declines of farmland bird populations in Britain, an appraisal of causal factors and conservation actions. Ibis 146:579–600
Novoa C, Aebischer NJ, Landry P (2002) Upland habitat use by Pyrenean grey partridge Perdix perdix hispaniensis during the breeding season. Wildlife Biol 8:99–108
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O'Hara RH, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2019) vegan: Community Ecology Package. R Package Version 2:5–5 https://CRAN.R-project.org/package=vegan
Opermanis O (2004) Appearance and vulnerability of artificial duck nests to avian predators. J Avian Biol 35:410–415
Panek M (1992) The effect of environmental factors on survival of grey partridge (Perdix perdix) chicks in Poland during 1987–89. J Appl Ecol 29:745–750
Panek M (1997) Density-dependent brood production in the grey partridge Perdix perdix in relation to habitat quality. Bird Study 44:235–238
Potts GR (1986) The partridge: herbicides, predation and conservation. Collins, London
Prokop P, Trnka A (2011) Why do grebes cover their nests? Laboratory and field tests of two alternative hypotheses. J Ethol 29:17–22
R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Rands MRW (1988) The effect of nest site selection on nest predation in grey partridge Perdix perdix and red-legged partridge Alectoris rufa. Ornis Scand 19:35–40
Rantanen EM, Buner F, Riordan P, Sotherton N, Macdonald DW (2010) Habitat preferences and survival in wildlife reintroductions: an ecological trap in reintroduced grey partridges. J Appl Ecol 47:1357–1364
Reif J, Vermouzek Z (2019) Collapse of farmland bird populations in an Eastern European country following its EU accession. Conserv Lett 12:e12585
Reitz F, Le Goff E, Fuzeau M (2002) Landscape selection by grey partridge (Perdix perdix) for nesting in the fields of French cereal agrosystems. Game and Wildlife Science 19:209–220
Ricklefs RE (1969) An analysis of nesting mortality in birds. Smithson Contrib Zool 9:1–48
Robinson RA, Sutherland WJ (2002) Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 39:157–176
Ronnenberg K, Strauß E, Siebert U (2016) Crop diversity loss as a primary cause of grey partridge and commnon pheasant decline in Lower Saxony, Germany. BMC Ecol 16:39. https://doi.org/10.1186/s12898-016-0093-9
Rymešová D, Šmilauer P, Šálek M (2012) Sex- and age-biased mortality in wild grey partridge Perdix perdix populations. Ibis 154:815–824
Rymešová D, Tomášek O, Šálek M (2013) Differences in mortality rates, dispersal distances and breeding success of commercially reared and wild grey partridges in the Czech agricultural landscape. Eur J Wildl Res 59:147–158
Šálek M, Marhoul P (2008) Spatial movements of grey partridge Perdix perdix: male-biased spring dispersal and effect of habitat quality. J Ornithol 149:329–335
Šálek M, Marhoul P, Pintíř J, Kopecký T, Slabý L (2004) Importance of unmanaged wasteland patches for grey partridge Perdix perdix in suburban habitats. Acta Oecol 25:23–33
Sasaki K, Hotes S, Kadoya T, Yoshioka A, Wolters V (2020) Landscape associations of farmland bird diversity in Germany and Japan. Glob Ecol Conserv 21:e00891
Siriwardena GM, Baillie SR, Buckland ST, Fewster RM, Marchant JH, Wilson JD (1998) Trends in the abundance of farmland birds: a quantitative comparison of smoothed common birds census indices. J Appl Ecol 35:24–43
Skutch AF (1949) Do tropical birds rear as many young as they can nourish? Ibis 91:430–455
Tapper SC, Potts GR, Brockless MH (1996) The effect of an experimental reduction in predation pressure on the breeding success and population density of grey partridge Perdix perdix. J Appl Ecol 33:965–978
Traba J, Morales MB (2019) The decline of farmland birds in Spain is strongly associated to the loss of fallowland. Sci Rep 9:9473. https://doi.org/10.1038/s41598-019-45854-0
Tworek S (2002) Different bird strategies and their responses to habitat changes in an agricultural landscape. Ecol Res 17:339–359
Vickery JA, Bradbury RB, Henderson IG, Eaton MA, Grice PV (2004) The role of agri-environment schemes and farm management practices in reversing the decline of farmland birds in England. Biol Conserv 119:19–39
Whittingham MJ, Evans KL (2004) The effects of habitat structure on predation risk of birds in agricultural landscapes. Ibis 146:210–220
Wübbenhorst D, Leuschner C (2006) Vegetation structure at the breeding sites of the partridge (Perdix perdix L.) in central Europe and its possible importance for population density. Pol J Ecol 54:57–67
Acknowledgements
We thank all our collaborators and the anonymous referees for their valuable comments. We also thank Connor Panter for English corrections.
Code availability
Not applicable
Funding
The study was supported by the Czech Science Foundation (GA206/08/1281) and appears through the institutional support of long-term conceptual development of research institutions provided by the Ministry of Culture of the Czech Republic (ref. MK000094862).
Author information
Authors and Affiliations
Contributions
MŠ and MČ designed the study. All authors collected data using radio tracking. MČ analysed the data. MČ contributed most to the writing of the paper, which was reviewed and approved by DR and MŠ.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interests.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Černý, M., Rymešová, D. & Šálek, M. Habitat scarcity forms an ecological trap for the grey partridge (Perdix perdix) within a central European agricultural landscape. Eur J Wildl Res 66, 83 (2020). https://doi.org/10.1007/s10344-020-01422-w
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-020-01422-w