
Abstract
Ruminants are classified into three groups, according to their feeding behaviour: browsers, intermediate feeders and grazers. Corresponding to their dietary preferences, multiple morphological and physiological adaptations have been described, resulting in another classification: ‘moose-type’ and ‘cattle-type’ ruminants. Digesta retention patterns in the gastrointestinal tract (GIT) and reticulorumen (RR) are considered major criteria to distinguish these types, as cattle-type ruminants show shorter retention of fluids (measured by a solute marker) than of particles, while in moose-type ruminants, both are retained for more similar periods. To what extent these digestive types are specific to phylogenetic lineages is still unclear. We measured mean retention times (MRTs) of solutes and particles (2 and 20 mm) in the strictest grazing cervid: the Père David’s deer (Elaphurus davidianus; n = 5; body mass = 155.0 ± 14.5 kg). The MRTs of solutes, small and large particles in the GIT were 34 ± 4, 60 ± 7 and 69 ± 9 h, respectively. The ratio of the MRT of small particles versus solutes in the RR was 2.0 ± 0.1, similar to other cattle-type ruminants. The results confirm the hypothesis that Père David’s deer can be classified as cattle-type ruminants, corresponding to both dietary preferences and previously described morphological traits. The results complement previous findings, showing that both cattle-type and moose-type physiologies are found among bovids as well as cervids, indicating that these digestion types can be considered convergent adaptations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ruminants have been classified into three feeding types (browser, intermediate feeders and grazers) (Hofmann and Stewart 1972) and two digestion types (‘moose-type’ and ‘cattle-type’) (Clauss et al. 2010). Moose-type ruminants are typically browsers, whereas cattle-type ruminants cover the spectrum of intermediate feeders and grazers (Codron and Clauss 2010). Some important morphological differences between moose- and cattle-type ruminants are associated with the function of the rumen. The moose-type’s ruminal mucosa shows papillation that is more homogeneously distributed over the whole surface, while the papillation in cattle-types is less distinct in the dorsal and ventral, but prominent in the middle rumen (Clauss et al. 2009b). The uneven papillation is an indication for stratified rumen contents, which are seen in cattle-type ruminants, with more moisture in the ventral part of the rumen (Clauss et al. 2009a; Codron and Clauss 2010; Tahas et al. 2017). Stratified rumen contents are linked to a higher rumen fluid throughput in cattle-type ruminants, and therefore, a major physiological difference between moose-type and cattle-type ruminants is that in the former, fluids and particles leave the rumen nearly in parallel, whereas in the latter, particles are retained much longer and hence ‘washed’ by rumen fluid (Dittmann et al. 2015a). Across ruminant species, the difference in moisture content between dorsal and ventral rumen (content stratification), the papillation gradient (papillation stratification) and the degree by which fluids leave the rumen sooner than particles (digesta washing) appear to be interrelated (Tahas et al. 2017).
Lechner-Doll et al. (1990) called the difference in mean retention time (MRT) of fluids and particles the ‘selectivity factor’ (SF), expressed as the ratio of MRTparticle/MRTsolute in the reticulorumen (RR). Low SF values are present in tragulids (Darlis et al. 2012), giraffids (Hummel et al. 2005), a number of cervids (Behrend et al. 2004; Lechner et al. 2010) and some bovid species (Clauss et al. 2011), suggesting either a convergence in this characteristic in browsing species, or that this characteristic represents the plesiomorphic state in ruminants. While high SF values have been reported in a variety of bovid species (Dittmann et al. 2015a), few SFs have been reported for cervid species that are not considered strict browsers, namely the wapiti (Cervus elaphus) with a SFRR of 2.1 (Renecker and Hudson 1990) and the reindeer (Rangifer tarandus) with a SFRR of 1.7 (Lechner et al. 2010). Notably, data on MRT and SF in a cervid species that is considered a grazer has been lacking so far.
One reason might be the general lack of truly grazing species among cervids. According to Hofmann (1985), some deer species are browsers and all other are intermediate feeders, with no strict grazing species. Others disagree and count for example the hog deer (Axis porcinus) (Asher 2011) and the axis deer (Axis axis) (Pérez et al. 2015) among grazers, as they primarily graze during the summer. In a later publication, Hofmann (1991) himself considered the Père David’s deer (Elaphurus davidianus) a grazer. While the literature is divided on whether Père David’s deer should be considered grazers or intermediate feeders (Beck and Wemmer 1983; Van Wieren 1996; Jiang and Takatsuki 1999), with a diet that contains approximately 75% of grass, they are considered the ‘most grazing’ species amongst cervids (e.g. Cantalapiedra et al. 2014), although this information most likely only derives from a very general statement in Nowak and Paradiso (1999). In addition, Père David’s deer show morphological traits that suggest that they are ruminants of the cattle-type, with a stratified rumen papillation (Clauss et al. 2009b). In the present study, the MRTs of solute and particle markers in the GIT of Père David’s deer were investigated, to test whether its digestive physiology matches the ruminal morphology and the vague botanical classification.
Materials and methods
This study was performed under animal experiment licence 84-02.04.2016.A363 at the Zoological Garden of Wuppertal, Germany. Five female Père David’s deer (body mass BM 155.0 ± 14.5 kg) were adapted to a diet of only grass hay, fed for ad libitum consumption, in their usual group enclosure. Animals had been weighed recently during immobilisation for the application of ear tags and for a general clinical examination. They were sequentially assessed in individual housing between September 20th and November 6th of 2017. Individual housing was either achieved in a 4.2 × 4.0 m compartment with paved floor (animals 1–2) or a 5.7 × 4.7 m compartment with concrete floor (animals 3–5), which were both located in larger stables and had empty neighbouring enclosures to allow a shifting of the animals during feeding, cleaning and faeces collection. The enclosure floors were covered with sawdust. Throughout the whole study, the animals had ad libitum access to clean drinking water and the grass hay. New hay was offered every day at the same hour, to maintain a constant feeding routine. In the individual compartments, animals were maintained first for an adaptation period of 7 days and a subsequent collection period of 8 days. During the adaptation period, animals were habituated to being offered, always at the same hour of the day, a separate meal of 80 g wheat bran mixed with small hay particles, water and red beet juice (imitating the colour of Co-EDTA), in order to habituate them to this meal and thus guarantee unproblematic ingestion when the passage markers were to be applied in a meal on the last day of adaptation. During the collection period, hay intake was monitored by weighing offered and refused hay on a daily basis. Every day, hay samples (offered and refused) were taken and kept at room temperature for later analysis.
Three markers were used to measure retention times: Co-EDTA as a solute marker for the fluid digesta fraction, Cr-mordanted fibre for small particles (< 2 mm) and La-mordanted fibre for larger particles (~ 20 mm), prepared according to Udén et al. (1980) and Schwarm et al. (2009). The particle markers contained 36.9 g Cr and 20.9 g La/kg dry matter, respectively. The markers were fed at approximately 0.1 g/kg BM for the fibre mordants and 0.02 g/kg BM for the Co-EDTA. The marker meal was offered at the last day of the adaptation period and was consumed within 15 min by all animals.
For faeces collection, animals were shifted from the main compartment to the neighbouring pen. All faeces were collected in toto from the compartment, cleaned of sawdust, and weighed. Thereafter, a representative sample of approximately 200 g fresh weight was stored in a tared aluminium dish at − 20 °C until further processing. Prior to marker feeding, three faecal samples were taken on 48 h, 24 h and 12 h before the marker was fed, to determine baseline concentrations of Co, Cr and La. After marker feeding, faeces were collected regularly for 8 days, with intensive sampling during the first 2 days (every 4 h) and increasing time intervals thereafter: on the third day every 6 h, on the fourth and fifth days every 8 h, and on the sixth, seventh and eighth days every 12 h. It was assumed that the markers would be excreted completely within 8 days. As this was not always the case (see ‘Results’), the resulting MRT values represent slight underestimates.
Representative samples of the offered and refused hay were ground to pass through a 2-mm mesh sieve. Thereafter, samples were subjected to standard nutrient analyses (AOAC 1995) for dry matter (DM) and total ash (AOAC no. 942.05), crude protein (AOAC no. 977.02), crude fibre (AOAC no. 930.10), crude fat (AOAC no. 920.39, only the hay offered), and neutral detergent fibre (NDF, AOAC no. 2002.04; corrected for residual ash). For each animal, a portion of every faecal sample (representing a constant proportion of the daily faeces output) was pooled, dried at 60 °C and then ground using a blender and analysed for DM, crude ash, crude protein, crude fibre, NDF) with the same methods.
The individual faeces samples were dried at 105 °C, ground with a blender and used for marker analysis by inductively coupled plasma optical emission spectrometer (model Optima 8000, PerkinElmer, Rodgau, Germany) according to Frei et al. (2015). The MRT in the whole digestive tract was calculated according to Thielemans et al. (1978) as:
with Ci = marker concentration in the faecal samples from the interval represented by time ti (h after marker administration, using the midpoint of the sampling interval) and dti = the interval (h) of the respective sample
Complete excretion of the markers was assumed once the faecal marker concentrations were similar to the background levels determined in pre-dose faecal samples. Mean retention time in the RR was estimated following Lechner-Doll et al. (1990). The MRTsoluteRR is determined by estimating the rate constant of the descending part of the marker excretion curve via an exponential equation:
with y = faecal marker concentration at time t (mg/kg DM), A = a constant, k = rate-constant (h−1) and t = time after marker dosing (h); the reciprocal of k represents the MRT for the RR. The MRTparticleRR is calculated based on the assumption that fluid and particles do not differ in passage characteristics distal to the RR (Mambrini and Peyraud 1997) and that ruminal dilution rate is covered by the rate constant of the decreasing part of the marker excretion pattern alone:
The selectivity factor (SF, the ratio of MRTparticle/MRTsolute) was calculated for both the total GIT and the RR.
Results
Nutritional values of the feed are presented in Table 1. Differences between offered and leftover hay reflect selective feeding. Of the total amount of offered hay, 52 ± 10% was refused on a dry matter basis. The refused hay contained more crude fibre (− 0.3–6.4%) and NDF (0.6–6.9%) and less protein (− 1.0 − − 0.3%) than the offered hay. Daily food intake averaged at 2002 ± 247 g DM, and faecal excretion at 769 ± 34 g DM. The resulting apparent digestibility coefficients for dry matter, organic matter, crude protein, crude fibre and NDF were 61.0 ± 6.0, 62.4 ± 6.1, 52.2 ± 6.4, 54.4 ± 10.9 and 55.0 ± 6.9%.
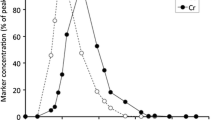
The excretion patterns of the passage markers are shown in Fig. 1. Approximately 16 h after marker administration, the excretion started, for both fluid and particles. Thereafter, maximum concentrations of Co-EDTA, Cr-mordanted fibre and La-mordanted fibre were detected 20.8 ± 3.3, 32.8 ± 3.3 and 50.4 ± 14.5 h after the marker was consumed, respectively. Complete excretion of the fluid marker was finished after an average of 6.3 days, while both particle markers were still detected in the faeces after the full experimental phase of 8 days. Yet, at that point, they were present at very low concentrations (4 ± 4% of peak concentration).

Faecal excretion patterns for solute, small particle (2 mm) and large particle (20 mm) markers in five Père David’s deer (% of peak concentration)
On average, the MRTparticle in the GIT was 59.8 ± 6.8 h for small particles (2 mm) and 69.3 ± 9.1 h for large particles (20 mm). Shorter retention times were measured for solutes, with an average MRTsoluteGIT of 33.8 ± 4.4 h. This was also found to be true for retention in the RR, where small particles, large particles and fluids were excreted approximately after 52.9 ± 6.6, 62.4 ± 8.7 and 26.9 ± 4.2 h. Based on MRT values of small particles versus those of solutes, the SF was 1.8 ± 0.1 for the whole GIT and 2.0 ± 0.1 for the RR (Table 2).
Discussion
The present study shows that the SF in Père David’s deer corresponds to values described for other grazing ruminants. This confirms the occurrence of cattle-type digestion in a cervid species, strongly suggesting that ruminant digestion types are convergent adaptations.
Typical constraints for zoo-based research apply: The number of animals was limited, and it was not considered justifiable to assess individual animals more than once, for example on different diets, different feed intake levels, or on the same diet to evaluate the repeatability of the results. Therefore, the results must be considered with caution, although the similarity between the five individuals at least suggests that the results could be representative for the species. It should be noted that in ruminants, apparent protein digestibilities do not mainly indicate protein degradability, but simply protein intake.
An important constraint of this study is the method of marker administration via voluntary consumption. This might have had an effect on particle markers, if the mordanted fibres were chewed on by the animal. Options to circumvent this situation were not given in the present study: On the one hand, the use of fistulae (Lechner et al. 2010) is rarely possible for wild ruminants, and fistulation for such just a single experiment does not appear ethically justified. On the other hand, the practice of feeding a marked forage whole (i.e., not chopped or ground) and determining the mean retention times of different faecal particle size fractions does not allow an estimation of large particle retention in ruminants (Hummel et al. 2018). Their comminution during rumination makes them indistinguishable from smaller particles that are excreted sooner. Another possible limitation could exist in the accuracy in faeces and leftover hay collection. To minimise errors in precision, collection was carried out by the same person during the whole trial. A third problem is the possible effect of the unusual stress of the experimental setup, including the fact that animals normally kept in a group are kept individually. While the adaptation period, which also included habituation to the investigators’ presence and the management routines of feeding and faeces collection, most likely reduced such stress to a considerable extent, its effect cannot be excluded or reasonably assessed.
As the animals were fed ad libitum hay with ample amounts of leftovers each day (2211 ± 641 g), they had the opportunity to feed selectively. The ingested hay had slightly lower levels of crude fibre and NDF and higher protein levels, indicating that the animals used this opportunity to a certain extent. In the present study, animals digested 55.0 ± 6.9% of NDF. In a data collection of 37 studies on 24 ruminant species, the NDF digestibility of very strict browsers ranged approximately between 25 and 50%, whereas grazers had a range of 50–65% (Pérez-Barbería et al. 2004). The values of our Père David’s deer therefore fall into the ‘grazer’ category. The difference in NDF digestibility between species is most likely a result of differences in retention time of the fibre, as longer retention facilitates a more thorough digestion by the commensal rumen bacteria (Hummel et al. 2006).
With respect to particle sorting, combining the results of the present study with the data collection of Dittmann et al. (2015b) (Fig. 2) indicates that the particle size discrimination mechanism in Père David’s deer is similar to that observed in other ruminants. For absolute retention times, Dittmann et al. (2015a) noticed that the MRTparticleRR was significantly related to the percentage of grass (%grass) in the natural diet across ruminant species, although significance became only marginal once BM was also accounted for in the analysis. In our Père David’s deer, MRTparticleRR were relatively high compared to values measured in other ruminants with comparable %grass in their natural diet, such as Oryx gazella (46.0 h) or Antilope cervicapra (45.0 h) (Dittmann et al. 2015a). An extended particle retention has often been suggested to be a logical consequence of a grass diet. Grass contains a higher content of digestible fibre, and therefore long retention times in the RR are an expected adaptation of grazing ruminants (Hummel et al. 2006; Dittmann et al. 2015a). This does not seem to be the case for the retention of fluid. MRTsoluteRR itself is neither linked to %grass in the diet nor to BM, but is likely species-specific (Dittmann et al. 2015a). In particular, the relationship between MRTsoluteRR and MRTparticleRR is considered an indicator for digestion type (with a higher difference in cattle-type ruminants) (Clauss et al. 2010): Cattle-type ruminants show SF values in the reticulorumen from 1.7 to 4.6, whereas the SF of moose-type ruminants is much lower (1.1–1.8) (Hummel et al. 2005). The relationship between MRTsoluteRR and MRTparticleRR has been qualitatively linked to categorical feeding types (Clauss and Lechner-Doll 2001), shown to differ quantitatively between categorical feeding types (Clauss et al. 2006), as well as shown to correlate significantly with the %grass in the natural diet as a continuous variable for feeding types (Dittmann et al. 2015a). Adding our data of the Père David’s deer to the data collection of Dittmann et al. (2015a) (Fig. 3) shows that Père David’s deer represent the most extreme cervid investigated so far and that cervid species are distributed across the whole data range covered by bovid ruminants. When plotting the average SF measured in our Père David’s deer against data from other animals on the intraruminal papillation pattern (Fig. 4), Père David’s deer match the overall ruminant pattern of an increased difference in intraruminal papillation with a higher SF. This has been explained by the triggering of rumen contents stratification by a high SF, consequently resulting in a stratification of the intraruminal papillation pattern (Tahas et al. 2017).

Relationship of the mean retention time (MRT) between small and large particles. Data collection for ruminants (RUM) from Dittmann et al. (2015b) with data from the current study on Père David’s deer (PDD, Elaphurus davidianus). The dotted line denotes y = x

Relationship between the mean retention time (MRT) of small particles and a solute marker in the reticulorumen (RR) of various ruminant species (from Dittmann et al. 2015a). Among the species consuming more than 20% grass in their natural diet, cervids are in grey

Relationship between the ruminal papillation gradient (expressed as the surface enlargement factor SEF of the dorsal rumen in % of the SEF of the atrium ruminis) and the selectivity factor (SF) in the reticulorumen (RR) in ruminant species from Tahas et al. (2017) and the Père David’s deer (PDD, Elaphurus davidianus) of the present study. Note that a lower SEF percentage indicates a more stratified ruminal papillation, and a higher SF a more distinct difference in the retention of particles and fluids in the RR
It remains unclear whether the moose-type or the cattle-type morphophysiology represents the original or a derived set of characteristics. Browsing ruminants have been considered the ‘original’ ruminants (Hofmann 1989), which was also based on the fact that many families that hold only few species (such as tragulid, giraffid, pronghorn and musk deer) involve browsers. However, various lines of evidence from analyses of the cervid and bovid families that include many more species have insinuated that current information is more compatible with considering the state of mixed feeders (and therefore, by derivation, a cattle-type digestive physiology) as the ancestral state of ruminants, with strict browsing and strict grazing both representing secondary adaptations (Codron et al. 2008; DeMiguel et al. 2008; Cantalapiedra et al. 2014; Cerling et al. 2015). At present, it cannot be decided whether high or low SF is the ancestral condition, but the fact that both occur in cervids and bovids opens the possibility that a high SF, aimed at microbial harvesting, was an early adaptation of ancestral ruminants.
References
AOAC (1995) Official methods of analysis of AOAC International. Association of Official Analytical Chemists, Arlington VA
Asher GW (2011) Reproductive cycles of deer. Anim Reprod Sci 124:170–175
Beck B, Wemmer C (1983) The biology and management of an extinct species: Père David’s deer. Noyes Publications, Parkridge
Behrend A, Lechner-Doll M, Streich WJ, Clauss M (2004) Seasonal faecal excretion, gut fill, liquid and particle marker retention in mouflon (Ovis ammon musimon), and a comparison with roe deer (Capreolus capreolus). Acta Theriol 49:503–515
Cantalapiedra JL, FitzJohn RG, Kuhn TS, Fernández MH, DeMiguel D, Azanza B, Morales J, Mooers AØ (2014) Dietary innovations spurred the diversification of ruminants during the Caenozoic. Proc R Soc B 281:20132746
Cerling TE, Andanje SA, Blumenthal SA, Brown FH, Chritz KL, Harris JM, Hart JA, Kirera FM, Kaleme P, Leakey LN, Leakey MG (2015) Dietary changes of large herbivores in the Turkana Basin, Kenya from 4 to 1 ma. PNAS 112:11467–11472
Clauss M, Lechner-Doll M (2001) Differences in selective reticulo-ruminal particle retention as a key factor in ruminant diversification. Oecologia 129:321–327
Clauss M, Hummel J, Streich WJ (2006) The dissociation of the fluid and particle phase in the forestomach as a physiological characteristic of large grazing ruminants: an evaluation of available, comparable ruminant passage data. Eur J Wildl Res 52:88–98
Clauss M, Fritz J, Bayer D, Nygren K, Hammer S, Hatt J-M, Südekum K-H, Hummel J (2009a) Physical characteristics of rumen contents in four large ruminants of different feeding type, the addax (Addax nasomaculatus), bison (Bison bison), red deer (Cervus elaphus) and moose (Alces alces). Comp Biochem Physiol A 152:398–406
Clauss M, Hofmann RR, Fickel J, Streich WJ, Hummel J (2009b) The intraruminal papillation gradient in wild ruminants of different feeding types: implications for rumen physiology. J Morphol 270:929–942
Clauss M, Hume ID, Hummel J (2010) Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 4:979–992
Clauss M, Lunt N, Ortmann S, Plowman A, Codron D, Hummel J (2011) Fluid and particle passage in three duiker species. Eur J Wildl Res 57:143–148
Codron D, Clauss M (2010) Rumen physiology constrains diet niche: linking digestive physiology and food selection across wild ruminant species. Can J Zool 88:1129–1138
Codron D, Brink JS, Rossouw L, Clauss M (2008) The evolution of ecological specialization in southern African ungulates: competition- or physical environmental turnover? Oikos 117:344–353
Darlis NA, Liang JB, Ho YW (2012) Effects of diets of differing fiber contents on digestibility, passage rate of digesta and heat production in lesser mouse deer (Tragulus javanicus). Mamm Biol 77:385–390
DeMiguel D, Fortelius M, Azanza B, Morales J (2008) Ancestral feeding state of ruminants reconsidered: earliest grazing adaptation claims a mixed condition for Cervidae. BMC Evol Biol 8:13
Dittmann MT, Hummel J, Hammer S, Arif A, Hebel C, Müller DWH, Fritz J, Steuer P, Schwarm A, Kreuzer M, Clauss M (2015a) Digesta retention in gazelles in comparison to other ruminants: evidence for taxon-specific rumen fluid throughput to adjust digesta washing to the natural diet. Comp Biochem Physiol A 185:58–68
Dittmann MT, Runge U, Ortmann S, Lang RA, Moser D, Galeffi C, Schwarm A, Kreuzer M, Clauss M (2015b) Digesta retention patterns of solutes and different-sized particles in camelids compared with ruminants and other foregut fermenters. J Comp Physiol B 185:559–573
Frei S, Ortmann S, Reutlinger C, Kreuzer M, Hatt J-M, Clauss M (2015) Comparative digesta retention patterns in ratites. Auk Ornithol Adv 132:119–131
Hofmann RR (1985) Digestive physiology of deer - their morphophysiological specialisation and adaptation. Roy Soc N Z Bull 22:393–407
Hofmann RR (1989) Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78:443–457
Hofmann RR (1991) Endangered tropical herbivores - their nutritional requirements and habitat demands. In: Ho YW, Wong HK, Abdullah N, Tajuddin ZA (eds) Recent advances on the nutrition of herbivores. Malaysia Society of Animal Production, UPM Serdang, pp 27–34
Hofmann RR, Stewart DRM (1972) Grazer or browser: a classification based on the stomach-structure and feeding habit of east African ruminants. Mammalia 36:226–240
Hummel J, Clauss M, Zimmermann W, Johanson K, Norgaard C, Pfeffer E (2005) Fluid and particle retention in captive okapi (Okapia johnstoni). Comp Biochem Physiol A 140:436–444
Hummel J, Südekum K-H, Streich WJ, Clauss M (2006) Forage fermentation patterns and their implications for herbivore ingesta retention times. Funct Ecol 20:989–1002
Hummel J, Scheurich F, Ortmann S, Crompton LA, Gerken M, Clauss M (2018) Comparative selective retention of particle size classes in the gastrointestinal tract of ponies and goats. J Anim Physiol Anim Nutr 102:429–439
Jiang Z, Takatsuki S (1999) Constraints on feeding type in ruminants: a case for morphology over phylogeny. Mammal Study 24:79–89
Lechner I, Barboza P, Collins W, Fritz J, Günther D, Hattendorf B, Hummel J, Südekum K-H, Clauss M (2010) Differential passage of fluids and different-sized particles in fistulated oxen (Bos primigenius f. taurus), muskoxen (Ovibos moschatus), reindeer (Rangifer tarandus) and moose (Alces alces): rumen particle size discrimination is independent from contents stratification. Comp Biochem Physiol A 155:211–222
Lechner-Doll M, Rutagwenda T, Schwartz HJ, Schultka W, von Engelhardt W (1990) Seasonal changes of ingesta mean retention time and forestomach fluid volume in indigenous camels, cattle, sheep and goats grazing in a thornbush savanna pasture in Kenya. J Agric Sci 115:409–420
Mambrini M, Peyraud JL (1997) Retention time of feed particles and liquids in the stomachs and intestines of dairy cows. Direct measurement and calculation based on fecal collection. Reprod Nutr Dev 37:427–442
Nowak RM, Paradiso JL (1999) Walker's Mammals of the world, 6th edn. The Johns Hopkins University Press, Baltimore and London
Pérez W, Erdogan S, Ungerfeld R (2015) Anatomical study of the gastrointestinal tract in free-living Axis deer (Axis axis). Anat Histol Embryol 44:43–49
Pérez-Barbería FJ, Elston DA, Gordon IJ, Illius AW (2004) The evolution of phylogenetic differences in the efficiency of digestion in ruminants. Proc R Soc B 271:1081–1090
Renecker LA, Hudson RJ (1990) Digestive kinetics of moose (Alces alces) , wapiti (Cervus elaphus) and cattle. Br Soc Anim Prod 50:51–61
Schwarm A, Ortmann S, Wolf C, Clauss M (2009) No distinct difference in the excretion of large particles of varying size in a wild ruminant, the banteng (Bos javanicus). Eur J Wildl Res 55:531–533
Tahas SA, Martin Jurado O, Hammer S, Arif A, Reese S, Hatt J-M, Clauss M (2017) Gross measurements of the digestive tract and visceral organs of addax antelope (Addax nasomaculatus) following a concentrate or forage feeding regime. Anat Histol Embryol 46:282–293
Thielemans MF, François E, Bodart C, Thewis A (1978) Mesure du transit gastrointestinal chez le porc a l'aide des radiolanthanides. Comparaison avec le mouton. Ann Biol Anim Biochim Biophys 18:237–247
Udén P, Colucci PE, Van Soest PJ (1980) Investigation of chromium, cerium and cobalt as markers in digesta. Rate of passage studies. J Sci Food Agric 31:625–632
Van Wieren SE (1996) Browser and grazers: foraging strategies in ruminants. In: Van Wieren SE (ed) Digestive strategies in ruminants and nonruminants. Thesis Landbouw. University of Wageningen, Wageningen, pp 119–146
Acknowledgements
We thank Heidrun Barleben for the marker analyses and Herman de Rycke for the analyses on nutrition and faeces. We also thank the zookeepers of Zoo Wuppertal for support and assistance in the handling of the animals, particularly the following: Jürgen Abend, Frank Soumagne, Andrea Becker, Boris Kostka, Jörg Effenberger, Kevin Büttner, Roland Stöter and Malena Reeh. Furthermore, special thanks to veterinary nurses Laura Cyrener and Katrin Gries, and zoo curators Silja Herberg and Severin Dressen, and three reviewers for comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Derix, J., Ortmann, S., Wiegmann, L. et al. Retention of solutes and particles in the gastrointestinal tract of a grazing cervid: Père David’s deer (Elaphurus davidianus). Eur J Wildl Res 65, 46 (2019). https://doi.org/10.1007/s10344-019-1288-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-019-1288-0