
Abstract
Italy is amongst the European countries for which data on badger Meles meles numbers and variation in population size are still too few to assess its conservation status. With the aim of estimating badger density in northern Italy, between February 2013 and March 2014, we extensively surveyed an isolated hilly area for badger setts and assessed group size by monitoring a sample of setts by two sessions of camera-trapping. The size and shape of each home range were assessed from main sett coordinates using the Dirichlet tessellation method. Then, assuming that badger home ranges tend to be delimited by hard boundaries or geographical features, the polygon shapes were successively modified by hand to draw more realistic home ranges. The overall trapping effort was 754 camera trap-days. The number of badger individuals sharing the same sett/entrances was recorded, distinguishing individuals based on the time and hole of emergence and, possibly, tail patterns. Groups consisted of two to four adult individuals, averaging 2.75 badgers/group. Camera-trapping proved to be a cost-effective and time-saving method. Depending on the method used to draw home ranges, in the study area, sett density was 0.34–0.51 sett/km2, whilst badger density was 0.93–1.4 adult individuals/km2. Both sett and badger densities were rather high with respect to most available data for continental Europe. As badger mean group size seems to be rather constant throughout both hilly and plain areas of the River Po basin, sett density may be used as an accurate estimator of badger density throughout this wide area.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The assessment of population density is crucial for planning conservation measures, evaluating the effectiveness of management strategies and assessing the impact of hunting and economic activities on species demography (Nichols and Williams 2006; Caro et al. 2009; Campbell et al. 2011).
In the case of the European badger (Meles meles), a widely distributed (Marmi et al. 2006) and relatively common species throughout Europe (Kranz et al. 2008), the estimation of density has both a management interest, due to its role in the transmission of bovine tuberculosis in the UK and Ireland (Garnett et al. 2005), and a theoretical interest, due to the relation between population density and social organisation.
Early research on badger social and territorial behaviour led to apparently opposite results, with badgers living in large social groups in England (Kruuk 1978) and almost solitarily in central Italy (Pigozzi 1987). Although the evidence supporting solitary territoriality has been heavily criticised (Revilla and Palomares 2002; Roper 2010), these findings, together with growing evidence that badgers do not cooperate for either hunting or breeding (Kruuk 1978; Woodroffe and Macdonald 1999), stimulated several attempts to explain its social organisation and yielded some general hypotheses about group formation and the origin of territoriality (Woodroffe and Macdonald 1993).
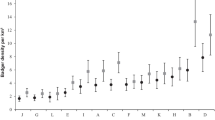
Available information on badger density throughout Europe has been revised by Kowalczyk et al. (2000), Johnson et al. (2002) and Roper (2010). All studies agreed that badger numbers were significantly larger in north-western Europe (up to 25 adults/km2 in the British Isles; Cheeseman et al. 1987; Rogers et al. 1997) than in the rest of the wide badger range (on average, 1.24 badgers/km2; Table 1).
The accuracy of density estimates depends on several factors, including the method used for number assessment. Moreover, for social species, to obtain an estimate of population density, the assessment of territory size must be coupled with that of social group size.
Whilst in the UK, badger populations have been the object of several long-term capture-mark-recapture studies, in continental Europe, group size has been mostly determined by sett-watching at dusk and/or live trapping (e.g. Goszczynski 1999; Rosalino et al. 2004; Acevedo et al. 2014), whilst radio telemetry (e.g. Rodríguez et al. 1996; Do Linh San et al. 2007a) and bait-marking (Delahay et al. 2000) have been the methods of choice to assess home range size. These approaches have some limitations; direct observation tends to underestimate the number of animals per group (Tuyttens et al. 2001; Scheppers et al. 2007), whilst home range size depends on both the estimator used (White and Garrott 1990) and time interval between successive fixes (De Solla et al. 1999), making it difficult to compare the results from different studies. Sample size is a further problem, as most available studies were based on a few individuals and groups due to the labour intensiveness of trapping and fix recording.
To our knowledge, published badger density data were not available for Italy. Some information is available for sett density, which has been reported to be ca. 0.2 setts/km2 for plain areas of northern Italy (Quadrelli 1993; Remonti et al. 2006a), whilst higher values have been reported for central Italy (0.7 setts/km2; Tinelli and Tinelli 1980) and both hilly (1.32 setts/km2; Remonti et al. 2006a) and Alpine (7.1 setts/km2; Prigioni and Deflorian 2005) areas. Group size is poorly known, although hair-trapping (Balestrieri et al. 2010), sett clearing (Balestrieri et al. 2006), radio-tracking (Remonti et al. 2006b) and camera-trapping (Mori et al. 2014) suggested the occurrence of small groups including one adult male and from one to three adult females.
With the aim of estimating badger population density, we extensively surveyed an isolated hilly area of northern Italy for badger setts and assessed group size by monitoring a sample of setts by camera-trapping. The size and shape of home ranges were estimated by the Dirichlet tessellation method (Doncaster and Woodroffe 1993), which is based on the assumption that setts are a substantial and limiting resource that rules badger spatial organisation. Although the suitability of the method has been criticised (Blackwell and Macdonald 2000; Doncaster 2001), it can be used to estimate territory size and shape from the location of main setts quite accurately (e.g. Macdonald et al. 2004).
Study area
The study area consisted of an isolated, 14.5-km2 wide, hill (up to 147 m above sea level (asl)) in the middle River Po plain (Lombardy region, ca. 45° 11′ N, 9° 29′ E). It consisted of summital Miocene marls, covered by clays and limestones from the Pliocene and Quaternary alluvial sediments, and was bordered to the north-east by the River Lambro and to the south by the River Po. Woodland (ca. 24 %) consisted of oaks (Quercus robur and Q. petraea), alder (Alnus glutinosa), chestnut (Castanea sativa) and black locust (Robinia pesudoacacia), with shrubs of Cornus sanguinea, Corlylus avellana and Crateaegus monogyna. Permanent cover also included grasslands, fallow fields and shrublands (14 %). Cropland (55 %), mostly vineyards (39 %), was mainly located in the south-eastern part of the hill. The surrounding matrix consisted of an intensively cultivated agricultural area, being mostly maize and rice fields and small urban areas. The climate was subcontinental temperate, with an average yearly temperature of 11.4 °C and an average yearly precipitation of 712 mm.
Methods
Badger setts were detected by extensive surveys carried out between February 2013 and March 2014, both on the hill and surrounding plain areas. As burrows are preferentially located in areas offering dense vegetation cover (Remonti et al. 2006a; Reid et al. 2012) and slope (Balestrieri and Remonti 2000; Revilla et al. 2001; Reid et al. 2012), potentially suitable sites were first located by land-cover digitalised maps (1:10,000) produced by a Geographic Information System (ArcView®) based on Coordination of Information on the Environment (CORINE) Land-Cover maps and satellite images. For each survey, an ca. 1-km2-large area was searched on foot for badger setts and signs by four trained surveyors. Badger footprints, paths and latrines were georeferenced and used to infer the location of setts. Setts were defined as ‘main setts’, multi-entrance (4–15) burrows showing signs of permanent occupation (footprints, bedding material and ‘fresh’ excavated soil), and ‘secondary setts’, which were close to a main sett, had a few entrances (1–4) and, generally, no or scarce signs of use. The latter category included annexes, outliers and subsidiary setts, as defined by Kruuk (1978) and Thornton (1988).
To analyse badger preference towards sett location, we recorded the habitat type where each main sett had been dug. The following four habitat types were considered: woods, vineyards, meadows and urban areas. Expected frequencies were calculated according to habitat availability, assessed from 100 random points created in ArcView by the tool ‘random point theme’ of the extension Animal Movement (with distance from boundary = 10 m and distance from other points = 50 m). Observed and expected frequencies were then compared by the chi-squared test, with Yates’ correction for small samples. Bonferroni’s confidence intervals for the proportion of use were then checked for each habitat type (Neu et al. 1974; Byers et al. 1984).
The size and shape of each home range were assessed from main sett coordinates using the Dirichlet tessellation method (DT; Doncaster and Woodroffe 1993), which provides convex, tessellated polygons having the property that every point within a polygon is nearer to the included sett than to any other neighbouring sett. Polygons were drawn using the extension ‘Animal Movement’ of ArcView 9.2 (Hooge and Eichenlaub 1997).
Assuming that badger home ranges tend to be delimited by hard boundaries (Powell and Mitchell 1998), and these are often left out by most methods for constructing home ranges (Getz et al. 2007), whenever the sides of a tessellation felt next to a clear barrier (e.g. major roads, rivers and canals) or geographical feature (e.g. hill ridges and urbanised areas or other inhospitable terrain), they were successively modified by hand as to draw more realistic home ranges (HRs; Getz et al. 2007; Frantz et al. 2010).
We then calculated the areas of both closed DTs (N = 9) and HRs (N = 12). For each HR, the percent cover of woods was assessed by ArcView based on CORINE Land-Cover maps and ground surveys, and the relationship between home range size and wood cover was tested by Spearman’s correlation coefficient.
To estimate group size, a sample of six setts located in the hilly area was monitored by digital scouting pocket cameras (Multipir 12 and SG550 with Passive Infrared motion sensor), recording 30-s-long videoclips (with a 60-s interval between two successive recordings) throughout the 24-h period. Camera traps were tied to trees about 5 m from each active entrance of each surveyed sett, monitoring both main setts and secondary setts with signs of badger presence simultaneously. Apparently, unused holes were gently closed with branches as to either prevent or detect their use during the trapping session.
Setts were surveyed twice, in March–April 2013 and February–March 2014, respectively, for 7–10 consecutive days each, using from 9 to 19 cameras. Trapping effort was expressed as the number of camera-days (= number of camera traps × number of working days).
The number of adult badger individuals sharing the same sett/entrances was recorded. Although badgers may emerge from one entrance and re-enter the sett through a different hole several times before going off to forage (Roper 2010), the synchronisation of the date and time of all cameras, together with the exam of tail pattern (Dixon 2003), helped to prevent double counts. All entrances having been monitored; each badger leaving the sett at dusk was considered as a group member until no badger re-entered the sett. Successively, whenever possible, to distinguish individual badgers, the tail pattern of any badger leaving the sett was compared to that of badgers known to have re-entered the sett. In case of ambiguity, leaving badgers were not considered as additional group members. We are confident that the survey period was sufficient to assess group size effectively.
Badger density in the study area was then assessed based on the resulting mean number of badgers per sett and the overall area of both DTs and HRs.
Results
Nineteen main setts were found, of which 10 were in the hilly area and 9 in the surrounding lowland (Fig. 1). In the hilly area, setts were not randomly distributed in the available habitats (χ 2 = 11.7, P = 0.011), with woods being used as sett sites more than expected according to their relative availability (Bonferroni’s confidence intervals, P < 0.001).

Badger territorial organisation in the study area (a 14.5-km2 wide hill—marked in light grey—and surrounding lowlands in Lombardy region, north Italy), as assessed by Dirichlet tessellations and, successively, modifying polygons’ borders by hand, based on the presence of geographical features (home ranges; numbers correspond to those in Table 2)
The mean area (±SE) of closed Dirichlet polygons (N = 9) was 2.96 ± 0.47 km2. Correcting polygons’ borders by hand, home range size could be assessed for the 10 badger groups with setts on the hill and also for two groups in the plain, whose ranges were clearly delimited by the course of the River Lambro (Table 2 and Fig. 1). Home range size was, on average, 1.95 ± 0.23 km2 and was inversely correlated to wood percent cover (r = −0.75, P = 0.005, N = 12).
Nineteen latrines were found, of which 15 were dug next to the main sett and the 4 latrines far from setts were successively found out to be next to Dirichlet sides.
The overall trapping effort was 311 camera trap-days for the first session and 443 for the second one (Table 3). Badgers were recorded a total of 643 times; although they were more active during the second trapping session (one videoclip per 2.9 vs. 0.8 camera-days), the minimum total number of individuals recorded scarcely differed (16 vs. 17 badgers, respectively). Groups consisted of two to four adult individuals (Table 3), averaging 2.75 badgers/group. No young of the year was observed.
Based on the overall area of either DTs (26.6 km2) or HRs (23.5 km2) and mean group size, in the hilly area, sett density was 0.34–0.51 sett/km2, whilst badger density was 0.93–1.4 adult individuals/km2, respectively.
Discussion
Whilst at continental scale, badger density may vary according to variation in climate and habitat availability (Kowalczyk et al. 2000), at local scale, density depends on the type of habitat surveyed amongst those available (Johnson et al. 2002) and can vary over time at the same study area (Cheeseman et al. 1987; Macdonald and Newman 2001). As a consequence, every density estimate is likely to be representative of a specific area and time and should not been extrapolated to a wide portion of a species’ range. On the other hand, whilst the factors associated with sett location are well known (e.g. Reid et al. 2012; Acevedo et al. 2014), those underlying variations in badger density throughout its wide distribution range are still poorly known, as a consequence of the low number of available estimates, and data on badger numbers and variation in population size are still needed to assess its conservation status in many European countries, including Italy (Roper 2010).
Keeping in mind the difficulty of comparing density estimates from different studies, in our study area, both badger (see Table 1) and sett densities (mean for continental Europe = 0.16 ± 0.03 setts/km2, calculated from data in Johnson et al. 2002) were rather high with respect to most available data for continental Europe. With respect to neighbouring Switzerland, badger density in our low-altitude area was similar to those recorded, in the pre-reproductive period, in rural areas at 430–630 m asl (1.04 ± 0.71 adults and/or subadults/km2; Do Linh San et al. 2007b) and 995–1288 m asl (1 ind/km2; Do Linh San et al. 2007a).
As recorded for other rural areas (Revilla et al. 2001; Remonti et al. 2006a; Do Linh San et al. 2011), woods were preferred as sett sites. In agricultural areas, wood patches offer cover seclusion and are often associated with unfit-for-cultivation, sloping soils, which offer suitable conditions for burrowing. In the intensively cultivated landscape of northern Italy, wood patches have been reported to be strongly selected by radiotracked badgers (Balestrieri et al. 2009). The negative correlation between home range size and percent wood cover supports the hypothesis that woods also offer profitable food resources (see Kowalczyk et al. 2003) with respect to crops and vineyards. Accordingly, in intensively cultivated areas, badgers have been reported to have unbalanced diets, disproportionately rich in carbohydrates, probably as a consequence of the scarcity of animal food (Remonti et al. 2011).
The accurate estimation of badger density ensues from precise as possible assessment of both group size and study area (i.e. the territory effectively used by monitored individual badgers).
Home range borders as assessed by Dirichlet tessellations matched well with both geographical barriers/features and the location of four latrines in the north-western half of the hill, where home ranges were small. The match was less precise at the foot of the hill, where the presence of urban areas, roads and wide canals suggested reduction by hand of some polygons areas, but, on the whole, the assumption that range borders lie at the halfway point between adjacent main setts was adequate and allowed almost unbiased estimates of range size.
To date, video cameras have been used to study the behaviour of Japanese badger Meles anakuma (Tanaka et al. 2002) and assess the role played by the European badger as a reservoir species of bovine tuberculosis (Tolhurst et al. 2009). Remote-controlled, infrared video surveillance equipment has been also used for continuous nocturnal observation of setts to investigate allo-marking of conspecifics with subcaudal gland secretions by M. meles (Buesching et al. 2003). In our study area, camera traps increased the efficacy of group size assessment with respect to direct observation by enabling the continuous monitoring of badger setts over several nights and, frequently, the observation and comparison of tail appearance.
The consistency of badger numbers per sett over the two sampling periods, notwithstanding wide variation in badger activity, indicates that the estimate of group size was accurate. Results were consistent with those obtained in plain areas of northern Italy by the clearing of four main setts (mean group size = 3 adult individuals; Balestrieri et al. 2006).
As available data indicate that badger group size ranges between two and four adult individuals throughout both hilly and plain areas of the River Po basin (Balestrieri et al. 2006; Remonti et al. 2006b; Balestrieri et al. 2010; this study), sett density may be used as an estimator of badger density throughout this wide area (see also Lara-Romero et al. 2012; Judge et al. 2014). The twofold higher sett density on hills with respect to plain cultivated areas (Quadrelli 1993; Remonti et al. 2006a) suggests that residual woods and/or sloping soils largely influence badger abundance. Further studies are needed to understand which factor—slope, cover and food, i.e. sett site or food availability—plays the main role in shaping badger density.
References
Acevedo P, Gonzalez-Quiros P, Prieto JM, Etherington TR, Gortazar C, Balseiro A (2014) Generalizing and transferring spatial models: a case study to predict Eurasian badger abundance in Atlantic Spain. Ecol Model 275:1–8
Balestrieri A, Remonti L (2000) Reduction of badger (Meles meles) setts damage on artificial elements of the territory. Hystrix Ital J Mammal 11:3–6
Balestrieri A, Remonti L, Prigioni C (2006) The reintroduction of the European badger (Meles meles) to a protected area of northern Italy. Ital J Zool 73:227–235
Balestrieri A, Remonti L, Prigioni C (2009) Habitat selection in a low-density badger Meles meles population: a comparison of radio-tracking and latrine surveys. Wildl Biol 15:1–7
Balestrieri A, Remonti L, Frantz AC, Capelli E, Zenato M, Dettori EE, Guidali F, Prigioni C (2010) Efficacy of passive hair-traps for the genetic sampling of a low-density badger population. Hystrix 21:137–146
Blackwell PG, Macdonald DE (2000) Shapes and sizes of badger territories. Oikos 89:392–398
Broseth H, Knutsen B, Bevanger K (1997) Spatial organization and habitat utilization of badgers Meles meles: effects of food patch dispersion in the boreal forest of central Norway. Mamm Biol 62:12–22
Buesching CD, Stopka P, MacDonald DW (2003) The social functions of allo-marking in the European badger (Meles meles). Behaviour 140:965–980
Byers CR, Steinhorst RK, Krausman PR (1984) Clarification of a technique for analysis of utilization-availability data. J Wildl Manag 48:1050–1053
Campbell G, Kuehl HS, Diarrassouba A, Ń’Goran PK, Boesch C (2011) Long-term research sites as refugia for threatened and over-harvested species. Biol Lett 7:723–726
Caro TM, Young CR, Cauldwell AE, Brown DDE (2009) Animal breeding systems and big game hunting: models and application. Biol Conserv 142:909–929
Cheeseman CL, Wilesmith JW, Ryan J, Mallinsin PJ (1987) Badger population dynamics in a high density area. Symp Zool Soc Lond 58:279–294
De Solla SR, Bonduriansky R, Brooks RJ (1999) Eliminating autocorrelation reduces biological relevance of home range estimates. J Anim Ecol 68:221–234
Delahay RJ, Brown JA, Mallinson PJ, Spyvee PD, Handoll D, Rogers LM, Cheeseman CL (2000) The use of marked bait in studies of the territorial organization of the European badger (Meles meles). Mammal Rev 30:73–87
Dixon DR (2003) A non-invasive technique for identifying individual badgers Meles meles. Mammal Rev 33:92–94
Do Linh San E, Ferrari N, Weber J-M (2007a) Spatio-temporal ecology and density of badgers Meles meles in the Swiss Jura Mountains. Eur J Wildl Res 53:265–275
Do Linh San E, Ferrari N, Weber J-M (2007b) Socio-spatial organization of Eurasian badgers (Meles meles) in a low-density population of central Europe. Can J Zool 85:973–984
Do Linh San E, Ferrari N, Fischer C, Weber J-M (2011) Ecology of European badgers (Meles meles) in rural areas of western Switzerland. In: Rosalino LM, Gheler-Costa C (eds) Middle-sized carnivores in agricultural landscapes. Nova Science Publishers, NY, pp 83–104
Doncaster CP (2001) What determines territory configurations of badgers? Oikos 93:497–498
Doncaster CP, Woodroffe R (1993) Den site can determine shape and size of badger territories: implications for group-living. Oikos 66:88–93
Frantz AC, Schley L, Schaul M, Balestrieri A, Roper TJ (2010) Spatial organisation of badgers (Meles meles) in a medium-density population in Luxembourg. Hystrix Ital J Mammal 21:3–18
Garnett BT, Delahay RJ, Roper TJ (2005) Ranging behaviour of European badgers (Meles meles) in relation to bovine tuberculosis (Mycobacterium bovis) infection. Appl Anim Behav Sci 94:331–340
Getz WM, Fortmann-Roe S, Cross PC, Lyons AJ, Ryan SJ, Wilmers CC (2007) LoCoH: nonparametric kernel methods for constructing home ranges and utilization distributions. PLoS ONE 2(2):e207. doi:10.1371/journal.pone.0000207
Goszczynski J (1999) Fox raccoon dog and badger densities in North Eastern Poland. Acta Theriol 44:413–420
Goszczyñski J, Skoczyñska J (1996) Density estimation, family group size and recruitment in a badger population near Rogów (central Poland). Miscellánia Zoológica 19(2):27–32
Graf M, Wandeler AI, Lups P (1996) Spatial organization and habitat utilization in a population of European badgers (Meles meles L.) in a hilly area of the Swiss midlands. Rev Suisse Zool 103:835–850
Hofmann T, Ebersbach H, Stubbe M (2000) Home range-Größe und Habitat-nutzung beim Europäischen Dachs (Meles meles L., 1758) im nordöstlichen Harzvorland (Sachsen-Anhalt). Beiträge zur Jagd- und Wildforschung 25:199–209
Hooge PN, Eichenlaub B (1997) Animal movement extension to ArcView, vol ver. 1.1. Alaska Biological Science Center, U.S. Geological Survey, Anchorage
Ivanter EV (1973) Notes on an investigation of the badger near the northern limit of its range. Trudy Gosudarstvennogo Zapovednika Kivach 2:164–173 [In Russian]
Johnson DDP, Jetz W, Macdonald DW (2002) Environmental correlates of badger social spacing across Europe. J Biogeogr 29:411–425
Judge J, Wilson GJ, Macarthur R, Delahay RJ, McDonald RA (2014) Density and abundance of badger social groups in England and Wales in 2011–2013. Sci Rep 4:3809. doi:10.1038/srep03809
Kauhala K, Holmala K, Lammers W, Schregel J (2006) Home ranges and densities of medium-sized carnivores in south-east Finland, with special reference to rabies spread. Acta Theriol 51:1–13
Kowalczyk R, Bunevich AN, Jędrzejewska B (2000) Badger density and distribution of setts in Białowieża Primeval Forest (Poland and Belarus) compared to other Eurasian populations. Acta Theriol 45:395–408
Kowalczyk R, Zalewski A, Jedrzejewska B, Jedrzejewski W (2003) Spatial organization and demography of badgers (Meles meles) in Bialowieza Primeval Forest, Poland, and the influence of earthworms on badger densities in Europe. Can J Zool 81:74–87
Kranz A, Tikhonov A, Conroy J, Cavallini P, Herrero J, Stubbe M, Maran T, Fernandes M, Abramov A, Wozencraft C (2008) Meles meles. The IUCN red list of threatened species, Version 2014.3. <www.iucnredlist.org>. Downloaded on 11/02/2015
Kruuk H (1978) Spatial organisation and territorial behavior of the European badger Meles meles. J Zool 184:1–19
Lara-Romero C, Virgós E, Revilla E (2012) Sett density as an estimator of population density in the European badger Meles meles. Mammal Rev 42:78–84
Lobachev YS (1976) The ecology of Meles meles in the mountains of southern Kazakhstan. Byulleten Moskovskogo Obshchestva Ispytatelei Prirody, Otdel Biologicheskiï 81(5):7–21 [In Russian with English summary]
Macdonald DW, Newman C (2001) Badger (Meles meles) population dynamics in Oxfordshire, UK: numbers, density and cohort life histories, and a possible role of climate change in population growth. J Zool 256:121–138
Macdonald DW, Newman C, Dean J, Buesching CD, Johnson PJ (2004) The distribution of Eurasian badger, Meles meles, setts in a high-density area: field observations contradict the sett dispersion hypothesis. Oikos 106:295–307
Marmi J, López-Giráldez F, Macdonald DW, Calafell F, Zholnerovskaja E, Domingo-Roura X (2006) Mitochondrial DNA reveals a strong phylogeographic structure in the badger across Eurasia. Mol Ecol 15:1007–1020
Martin-Franquelo R, Delibes M (1985) Biology of the badger (Meles meles) in Donana, Mediterranean Spain. In: Fuller WA, Nietfeld NT, Harris MA (eds) 4th International Theriological Congress, Edmonton. Texas Tech University Press, Lubbock
Matyástík T, Bicík V (1999) Distribution and habitat selection of badger (Meles meles) in northern Moravia. Acta Universitatis Palackianae Olomucensis Biologica 37:77–88
Mori E, Menchetti M, Balestrieri A (2014) Interspecific den sharing: a study on European badger setts using camera traps. Acta Ethol 18:121–126
Neu CW, Byers CR, Peek JM, Boy V (1974) A technique for analysis of utilization-availability data. J Wildl Manag 38:541–545
Nichols JD, Williams BK (2006) Monitoring for conservation. Trends Ecol Evol 21:668–673
Pelikan J, Vackar J (1978) Densities and fluctuation in numbers of red fox, badger, and pine marten in the Buèin Forest. Folia Zool 27:289–303
Pigozzi G (1987) Behavioural ecology of the European badger (Meles meles L.): diet, food availability and use of space in the Maremma Natural Park, central Italy. PhD dissertation, University of Aberdeen
Powell RA, Mitchell MS (1998) Topographical constraints and home range quality. Ecography 21:337–341
Prigioni C, Deflorian MC (2005) Sett site selection by the Eurasian badger (Meles meles) in an Italian Alpine area. Ital J Zool 72:43–48
Quadrelli G (1993) Densità e distribuzione delle tane di tasso Meles meles nel basso Lodigiano. Natura Bresciana 28:429–431
Reid N, Etherington TR, Wilson GJ, Montgomery WI, McDonald RA (2012) Monitoring and population estimation of the European badger Meles meles in northern Ireland. Wildl Biol 18:46–57
Remonti L, Balestrieri A, Prigioni C (2006a) Factors determining badger Meles meles sett location in agricultural ecosystems of NW Italy. Folia Zool 55:19–27
Remonti L, Balestrieri A, Prigioni C (2006b) Range of the Eurasian badger (Meles meles) in an agricultural area of northern Italy. Ethol Ecol Evol 18:61–67
Remonti L, Balestrieri A, Prigioni C (2011) Percentage of protein, lipids and carbohydrates in the diet of badger (Meles meles) populations across Europe. Ecol Res 26:487–495
Revilla E, Palomares F (2002) Spatial organization, group living and ecological correlates in low-density populations of Eurasian badgers, Meles meles. J Anim Ecol 71:497–512
Revilla E, Delibes M, Travaini A, Palomares F (1999) Physical and population parameters of Eurasian badgers (Meles meles L.) from Mediterranean Spain. Mamm Biol 64:269–276
Revilla E, Palomares F, Fernandez N (2001) Characteristics, location and selection of diurnal resting dens by Eurasian badgers (Meles meles) in a low density area. J Zool 255:291–299
Rodríguez A, Martín R, Delibes M (1996) Space use and activity in a Mediterranean population of badgers Meles meles. Acta Theriol 41:59–72
Rogers LM, Cheeseman CL, Mallinson PJ, Clifton-Hadley R (1997) The demography of a high-density badger (Meles meles) population in the west of England. J Zool 242:705–728
Roper TJ (2010) Badger. Harper Collins Publishers, London
Rosalino LM, Macdonald DW, Santos-Reis M (2004) Spatial structure and land-cover use in a low density Mediterranean population of Eurasian badgers. Can J Zool 82:1493–1502
Scheppers TLJ, Frantz AC, Schaul M, Engel E, Breyne P, Schley L, Roper TJ (2007) Estimating social group size of Eurasian badgers Meles meles by genotyping remotely plucked single hairs. Wildl Biol 13:195–207
Tanaka H, Yamanaka A, Endo K (2002) Spatial distribution and sett use by the Japanese badger, Meles anakuma. Mammal Study 27:15–22
Thornton PS (1988) Density and distribution of badgers in S.W. England—a predictive model. Mammal Rev 18:11–23
Tinelli A, Tinelli P (1980) Le tane di istrice e di tasso. Censimento e densità delle tane nella Riserva Presidenziale di Castelporziano per la conservazione dell’istrice e del tasso. Segretariato generale della Presidenza della Repubblica. Tenuta di Castelporziano, Rome [in Italian]
Tolhurst BA, Delahay RJ, Walker NJ, Ward AI, Roper TJ (2009) Behaviour of badgers (Meles meles) in farm buildings: opportunities for the transmission of Mycobacterium bovis to cattle? Appl Anim Behav Sci 117:103–113
Tuyttens FAM, Long B, Fawcett T, Skinner A, Brown JA, Cheeseman CL, Roddam AW, Macdonald DW (2001) Estimating group size and population density of Eurasian badgers Meles meles by quantifying latrine use. J Appl Ecol 38:1114–1121
Van Apeldoorn RC, Vink J, Matyáštík T (2006) Dynamics of a local badger (Meles meles) population in the Netherlands over the years 1983–2001. Mamm Biol 71:25–38
White GC, Garrott RA (1990) Analysis of wildlife radio-tracking data. Academic Press, San Diego
Woodroffe R, Macdonald DW (1993) Badger sociality—models of spatial grouping. Symp Zool Soc Lond 65:145–169
Woodroffe R, Macdonald DW (1999) Helpers provide no detectable benefits in the European badger, Meles meles. J Zool 250:113–119
Zoss A (1992) Some aspects of the ecology of European badger (Meles meles L.) in the Slitere nature reserve. Acta et Commentationes Univ Tartuensis 995:176–185
Acknowledgments
We thank Maurizio Macchetta, who helped with field work, and Giuseppe Bogliani, for the logistic support. Lesley C. Wright kindly revised the paper for English language use.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Balestrieri, A., Cardarelli, E., Pandini, M. et al. Spatial organisation of European badger (Meles meles) in northern Italy as assessed by camera-trapping. Eur J Wildl Res 62, 219–226 (2016). https://doi.org/10.1007/s10344-016-0996-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-016-0996-y