
Abstract
Water use efficiency (WUE) was compared in three upland South Moravian forested microwatersheds in the light of effects of global climate change on forest ecosystems (GCC). The experimental catchments were characterized as upland headwater forested microwatersheds of similar size and morphology and silvicultural system, but each with different dominant tree species in the stands (over 50% of forest stand composition in living stock): Norway spruce, European beech and mixed forest. WUE was evaluated according to mean daily streamflow reduction, measured at the discharge points of the recipients of the individual catchments in precipitation-free periods lasting more than 5 days. During these times, streamflow dynamics are mainly influenced by evapotranspiration processes occurring in the forest stands. Four precipitation-free periods were observed, two in the middle of the growing season and two at its end. Two of these periods were long (15 days or more), and two were shorter (6 days). The results indicated that WUE of upland forested catchments can be very different, depending upon the dominant tree species and the seasonal phase. Highest WUE at the catchment scale (never decreasing below 80%) was exhibited by beech predominating site. WUE of mixed forest was high as well, never decreasing below 69%. The lowest WUE was exhibited by spruce predominating site, especially during a long precipitation-free period in the summer where it decreased down to 39%. In the context of the landscape, upland microwatersheds with pure spruce stands could cause its accelerated dry out in the summer and pose a significant threat to sustainable water and forest management of these areas. In comparison, mixed forests stands where spruce is not the dominant species or beech stands should still be a viable option even under the effects of GCC.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Forest ecosystems are currently affected by global climate change (GCC), and there are projections of changes in amount and distribution of precipitation across the whole of Europe (Bates et al. 2008). In Central Europe, a significant decrease in summer precipitation is to be expected (Trenberth et al. 2003; IPCC 2013; Dubrovský et al. 2015), as well as a lower frequency and duration of precipitation periods, which could lead to longer and more frequent precipitation-free periods (IPCC 2013). GCC impacts not only the forest ecosystems but also the length of the growing season, increasing the latter in the middle latitudes (Frich et al. 2002) with projected higher minimum daily temperatures and reduced number of frost days. During the last 20 years, the length of the growing season in Northern and Central Europe increased on average by 10–20 days, mostly because of the earlier arrival of spring (Linderholm 2006; Vitasse et al. 2011; Kolář et al. 2016). These changes pose a potential risk to forest ecosystems, as trees become more exposed to frost, which can cause damages and the dieback of entire forest stands (Bokhorst et al. 2008). Among the expected reactions of forest ecosystems to GCC are wide changes to their species composition, with a range of overlapping effects on their hydrological regime (Adams et al. 2009; Troch et al. 2009). In the managed forests of Central Europe, where forest stands are often homogeneous in stand tree species composition with specific dominant tree species, one of the key issues in altering the negative impacts of GCC is a proper understanding of individual tree species and how they can affect water use efficiency (WUE) of whole catchments (Richter et al. 2006; Arthington et al. 2010). WUE at the catchment scale was considered as the ability of forest stands and the dominant tree species to maintain balanced streamflow even during precipitation-free periods or the catchment potential to limit water losses during dry periods of the growing season and thus sustain balanced streamflow. This paper addresses the evaluation of WUE in three South Moravian upland forested microwatersheds under these conditions. The catchments were chosen to represent upland headwater microwatersheds of similar size and morphology, with forest stands dominated by Norway spruce (Picea abies L.), European beech (Fagus sylvatica L.) or mixed forest as the typical forest stands in the region (Pretzsch et al. 2014).
Forest vegetation exhibits the highest WUE in times of limited water resources (Troch et al. 2009; Huxman et al. 2004). During sunny warm days in forested areas, forest transpiration reaches more than 90% of total evapotranspiration (Grelle et al. 1997; Schlesinger and Jasechko 2014; Deutscher et al. 2016; Wang et al. 2017). Therefore, the streamflow dynamics were analyzed during precipitation-free periods of the growing season, when the effect of the vegetation on the quantity of water in the recipient channel was greatest and could be best observed (Graham et al. 2013; Kovář and Bačinová 2015).
Materials and methods
The three experimental catchments are situated in the Training Forest Enterprise Masaryk Forest Křtiny (TFE), a forest enterprise of Mendel University in Brno. It is located in the South Moravian region (SMR) of the Czech Republic (CR) (Fig. 1). It is situated in the uplands of the Bohemian Massif (Demek 1987). The experimental catchments are located on the granodiorite complexes of Brno Massif, consisting mainly of granodiorites and, to a lesser extent, acidic granodiorites (Bajer 2015). The whole region can be characterized as a mild warm climate (Quitt 2009), with a mean annual temperature of 7.5 °C and mean annual precipitation of 610 mm. The natural vegetation consists of deciduous forests of the temperate zone with dominant European beech (Fagus Sylvatica L.), with an admixture of Sessile oak (Quercus petraea Matusch), European hornbeam (Carpinus betulus L.) and Silver fir (Abies alba Mill.). Currently, the species composition of managed forests has changed in favor of coniferous species such as Norway spruce (Picea abies L.), Scotch pine (Pinus sylvestris L.) and European larch (Larix decidua Mill.) (Ministry of Agriculture of the Czech Republic 2013).

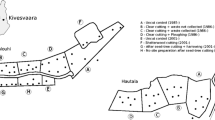
Location of the experimental catchments—Útěchov (beech), Kanice (mixed forest) and Křtiny (spruce) including digital terrain models and age distribution according to Forest Spatial Alignment maps (big bold numbers identify forest compartments; within those, capital bold letters identify forest stands; within those, big numbers identify forest units that also describe age in 10 year intervals, for example, 7 indicates age of 71–80 years)
The experimental catchments were named according to the closest municipalities: Křtiny, Kanice and Útěchov. They were selected as close to each other as possible (less than 10 km) according to three main attributes: area of approximately 50 ha; similar geomorphology of the Bohemian Massif; and different predominant tree species in the forest stands. They are all headwater microwatersheds, with an upslope surface headspring, without contribution from deep hydrogeological aquifers. The relative terrain altitude difference reaches 30–150 m. The dominant tree species are Norway spruce, European beech and mixed forest in the microwatersheds of Křtiny, Útěchov and Kanice (further referred to as spruce, beech and mixed), respectively (Table 1). In spruce, a considerable admixture of beech is present; however, it is scattered across the catchment area (in 60% of forest units with beech the proportion of beech is less than 50% in living stock), whereas spruce can be mostly found in pure stands (in 70% of forest units with spruce the proportion of spruce is more than 75%) and therefore it was considered the dominant species (Lesprojekt Brno 2013). All the stands are even-aged, with the origin from a combination of both artificial and natural regeneration. The same silvicultural system—small-scale clearcutting (0.5–1 ha) and shelterwood cutting—is applied to all catchments. The age structure of the predominant tree species is similar in beech and spruce where the highest living stock can be found in young (up to 40 years) and mature stands while an even distribution of age classes can be found in mixed (Fig. 2).

Age structure of the predominant species in the experimental catchments. Age classes are presented in 10-year intervals
Selected catchment areas were observed from 1.7. to 31.10. 2015. WUE was evaluated by a trend analysis of daily streamflow reduction, measured at the discharge points of individual microwatersheds. The methodology consisted of two basic steps:
-
1.
Measurements of air temperature, precipitation and streamflow.
-
2.
Data processing—identification of precipitation-free periods, trend analysis.
Precipitation (mm) and temperature (°C) were obtained from a climatic station with a 15-minute writing interval (AMET, Velké Bílovice, CR) situated in a clear and fenced site 532 m above sea level (GPS 49°19′06.9′′N, 16°43′49.2′′E; Fig. 1) locally called Dyk′s Enterprise. All studied catchments are located within 10 km distance, so a single station was used for all three forests. Long-term climatic data for SMR and CR were obtained from the Czech Hydro-meteorological Institute (portal.chmi.cz). The streamflow was measured at the discharge points of the individual catchments by measuring systems, consisting of a Thomson weir with a pressure water-level sensor (TSH22-3-1), connected to a datalogger (Hydro Logger H40D, both Fiedler Mágr, České Budějovice, CR). The water-level values were automatically converted to streamflow in the datalogger every 15 min by a preset rating curve for the Thomson weir (Deutscher and Kupec 2014).
Mean daily temperatures and daily sums of precipitation were established from measured climatic data (Fig. 3). Periods with at least five consecutive days without precipitation were identified. The measured streamflow was used to create a hydrograph for the study period from mean hourly streamflow values (Fig. 4). The beginning of the precipitation-free periods was then tracked back to the point where the balanced streamflow (base flow) was reached, after the last antecedent precipitation event, generally 1 day afterward. The very beginning of each precipitation-free period was defined by the maximum morning streamflow (Deutscher and Kupec 2014) reached between 8:00 and 9:00 of the first day of balanced streamflow. Its end was defined as the same hour on the last day without precipitation. In this way, a total of four precipitation-free periods were identified during the studied period (Figs. 3, 4).

Climagraph for the study period, arrows indicating the observed precipitation-free periods

Hydrograph for the study period, arrows indicating the observed precipitation-free periods
The basic characteristics of each individual precipitation-free period were obtained to enable their description and comparison (Table 2). During these periods, the evaluation of WUE of individual catchments was carried out. This was based on a trend analysis of daily streamflow reduction during each period and in each locality, twelve times in total. Basic linear regression was used to identify the trends and their overall accuracy (R squared) and its statistical significance was tested by the t test and its nominal p value to avoid bias by autocorrelation (p value 0.05 was considered a significant trend) (example—Fig. 5). The daily streamflow reduction was then standardized to the maximum streamflow in the morning of the first day of the period. WUE was then calculated from the mean daily reduction of all the days within the period subtracted from 100%.

An example of linear regression model used to describe the trend in daily streamflow reduction (locality Kanice mixed, period 1). Black squares indicate mean daily streamflow values interlaced by the linear trend line. Such regression was made for each locality and each period, 12 in total (R2 and nominal p values are shown in Table 3)
Results
Local climate in 2015
Mean temperature measured at Dyk’s Enterprise over the study period (July–October) reached 16.6 and 17.0 and 15.6 °C in SMR and CR, respectively, while the mean long-term average (ML-TA) 1961–1990 and 1981–2010 in SMR for this period reaches 14.6 and 15.3 °C, respectively, which indicates increasing temperature in the region but also that the local climate was even warmer (Fig. 6).

Mean temperature in the observed period from July to October 2015 in different spatial scales (local, South Moravian Region, Czech Republic) as compared to the mean long-term averages 1961–1990 and 1981–2010 for South Moravian Region (ML-TA SMR)
The total precipitation during the same period measured at Dyk′s Enterprise reached 193 and 207 and 187 mm in SMR and CR, respectively. ML-TA precipitation 1961–1990 and 1981–2010 in SMR for this period reaches 200 and 223 mm, respectively, which is close to measured precipitation in 2015. However, the total precipitation for the whole year in SMR reached 430 mm (77% of ML-TA 1981–2010) and only 270 mm at Dyk’s Enterprise, of which more than 60% (193 mm) fell during the study period (Fig. 7). The ratio of monthly precipitation and its distribution was the most extreme during August and September. During August, precipitation reached 69.6 mm all of which fell during 5 days. During September, precipitation reached 20.2 mm spread throughout 10 days. A total of four precipitation-free periods were observed, two of which occurred in the middle of the growing season in August, and two during its end, in September and November. Two were evaluated as long (15 days and more) and two as short (6 days) (Table 2).

Total precipitation for the observed period from July to October 2015 in different spatial scales (local, South Moravian Region, Czech Republic) as compared to the mean long-term averages 1961–1990 and 1981–2010 for South Moravian Region (ML-TA SMR)
Trend analysis of streamflow during observed precipitation-free periods in experimental catchments
The streamflow over the entire study period is presented in a hydrograph of mean daily values (Fig. 4). The mean streamflow during the study period reached 0.50, 0.39 and 0.44 l/s in spruce, beech and mixed, respectively. During the precipitation-free periods, a decreasing trend in streamflow was identified reaching on average 0.034, 0.006 and 0.014 l/s/day in spruce, beech and mixed, respectively (Table 3). In most of the cases, the trend was significant with the exceptions of the two last precipitation-free periods at the end of September and November in beech and the long precipitation-free period at the end of September in spruce (see p value 0.06; 0.71 and 0.76 in Table 3). Basic linear trend was sufficient to explain more than 50% of the variability of the decrease in most of the cases with the same exceptions as above (see R squared value 0.24; 0.01 and 0.03 in Table 3).
Period 1 (31.7.–16.8.) lasted for 16 days with six antecedent days of rainfall with a mean intensity of 5 mm/day. The period occurred in the middle of the growing season with the mean daily temperature reaching 25.1 °C, with mean daily maximum and minimum temperature reaching 31.9 and 17.1 °C, respectively. The trend in mean daily streamflow decrease was significant in all catchments, but the lowest WUE of 39% was exhibited by spruce. In contrast, WUE in beech and mixed reached 79 and 69%, respectively (Fig. 8 and Table 4).

WUE at the catchment scale according to the trend analysis of streamflow during observed precipitation-free periods in experimental catchments based on mean daily streamflow reduction, which was standardized to the initial maximum streamflow in the morning of the first day of the period
Period 2 (27.8.–1.9.) was short (lasting 6 days), with relatively high antecedent precipitation lasting 10 days, with the mean intensity approaching 7 mm/day. The period occurred in the middle of the growing season with the mean daily temperature reaching 23.7 °C, with mean daily maximum and minimum temperature reaching 29.7 and 15.9 °C, respectively. The trend in mean daily streamflow decrease was linear in all of the catchments. However, the highest WUE of 90% was again exhibited by beech. In mixed and spruce, WUE reached 87 and 79%, respectively (Fig. 8 and Table 4).
Period 3 (20.9.–5.10.) was long (lasting 15 days), with low antecedent precipitation lasting 6 days, with a mean intensity of under 1 mm/day. This period occurred during the second half of September at the end of the growing season, with mean daily temperatures approaching 12 °C, with mean daily maximum and minimum temperature reaching 16.0 and 6.3 °C, respectively. The trend in daily streamflow decrease was not significant in beech and to a lesser extent in spruce as during the end of the period, streamflow seemed to slowly increase, which was probably caused by decreasing temperature causing limited transpiration. WUE of all catchments was remarkably similar, reaching 81–84% in all three forests (Fig. 8 and Table 4).
Period 4 (22.10.–28.10) was short (lasting 6 days), with antecedent precipitation lasting 8 days, with the mean intensity reaching 6.3 mm/day. This period occurred during the end of the growing season, with the mean daily temperature below 8 °C, with mean daily maximum and minimum temperature reaching 12.0 and 3.6 °C, respectively. The trend in daily streamflow decrease was again not significant in beech and seemed to follow no trend at all. This was possibly as a result of an interaction of several factors, most importantly short duration of the period, high and intensive antecedent precipitation, low temperature inhibiting transpiration in forest stands dominated by broadleaves as well as the seasonal phase. During this period, WUE in beech and mixed forest reached 99 and 94%, respectively, while spruce exhibited WUE of 72% (Fig. 8 and Table 4).
Discussion
Our results confirmed the differences in the current state of WUE of three managed South Moravian upland forest microwatersheds, based on their dominant tree species (spruce, beech and mixed forest). Spruce catchment exhibited a significant decrease in WUE during a long precipitation-free period in the summer (down to 39%). In connection to GCC, the overall stability and health status of forests stands as well as their expected hydrological effects on the landscape are at premium importance. Low WUE of upland microwatersheds dominated by spruce poses a significant threat to future existence of forests.
WUE was evaluated in a novel way, based on a trend analysis of mean daily streamflow reduction occurring during precipitation-free periods, standardized to the maximum initial streamflow at its beginning. For a clearer interpretation of our results, it is important to distinguish between plant water use efficiency (P-WUE) for individual tree species and the water use efficiency of the entire microwatershed at the catchment level (C-WUE). The higher the P-WUE, the less water is needed by the plant to sustain its growth (Binkley et al. 2004). In relation to C-WUE, as viewed in this paper, as the ability to minimize water losses during precipitation-free periods, it is the very ability of plants to increase their P-WUE under these specific conditions (Troch et al. 2009) that is essential to sustaining a high C-WUE while limiting water uptake at the same time. It is known that plants usually exhibit their highest potential P-WUE during periods of low water availability (Huxman et al. 2004). However, there is no direct correlation between P-WUE and C-WUE, as it is the water uptake that is the most important factor in altering C-WUE, not just the P-WUE of the dominant species.
Our results indicate that WUE of upland catchments in SMR dominated by pure spruce stands could be limited, especially during the longer precipitation-free periods in the middle of the growing season, resulting in a depletion of water resources. This contrasts with alpine regions, where spruce stands are usually perceived as a species with a positive effect on C-WUE, where in combination with higher annual precipitation and lower temperature they effectively function as a source of water during dry parts of the year (Švihla 2001; Šach et al. 2014). At the same time, an interesting correlation between WUE and the seasonal phase was found in the beech catchment (according to the comparison of first two and the last two precipitation-free periods—see Fig. 8). While in the mixed forest and in spruce, WUE remained more or less the same indicating no seasonality, there was a big difference in beech between the middle of the growing season and its end when the beech stands seemed to influence WUE little without significant trend in streamflow decrease (Table 3). This could be explained by the fact that the end of the growing season can be associated with decreasing daily temperatures and phenological changes (such as leaf coloring and leave fall) resulting in limited transpiration which in these altitudes usually occurs at the end of October in beech (Schieber et al. 2013) as compared to spruce where the phenophases are more reliant on the length of the photoperiod and usually happen later in the year (Kolářová et al. 2013). Using this knowledge about the response of forest stands dominated by different tree species throughout the year in forest planning and management could serve as an important tool in mitigating the negative effects of GCC (Graham et al. 2013; Kovář and Bačinová 2015).
Local climate during the year 2015 was different to the recent mean long-term average 1981–2010 and exhibited most of the profound changes connected to GCC (Trenberth et al. 2003; IPCC 2013; Dubrovský et al. 2015). Under the effects of GCC, an important question arises regarding how these changes would affect WUE in the future. It is expected that P-WUE would decrease due to increasing concentrations of CO2 in the atmosphere, which limit transpiration and stomata conductivity (Hasper et al. 2016). This would lead to higher discharge from forested catchments, resulting in a faster depletion of water resources during dry periods and therefore a loss of C-WUE (Beerling 1999). At the same time, the increase in temperature and prolonging of the growing season is expected to cause an increase in evapotranspiration and would result in the reduction of discharge from these areas. However, there are some results that indicate these two reverse effects can cancel each other out, without significant change to WUE at all (Hasper et al. 2016).
Conclusions
Highest WUE at the catchment scale was exhibited by the beech catchment Útěchov. Even during the long precipitation-free periods in any part of the year, WUE never decreased below 80%. WUE of the mixed forest catchment Kanice was high as well, never decreasing below 69%. The spruce catchment Křtiny exhibited the lowest WUE, a significant reduction in WUE was observed during the long precipitation-free period in the summer, where it decreased down to 39%.
In the summer, WUE of forested upland catchments during precipitation-free periods was very different depending on the dominant tree species. The results also indicated that during long precipitation-free periods (more than 15 days) in the autumn, WUE of the catchments dominated by beech and spruce was similar (81 and 83%, respectively), while in the summer, WUE of the spruce catchment reached only half of that of beech (39 and 79%, respectively). This dramatic reduction in WUE in the spruce catchment indicates that a significant depletion of water resources occurred.
These results indicate that from the perspective of forest and water management of upland areas, silviculture of pure spruce stands will be more and more challenging in the near future. Given the current effects of GCC such as longer and more frequent precipitation-free periods in the summer (IPCC 2013; Dubrovský et al. 2015), upland spruce stands will be heavily stressed by the lack of water (according to our results WUE decreased down to 39% in spruce stands during long summer precipitation-free period, even though they were able to tolerate short dry periods). In the context of the landscape, upland microwatersheds dominated by spruce could cause its accelerated dry out in the summer. In comparison, silviculture of mixed forests stands where spruce is not the dominant species (in our case less than 30% spruce) should still be a viable option for forest management (according to our results their WUE was not affected as negatively even during the long dry precipitation-free periods decreasing only down to 79%). At the same time under the described conditions, beech stands should be able to resist the aforementioned changes in summer precipitation without negative influences.
References
Adams HD, Guardiola-Claramonte M, Barron-Gafford GA, Villegas JC, Breshears DD, Zou CB, Troch PA, Huxman TE (2009) Temperature sensitivity of drought-induced tree mortality: implications for regional die-off under global-change-type drought. Proc Natl Acad Sci USA 106(17):7063–7066
Arthington AH, Naiman RJ, Mcclain ME, Nilsson C (2010) Preserving the biodiversity and ecological services of rivers: new challenges and research opportunities. Freshw Biol 55:1–16
Bajer A (2015) a kol. Krajina a geodiverzita: neživá příroda jako základ krajinných a kulturních hodnot. Brno: Mendelova univerzita v Brně. 159 s. ISBN 978-80-87443-11-8
Bates BC, Kundzewicz ZW, Wu S, Palutikof JP (eds) (2008) Climate change and water. Technical paper of the intergovernmental panel on climate change. IPCC Sec-retariat, Geneva. Accessed July 10, 2013. www.ipcc.ch/pdf/tec
Beerling DJ (1999) Long-term responses of boreal vegetation to global change: an experimental and modelling investigation. Glob Change Biol 5:55–74
Binkley D, Stape JL, Ryan MG (2004) Thinking about efficiency of resource use in forests. For Ecol Manag 193:5–16. https://doi.org/10.1016/j.foreco.2004.01.019
Bokhorst S, Bjerke JW, Bowles FW, Melillo J, Callaghan TV, Phoenix GK (2008) Impacts of extreme winter warming in the sub-Arctic: growing season responses of dwarf shrub heathland. Glob Change Biol 14:2603–2612. https://doi.org/10.1111/j.1365-2486.2008.01689.x
Demek J (1987) Obecná geomorfologie. Praha: Academia, 476 s
Deutscher J, Kupec P (2014) Monitoring and validating the temporal dynamics of interday streamflow from two upland head microwatersheds with different vegetative conditions during dry periods of the growing season in the Bohemian Massif, Czech Republic. Environ Monit Assess. https://doi.org/10.1007/s10661-014-3661-5
Deutscher J, Kupec P, Dundek P, Holík L, Machala M, Urban J (2016) Diurnal dynamics of streamflow in an upland forested micro-watershed during short precipitation-free periods is altered by tree sap flow. Hydrol Process 30(13):2042–2049. https://doi.org/10.1002/hyp.10771
Dubrovský M, Trnka M, Holman IP, Svobodová E, Harrison PA (2015) Developing a reduced-form ensemble of climate change scenarios for Europe and its application to selected impact indicators. Clim Change 128:169–186. https://doi.org/10.1007/s10584-014-1297-7
Frich P, Alexander LV, Della-Marta P, Gleason B, Haylock M, Klein Tank AMG, Peterson T (2002) Observed coherent changes in climatic extremes during the second half of the 20th century. Climate Res 19:193–212
Graham CB, Barnard HR, Kavanagh KL, McNamara JP (2013) Catchment scale controls the temporal connection of transpiration and diel fluctuations in streamflow. Hydrol Process 27:2541–2556. https://doi.org/10.1002/Hyp.9334
Grelle A, Lundberg A, Lindroth A, As Moren, Cienciala E (1997) Evaporation components of a boreal forest: variations during the growing season. J Hydrol 197:70–87
Hasper TB, Wallin G, Lamba S, Hall M, Jaramillo F, Laudon H, Linder S, Medhurst JL, Rantfors M, Sigurdsson BD, Uddling J (2016) Water use by Swedish boreal forests in a changing climate. Funct Ecol 2016(30):690–699. https://doi.org/10.1111/1365-2435.12546
Huxman TE, Smith MD, Fay PA, Knapp AK, Shaw MR, Loik ME et al (2004) Convergence across biomes to a common rain-use efficiency. Nature 429:651–654
IPCC (2013) Climate change 2007: the physical science basis. Technical report. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Mille HL (eds) Conntribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Ambridge University Press, Cambridge, p 996
Kolář T, Giagli K, Trnka M, Bednářová E, Vavrčík H, Rybníček M (2016) Response of the leaf phenology and tree-ring width of European beech to climate variability. Silva Fennica 50(2):1520 ISSN: 00375330
Kolářová E, Nekovář J, Adamík P (2013) Long-term temporal changes in central European tree phenology (1946–2010) confirm the recent extension of growing seasons. Int J Biometeorol. https://doi.org/10.1007/s00484-013-0779-z
Kovář P, Bačinová H (2015) Impact of evapotranspiration on diurnal discharge fluctuation determined by the Fourier series model in dry periods. Soil Water Res 10(4):210–217
Lesprojekt Brno AS (2013) Forest Management Plan of the Training forest Enterprise Masaryk Forest Křtiny 2013–2022
Linderholm HW (2006) Growing season changes in the last century. Agric For Meteorol 137(2006):1–14
Ministry of Agriculture of the Czech Republic (2013) Information on forests and forestry in the Czech Republic by 2012. Prague 2013, p 35. www.eagri.cz
Pretzsch H, Biber P, Schütze G, Uhl E, Rötzer T (2014) Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat Commun. https://doi.org/10.1038/ncomms5967
Quitt E (2009) Klimatické oblasti (1901–2000). Barevná mapa č. 38, 1: 1 000 000. In: Hrnčiarová T., Mackovčin P., Zvara I.: Atlas krajiny ČR. Ministerstvo životního prostředí ČR a Ústav Silva Taroucy pro krajinu a okrasné zahradnictví, Praha
Richter BD, Warner AT, Meyer JL, Lutz K (2006) Acollaborative and adaptive process for developing environmental flow recommendations. River Res Appl 22:297–318
Šach F, Švihla V, Černohous V, Kantor P (2014) Management of mountain forests in the hydrology of a landscape, the Czech Republic. J For Sci 60(1):42–50 (ISSN 1212-4834)
Schieber B, Janík R, Snopková Z (2013) Phenology of common beech (Fagus sylvatica L.) along the altitudinal gradient in Slovak Republic (Inner Western Carpathians). J For Sci 59(4):176–184
Schlesinger WH, Jasechko S (2014) Transpiration in the global water cycle. Agric For Meteorol 189–190:117–155
Švihla V (2001) Vliv lesa na odtokové poměry namalém povodí. Lesnická práce 2. http://www.silvarium.cz/lesnicka-prace-c-2-01/vliv-lesa-naodtokove-pomery-na-malem-povodi. Accessed Feb 5, 2014
Trenberth KE, Dai A, Rasmussen RM, Parsons DB (2003) The changing character of precipitation. Bull Am Meteor Soc 84:1205–1217
Troch PA, Martinez GF, Pauwels VRN, Durcik M, Sivapalan M, Harman C, Brooks PD, Gupta H, Huxman T (2009) Climate and vegetation water use efficiency at catchment scales. Hydrol Process 23:2409–2414
Vitasse Y, François C, Delpierre N, Dufrêne E, Kremer A, Chuine I, Delzon S (2011) Assessing the effects of climate change on the phenology of European temperate trees. Agric For Meteorol 151(7):969–980. https://doi.org/10.1016/j.agrformet.2011.03.003
Wang H, Tetzlaff D, Dick JJ, Soulsby Ch (2017) Assessing the environmental controls on Scots pine transpiration and the implications for water partitioning in a boreal headwater catchment. Agric For Meteorol 240–241(2017):58–66
Acknowledgments
Supported by the Project FRAMEADAPT Frameworks and possibilities of forest adaptation measures and strategies connected with climate change EHP-CZ02-OV-1-019-2014 and by Internal Grant Agency of the Faculty of Forestry and Wood Technology (IGA LDF), MENDELU in Brno as Project 17/2016.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Agustín Merino.
Rights and permissions
About this article

Cite this article
Petr, K., Libor, Š. & Jan, D. Tree species composition influences differences in water use efficiency of upland forested microwatersheds. Eur J Forest Res 137, 477–487 (2018). https://doi.org/10.1007/s10342-018-1117-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-018-1117-0