
Abstract
While the effects of seed size on survival and early seedling growth have been demonstrated in Quercus spp., there is a knowledge gap regarding these effects under water limiting conditions. To address this need, we studied cork oak (Quercus suber L.) seedling development among treatments including three acorn classes (small: < 4 g, intermediate: 4–6 g), and large: ≥ 6 g) and two water regimes (well-watered versus water stressed). The following hypotheses were proposed: the reserve effect (larger seeds retain a larger proportion of reserves after germinating), the metabolic effect (seedlings from larger seeds have slower relative growth rates), the seedling-size effect (larger seeds produce larger seedlings), and that plant tolerance to water stress increases with increasing seed size. Acorn size had no effect on duration and rate of acorn germination nor on seedling survival, but it significantly affected almost all growth parameters, excepting the number of growth unit and leaves and leaf area. Seedlings coming from large acorns always showed the highest values for the affected growth parameters, followed by those coming from acorns of intermediate size; seedlings coming from small acorns always exhibited the lowest values. Water stress significantly affected survival rate (88.2% for well-watered seedlings versus 64.7% for water stressed seedlings) and biomass of fine roots and stems, but independently of acorn size. Our results are in line with both hypothesizes of the reserve effect and seedling-size effect, but not for the metabolic effect nor the plant tolerance to water stress increases with increasing seed size hypotheses. In general, our results show that large acorns had no advantage in survival or seedling growth under water stress comparatively to intermediate or small acorns, at least under the conditions of our experiment.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Oaks (Quercus spp.) display great variation in seed size within a species (Khan and Shankar 2001). Seed size variation may affect seed germination, survival, emergence (Khan and Shankar 2001), seedling establishment (Quero et al. 2007), growth rate, and final seedling size (Karrfalt 2004; Merouani et al. 2001). The influence of acorn size is most pronounced during germination and the first growing season. The mechanisms by which seed size influences seedling performance appear to be variable. Thus, three mechanisms have been postulated to help explain functional seed–seedling relationships (Westoby et al. 1996; Leishman et al. 2000). The first is the reserve effect by which larger seeds hold a large amount of stored reserves, which can then be used to fuel seedling growth, maintenance, and repair. This may improve the early development of the growing plant (Jurado and Westoby 1992; Bonfil 1998) and increase its chances of survival (Bonfil 1998). Second is the metabolic effect, which reports that seedlings from larger seeds have slower relative growth rates and use reserves more slowly. This improves long-term survival (Wright and Westoby 1999). Third is the seedling-size effect by which larger seeds produce larger seedlings, which are more capable i) of reaching deeper soil layers (Bond et al. 1999), allowing them to gain greater access to soil water resources, and ii) of overtopping neighboring seedlings and capturing more light (Foster 1986).
In this study we focused on a Mediterranean oak species, the evergreen sclerophyllous cork oak (Quercus suber L.). In Tunisia, cork oak forests are mainly plentiful in the north-west of the country that corresponds to the natural area of the species. The species has great importance from an ecological, social and economic point of view. Cork oak forests represent 10% of the total forest area in Tunisia and 4.3% of the world’s cork oak forest area. However, area of cork oak forests has decreased dramatically, from 148,000 to 70,000 ha, between 1920 and 2005 (Selmi 2006). Multiple and diverse factors have caused this reduction including aging, the effect of a complex disease known as cork oak decline, cork oak dieback or cork oak mortality (Ja et al. 2011), and difficulty of natural and artificial regeneration mainly linked to overgrazing (Hasnaoui 1992), vegetation competition (Chaar et al. 2008), acorn predation (Vieira 1951), slow growth (Chaar et al. 2008), and low survival in particular during the first summer drought (Vieira 1951). In a previous study conducted on the same species, we found that use of tree shelters may help to overcome the problem of grazing and slow growth of the species and that vegetative competition may have no effect on seedling growth on fertile sites (Mechergui and Pardos 2017).
Despite efforts to understand the effects of seed size on survival and early seedling growth in Quercus spp., there is a knowledge gap regarding these effects under low water availability. Thus, our objective was to study the effect of seed size on survival and early development of cork oak seedlings subjected to severe water stress. In accordance with the possible mechanisms by which seed size influences seedling performance as described above, the following hypotheses were proposed: the reserve effect (larger seeds retain a larger proportion of reserves after germinating), the metabolic effect (seedlings from larger seeds have slower relative growth rates), and the seedling-size effect (larger seeds produce larger seedlings). We also hypothesized that plant tolerance to water stress increases with increasing seed size.
Materials and methods
Plant material and experimental design
The experiment was established at the research station of Sejnane (37° 03′ 33″ N, 9° 13′ 24″ W, 138 m a.s.l.) in northwestern Tunisia. The climate is Mediterranean with an annual mean temperature of 18.2 °C (1975–2004), average maximum and minimum temperatures of 34.4 °C and 5.6 °C, respectively, and average annual rainfall of 912 mm. Mature acorns were collected during September from six trees in a natural population of Quercus suber L. growing in Ain Zena in Northwestern Tunisia (36° 44′ 57.53″ N, 8° 54′ 33.57″ E, elevation 200 m a.s.l.) where the climate is Mediterranean with a mean annual temperature of 18.2 °C and a mean rainfall of 923 mm (Boussaïdi 2012). Acorns were surveyed and dry and infected ones were discarded. They were then floated in water to separate viable from parasitized acorns. Healthy acorns were individually weighed before sowing to determine their fresh mass.
Fifty acorns of each size class (small (< 4 g), intermediate (4 to < 6 g), and large (≥ 6 g), were sown separately (December-2017) at 3-cm depth in the topsoil, corresponding to recommended sowing depth (Hasnaoui 1992), in 8-L cylindrical plastic pots (23 cm height, 397.7 cm2 and 298.7 cm2, upper and lower cross-sectional area, respectively). Pots were filled with 1/3 sand and 2/3 forest soil. They were kept outdoors under natural conditions. They were randomly divided in two groups according to the water regime, one group was watered to field capacity four days per week (W +), while the other group (W–) was subjected to a series of two soil-drying cycles of 18 and 19 days, respectively, beginning on 6 July and extending to 15 August. The timing and duration of the drought correspond to that occurring naturally in the area of occurrence of the studied species. Water-stressed seedlings were watered at field capacity, only twice (25 and 27 July) at the end of the first drying cycle. Volumetric soil water content was monitored four days per week at 15 cm depth with a Soil Moisture Meter (Model: PMS-714, Range: 0 to 50% moisture content with 0.1% resolution). Volumetric soil water content in the well-watered seedlings (W +) was maintained at 100% of field capacity, while seedlings subjected to severe water stress (W–) were allowed to dry to a water content at 25% of field capacity. Under each water regime, pots were randomly divided in three groups according to the three acorn size classes. Before water stress application, all seedlings (W + and W-) were kept well-watered four days per week.
The experiment was a split–plot design with Whole Plots in Randomized Blocks with five replications (or blocks). The whole-plots were formed by the two irrigation regimes and the subplots by the three seed size classes. Each subplot was represented by five pots with one plant per pot. Therefore, 150 plants were used in total (5 blocks × 6 treatments × 5 plants per treatment). The two water regimes were well watered versus water stressed.
Measurements
Measurements were made on all surviving seedlings (all with terminal buds) at the end of the experiment (15 August 2018), except for germination which was assessed regularly, each day. An acorn was considered germinated after the emergence of the epicotyl from the soil. Measured variables were: (1) qualitative including percentage of germination, survival, number of growth units (GU; a distinct portion of the main stem established during the growth phase) and seedling posture (erect posture or bent posture (horizontal growth, phenomena which was observed for cork oak)) and (2) quantitative including duration of germination (date of germination minus date of sowing) and growth period (date of the harvest of seedlings at the end of the experiment minus the date of germination), basal diameter, plant height, seedling vigor index (germination percentage × seedling dry weight), number of ramifications and leaves per seedling, individual and total area of leaves, leaf mass per area, plant biomass, shoot/root dry weight ratio, amount of mobilized and remnant reserves and the reserve mobilization capacity.
Number and length of GU were determined. Basal diameter was measured using an electronic caliper. Leaf area was calculated using a scanner and MESURIM PRO software. The leaf mass per area (LMA) was determined by dividing total dry weight of leaves by total area. To determine plant biomass, leaves were removed from stems, stems were severed at the root-collar, and all roots were then rinsed free of soil. Weight of above- (shoots and leaves) and below-ground (taproot and fine roots) biomass was then determined after drying at 70 °C until weight became constant. Total biomass was calculated from above- and below-ground biomass. Shoot/root ratio was calculated for each plant from dry weights of the biomass components. The amount of mobilized reserves (i.e., cotyledonary reserves mobilized into seedlings) depending on acorn size was determined by subtracting dry mass of the remnant cotyledons at harvest to the estimated initial dry mass of cotyledons (g). The reserve mobilization capacity of each acorn into seedlings was determined as follows:
To estimate seed dry mass (cotyledonary dry mass), a subsample of each acorn class was weighed and then oven-dried at 70 °C until biomass became constant. They were then separated from their pericarp and weighed to determine seed dry mass. Acorn specific regression equations were made for seed dry mass vs. acorn fresh mass. The regression used to estimate for each plant the seed dry mass from the acorn fresh mass were:
-
1
Seed dry mass for large acorn = 0.158 + 0.826 × acorn fresh mass (N = 50, R2 = 0.99);
-
2
Seed dry mass for acorns of intermediate size = -0.102 + 0.883 × acorn fresh mass (N = 50, R2 = 0.99);
-
3
Seed dry mass for small acorns = 0.043 + 0.169 × acorn fresh mass (N = 50, R2 = 0.97).
Statistical analysis
All qualitative variables were expressed in percentage (%) and all statistical analyses were performed using Statistical Analysis System (SAS Institute Inc., Cary, NC, Version 9.0). A comparison of mean proportions (%) was then performed through the χ2-test by using the PROC FREQ procedure. Differences were considered significant at P ≤ 0.05. For each analysis, when statistically significant differences between treatments were found, likelihood confidence intervals at 5% level are calculated for the proportions.
With regard to quantitative variables, duration of acorn germination and seedling vigor index were analyzed according to model 1, using proc GLM, where the effect of water regime was excluded from this model, since water stress was applied later, after acorn germination occurred, while for the duration of growth period merely a simple comparison of mean values between acorn size treatments was done. The other quantitative variables were analyzed according to model 2, using PROC MIXED. For each analysis, when the ANOVA was significant, statistically significant differences between means were identified using Tukey–Kramer multiple comparison test, at P ≤ 0.05.
where Yij = is the mean value measured in the plant on the acorn size treatment i, on the block j. µ = the grand mean.
where Yijk = is the mean value measured in the plant on the water regime treatment i, and the acorn size treatment j, on the block k. µ = the grand mean.
To test the hypothesizes of the reserve effect, the metabolic effect, and the seedling-size effect (see introduction) we respectively tested, as described by Quero et al. (2007), the significance of the relationship between reserves used and seed dry mass, the relationship between relative growth (RGR) and seed dry mass, and the relationship between seedling biomass and seed dry mass. The relationship between the reserves used and seed mass was parameterized as follows:
where a1 was the intercept and b1 was the slope. If b1 was inferior to 1, condition 1 of the reserve effect hypothesis is accepted. To accept condition 2 of the reserve effect, the slope of the ln of seedling mass vs. the ln of seed mass should also be significantly greater than that of the ln of used reserve vs. the ln of seed mass.
The relationship between relative growth (RGR) and seed mass was parameterized by:
where RGR, rate of biomass increase per unit of plant biomass, was calculated as the natural log (ln) of seedling biomass minus the natural log of used seed reserve (Steege et al. 1994) divided by the growth period. If b2 was inferior to 0, the metabolic-effect hypothesis was accepted.
The relationship between seedling biomass and seed mass was parameterized by:
If b3 was superior to 0, the hypothesis of seedling-size effect was accepted.
All analyses of the correlation were done with the test of Pearson correlation, using the PROC CORR procedure of SAS.
Results
Effect of acorn size on germination and survival rate
Seed mass varied from 1.75 to 3.94 g for small acorns, 4 to 5.97 g for acorns of intermediate size, and from 6 to 9.44 g for large acorns, with a mean size of 3 g, 5 g, and 7.2 g, respectively. Differences among seed mass classes were significant (P < 0.0001). Acorns moisture concentration (acorn fresh weight minus dry weight) was 48.6–50.1% (mean: 49%) for small acorns, 48.2–48.6% (mean: 48.4%) for intermediate size and 48–48.2% (mean: 48.1%) for large acorns.
Seedlings began to germinate by the beginning of March. All acorn classes exhibited only one peak of germination, during the first week (Fig. 1). During this first week more than half of acorns, for each acorn class, germinated; germination of the remainder was spread out over five weeks for both intermediate and large size of acorns and seven weeks for small acorns. Duration of acorn germination was of 61–105 days (mean: 67 days) for large acorns, 61–104 days (mean: 68 days) for intermediate acorns and 61–102 days (mean: 72 days) for small acorns, with no significant differences between seed size. Seed germination capability was not affected by seed size, and final germination rates were similar for all acorn classes (P = 0.10), with a germination rate of 46% for large acorns, 47% for intermediate acorns and 44% for small acorns.

Percentage of emerged acorns according to acorn size (S, small acorn; I, acorn of intermediate size; L, acorn of large size)
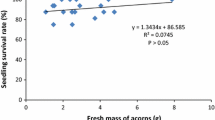
At the end of the experiment, survival rate was 76.4%. Survival was independent of acorn size (P = 0.96) and its interaction with water regime (P = 0.20), but significantly affected by water regime (P = 0.0012), resulting in a higher value under non-limiting water conditions than under water stress (88.2 ± 5.4 vs. 64.7% ± 5.7 (± likelihood confidence limits)).
Effect of acorn size on seedling growth
Duration of growth period (date at the end of experiment minus date of germination) varied between 156 and 161.4 d, with an overall mean of 159.3 d, which was not significantly different between all seed size classes (P = 0.07). Seedling height was independent of water regime (P = 0.18) and water regime × acorn size interaction (P = 0.10), but significantly affected by acorn size (P < 0.0001). At the end of the experiment, i.e., after a growth mean period of ca. 159.3 d, height varied between a minimum of 14.2 cm and a maximum of 60.2 cm for seedlings from large acorns, 14.9 cm and 48.2 cm for seedlings from acorns of intermediate size, and 5.8 cm and 44.1 cm for those from small acorns. Mean height of seedlings coming from large acorns was 15.6% greater than that of those from intermediate, although the difference was not significant, and 57% greater than that of those from small acorns, which was significant (Fig. 2). Difference between small acorns and acorns of intermediate size was also significant.

Height, diameter, seedling vigor index and length of established growth unit (GU), after ca. 159.3 d, according to acorn size. Data are expressed as mean ± SE. Means marked with different letters were significantly different according to the Tukey–Kramer multiple comparison test, at P ≤ 0.05 level
Similar to height growth, neither water regime (P = 0.37) nor its interaction with acorn size (P = 0.18) had a significant effect on diameter growth, while a significant effect of acorn size was found (P < 0.0001). Basal stem diameter of seedlings coming from large acorns ranged from 2.2 to 6.4 mm, while that of those coming from intermediate and small acorns ranged from 2.7 to 4.7 mm and from 1.4 to 5.1 mm, respectively. On average, basal stem diameter of seedlings coming from large acorns was 18.2% and 34.5% larger than that of those coming from intermediate and small acorns, respectively (Fig. 2). All differences were significant.
Seedling vigor index, which was analyzed only for seed size (see details in statistical analyses), showed significant differences between treatments (P < 0.0001). It was significantly greater for seedlings coming from large acorns than for seedlings coming from acorns of intermediate size, which were significantly greater than those coming from small acorns (Fig. 2).
Analyses on GU showed that plants formed one to three GU along the main stem, with a trend of all plants to establish only 2 GU (Table 1). This number of GU was not influenced by water regime (P = 0.64) nor acorn size (P = 0.21) and their interaction (P = 0.22). With regard to GU length, there was no significant effect of water regime (P = 0.35) nor its interaction with acorn size (P = 0.99), while a significant effect of acorn size was found (P = 0.0044). Seedlings coming from large acorns had the longest mean GU length, which was not different from that of seedlings coming from acorns of intermediate size, but were significantly different from that of seedlings coming from small acorns. Differences between seedlings coming from small acorns and those coming from acorns of intermediate size were not significant (Fig. 2).
Both individual leaf area and leaf mass per area (LMA) were found to be independent of water regime (P = 0.97; P = 0.53, respectively) and acorn size (P = 0.76; P = 0.88, respectively) and their interaction (P = 0.93; P = 0.24, respectively). With regard to number and total area of leaves, they were found also to be independent of water regime (P = 0.21; P = 0.14, respectively) and its interaction with acorn size (P = 0.13; P = 0.0650, respectively), but they were influenced by acorn size (P = 0.0495; P = 0.0057, respectively). They were highest for seedlings from large acorns, followed by those from acorns of intermediate size, but the difference was not significant (Fig. 3). The lowest number was found in the seedlings from small seedlings, but did not significantly differ from that of seedlings from acorns of intermediate size. Number of ramifications was also significantly influenced by acorn size (P < 0.0001), while no effects were found for water regime (P = 0.88) and its interaction with acorn size (P = 0.76). It was significantly higher for seedlings from large acorns than for those from intermediate and small acorns, for which the difference was not significant (Fig. 3). Percentage of seedlings showing horizontal growth instead of vertical growth was significantly higher for seedlings from large acorns than for those from acorns of intermediate and small size for which the difference was not significant (Fig. 3).

Number and total area of leaves, number of ramifications (mean ± SE) and percentage of seedlings having horizontal growth (mean percentage ± likelihood confidence limits), after ca. 159.3 d, according to acorn size. Means marked with different letters were significantly different according to the Tukey–Kramer multiple comparison test, at P = 0.05 level
Both total above-ground biomass and total below-ground biomass were significantly affected by acorn size (Table 2). However, while total above-ground biomass was significantly affected by water regime, total below-ground biomass was not. In addition, more detailed analyses showed that at the level of above-ground biomass there was a significant effect of water regime for shoot biomass, but not for biomass of leaves, while at the level of below-ground biomass there was a significant effect of water regime for fine roots biomass, but not for taproot biomass. Acorn size affected the different fractions of both below- and above-ground biomass (leaves, shoots and roots). With regard to seedling biomass (total biomass), there was a significant effect only of acorn size, with seedlings produced by large acorns showing a greater value (9.4 ± 0.46 g) than those produced by acorns of intermediate size, themselves had a greater value (7.06 ± 0.46 g) than seedlings produced by small acorns (4.6 ± 0.48 g). All differences were significant. For the weight of the total above- and below-ground biomass and the different fractions of above- and below-ground biomass, seedlings from large acorns had the highest biomass values, followed by seedlings from acorns of intermediate size; seedlings from small acorns had always the lowest values, with significant differences between all acorn size treatments (Fig. 4). Water regime effect resulted in significantly higher fine roots biomass, shoot biomass and total above-ground biomass in well-watered seedlings (0.55 ± 0.04 g, 1.73 ± 0.12 g, 3.57 ± 0.23 g, respectively) than in stressed ones (0.38 ± 0.04, 1.22 ± 0.13 g, 2.69 ± 0.26 g, respectively). For shoot/root ratio, there was no effect of water regime (P = 0.13) and its interaction with acorn size (P = 0.17), but a significant effect of acorn size (P = 0.0153). The highest value of this ratio was found for seedlings from large acorns (0.42 ± 0.30), followed by those from acorns of intermediate size (0.36 ± 0.30), but the difference was not significant. Seedlings from small acorns had the lowest ratio (0.31 ± 0.31), but did not significantly differ from seedlings from acorns of intermediate size.

Different fractions of above- and below-ground biomass and total above- and below-ground biomass after ca. 159.3 d, according to acorn size. Data are expressed as mean ± SE. Means marked with different upper case letters for total above- and below-ground biomass and lower-case letters for each above-ground (leaves, shoots) or below-ground (taproot, fine roots) component were significantly different according to the Tukey–Kramer multiple comparison test, at P ≤ 0.05 level
Mobilization and remnant of cotyledon reserves
Amounts of mobilized reserves were significantly different between acorns size treatments (P < 0.0001). By contrast, there were no significant differences between water regime treatments (P = 0.54). Water regime × acorn size interaction was also not significant (P = 0.47). The same pattern was also observed for the mobilization capacity of cotyledon reserves (proportion of reserves mobilized), which showed significant differences for acorn size treatments (P < 0.0001), but not for water regime treatments (P = 0.78). Water regime × acorn size interaction was not significant (P = 0.47). Neither water regime (P = 0.48) nor its interaction with acorn size (P = 0.52) were found to have a significant effect on the remnant of cotyledon reserves, but a significant effect of acorn size was found (P < 0.0001). Both mobilized and remnant reserves increased with increasing seed size, with large acorns always having the highest and small acorns the lowest values; intermediate values were associated with acorns of intermediate size, with significant differences between all treatments for both parameters (Fig. 5). The mobilization capacity of cotyledon reserves increased with decreasing seed size, where large acorns had the lowest value, and small acorns had the highest value (Fig. 5). Intermediate value was recorded for acorns of intermediate size. All differences between treatments were significant.

Amount of mobilized and remnant cotyledon reserves and reserve mobilization capacity of cotyledons, according to acorn size. Data are expressed as mean ± SE. Means marked with different letters were significantly different according to the Tukey–Kramer multiple comparison test, at P = 0.05 level. The amount of mobilized reserves was determined by subtracting dry mass of the remnant cotyledons at harvest (g) to the estimated initial dry mass of cotyledons (g). The amount of remnant reserves (g) corresponds to dry mass of the remnant cotyledons at harvest (g). Reserve mobilization capacity corresponds to the estimated dry mass of cotyledons (g) minus dry mass of the remnant cotyledons (g) divided by estimated dry mass of cotyledons (g) then multiplied by 100
Hypothesis tests
Reserve-effect hypothesis— seed reserves used for the 159 d of growth were significantly (P < 0.0001) and positively related to the initial seed mass under both water regimes (Fig. 6). Furthermore, condition 1 of the reserve effect was fulfilled (the Standardized Major Axis (SMA) slope was 0.76, inferior to 1 under both water regimes). To fulfill condition 2 of the reserve effect, however, the slope of the ln of seedling mass vs. the ln of seed mass should also be significantly greater than that of the ln of used reserve vs. the ln of seed mass, which was the case in our study (P < 0.0001). Hence, condition 2 was also fulfilled and consequently the reserve-effect hypothesis was accepted for both well-watered and stressed seedlings.

Used seed reserve, relative growth rate (RGR), and seedling biomass after ca. 159 d of growth vs. ln initial seed dry mass. Pearson correlation and significance are indicated as: ns, not significant; (*), significant at P ≤ 0.05 level. R, Pearson correlation coefficient. The standardized major axis regression (SMA) lines are given when they are significant: continuous line for well-watered seedlings (W +) and dotted line for stressed seedlings. The slope of each SMA regression (S) and their significance against the null model (S = 1 for the reserve effect, and S = 0 for the metabolic effect and the seedling-size effect) is given
Metabolic-effect hypothesis—RGR was not correlated to seed mass for both water regimes (P = 0.14 and P = 0.10) stressful and non-stressful water conditions, respectively) (Fig. 6), which consequently did not support the metabolic-effect hypothesis.
Seedling-size effect hypothesis—After 159 d of growth, seedling biomass was significantly (P < 0.0001) and positively correlated to seed mass under both water regimes (Fig. 6). The SMA slopes were superior to 0, under both water regimes. So, the hypothesis of seedling-size effect was accepted for both well-watered and stressed seedlings.
Acorn-size effect on resistance on water stress hypothesis—The water stress × seed size interaction had no significant on survival nor growth parameters. This indicates that the water stress effect was independent of acorn size, which consequently is not in line with the acorn size effect hypothesis on subsequent seedling resistance to water stress.
Discussion
Effect of acorn size on germination and survival rate and its role in mobilization of cotyledon reserves
Many factors, including rodent seed predation and summer drought (Vieira 1951), vegetative competition and slow growth of seedlings during the early stages (Chaar et al. 2008) and overgrazing (Hasnaoui 1992) are cited as factors that affect cork regeneration in the Mediterranean region. In our study, cork oak acorns showed high germination rates for all acorn size (mass) classes (88–94%), with no significant differences related to acorn size, indicating that acorn germination does not constitute a problem for regeneration of this species, if acorns are healthy (Hasnaoui 1992). These results are, however, contradictory with those of Pandey et al. (2017) in Quercus leucotrichophora, who reported a significant effect of seed size on final germination percentage, where decreasing acorn size resulted in lower germination. In addition, we did not find that acorn size influenced duration of germination, inconsistent with studies reporting that large sized seeds germinate faster (Norden et al. 2008). Furthermore, survival rate was high (75.5–77.7%) and did not show significant differences between all acorn size classes, suggesting that, in our study, acorn size did not determine acorn survival at least during the first six months following germination. These findings agree with results reported by Zolfaghari et al. (2013) and by Yi et al. (2015). Past studies have shown that acorns of various oak species germinate and successfully establish after partial cotyledon loss (Yi and Yang 2010; Perea et al. 2011; Yang and Yi 2012). Giertych and Suszka (2011) observed that Q. robur acorns can germinate and establish even after the excision of 2/3 from their cotyledon weight. This suggests that small acorns contain enough cotyledon reserves to support seedling survival, which could explain why they did not show significant differences in survival compared to acorns of intermediate or large size.
Yi et al. (2015) observed that large acorns mobilized more cotyledon reserves to produce seedlings, compared to small acorns. Our study confirmed these results, showing that the amount of mobilized reserves increases as seed size increases (Fig. 6), highlighting the importance of seed size in an evolutionary context. However, contrary to these authors, who found no significant difference in the reserve mobilization capacity of cotyledons between large and small acorns, our results show significant differences between all acorn size classes, indicating that acorns had varying ability to mobilize reserves into seedlings based on acorn size (i.e., the smaller the acorn, the greater their ability to mobilize cotyledon reserves).
Remnant reserves in the cotyledons
Although early seedling development is based mainly on seed reserves, all acorn size classes showed, after almost six months of growth, that more than 30–40% of dry mass reserves were stored in the remnant cotyledons, in agreement with results of Yi et al. (2015). However, while mobilized reserves increased with increasing seed size, the amount of reserves in the remnant cotyledons followed the same trend and increased with increasing seed size. This is because larger seeds have more capacity to store greater amounts of carbohydrate in their endosperm or cotyledons than small seeds (Milberg and Lamount 1997), which could be confirmed by the reserve effect (larger seeds retain a larger proportion of reserves after germination) found in this study, both under stressful (water stress) and non-stressful conditions. The reserves in the remnant cotyledons were found to be more important for ensuring seedling resprouting than for supporting seedling growth and survival (Sonesson 1994; Yi and Liu 2014). Indeed, prior studies point out that up to 50% (Giertych and Suszka 2011; Perea et al. 2011; Liu et al. 2012) of the cotyledon reserves and perhaps more (Yi et al. 2015) are unnecessary for acorn germination and seedling survival (Giertych and Suszka 2011; Perea et al. 2011; Liu et al. 2012). Nevertheless, it is important to note that the capacity of Quercus seedlings to resprout is not dependent on seed size (Erniwati 2006), probably because the remaining reserves are sufficient for resprouting. It may be that repeated clipping would have revealed a seed-size effect (Quero et al. 2007).
Effect of acorn mass on seedling growth
While no significant effect on survival was found, acorn mass significantly influenced seedling growth, which resulted in the tallest, largest and greatest biomass (total biomass) seedlings coming from large seeds comparatively to those coming from intermediate and small seeds, consistent with the results reported by Khan and Shankar (2001) in Quercus semiserrata. These results verify the seedling-size effect indicating that larger seeds produce larger seedlings. The decomposition of the main stem on further morphological entities showed no difference between acorn size treatments with regard to the number of growth unit (GU), a distinct portion of the main stem established during the growth phase, while for GU length large acorns produced seedlings with longer GU compared to those from small acorns. Thus, according to these results, the greater height of seedlings from large seedlings in comparison with those from small acorns is the result of longer GU. Benefits offered by large seeds are due to the fact that large seeds contain greater reserves of protein, carbohydrate, lipid and energy than intermediate size and small seeds (Khan and Shankar 2001). Large seeds provide advantages not only for producing large seedlings, which are better competitors (Metcalfe and Grubb 1997; Lloret et al. 1999), and thus, better able to gain greater access to resources, but also for producing robust seedlings as proved by the highest value of seedling vigor index associated with seedlings coming from large seeds. Increased seedling vigor index with increasing seed size was also observed by Pandey et al. (2017). Coupling with the results of the present study, seed size clearly plays a strong role in the vigor of the future plant.
Our data show also that large acorns result in seedlings with a higher shoot/root ratio than did small acorns, suggesting that more cotyledon reserves in large acorns were more mobilized into shoots than in roots. Thus, due to their well-developed shoots, seedlings from large acorns can sustain higher photosynthetic activity relative to those from small seeds (Yi et al. 2015). Indeed, total biomass (Dupraz 1997) and number of leaves per plant, which are good indicators of photosynthetic activity, were significantly higher for seedlings from larger than smaller acorns. Oak in general and cork oak in particular are recognized by the development of a large taproot (Hasnaoui 1992). This trend was, however, in our case more pronounced for seedlings from large acorns, conferring them the capacity to explore deep layers of the soil profile, under drought conditions (Leskovar 1995) and to be well anchored to the soil (Zobel 2005), which highlights another advantage of large acorns.
Effect of water stress on survival and seedling growth
Prior experiments have compared oak seedling development according to acorn size, without including external factors such as water stress. In our study, both survival and seedling growth were found to be affected by water stress but independent of seed size, showing that seed size had no advantage, subsequently, on the resistance of plant to water stress. These results are in contrast with those of Khurana and Singh (2000), who reported that under severe water stress seedlings from large seeds were more tolerant to water stress than those from smaller seeds. However, our results correspond with those of Jacobs et al. (2009), who found that larger Quercus rubra seedlings (graded by root volume) did not show greater drought avoidance during the first season after transplant. The lack of significant water stress effects on seedling growth in our study indicates that seedling growth may be more reliant on cotyledon reserves than on external environmental factors. The fact that water stress affects stem biomass, but not leaf biomass, suggests that leaves are more resistant to this stress than shoots, which may be a strategy to sustain higher photosynthetic activity. Indeed, we did not find significant differences in leaf morphology between water regime treatments, where all seedlings displayed leaves with similar area and thickness. However, while individual leaf area was similar between the two treatments (water regime and acorn size), there was a significant difference in the total leaf area between seedlings coming from small and those coming from large acorns with a greater value for the latter. An increase in the total leaf area allows seedlings to intercept a higher amount of light, improving consequently photosynthetic activity which could confirm our suggestion that seedlings coming from large seedlings can sustain a higher photosynthetic activity relative to those coming from small seeds.
The taproot showed more resistance to water stress than fine roots, since only fine roots were affected by water stress. Taproots constitute the main storage organ in oak seedlings (Grünzweig et al. 2008), which makes it, perhaps, richer in water than fine roots, and consequently more resistant to water stress. Overall, none of seed size treatment seemed to be more resistant than the other to water stress (water stress × seed size interaction was not significant), showing no role of seed size in the resistance of the future plant to this stress. This is in contradiction with our last hypothesis, which says that plant tolerance to water stress increases with increasing seed size.
Conclusion
Our study shows that cork oak acorns can germinate easily, regardless of acorn size. We did not find an effect of acorn size on seedling survival, indicating that small acorns contain enough cotyledon reserves to support seedling survival. Seed size did, however, plays a key role in initial seedling growth, which increased with increasing acorn size. This indicates the benefit of selecting large acorns for plantation programs to produce taller plants with greater above- and below-ground biomass, which promotes their competitive ability for light, water and nutrient resources, since benefits of acorn reserves are temporal. The role of metabolic resources stored in the cotyledons in seedling establishment has been well studied (Khan and Shankar 2001; Aizen and Woodcock 1996; Bonfil 1998). However, there was a lack of published information on the difference in cotyledon mobilization between seeds of different sizes. Our study showed a significant increase in mobilization of seed reserves as seed size increases. Finally, our results demonstrated that water stress adversely affects survival and seedling growth independent of seed size, indicating a lack of influence of acorn size on subsequent resistance of plant to water stress, at least under the conditions of our experiment.
References
Aizen MA, Woodcock H (1996) Effects of acorn size on seedling survival and growth in Quercus rubra following simulated spring freeze. Can J Bot 74:308–314
Bond WJ, Honing M, Maze KE (1999) Seed size and seedling emergence: an allometric relationship and some ecological implications. Oecologia 120:132–136
Bonfil C (1998) The effects of seed size, cotyledon reserves and herbivory on seedling survival and growth in Quercus rugosa and Q. laurina (Fagaceae). Am J Bot 85:79–87
Boussaïdi N (2012) Impacts de l’action anthropique sur la subéraie tunisienne: essai de projection dans le futur d’un écosystème (cas de la subéraie de Kroumirie – nord-ouest de la Tunisie). Thèse de doctorat, Institut National Agronomique de Tunisie, p 203
Chaar H, Mechergui T, Khouaja A, Abid H (2008) Effects of tree shelters and polyethylene mulch sheets on survival and growth of cork oak (Quercus suber L.) seedlings planted in North-western Tunisia. For Ecol Manage 256:722–731
Leskovar DI (1995) Seedling Morphological and Physiological Adaptation to Abiotic Stress: Introduction to the Colloquium. HortScience 30(6):1151–1159
Dupraz C (1997) Les Protections de plants à effet de serre. Première partie: ce qu'en pensent les arbres. Rev For Fr XLIX-5:417–432
Erniwati E (2006) The role of seed size in the re-sprouting ability of oak seedlings. Do larger seeds of Q. robur and Q. petraea have an advantage by saving more resource for re-sprouting? Master’s thesis, Wageningen University, Wageningen, Netherlands
Foster SA (1986) On the adaptive value of large seeds for tropical moist forest trees—a review and synthesis. Bot Rev 52:260–299
Giertych MJ, Suszka J (2011) Consequences of cutting off distal ends of cotyledons of Quercus robur acorns before sowing. Ann For Sci 68:433–442
Grünzweig JM, Carmel Y, Riov J, Sever N, McCreary DD, Flather CH (2008) Growth, resource storage, and adaptation to drought in California and eastern Mediterranean oak seedlings. Can J For Res 38:331–342
Hasnaoui B (1992) Chênaies du Nord de la Tunisie. Écologie et régénération. Thèse de doctorat d’État es Sciences Naturelles, Université de Provence, Aix-Marseille I, p 202.
Ja KM, Han LS, Young WS (2011) Cork oak (Quercus suber L.) forest decline in Tunisia: a linkage between physiological adaptation and stress. Sci Res Essays 6(6):1143–1146
Jacobs DF, Salifu KF, Davis AS (2009) Drought susceptibility and recovery of transplanted Quercus rubra seedlings in relation to root system morphology. Ann For Sci 66:1–12
Jurado E, Westoby M (1992) Seedling growth in relation to seed size among species of arid Australia. J Ecol 80:407–416
Karrfalt RP (2004) How acorn size influences seedling size and possible seed management choices. In: Riley IE (ed) National proceedings: forest and conservation nursery associations. USDA Forest Service, Washington DC, pp 117–118
Khan ML, Shankar UMA (2001) Effect of seed weight, light regime and substratum microsite on germination and seedling growth of Quercus semiserrata Roxb. Trop Ecol 42(1):117–125
Khurana E, Singh JS (2000) Influence of seed size on seedling growth of Albizia procera under different soil water levels. Ann Bot 86:1185–1192
Leishman MR, Wright IJ, Moles AT, Westoby M (2000) The evolutionary ecology of seed size. In: Fenner M (ed) Seeds: ecology of regeneration in plant communities. CAB International, Wallingford, pp 31–57
Liu Y, Liu G, Li Q, Liu Y, Hou L, Li G (2012) Influence of pericarp, cotyledon and inhibitory substances on sharp tooth oak (Quercus aliena var. acuteserrata) germination. PLoS ONE 7:e47682
Lloret F, Casanovas C, Peñuelas J (1999) Seedling survival of Mediterranean shrubland species in relation to root:shoot ratio, seed size and water and nitrogen use. Funct Ecol 13:210–216
Mechergui T, Pardos M (2017) Impacts of mulching and tree shelters on cork oak (Quercus suber L.) seedling survival and growth after four growing seasons. Rev Ecol (Terre et Vie) 72:410–424
Merouani H, Branco C, Almeida MH, Pereira JS (2001) Effects of acorn storage duration and parental tree on emergence and physiological status of Cork oak (Quercus suber L.) seedlings. Ann For Sci 58:543–554
Metcalfe DJ, Grubb PJ (1997) The response to shade of seedling of very small-seeded tree and shrub species from tropical rain forest in Singapore. Funct Ecol 11:215–221
Milberg P, Lamount BB (1997) Seed/Cotyledon size and nutrient content play a major role in early performance of species n nutrient-poor soil. New Phytol 137(4):665–2672
Norden N, Matthew ID, Antoine C, Gonzalea MA, Garwood NC, Chave J (2008) The relationship between seed mass and mean time to germination for 1037 tree species across fire tropical forests. Funct Ecol 23:203–210. https://doi.org/10.1111/j.1365-2435.2008.01477
Pandey R, Bargali K, Bargali SS (2017) Does Seed Size Affect Water Stress Tolerance in Quercus leucotrichophora A Camus at Germination and Early Seedling Growth Stage? Biodiversity Int J 1(1):24–30. https://doi.org/10.15406/bij.2017.01.00005
Perea R, San Miguel A, Gil L (2011) Leftovers in seed dispersal: ecological implications of partial seed consumption for oak regeneration. J Eco 199:194–201
Quero JL, Villar R, Maranon T, Zamor R, Poorter L (2007) Seed mass effects in four Mediterranean Quercus species (Fagacea) growing in contrasting light environments. Am J Bot 94:1795–1803
Selmi K (2006) Use of the data of the national forest inventory for the monitoring and the management of cork oak forest in Tunisia. Ann Inrgref 9:21–30
Sonesson LK (1994) Growth and survival after cotyledon removal in Quercus robur seedlings, grown in different natural soil types. Oikos 69:65–70
Steege HT, Bokdam C, Boland M, Dobbelsteen J, Verburg I (1994) The effects of man-made gaps on germination, early survival, and morphology, of Chlorocardium rodiei seedlings in Guyana. J Trop Ecol 10:245–260
Vieira Natividade J (1951) Subericultura., Direc ao Geral dos Sevi os Florestais e Aquicolas, Portugal.
Westoby M, Leishman M, Lord J (1996) Comparative ecology of seed size and dispersal. Philos Trans R Soc Lond B Biol Sci 351:1309–1318
Wright IJ, Westoby M (1999) Differences in seedling growth behaviour among species: trait correlations across species, and trait shifts along nutrient compared with rainfall gradients. J Ecol 87:85–97
Yi X, Zhang J, Wang Z (2015) Large and small acorns contribute equally to early-stage oak seedlings: a multiple species study. Eur J Forest Res 134:1019–1026. https://doi.org/10.1007/s10342-015-0906-y
Yang Y, Yi X (2012) Partial acorn consumption by small rodents: implication for regeneration of white oak, Quercus mongolica. Plant Ecol 213:197–205
Yi X, Liu C (2014) Retention of cotyledons is crucial for resprouting of clipped oak seedlings. Sci Rep 4:5145
Yi X, Yang Y (2010) Large acorns benefit seedling recruitment by satiating weevil larvae in Quercus aliena. Plant Ecol 209:291–300
Zobel RW (2005) Tertiary root systems. In: Zobel RW, Wright SF (eds) Roots and soil management: interactions between roots and the soil. American Society of Agronomy, Madison, pp 35–56
Zolfaghari R, Fayyaz P, Nazari M, Valladares F (2013) Interactive effects of seed size and drought stress on growth and allocation of Quercus brantii Lindl. seedlings from two provenances. Turk J Agric For 37:361–368
Acknowledgements
We would like to thank the research station and the direction of forests in the district of Sejnane for their assistance in all phases of this research project, including soil and pots supply. Special thanks to Amor Houatmia for helping in the setup of the experiment.
Funding
This study was funded by the direction of forests in the district of Sejnane.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Oliver Gailing.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Mechergui, T., Pardos, M. & Jacobs, D.F. Effect of acorn size on survival and growth of Quercus suber L. seedlings under water stress. Eur J Forest Res 140, 175–186 (2021). https://doi.org/10.1007/s10342-020-01323-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-020-01323-2