
Abstract
The study of nocturnal mammals relies on indirect evidence or invasive methods involving capture and tagging of individuals. Indirect methods are prone to error, while capture and tagging mammals have logistical and ethical considerations. Off-the-shelf camera traps are perceived as an accessible, non-intrusive method for direct data gathering, having many benefits but also potential biases. Here, using a 6-year camera-trap study of a Eurasian otter holt (den), we evaluate key parameters of study design. First, we analyse patterns of holt use in relation to researcher visits to maintain the camera traps. Then, using a dual camera-trap deployment, we compare the success of data capture from each camera-trap position in relation to the dual setup. Finally, we provide analyses to optimise minimum survey effort and camera-trap programming. Our findings indicate that otter presence and resting patterns were unaffected by the researcher visits. Results were significantly better using a close camera-trap emplacement than a distant. There was a higher frequency of otter activity at the holt during the natal and early rearing period which has implications for determining the minimum survey duration. Reducing video clip duration from 30 to 19 s would have included 95% of instances where sex could be identified, and saved 35–40% of memory storage. Peaks of otter activity were related to sunrise and sunset; exclusion of diurnal hours would have missed 11% of registrations. Camera-trap studies would benefit by adopting a similar framework of analyses in the preliminary stages or during a trial period to inform subsequent methodological refinements.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The study of terrestrial carnivores encompasses a great variety of direct and indirect monitoring methods, such as telemetry, capture-mark-recapture, distribution of field signs, harvest reports and questionnaire surveys (Gese 2001). The development of remote trail cameras, or camera traps, opened new avenues of study, and the twenty-first century marked a rapid proliferation in their use in ecological research (Rowcliffe and Carbone 2008). Camera-trap technology has been applied to biodiversity monitoring (Mugerwa et al. 2013; Tobler et al. 2015), estimating population size (Rowcliffe et al. 2008; Tobler et al. 2015) and behavioural observation (Brzeziński et al. 2014; Huang et al. 2014). There are, however, acknowledged potential sources of bias in camera-trap studies including disturbance, detectability, sampling design and trapping effort which may affect the use of camera traps as a research tool (Sollmann and Kelly 2013; Gužvica et al. 2014).
The ability of a camera trap to detect and record its target has been shown to be affected by the mass of the target, the distance between the camera trap and the target, the speed the target moves at and the season (Rowcliffe et al. 2011). Differences in detectability have also been found between camera-trap models (Swann et al. 2004; Wellington et al. 2014). Imperfect detection by a single camera trap has been improved by using two camera traps in different configurations, e.g. where camera traps are set at different distances from the target (Kilshaw and MacDonald 2011), adjacent to each other (Glen et al. 2013) or at 90° to the target (Newey et al. 2015). Most camera traps use passive infra-red (PIR) motion detectors which monitor ambient infra-red radiation and are triggered by changes due to infra-red radiation emitted by a passing animal. The otter’s adaptations to a semi-aquatic life such as fur structure and thermoregulation may reduce their infra-red footprint when exiting water, thus reducing their visibility to PIR suggesting that they may not be suitable for PIR motion detectors (Lerone et al. 2015). However, when the efficacy of camera traps was compared to scat surveys to quantify visitation rates by North American river otter Lontra canadensis to scat sites, the camera traps produced fewer false negatives than presence indicated by field signs (Day et al. 2016). In addition to investigating presence of otter at spraint sites, camera traps have been used to assist field sign interpretation in areas where several otter species co-exist but have similar spoor (Kanchanasaka 2001).
Camera trapping has been perceived as a non-intrusive “hands-off” method of direct observation (Rowcliffe et al. 2008; Adamič and Smole 2011). However, evidence is emerging that challenges this assumption: camera-trap shyness has been exhibited by tiger Panthera tigris (Wegge et al. 2004), and a startle reflex has been observed in the grey wolf Canis lupus (Gibeau and McTavish 2009). Behavioural responses to camera traps vary between species, and between individuals within species (Meek et al. 2016). Suggested sources of disturbance include the deposition of scent from ecologists undertaking maintenance visits (Munoz et al. 2014) and also noise (mechanical and ultrasonic) and infra-red illumination emitted by the camera traps (Meek et al. 2014). Potential bias from observation-effects should also therefore be considered in camera-trap studies.
Camera trapping is time-efficient in the field but analysis time can be onerous due to capture of non-target species and superfluous triggering caused by extraneous stimuli. In conservation and research, resources are inevitably limited; considering methodology efficiencies is important in terms of resources saved against any impact on data quantity or quality.
When deploying camera traps at den sites, key considerations therefore include (A) potential bias from disturbance, (B) the optimal number and placement of camera traps, (C) study duration and (D) the optimal camera-trap settings (e.g. clip duration, hours of operation/duty time). This study presents an empirical approach to address these using a 6-year study of the holt of a semi-aquatic mammal, Eurasian otter Lutra lutra.
Firstly (A), we investigate any effect on otter activity levels caused by regular visits by the researchers to maintain the camera traps. We hypothesise that if researcher visits caused disturbance to otters using the holt, a positive relationship between frequency of resting or scent-marking behaviours and number of days elapsed since the maintenance visit would be expected.
Secondly (B), we investigate how the position of a camera trap in relation to the recording area can affect the amount and type of data recorded. We hypothesise that data gain would improve using dual camera traps compared with one camera trap and that camera-trap position relative to the holt would affect both the probability of capturing an event and also the ability to record more specific observations such as sex and behaviour.
Thirdly (C), we investigate optimisation of study duration by quantifying the minimum number of days camera traps would need to be employed to observe specific activity types which would contribute to defining the minimum survey duration (MSD).
Finally (D), we examine whether camera-trap settings could be informed by an analysis of optimal video clip duration, and of duty time (time during the daily cycle when camera traps are armed) in relation to parameters which would be commonly recorded.
Our approach provides a framework whereby camera-trap studies in other locations or for other species could be optimised from the analysis of pilot study data. While we do not aim to define specific deployment and programming criteria for otter holt camera-trap studies, our results provide a baseline against which other sites can be compared.
Methods
Study species and context
The Eurasian otter (hereafter “otter”) is on Annex IV of the Habitats Directive (Council Directive 92/43/ECC) which affords it strict protection. Article 12 of the Directive frames protection in terms of the species’ wider habitat and also in relation to a species’ breeding and resting sites. With wide-ranging species such as otter, the actual place of rest is considered protected (EU 2007), and the Directive states that such sites must be “clearly perceptible” or “perfectly known and identified as such” (European Commission 1992). It is therefore important that breeding and resting sites can reliably be identified for the purpose of Environmental Impact Assessment and derogation licencing. Camera traps have been used to confirm the use of structures as dens for other species such as the Asiatic black bear Ursus thibetanus gedrosianus (Fahimi et al. 2011) as well as examining circadian activity of neotropical otter Lontra longicaudis at holts (Rheingantz et al. 2016). For species with unpredictable denning and breeding habits such as otter, camera trapping offers an accessible monitoring method to compliment traditional field-evidence surveys.
Study holt
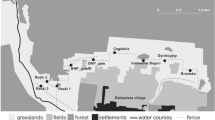
The study holt is in southern Scotland at a latitude of 56° 6′ 26″ N and is at an altitude of 125 m AMSL. The holt is adjacent to a small watercourse 3–4 m across in a secluded and undisturbed valley with near-continuous cover from scrub and thickets of bracken Pteridium aquilinum. It is 600 m downstream from a eutrophic loch, formerly a mixed fishery which is still fished informally and used for dog walking. Therefore, the holt and its immediate environs are undisturbed, but recreational disturbance is present around prey-rich habitat relatively close by. The holt is approximately 20 m from water and did not flood during the study period. The structure is a partially blocked drift mine with a tunnel (cross-section approximately 34 cm high and 140 cm wide) in rock which narrows and divides into two smaller tunnels after approximately 6 m.
Sampling period and summary of holt use
The study was undertaken between December 2009 and September 2015. Of the 2120 potential camera-trap days, cameras were operative on 1720 (81%). A large gap in recording in 2011 was due to stolen camera traps; to avoid further loss of equipment, monitoring ceased for approximately 6 months which also gave time to install more secure housing. Other gaps were due to battery depletion and delays in procuring replacement of defunct units. Maintenance visits during periods of continuous monitoring were on average every 15.2 (± SD 6.6) days, with approximately 15 min at the study site per visit. The same two researchers shared the maintenance visits throughout the study period; usually, just one researcher attended at each visit according to availability. Researcher visits avoided peak activity times of dawn and dusk.
The holt was assigned one of a set of mutually exclusive functions (pre-natal, natal, early rearing, mid rearing, late rearing and non-breeding) according to the status and/or absence of a breeding female (Fig. 1). The natal period, before emergence of the cubs, was taken as the 10 weeks preceding the first record of small cubs (Durbin 1996; Kruuk 2006) provided that there had been near-daily activity of adult otter recorded for at least 8 weeks. The early rearing period was defined as 60 days following the first day of emergence, mid rearing as 60 days following the end of early rearing and late rearing as 60 days following the end of mid rearing. At the end of the late rearing period, the cubs would be at least 8 months old, difficult to distinguish from the adult female and approaching dispersal (Jenkins 1980; Kruuk 2006). The pre-natal period was defined as 30 days before the estimated birth date, which is approximately the second half of the 63-day gestation period (Roos et al. 2015). Atypical activity was recorded during the single pre-natal period recorded (Nov–Dec 2011) when a female, a sub-adult male thought to be her cub from the last litter, and an adult male frequently rested in the holt as single otters, dyads or triads. Non-breeding was defined as none of the above. There was a minimum of two different breeding females during the study period: for the first two winters, the holt was used for birthing (natal), possibly by the same female, subsequently in the winters of 2011–2012, 2012–2013 and 2013–2014 a female with a distinctive broken/malformed tail used the holt for rearing but not birthing.

Timeline of holt function as defined by the status of the breeding female throughout study period (November 2009–September 2015) and times when camera traps were not recording. Holt function is defined in the text
Camera-trap deployment and setup
Over the study, two camera-trap positions were used, “close” and “distant” (Fig. 2). The close position was 1.6 m from the centre of the holt entrance at a height of 40 cm, and the distant was 4.2 m from the holt and 30 cm from the ground. No bait or lure was used at any time.

Camera-trap positions and approximate fields of view. a Camera traps were deployed in a modified plastic drain pipe (close camera) and wooden housing (distant camera) which emulated an old fence post for camouflage. The height and angle of the camera traps were consequently consistent each time they were reset. b Field-of-view of close camera trap. c Field-of-view of distant camera trap
The time-scale of the study resulted in two different camera-trap models being used which had different programming capabilities. Initially, a Moultrie I40 was deployed in the close position in December 2009. This model had an IR frequency < 850 nm and was programmed to record the maximum length of video possible (5 s) with the minimum programmable re-arm time between videos of 1 min. A second camera trap was added in November 2010 in the distant position to create the dual camera-trap deployment. The second camera trap was a Uway Night Trakker 50B (IR frequency 950 nm). This had better programming flexibility and so was set to record videos of 30 s with the minimum of 6 s to re-arm between videos. The close camera trap was replaced by a Uway in February 2011.
Camera traps were placed in unobtrusive locations dictated by the local topography and did not interfere with the otter runs. The holt was in a linear hollow which limited the extent of the detection and recording areas and also naturally contained otter activity.
Filtering videos and extraction of data
The date, time (GMT), number of otters and movement in or out of the holt were recorded, as was sex where possible. Sex was identified using primary characteristics (presence of scrotum, presence of nipples, source and direction of urine stream) and/or secondary characteristics (size and body shape). Selected behaviours including scent-marking (spraint and urine), vocalisation, play, grooming, loafing and bedding collection were recorded. Video clips from both cameras were cross-referenced using the date and time to compile a database of “events”. An event was defined as a unit of continuous activity, varying from the rapid pass of an otter, to an otter loafing for an extended period comprising numerous video clips. The event record contained the combined data gained from both camera traps.
Time spent in the holt was calculated where an otter was observed both entering and leaving, termed a “paired event”. On occasions, a group of otters would use the holt but entered and exited individually at different times. In such cases, the time in the holt of individual otters could not be tracked, and the minimum time spent in the holt was calculated from the last entry time to the first exit time (such occurrences accounted for 7% [36/492] of paired events). A bimodal frequency distribution of time spent in the holt indicated two natural categories of rest type: “visits” of 15 min or less (n = 305), or “rests” of greater than 15 min (n = 492). Thermal imaging has shown that wet otters can have a limited heat footprint due to their highly insulating fur (Kuhn and Meyer 2009). Single events of an otter exiting the holt at dusk with a dry coat without a corresponding record of it entering the holt were attributed to detection failure of the camera traps of a wet otter upon entry. These events were excluded from the analysis of time spent in the holt but were included as a rest in further analysis (17% of all rests) since it was assumed that to become dry the otter would have to have been in the holt for at least 15 min.
Analysis
Statistics were carried out in R version 3.2.2 (R Core Team 2015) within R Studio (RStudio Team 2016). Fitting of generalised linear mixed models used packages lme4 (Bates et al. 2014). A function to calculate sunrise and sunset was written using the packages rgeos (Bivand and Rundel 2016) and maptools (Bivand and Lewin-Koh 2016). We used the manipulate package (Allaire 2014) to fit the first apparent in frame models and the package lubridate (Grolemund and Wickham 2011) to facilitate use of dates and times.
Potential bias from disturbance
If otter activity was influenced by the researchers’ camera-trap maintenance visits, there would be a relationship between key otter behaviours such as resting and scent-marking and the number of days elapsed since a researcher visit. Additionally, it was hypothesised that propensity to disturbance might be influenced by the current function of the holt and that any disturbance would potentially be greatest during the natal and early rearing periods when cubs were small. Thus, generalised linear models (GLM) with binomial error distributions were constructed with the probability of rests (i.e. > 15 min in duration) occurring on any day as the binary response variable (1 = rest occurred, 0 = no rest occurred). The date of the rest was recorded as the date of entry to the holt. Models were tested for over dispersion.
Three explanatory variables were generated: (i) a binary variable indicating the holt function at the time of that rest as either “breeding” (pre-natal, natal, and early, mid and late rearing) or “non-breeding”, (ii) a binary variable indicating the holt function at the time of that rest as either “natal or early breeding” or “all other functions” (non-breeding, pre-natal, mid and late rearing) and (iii) a continuous variable indicating the number of days elapsed between the last researcher visit and the rest (the date of the rest was recorded as the date of entry to the holt). We then tested two models: one containing the interaction between (i) and (iii), and one containing the interaction between (ii) and (iii). For each model, if no interaction was found, the interaction was removed and the main effects were tested.
We used a likelihood ratio test with the Χ 2 distribution to compare models with and without the interaction term. If the test was not significant, we removed the interaction terms and tested the main effects within the non-interactive model.
Similarly, any relationships between the frequency of scent-marking at the holt and days elapsed since maintenance visit were tested for the response variable described whether scent-marking was detected on a particular day (1 = yes, 0 = no). Season was also included as a categorical explanatory variable (four levels: spring, summer, autumn, winter) as sprainting on land has been shown to be affected by season (Yoxon and Yoxon 2014). We tested whether the probability of scent-marking was related to an interaction between season and days elapsed since researcher visit. Again, we used a likelihood ratio test with the Χ 2 distribution to compare models with and without the interaction term, and then tested the main effects in the non-interactive model if the test was not significant.
The optimal number and placement of camera traps
The probabilities of data capture by each of the two individual camera traps were compared with the combined data gained from both camera traps. The dual camera-trap setup would always capture at least as much data as a scenario where only one of the camera traps was operational. Thus, we could examine the efficiency of each camera-trap position relative to each other and relative to the dual setup as a baseline, although not relative to perfect detection. This analysis can be conceptualised as the hypothetical removal of each camera in turn to retrospectively examine what the impact on our data would have been had we only had either the close or distant camera in place, thus comparing both cameras to the dual setup, and both cameras to each other. We examined the relative performance of both camera locations using three criteria: (i) count of otters, (ii) detection of sex of adult otter using primary characteristics (note that in the sample there were no events including more than one adult) and (iii) detection of selected behaviours (vocalisation, play, scent-marking, bedding collection, groom/rolling and loafing). Behaviour was recorded as the count of different behaviours observed; this was applied to both single otters and groups.
A random subsample of 200 events was selected when both Uway camera traps were in operation (i.e. post February 2011). These criteria required careful, multiple watching of footage, so from the large total of 2301 events, we randomly subsampled 200 (9%) events to provide a representative sample. Each event was given a categorical identifier. Microsoft Excel was used to generate random numbers, and events were selected using these numerical identifiers. Where the event was paired, the individual pass (in or out of the holt) was randomly selected by flipping a coin. The analysis included instances when one camera trap failed to trigger, or one camera trap triggered but did not record otter. For each pass of otter, the selected criteria (count, detection of sex using primary characteristics, behaviours) were recorded for each camera trap.
Generalised linear mixed models (GLMM) with binomial error distributions were constructed to investigate effects of camera-trap position and group size on the probability of capturing these three criteria. We hypothesised that the relative efficiency of each position could interact with group size, because a large group size may be a greater trigger stimulus than a single otter and therefore may increase detectability over longer distances. The categorical identifier was always included as a random effect to account for non-independence of the two camera-positions within each event. We used a likelihood ratio test with the Χ 2 distribution to compare models with and without the interaction term. If the test was not significant, we removed the interaction terms and tested a model just containing the main effects.
The first model used probability of detecting an otter as the response variable. The measure of success for each camera trap in detecting an otter was represented by a dual vector comprising the number of otters seen on the single camera trap (binomial numerator), and the number observed by the dual camera traps (binomial denominator).
A second GLMM was constructed using the ability to determine sex as the response variable (1 = sex identified, 0 = sex not identified). Finally, this was repeated using observation of behaviour as a response variable, represented by a dual vector of the numbers of behaviours observed on a single camera trap (binomial numerator) and the number of behaviours observed on the dual camera-trap system (binomial denominator). Again, an interaction between camera-trap position and number of otters on the dual system was tested for, and if this was not significant, the interaction term was removed to test the significance of the main effects within the non-interactive model.
Within the subsample of 200 random passes, redundancy of the two camera-traps positions in the dual camera setup was assessed for each pass by determining whether a particular data type was recorded by (a) both camera traps, (b) only the close camera trap or (c) only the distant camera trap. The higher the percentage of events that fall into (a), the more redundancy there is in the dual camera setup. The data types considered were (i) presence of otter(s) (yes/no), (ii) count of otters, (iii) observation of behaviour (yes/no) and (iv) determination of sex (yes/no). For (ii), we took the count as the minimum number of otters seen on the dual camera-trap setup.
Study duration
Given the status of otter as a European-protected species, there is a requirement to ascertain whether or not a structure is used for resting but there are no explicit criteria for identifying an otter resting site. Based on the distribution of duration of time spent at the holt, three hierarchical categories of otter use of the holt per study day were generated: absence of otter, any presence of otter (all registrations) and a rest by an otter (a stay within the holt of > 15 min). The last two categories broadly align with two potential aims of a camera-trap study at a holt, i.e. either to (a) simply determine presence of otter at a site or (b) to determine whether a site can be defined as a “resting site”. The number of days between a specified event type (i.e. presence or rest) would inform the minimum study duration required for that specific aim.
For each period of holt function (Fig. 1), the intervals (days) between consecutive instances of the same activity-type (presence or rest) were calculated. If the interval spanned more than one holt function, such as the last rest in the early rearing period of 2010 to the first rest in the early rearing period of 2011, then it was excluded.
A GLM with Poisson error distributions was constructed with the number of days between successive visits as the response variable and holt function as the explanatory variable. We repeated this using the number of days between otter rests as the response variable using a quasi-Poisson error distribution due to over dispersion. A likelihood ratio test with a Χ 2 distribution was used to assess model significance.
We calculated the 90th and 95th percentiles of intervals between events (separately for presence and rests) as a contributor to minimum survey duration which represents a 90–95% probability we would record one of each activity-type if our study was at least that long. Because holt function significantly influenced the intervals between events for both presence and rests, we calculated separate percentiles for each holt function (natal, early rearing, mid rearing, late rearing and non-breeding).
The optimal camera-trap settings: clip duration and duty time
Setting a camera trap to record longer video clips may increase data gain, but results in greater battery depletion and memory storage each time a camera triggers (often by non-target species or false-triggers), as well as longer time required to review the clips. Thus, selection of the duration of video clips represents a trade-off that ecologists have to make for each study. Reducing clip duration without losing significant data has the potential to increase efficiency of camera-trapping studies.
We specified a set of three observable criteria that ecologists are likely to record using camera-trap footage: (1) determining sex of an otter using primary sexual characteristics only or (2) both primary and secondary sexual characteristics (primary characteristics plus body shape), and (3) scent-marking activity (sprainting, urination). Using events recorded by the dual camera-trap setup with two Uway camera traps, we extracted all events containing the desired criteria (scent-marking n = 274, primary sexual characteristics n = 373, primary and secondary sexual characteristics n = 171). Some otters in this study had characteristic tail abnormalities which identified them as individuals. To avoid bias from individuals being recognised and influencing observations of sex, these were omitted from the subsampling for observation of primary sexual characteristics (reduction of n = 373 to 123). Excluding the period when the Moultrie camera trap was operating, we randomly selected and rewatched 60 events from each subset to provide a representative sample. For each event, the data from either the close or distant camera trap was randomly selected, as was the individual pass if it was a paired event (either going in, or coming out of the holt). For each pass, we observed a maximum of 30 s of video and recorded the time to the nearest second when each observable criteria was first apparent in frame (hereafter “FAF time”).
For each criteria, the FAF times were ranked in ascending order. The rank of each data point was then divided by the sample size for that criteria to form a cumulative proportion. The cumulative proportion (y-axis) was plotted against the FAF (x-axis) for each pass. To describe the asymptotic relationship that was apparent for each criteria, we fitted an exponential model of the form y = a.(1-e-b.x) + c where y is the predicted cumulative proportion of that observable criteria that would have been recorded given a hypothetical clip duration (s) of x, and a, b and c are parameters estimated by the model. There is a short delay between a subject triggering a PIR detector and the camera trap initiating recording. A recorded FAF time of zero can actually represent a range of true FAF times within that delay range. As such, the plotted cumulative distributions appeared truncated at t = 0. To avoid truncation influencing model fit, values of FAF = 0 were excluded from the model.
The fitted models were used to predict the minimum clip duration that would be required to record 95% of passes containing each observable criteria since we determined that 5% data loss would be acceptable if it could result in a proportionally greater reduction in superfluous video, battery depletion or memory depletion.
Many camera-trap models have the ability for duty time to be programmed, i.e. daily periods of time when the camera trap is active or inactive. They are prone to being triggered by precipitation, strong light conditions or vegetation moving in the wind (Swann et al. 2004), termed “false-triggers”. Runs of false-triggers were experienced almost exclusively during the daytime depleting the limited supply of memory storage capacity and increasing analysis time. Efficiency in analysis time would be improved if false-triggers could be substantially reduced by the camera trap being in “sleep-mode” during some, or all of the day, if it could be demonstrated that this would not lead to a significant loss of data. In describing mammal activity in relation to sunrise and sunset, four activity periods have been described (Lucherini et al. 2009): (a) day, (b) night, (c) dawn (1 h before sunrise to 1 h after) and (d) dusk (1 h before sunset to 1 h after). Otters are predominantly nocturnal (Green et al. 1984) but they can be active during the day. For each otter registration, the times of the closest sunset and sunrise were back-calculated using the date and time in conjunction with the holt’s latitude and longitude. The time of each registration was then compared to the time of the closest sunrise and sunset and assigned to whichever one it was closest to. We then plotted the distribution of hours relative to sunrise and sunset for each registration and calculated the proportion of registrations that occurred in the four activity periods (day, night, dawn, dusk). This was repeated for registrations within each holt function category.
Results
Potential bias from disturbance
The GLM which tested for any effect on the probability of a rest during periods when the holt function was breeding or non-breeding and days since maintenance found no significant interaction (Χ 2 = 1.16, df = 1, p = 0.281). When the interaction term was removed, there was no significant effect of holt function and days since maintenance check on the probability of a rest (Χ 2 = 0.859, df = 1, p = 0.354). Similarly, the GLM which defined the holt function as early breeding (natal and early rearing periods) or not early breeding found no significant interaction between holt function and days elapsed since maintenance visit on the probability of a rest (Χ 2 = 0.65, df = 1, p = 0.418). When the interaction term was removed, there was no significant effect of holt function and days since maintenance check on the probability of a rest (Χ 2 = 0.22, df = 1, p = 0.637).
The GLM using the probability of scent-marking as the response variable found no significant interaction between the season and days elapsed since maintenance check (Χ 2 = 6.84, df = 3 p = 0.077). When the interaction term was removed, the probability of scent-marking on a given day was not significantly related to days elapsed since maintenance check (Χ 2 = 0.57, df = 1, p = 0.520).
The optimal number and placement of camera traps
The GLMM investigating effects on the probability of detecting an otter found no significant interaction between camera-trap position and group size (Χ 2 = 0.04, df = 1, p = 0.852). When the interaction term was removed, the probability of detecting an otter was significantly related to camera-trap position and group size (Χ 2 = 25.86, df = 1, p < 0.001) (Fig. 3a). When investigating the effects on the probability of detecting the sex of an adult otter, no significant interaction was found between camera-trap position and group size (Χ 2 = 1.80, df = 1, p = 0.179). Removal of the interaction term resulted in a significant effect of camera-trap position and group size (Χ 2 = 21.96, df = 1, p < 0.001) (Fig. 3b). The GLMM investigating effects on the probability of observing behaviour found no interaction between group size and camera-trap position (Χ 2 = 0.52, df = 1, p = 0.469), and when the interaction term was removed, there was no significant difference from the camera-trap position and group size (Χ 2 = 0.04, df = 1, p = 0.842); however, there was a significant effect of camera-trap position (Χ 2 = 28.07, df = 1, p < 0.001). The probability of the close camera trap recording behaviours was 0.81 (± 0.04 SE) substantially greater than for the distant camera trap (0.47 [± 0.08 SE]).

The probability of data capture was different between the two camera-trap positions when considering a the probability of detecting an otter and b the ability to sex the adult otter
The close position substantially out-performed the distant camera trap both in terms of registering presence, count of otters and facilitating the identification of otter sex (Fig. 4) and also recording behaviour. There was the highest degree of redundancy between cameras when recording behaviours, with 91% of behaviours being recorded by both cameras. However, there was substantially less redundancy between cameras for presence (57%), count (48%) and sexing (52%).

Comparison of the uniqueness of data capture between the two camera-trap positions in a random sample of 200 events. Pale grey indicates the proportion of events where only the close camera trap recorded data in each category which was unique, and black indicates the proportion of events where only the distant camera trap recorded data in each category. The hatched area represents the proportion of events where both camera traps recorded the same data in each category
Study duration
There was a significant effect of holt function on days between consecutive records of otter presence (Χ 2 = 195.35, df = 5, p < 0.001). There was also a significant effect of holt function on days between consecutive records of otter rest (Χ 2 = 158.47, df = 5, p < 0.001).
The number of days between consecutive records of otter presence at the holt increases with decreasing breeding status relative to the natal period (Fig. 5); this is more pronounced with resting patterns than presence.

95 and 90% percentiles of intervals in days between consecutive rest types for each holt function excluding pre-natal as sample was too small; a for presence of otter at holt and b for a rest of over 15 min.
Optimal camera-trap settings
Clip duration
The 95th percentile for sexing otters using primary characteristics only was 22 s, for sexing otters using a combination of primary and secondary characteristics was 19 s and for recording scent-marking behaviour was 24 s (Fig. 6).

Minimum clip durations illustrating 95th percentile for three types of observation: a for sexing otters using primary characteristics only, b for sexing otters using a combination of primary and secondary characteristics and c for recording scent-marking behaviour (solid line: fitted model, dotted lines: standard errors of the relationship, dashed lines: 95th percentile readings)
Duty time
Frequency of registrations peaked approximately 2 h before sunrise and 2 h after sunset (Fig. 7). Nocturnal activity accounted for 81% (n = 2301) of all registrations. Inclusion of dawn and dusk periods increases the proportion of registrations to 89%. However, when the holt was functioning as a natal holt, 86% of registrations were nocturnal and 100% of registrations occurred in the nocturnal and dawn and dusk periods, i.e. there was no diurnal activity. There was a slight increase in diurnal activity during the early rearing period (nocturnal 86%, nocturnal, dawn and dusk 93%). Diurnal activity increased during late rearing and non-breeding status too (late rearing nocturnal 64%; nocturnal, dawn and dusk 81%; non-breeding nocturnal 78%; nocturnal, dawn and dusk 87%).

Histograms of otter registrations at the holt in relation to hours around a sunrise and b sunset. Each registration is included within a single histogram depending on whether it was closer to a sunrise or b sunset
Discussion
Before interpreting the results from any camera-trap study, potential sources of observer bias must be considered. There are two primary potential causes of observer bias in our study: (i) regular maintenance visits and (ii) any effect from the camera traps themselves. We did not find any effect of maintenance visit on the probability of resting or scent-marking at the study site and this was unaffected by the breeding status of the holt. The maintenance visits at the study site were, on average, 2 weeks apart and did not include scent masking, so the deposition of human scent at this interval does not appear to have affected otters’ use of the holt. There may be a threshold of shorter intervals between maintenance visits which would cause disturbance and affect patterns of activity, and future studies might be able to quantify this. Over such a long-term study, there may have been habituation to the visits which were by the same researchers throughout the study period. Additionally, the sleeping chambers of the holt are at least 6 m from the entrance so disturbance at the entrance to the tunnel may not be critical given the size and security of the structure. As such, further investigation using different types of structure in areas of higher/lower ambient disturbance levels (e.g. urban vs. rural sites) may be required to assess impacts of disturbance and habituation. Behavioural reactions to the camera traps were not quantified in this study, although no adverse reactions to the close camera trap were observed on the distant camera trap. Studies on predatory species found that animals could readily detect camera traps (Meek et al. 2014) with some nocturnal predators, such as felid species being particularly sensitive. Individuals of some species have been observed exhibiting adverse reactions such as backing away (Meek et al. 2016), and this could potentially affect detectability. However, neotropical otters continued to use holts after camera traps were deployed facing the holts, and this was observed in both areas of the study (Rheingantz et al. 2016), also giant otter Pteronura brasiliensis were almost indifferent to camera traps placed at the edge of latrine sites (Pickles et al. 2011) suggesting that this otter species may not be sensitive to camera traps. For these reasons, the unaffected activity patterns may not necessarily be applicable at other sites, but the lack of any change in activity indicates that observer effect need not be considered in our subsequent analyses.
Very poor detectability of otter by PIR triggered camera traps led researchers to question whether such camera traps are appropriate for semi-aquatic species and whether active triggers would be more effective (Lerone et al. 2015). Our study illustrates that PIR camera traps can successfully be used to study otter (see also Rheingantz et al. 2016), but differences in deployment can cause variation in detectability. In the comparison of data capture between the dual setup and individual camera traps, the close position provided the most information, both in terms of detecting otter and the ability to identify sex. With perfect detection, we would have observed one otter entering the holt for every otter exiting the holt (i.e. every event would be paired), but this only occurred in 61% (1610 of 2639) of events where holt entry/emergence occurred. Single events were thought to be due to missed registrations either when the otter did not trigger the PIR, where the PIR was triggered but the otter was not recorded possibly if the otter was moving quickly, or if an otter passed during the time when the camera trap re-armed between videos. The high proportion of missing passes and the poorer detectability of the distant camera trap are notable, although probability of detection has previously been shown to be affected by distance (Rowcliffe et al. 2011; Howe et al. 2017). A greater source of bias would have been experienced if only the distant camera had been used; this large discrepancy suggests a cause for concern when management/derogation licencing decisions are made based on camera-trap monitoring.
Setting the distance between the camera trap and the holt is a compromise. Increased distance gives a better overview of the den area and has a perceived, though not evidenced, potential reduction in disturbance, but has a negative effect on detection probability. The sensitivity of the target species to disturbance coupled with the individual characteristics of the den structure and the species’ effects on detection therefore all need to be balanced and understood when setting camera traps at den sites. Detection improved when family groups used the holt which indicates distance to the target may be more critical for solitary species than species living in a social group. A group of otters will present as a larger stimulus for PIR. This may have been a contributing factor in the success of other camera-trap studies of otter species which live in groups (Pickles et al. 2011; Day et al. 2016; Rheingantz et al. 2016) and the poor PIR detection reported for the Eurasian otter (Lerone et al. 2015) which is often solitary.
If a close camera trap is deployed, the addition of a second camera trap should offer significant data gain or have other tangible benefits to justify the capital cost and substantial increase in analysis time. In this case study, the distant camera trap offered little extra gain of data (Fig. 4), and its loss would have been acceptable in light of this and also its limitations as a back-up if the close camera trap failed. However, a second camera trap placed on the other side of the holt may have reduced the amount of missing passes.
False-triggers are a drain on power and memory storage. Duty time can be set on many models; a dormant camera trap during the daytime for a nocturnal species would likely increase the longevity of a camera trap in the field and reduce the likelihood of battery or memory depletion before maintenance visits. Otter activity at the holt was concentrated between 1 h before dusk and 1 h after sunrise, with 89% of all registrations occurring in this period and 100% of registrations when the holt was in the natal or early rearing phase. This conforms with studies on activity from radio telemetry where emergence occurred just after sunset and retirement was related to sunrise, but some diurnal activity was also recorded (Green et al. 1984). If duty times were set so that the camera trap recorded from 1 h before sunset and finished at 1 h after sunrise, the loss of data (11%) in this study would have been considered acceptable in context with the considerable time it took to filter daytime footage and compile the event database, and likely would have reduced instances of battery or memory depletion. However, it has been suggested that resource partitioning may occur in areas of high density with single otters foraging in areas during the daytime and families of otter using the same area during the night (Jenkins 1980). It has also been demonstrated that the circadian activity of neotropical otter varies between regions (Rheingantz et al. 2016). Caution is therefore needed, before restricting the duty time of camera traps even for perceived nocturnal species without knowledge of the study population, and our approach could be used on a set of pilot data before setting any restrictions on recording.
The frequency of resting at the holt was significantly related to the holt function. To determine the current function of the holt, the minimum study duration should consider the number of days between desired events, such as rests, with an additional period of habituation likely to be determined by the type of structure. This study of a very busy and secure holt in rich habitat indicates a minimum of 28 days to have a 95% probability of recording at least one rest regardless of holt function, which would be unlikely to be known when initiating a study. A period of habituation also needs to be factored in. A minimum of 28 days would have been long enough to detect a more significant function such as cub-rearing or birthing (natal) if the holt currently had that function. If the aim is to determine breeding, the monitoring should coincide with any known local breeding season, although this varies considerably across the species’ geographic range: summer on Skye in Scotland (Yoxon and Yoxon 1990), spring in southern Sweden (Erlinge 1967), locality-specific seasons in the Mediterranean (Ruiz-Olmo et al. 2002) and aseasonal in England and Wales (Chadwick and Sherrard-Smith 2010). A female is unlikely to give birth more frequently than once per year giving a window of opportunity for recording natal behaviour of 9–10 weeks out of 52. Where no breeding season is known, sampling through the year would be required if determining the status of a structure is a requirement for Environmental Impact Assessment or other assessments; however, fidelity to natal holts is not guaranteed leaving some residual uncertainty.
Long video clips will fill up memory space, drain batteries and increase analysis time. In the analyses for optimising clip duration, a survey simply for presence of otter would require the shortest clip duration, and it could be argued that still images would be more appropriate. To gain additional data such as identification of sex and recording scent-marking behaviour, a balance needs to be found between analysis time and data gain. If video clips had been reduced to 19 s (the 95th percentile of the FAF analysis) from 30, to facilitate sexing of otter using both primary and secondary characteristics, then this would proportionately have reduced memory storage by 35–40% (11/30 s) and reduced power consumption, which would have the benefit of extending the number of days that the camera trap could run untended. It would also have reduced video analysis time and so, on balance, the loss of the 5% of instances where the sex can be determined against the reduction of analysis time and greater field longevity of the camera trap would have been an acceptable trade-off. The FAF approach could therefore be applied to optimise settings for specific data collection; a trial period would enable the most efficient video duration to be estimated.
Recommendations
The study holt was a well-used otter breeding structure in rich habitat, and so there are limitations to the generality of the findings across all possible otter structures that practitioners may monitor. However, these results do present some general considerations for camera-trapping otters and other species of semi-aquatic and terrestrial mammals, as well as presenting a framework whereby efficiency and efficacy of camera trapping can be investigated and improved via the analyses of prior data collected. At den sites, we recommend analyses to assess any observer effect. If more than one camera trap is used on the same target area, the effect of distance on detectability should be considered, which could result in two close camera traps. However, data redundancy should be evaluated and a high level of redundancy may indicate that one camera trap could be removed or could be run as a back-up. Any local variation in activity should be taken into account when determining duty time and minimum survey duration; factors such as breeding status should also be considered. An adaptive approach, whereby data is evaluated in the early stages of a study and appropriate modifications made to study design, could improve both data quality and use of resources.
There are many potential biases within camera-trapping studies, and further research is required to understand how environmental, spatial and animal-based factors interact to influence the detection probability of animals to camera traps. These may vary between taxa and functional groups (e.g. semi-aquatic versus terrestrial mammal species), between solitary and social species and between habitats and environments, and so a one-size-fits-all approach is unlikely to be appropriate.
References
Adamič MH, Smole J (2011) Phototraps as a non-invasive method of monitoring otters (Lutra lutra): what can we expect? IUCN Otter Spec Gr Bull 28(A):60–69
Allaire JJ (2014) Manipulate: interactive plots for RStudio. R package version 1.0.1. http://CRAN.R-project.org/package=manipulate
Bates DM, Machler M, Bolker BM, Walker SC (2014) Fitting linear mixed-effects models using lme4. J Stat Softw 67:51
Bivand R, & Lewin-Koh N (2016) Maptools: tools for reading and 845 handling spatial objects. http://CRAN.Rproject.org/package=maptools
Bivand R, Rundel C (2016) Rgeos: interface to Geometry Engine - Open Source (GEOS). R package version 0.3-21. http://CRAN.R-project.org/package=rgeos
Brzeziński M, Rodak Ł, Zalewski A (2014) “Reversed” intraguild predation: red fox cubs killed by pine marten. Acta Theriol (Warsz) 59:473–477
Chadwick EA, Sherrard-Smith E (2010) Pregnancy among otters (Lutra lutra) found dead in England and Wales: foetal development and lack of seasonality. IUCN Otter Spec Gr Bull 27:33–41
Day CC, Westover MD, Hall LK, Larsen RT, Mcmillan BR (2016) Comparing direct and indirect methods to estimate detection rates and site use of a cryptic semi-aquatic carnivore. Ecol Indic 66:230–234
Durbin LS (1996) Some changes in the habitat use of a free-ranging female otter Lutra lutra during breeding. J Zool 240:761–764
Erlinge S (1967) Home range of the otter Lutra lutra L. in southern Sweden. Oikos 18:186–209
EU (2007) Guidance document on the strict protection of animal species of community interest under the habitats directive 92/43/EEC
European Commission (1992) Council directive 92/43/ECC. Off J Eur Union 94:40–52
Fahimi H, Yusefi GH, Madjdzadeh SM, Damangir AA, Sehhatisabet ME, Khalatbari L (2011) Camera traps reveal use of caves by Asiatic black bears (Ursus thibetanus gedrosianus) (Mammalia: Ursidae) in southeastern Iran. J Nat Hist 45:2363–2373
Gese EM (2001) Monitoring of terrestrial carnivore populations. In: Gittleman J, Funk S, Macdonald D, Wayne R (eds) Carnivore conservation. Cambridge University Press, Ithaca, New York, pp 372–396
Gibeau ML, McTavish C (2009) How to prevent camera traps from skewing animal behavior. Wildl Prof 3:35–37
Glen AS, Cockburn S, Nichols M, Ekanayake J, Warburton B (2013) Optimising camera traps for monitoring small mammals. PLoS One 8:1–7
Green J, Green R, Jefferies D (1984) A radio-tracking survey of otters Lutra lutra on a Perthsire river system. Lutra 27:85–145
Grolemund G, Wickham H (2011) Dates and times made easy with lubridate. J Stat Softw 40:1–25
Gužvica G, Bošnjak I, Bielen A, Babić D, Radanović-Gužvica B, Sver L (2014) Comparative analysis of three different methods for monitoring the use of green bridges by wildlife. PLoS One 9:1–12
Howe EJ, Buckland ST, Després-Einspenner M-L, Kühl HS (2017) Distance sampling with camera traps. Methods Ecol Evol 8:1558–1565
Huang ZP, Qi XG, Garber PA, Jin T, Guo ST, Li S, Li BG (2014) The use of camera traps to identify the set of scavengers preying on the carcass of a golden snub-nosed monkey (Rhinopithecus roxellana). PLoS One 9:1–6
Jenkins D (1980) Ecology of otters in northern Scotland: I. Otter (Lutra lutra) breeding and dispersion in mid-Deeside, Aberdeenshire in 1974-79. J Anim Ecol 49:713–735
Kanchanasaka B (2001) Tracks and other signs of the hairy-nosed otter (Lutra sumatrana). IUCN Otter Spec Gr Bull 18:2–7
Kilshaw K, MacDonald DW (2011). The use of camera trapping as a method to survey for the Scottish wildcat. Scottish Natural Heritage Commissioned Report 479:1–32
Kruuk H (2006) Otters: ecology, behaviour and conservation. Oxford University Press, Oxford
Kuhn R, Meyer W (2009) Infrared thermography of the body surface in the Eurasian otter Lutra lutra and the giant otter Pteronura brasiliensis. Aquat Biol 6:143–152
Lerone L, Carpaneto GM, Loy A (2015) Why camera traps fail to detect a semi-aquatic mammal: activation devices as possible cause. Wildl Soc Bull 39:193–196
Lucherini M, Reppucci JIJ, Walker RS, Villalba ML, Wurstten A, Gallardo G, Iriarte A, Villalobos R, Perovic P (2009) Activity patterns and segregation of carnivores in the High Andes. J Mammal 90:1404–1409
Meek P, Ballard G, Fleming P, Falzon G (2016) Are we getting the full picture? Animal responses to camera traps and implications for predator studies. Ecol Evol 6:3216–3225
Meek PD, Ballard GA, Fleming PJS, Schaefer M, Williams W, Falzon G (2014) Camera traps can be heard and seen by animals. PLoS One 9:1–16
Mugerwa B, Sheil D, Ssekiranda P, Van Heist M, Ezuma P (2013) A camera trap assessment of terrestrial vertebrates in Bwindi Impenetrable National Park, Uganda. Afr J Ecol 51:21–31
Munoz D, Kapfer J, Olfenbuttel C (2014) Do available products to mask human scent influence camera trap survey results? Wildlife Biol 20:246–252
Newey S, Davidson P, Nazir S, Fairhurst G, Verdicchio F, Irvine RJ, van der Wal R (2015) Limitations of recreational camera traps for wildlife management and conservation research: a practitioner’s perspective. Ambio 44:624–635
Pickles R, Zambrana V, Hoffmann-Heap I, Salinas A, Groombridge J, van Damme P (2011) An evaluation of the utility of camera traps in monitoring giant otter populations. IUCN Otter Spec Gr Bull 28:39–45
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Rheingantz ML, Leuchtenberger C, Zucco CA, Fernandez FAS (2016) Differences in activity patterns of the Neotropical otter Lontra longicaudis between rivers of two Brazilian ecoregions. J Trop Ecol 32:170–174
Roos, A., Loy, A., De Silva, P., Hajkova, P. & Zemanová, B. (2015). Lutra lutra. The IUCN Red List of Threatened Species 2015: e.T12419A21935287. [WWW Document]. URL Doi: https://doi.org/10.2305/IUCN.UK.2015-2.RLTS.T12419A21935287.en
Rowcliffe JM, Carbone C (2008) Surveys using camera traps: are we looking to a brighter future? Anim Conserv 11:185–186
Rowcliffe MJ, Carbone C, Jansen PA, Kays R, Kranstauber B (2011) Quantifying the sensitivity of camera traps: an adapted distance sampling approach. Methods Ecol Evol 2:464–476
Rowcliffe JM, Field J, Turvey ST, Carbone C (2008) Estimating animal density using camera traps without the need for individual recognition. J Appl Ecol 45:1228–1236
RStudio Team (2016) RStudio: integrated development for R. RStudio, Inc., Boston. http://www.rstudio.com/
Ruiz-Olmo J, Olmo-vidal JM, Manas S, Batet A (2002) The influence of resource seasonality on the breeding patterns of the Eurasian otter (Lutra lutra) in Mediterranean habitats. Can J Zool 80:2178–2189
Sollmann R, Kelly MJ (2013) Camera trapping for the study and conservation of tropical carnivores. Raffles Bull Zool 28:21–42
Swann DE, Hass CC, Dalton DC, Wolf SA (2004) Infrared-triggered cameras for detecting wildlife: an evaluation and review. Wildl Soc Bull 32:357–365
Tobler MW, Zúñiga Hartley A, Carrillo-Percastegui SE, Powell GVN (2015) Spatiotemporal hierarchical modelling of species richness and occupancy using camera trap data. J Appl Ecol 52:413–421
Wegge P, Pokheral CP, Jnawali SR (2004) Effects of trapping effort and trap shyness on estimates of tiger abundance from camera trap studies. Anim Conserv 7:251–256
Wellington K, Bottom C, Merrill C, Litvaitis JA (2014) Identifying performance differences among trail cameras used to monitor forest mammals. Wildl Soc Bull 38:634–638
Yoxon G, Yoxon P (1990) Otter survey of the Isle of Skye, Scotland (Lutra lutra). IUCN Otter Spec Gr Bull 5:70–75
Yoxon P, Yoxon K (2014) Estimating otter numbers using spraints: is it possible? J Mar Biol 2014:1–6
Acknowledgements
Many thanks to Roger Ingledew who shared the task of camera-trap maintenance throughout the study period.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Findlay, M.A., Briers, R.A., Diamond, N. et al. Developing an empirical approach to optimal camera-trap deployment at mammal resting sites: evidence from a longitudinal study of an otter Lutra lutra holt. Eur J Wildl Res 63, 96 (2017). https://doi.org/10.1007/s10344-017-1143-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-017-1143-0