
Abstract
In the analysis of old-growth forest dynamics, the continuous process of tree aging and forest structural change is split up into several distinct forest development stages. The criteria for distinguishing the stages vary among the different approaches. In most of them, vertical canopy heterogeneity is only coarsely addressed and horizontal forest structure is quantified at spatial scales far exceeding the size of conventional forest inventory plots. In order to describe and analyze the complex mosaic structure of temperate old-growth forests with objective and quantitative measures in the context of forest inventories, we propose the Development Stage Index I DS . It employs two easily measured stand structural parameters (stem density and basal area) for quantifying the abundance of trees in three conventionally recognized tree diameter classes (premature < 40 cm; mature 40–70 cm; and over-mature ≥ 70 cm) in plots of 500 m2 size, systematically distributed in the forest. This allows quantifying the spatial extension of the Initial, Optimum and Terminal stages of forest development at plot, stand and landscape levels. Based on thorough stand structural analyses in three virgin beech (Fagus sylvatica) forests in Slovakia, we demonstrate that I DS is a promising tool for (1) quantifying the proportion of the three stages on different scales, (2) visualizing the complex mixing of stages, and (3) analyzing dynamic changes in old-growth forest structure. We conclude that the Development Stage Index has the potential to improve the empirical foundation of forest dynamics research and to allow this discipline to proceed to more rigorous hypothesis testing.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The forest development cycle is a theoretical concept to describe the structural dynamics of natural forests driven by demographic processes and the action of external disturbances, which repeats itself with the formation of a new generation (Watt 1947; Remmert 1991). Forest dynamics processes can take place on different spatial scales, from the single tree to the stand level (Oliver and Larson 1996). What spatial scale is the most appropriate for study depends on the disturbance regime and tree species composition, both of which vary with climate and soil conditions (Peters 1997; Král et al. 2010a). North American conceptual models generally concentrate on forest succession following relatively frequent large-scale disturbances that peak in a steady-state or dynamic equilibrium (which can last for quite long) before another disturbance resets the system (Franklin et al. 2002). In Europe, where large-scale disturbances are less frequent and forests are often formed by late-successional tree species, concepts were developed to describe the spatial pattern and processes in the anticipated status of dynamic equilibrium at the stand level. Král et al. (2016) found the European conceptual model to be applicable to late-successional forests in North America as well, indicating some similarities in structural dynamics.
For describing the temporal dynamics of stand development in old-growth forests, most researchers split up the continuous process of tree aging and forest structural change into distinct forest development stages, which sometimes were further divided into different development phases. Verbal descriptions of development stages or phases in European temperate forests are given by, e.g., Leibundgut (1993), Korpel (1995), Meyer (1999), Tabaku (2000) and Král et al. (2010b). The categories were traditionally defined by expert decision. They were used to map the occurrence of different development stages or phases in old-growth forest landscapes in order to quantify the horizontal variability of forest structure and to draw conclusions on the driving forces of change (e.g., Neumann 1979; Leibundgut 1993; Korpel 1995). Even though the distinction of development stages is a subjective process, it may allow comparing stand-level dynamics across different forest communities and biomes, if the criteria for stage identification are sufficiently comparable across studies.
More recently, methods based on empirical stand structural data have been developed, which help to make the distinction of forest development stages or phases more objective and repeatable (Meyer 1999; Tabaku 2000; Emborg et al. 2000; Grassi et al. 2003; Král et al. 2010b, 2016). While the required amount and quality of data differ, all such approaches allow the unambiguous assignment of forest plots to certain development stages or phases.
The information quality of stand structural data largely depends on the chosen spatial scale, and this does also apply to the outcome of development stage categorizations (Commarmot et al. 2005; Král et al. 2010a; Winter and Brambach 2011; Zenner et al. 2014). The smallest possible unit is a single tree with its ontogenetic development and associated change in height and stem diameter. Information on these tree dimensional data has frequently been used in the study of natural forest dynamics (Emborg et al. 2000; Grassi et al. 2003; Král et al. 2010b, 2016; Peck et al. 2015; Peterken 1996; Tabaku 2000; Winter and Brambach 2011; Zenner et al. 2016). All the aforementioned approaches are based on selected structural features; among them are the diameter or height of the largest trees, canopy cover, regeneration cover, or the amount of deadwood. The identification of development stages is mostly done on the plot level (156.25–500 m2) by determining which structural feature seems to have the strongest indicative value, while ignoring less obvious structural properties. This puts strong emphasis on a single ‘structural master factor’ in the classification process, while information about other stand characteristics is lost. Thus, these approaches implicitly assume structural uniformity at the chosen spatial scale (Pretzsch 2009), while heterogeneity in stand structure in a patch as a characteristic of primeval forests is largely ignored. One consequence of such approaches is that end-life stages tend to be mapped more often than early ones (e.g., Zenner et al. 2016), and the forest appears more homogeneous than it really is. Further, Christensen et al. (2007) showed that the focus on only the dominant structural elements can result in misleading conclusions on how forest structure changes with time. In reality, the tree individuals in a patch of old-growth forest often differ in age and size and represent different ontogenetic phases, even in small plots of only 156.25 m2 size. Thus, stages or phases might intermingle horizontally as well as vertically even in forest patches not exceeding the size of one or two trees. Korpel (1995) also observed that different stages and phases frequently overlapped in space and time in Slovakian virgin forests, and only his Optimum stage was found to occur in more or less pure form. Similar observations were made by Grassi et al. (2003) and Drößler and Meyer (2006). Paluch (2007) emphasized the spatially highly variable vertical stratification in the canopy of a natural beech–fir forest.
Thus, it seems desirable to advance the structural classification approach toward a concept that views tree populations as the sum of all tree individuals and allows conclusions on the development stage which dominates the patch, but provides information on subdominant structures as well. Further progress would be achieved, if the approach facilitated the assessment of development stages on different spatial scales. This would allow the application on various data sets including standardized forest structure inventories, which are often conducted in plots of 500-1000 m2 size that are placed randomly or systematically in the stand (e.g., Commarmot et al. 2013; Meyer et al. 2001). To do so would further restrict the input data to parameters that are widely available.
Here, we propose a method for quantifying the extension of three commonly recognized forest development stages (‘Initial’, ‘Optimum’ and ‘Terminal’) on the plot level in temperate old-growth forests using empirical data on tree size and stem density. We select true virgin forests (sensu Hunter 1990) of Fagus sylvatica L. in the western Carpathians as test systems. The proposed approach aims at defining forest development stages by objective criteria, allowing to compare the dynamics of different forest stands and communities and providing a solid database for the multivariate analysis of stand structure. As criteria for development stage distinction, we choose easy-to-measure parameters and express them in relative terms in order to account for large structural differences between forest communities and biomes. The approach is therefore not restricted to our test systems, but could be adapted for use in other forest types with different structure and environmental conditions as well.
Methods
Study sites
Since our study addresses the dynamics of natural forests, and management activities often imprint on forest structure for centuries (e.g., Tabaku 2000), we selected true virgin forests for study, for which no forest management is known for the past 500 years or so. We studied three beech-dominated (F. sylvatica) forests at montane elevation in the Carpathians of eastern Slovakia which are protected as National Nature Reserves; two of them are listed in UNESCO’s World Heritage. They represent some of the last remnants of temperate broad-leaved virgin forest in the western Carpathians, where they still covered extended areas in the early twentieth century. While the reserves Havešová (HA) and Kyjov (KY) are pure beech stands, Stužica (ST) contains a considerable proportion (11% by stem numbers) of silver fir (Abies alba). However, all three stands belong to the Fagetum dentarietosum glandulosae forest community (Bohn et al. 2003). The physiography of the three forests is summarized in Table 1. Some basic stand structural characteristics are given in Table 3.
Data collection
In every stand, we installed 40 circular plots of 500 m2 size (25.24 m in diameter) arranged on the nodes of rectangular grids. The grid spacing was 65 m in Kyjov, 100 m in Stužica and 140 m in Havešová. Different spacings had to be chosen so that the 40 plots could be accommodated within each of the stands and no plot was within a 100-m distance to the nearest reserve border. As ST is comparably large and hosts different forest types (Korpel 1995), we concentrated our study on an area of approximately 70 ha in the southeast of the reserve. Two of the 40 plots in ST close to the stand edge were subsequently excluded from the analysis, as it turned out that they may have been affected by forest management activities in the early twentieth century. In every plot, we mapped all living and dead trees with diameter at breast height (dbh) ≥ 7 cm and recorded their dbh. Lying deadwood was also measured if the stump was located in the plot. Every dead tree trunk was assigned to one of five decay classes, adopting the classification system of Meyer et al. (2001) that agrees well with the systems proposed by Hunter (1990), Nagel and Svoboda (2008) and others. On every plot the height of three tree individuals of the main tree species (beech in Kyjov and Havešová; beech and fir in Stužica) and of all individuals of the less abundant tree species was recorded with a Vertex IV height meter (Haglöf Sweden AB, Långsele, Sweden). The volume of living trees was then calculated according to Petráš and Pajtík (1991), and for dead trees, we applied a reduction factor in dependence on their decay class, as proposed by Meyer et al. (2001). All trees were inspected for the occurrence of microhabitats formed by bark injury or the bracket fungus, and cavities at the stem base and the upper stem and signs of necrosis on the trunk. Tree saplings > 1.5 m in height (termed ‘advanced regeneration’) were counted in two belt transects of each 13 m2 area per plot.
The Development Stage Index (I DS )
In temperate broad-leaved old-growth forests, plots of 500 m2 size typically contain tree individuals of variable age and dimension, which from an ontogenetic perspective, may be assigned to different forest development stages. Even in monospecific stands, the forest structure in a plot of this size can be quite heterogeneous. Groups of trees of different ages and dimensions may occur in close neighborhood to each other, and identified development stages thus may overlap through vertical canopy stratification. To account for this heterogeneity, we introduced the Development Stage Index I DS , which is derived from measured tree dimensional data. It calculates the relative extension of development stages in a plot and allows assigning a plot to a prevailing stage by identifying the stage with largest extension.
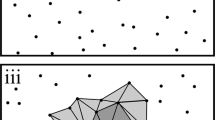
Stem diameter was used as the principal criterion for assigning a tree or tree group to one of the three development stages ‘Initial’, ‘Optimum’ or ‘Terminal’. The dbh thresholds were chosen by approximating points that enclose a section of the mean stem diameter–density distribution curve of the three virgin forests with low curve steepness (Fig. 1). According to Goff and West (1975) the beginning and end points of such a section (benchmarks) indicate transitions between three distinct phases in the tree life cycle in temperate old-growth forests of shade-tolerant North American tree species. The resulting dbh classes and their main characteristics are given in Table 2.

Semi-logarithmic stem diameter–density distribution curve of living trees (N ha−1) per 5 cm dbh class for the three forest stands (dashed lines) and a fitted line (continuous), representing a third-degree polynomial function calculated on the mean of the three stands across the dbh classes. The continuous vertical lines mark the locations of the thresholds set for the assignment of trees to the three stages ‘Initial’, ‘Optimum’ and ‘Terminal’. The dashed vertical lines mark the altered thresholds in the sensitivity analysis
These diameter thresholds largely fit to a categorization of tree dimensions often used in silviculture (e.g., Röhrig et al. 2006). For simplicity, the premature trees in a plot were subsequently assigned to the Initial development stage, mature trees to the Optimum stage, and over-mature trees to the Terminal stage of forest development (Fig. 2). Dead trees have until recently influenced the structure of the living stand and still are imprinting on the recent stand structure through competitive effects that were exerted by the now-dead trees in the past. To include dead trees in the I DS thus can help to understand the stand development in the more recent past. Therefore, not only live trees were considered in the calculation of I DS , but dead trees as well to account for their contribution to forest structure and space filling. We used the same dbh-based classification as above, assuming that mortality is to a considerable extent caused by community-level processes characteristic of the considered life history phase (Holzwarth et al. 2013). With proceeding decay, dead trees were reduced in their count and for calculating basal area by applying specific reduction factors for the five wood decay classes (DC) (DC 1 = 1.0, DC 2 = 0.95, DC 3 = 0.85, DC 4 = 0.7 and DC 5 = 0.5). Thus, deadwood represents a structural memory of the recent past in the I DS score that is fading with increasing wood decay. This procedure considers that the space once occupied by the live tree is increasingly filled by neighbors and recently established individuals. Beech logs up to class DC 5 are present in the stands for about 35 years (Müller-Using and Bartsch 2009; Prívetivý et al. 2016); according to Lombardi et al. (2008) beech and fir logs decompose at similar rates.

Scheme illustrating the assignment of stems of variable diameter in an exemplary 500-m2 plot to the three development stages (Initial, Optimum and Terminal) using the dbh categories 7–39, 40–69 and ≥ 70 cm as selection criteria. Thus, the plot contains elements of all three stages
The relative importance of trees of these three diameter categories in a plot was quantified through their abundance in terms of stem number (N) and basal area (BA) to account for tree size differences. To solely rely on BA as a measure of ‘natural stocking density’ (Assmann 1961) is not sufficient here. Trees at the lower end of a size class have a low BA, but can reach high N; the opposite is valid for trees at the upper end of the class. Both dimension measures were expressed in relative terms using forest patches with more or less exclusive presence of one of these tree size classes as a reference (see Table 4).
Calculation of I DS
For all plots i, the summed number of trees (N DS i ) and the summed basal area (BA DS i ) in the respective stages (DS) were calculated. To define the highest achievable tree number, which a given diameter category or development stage may reach in a plot (N DSref ), we identified the plots with the highest tree number (N DSmax ) in that stage in the three forest stands and averaged over these maximum densities:
The same was done for the parameter basal area:
The index value I of a given development stage DS (I DS ) in plot i was then calculated as the summed quotients of actual tree number (N DS i ) to potential maximum tree number (N DSref ), and actual basal area (BA DS i ) to potential maximum basal area (BA DSref ) in this diameter category:
with DS being the Initial (Ini), Optimum (Opt) or Terminal (Ter) stage.
The stage with the highest index score is per definitionem the dominating development stage in that plot. Dominance refers only to abundance and not to interactions between trees of different diameters. A main purpose of the I DS score is to measure the complex structural mixing of the three development stages on the plot level. The relative proportion of I DS scores of the three stages informs more about the current structural composition of the stand than about the stages’ conditions and the development potential. However, it can also be used to infer on processes of dynamic structural change, when repeated surveys with the index are conducted.
Data analysis
The index I DS was used for two purposes: (1) to generate data on the relative abundance of the three development stages in the individual plots and the forest stands and (2) to identify the most extended (or ‘dominant’) stage in a plot. These data can be used for mapping principal forest structural categories in old-growth forests, and they served for calculating the relative extension of previously defined development stages in the three primeval forests. As a measure of structural heterogeneity (and not species diversity) in the sample plots, we calculated Shannon’s diversity index (H′) and evenness with the relative stage extension data of the three stages in the plot. In the calculations, we assigned zero values to stages that were currently not present on a plot; the calculation of evenness was done with reference to conditions with equal abundance of all three stages in a plot.
There is the possibility that the I DS approach works only in the frame of the dbh classes and plots sizes which exist in the studied forests. To explore whether alterations of the proposed I DS index to modification in the chosen dbh classes and plot size lead to unexpected results which might be in conflict with the underlying assumptions and thus challenge the validity of the index, we performed a sensitivity analysis. We tested for the effect (1) of altered dbh thresholds used for distinguishing the tree diameter classes and (2) of different plot sizes on the I DS values calculated for the plots. In these runs, the dbh thresholds were shifted from 40 to 30 cm and from 70 to 80 cm, and plot size was reduced from 500 to 156.25 m2.
We further analyzed the relationships between the Development Stage Index I DS and selected structural characteristics of the trees and stands, notably:
-
the number of live, dead and all trees (N l , N d and N tot )
-
the BA of live, dead and all trees (BA l , BA d and BA tot )
-
the volume of live, dead and total wood (V l , V d and V tot )
-
the density of advanced regeneration (REG), and
-
the number of microhabitats (HAB).
The three stands were compared for significant differences in their structural characteristics. If this was the case, then further analyses were performed separately for the three stands. We performed correlation analyses for the I DS scores of the three stages and for their relative proportions (I DS /I Total ) on the plots. The first may better indicate stage-specific density effects, and the latter the effects of apparent structural complementarity among the stages. The fit and significance of correlations was analyzed by performing Pearson’s product-moment correlation. After stratification of the plots by assigning them to the prevailing development stage, the three development stages were tested for significant differences in the above-mentioned structural characteristics. As the data for stands and stratified stages did not fit to normal distribution, a Mann–Whitney U test was employed. All analyses were performed with R software, version 3.2.2 (R Core Team 2015), using a confidence level of 0.95 throughout. The p values for multiple testing were adjusted by the Bonferroni–Holm method.
Results
Stand structural characteristics
The three virgin forests had similar mean densities of live trees and nearly equal basal areas (Table 3), while the number of dead trees increased in the sequence KY < HA < ST (HA and ST significantly different from KY). The difference in total tree density (live and dead) was also small between the stands. There was no significant difference in the volume of above-ground wood biomass and dead wood volume among the three stands, but the volume of live and dead wood taken together was ~ 20% smaller in KY than in HA and ST (difference significant). The density of advanced regeneration (height > 1.5 m) was highest in ST, but the stands differed not significantly in this respect.
The volume of above-ground wood biomass increased from the Initial to the Optimum stage (Fig. 2 in supplementary material). Between the Optimum and the Terminal stage, it remained stable in the HA and KY forests, but increased in the ST stand. When deadwood is added to obtain total wood volume, all stands showed increasing volumes from the Initial to the Terminal stage. Among the plots of a development stage, wood volume generally varied greatly.
Characterizing stand structure by the Development Stage Index I DS
The minimum number of trees found on a 500-m2 plot was n = 7. All plots contained trees of two, and often three, different stem diameter classes. Thus, the same plot was often assignable to two or more development stages. Ninety-nine percent of the plots contained trees assignable to the Initial stage (7–39 cm dbh), 98% trees of the Optimum stage (40–69 cm), and 88% contained over-mature trees (≥ 70 cm) assignable to the Terminal stage. In the KY stand, over-mature trees were present in 98% of the plots, indicating a relatively even distribution of large trees across the stand. Even though mean stem density and basal area were not significantly different between the three stands, the observed maximum N and BA values in the respective stages differed between the HA, KY and ST forests (Table 4). N Ini and BA Ter reached exceptionally high maxima in certain HA plots, but much lower peak values in KY (except for BA Ini ). With the means of the stage-specific maxima being used as a reference, the relative stem density and basal area values of the three stages (0.31–0.47, mostly in the range 0.35–0.44) were remarkably similar in the three stands (supplemental Table 1). The calculated I DS scores reached very similar proportions of I Ini , I Opt and I Ter in I Total (the summed indices of all three stages) regarding all 118 plots, but varied to some extent in their means (Table 6) and distribution (Fig. 1 in supplementary material) between the three stands. As follows from the logics of the I DS score approach, we found significant negative correlations between the I DS score of a single stage with the summed scores of the respective two other stages (I Ini : r = − 0.43, p ≤ 0.001; I Opt : r = − 0.55, p ≤ 0.001; I Ter : r = − 0.64, p ≤ 0.001).
As expected, I DS was closely related to various plot-level structural attributes such as the basal area and wood volume of the living trees (Table 5). This relation was positive for I Ter and I Opt , but negative for I Ini , as trees of the latter stage profit from a reduced biomass of the older and larger trees. A close negative relation was also found between the I Ini score and the abundance of tree saplings > 1.5 m (REG), reflecting that dense stands of young trees generally suppress the offspring most effectively. A strong correlation (r: 0.87–0.94) exists between the I DS scores of the three stages and their corresponding proportions (I DS /I Total ) in a plot. Accordingly, we found similar tendencies for the relations between the stages´ proportions and the structural attributes (Table 5). The Initial stage on the one hand and the Optimum and Terminal stages on the other revealed significant differences in the structural parameters (N l , BA l , BA tot and V l ) (Table 2 in supplementary material).
Spatial extension of development stages
The I DS score allows quantifying the extension of different development stages in the forest, either through the relative extension of the stage in the total investigated plot area or via the proportion of all plots that are dominated by that stage. As shown by the box plots in supplemental Fig. 1, the I DS scores characterizing the abundance of the three stages vary largely among the plots, reflecting small-scale heterogeneity in the stands. Structural differences between the three forests become visible when comparing the median values (supplemental Fig. 1) and arithmetic means (Table 6) of the I Ini , I Opt and I Ter scores of the three stands. The Terminal stage with over-mature trees is more widely distributed in the KY forest (34% of the total plot area), while the ST forest has a greater presence of the Optimum and Initial stages in its plots (present on 34 and 39%, respectively); in contrast, the Terminal stage occurs in ST on only 28% of the area (Table 6).
A similar picture emerges when the proportion of plots dominated by a given stage is compared among the three forests: The ST forest has a lower frequency of the Terminal stage (dominant in 32% of the plots), which is more widespread in the KY and HA forests (35 and 38%; Table 7). While plots dominated by the Terminal stage prevail in the HA forest, the most frequent plots in ST are those dominated by the Optimum stage. The KY forest ranges between HA and ST with respect to the dominance of Optimum and Terminal phase.
Sensitivity analysis of the classification scheme
Shifting the dbh thresholds for the premature tree class from 7–40 cm in the original scheme to 7–30 cm and that of the over-mature tree class from > 70 to > 80 cm increased the dbh range of the trees which are included in the Optimum (> 30–≤ 70 cm) stage with the result that the mean I Opt score increased in the three stands by 21%, while the mean scores of the Initial and Terminal stages (I Ini and I Ter ) decreased by 4 and 18% compared to the original scheme. Thirty-one instead of 13% of the plots contained no trees assignable to the over-mature dbh category (Terminal stage). When reducing the plot size from 500 to 156.25 m2, still more than 95% of the plots contained two or more trees (median: n = 5.5). The number of plots, in which only two development stages were present, strongly increased from 19 to 81 of 118; 16 plots contained just a single stage (in most cases the Initial). Thus, smaller plots resulted in less spatial overlap of the three development stages, but the average proportion of the three development stages in the total investigated plot area remained similar to that derived for the 500-m2 plots.
Visualizing the mingling of stages in the study plots
We used ternary graphical plots to visualize for our 118-plot sample the variable mixing of trees assignable to the three development stages. For every plot, the I DS scores of the three stages were expressed in percent of the summed three I DS values of that plot (= I Total ) along three axes from 0 to 100%, which define the contribution of the Initial, Optimum and Terminal stages to the I Total score (see Fig. 3 for ternary plots of the HA, KY and ST stands, and the pooled plots of all stands). The majority of plots was located in moderate distance to the center of the graph, where the three stages contribute equally to I Total (by about 33%); this is also expressed by the evenness of the three I DS scores which reaches high means in all three stands (Table 6). Plots with high percentages of I Ini , I Opt and I Ter , and low percentages of I Ini and I Opt , are rare. In contrast, plots with low or zero scores of I Ter were present in all three forests. (I Ter was zero in 12% of the 118 plots.) The HA forest differed from the other two stands in that many plots were located left of the plot center at higher I Ter scores, while the ST plots were more frequent right of the center toward lower I Ter scores. The 40 KY plots showed the most even distribution across the ternary plot, reflecting the relatively balanced proportions of stages on the stand level (Table 6) in this forest.

Ternary diagrams visualizing the relative importance of the Initial, Optimum and Terminal stage in the 40 (or 38) plots in the stands HA, KY and ST and in all plots pooled (all stands). The relative importance of a stage is expressed by its I DS score related to the plot I Total (the summed I DS scores of all three stages in a plot). The three arrows in the diagram indicate the position of a plot along the 0–100% axes for the Initial stage (upper corner of triangle), the Optimum stage (lower right corner) and the Terminal stage (lower left corner). Plots located in the dark-gray area have a relatively similar share of all three stages. (No stage exceeds the other two in terms of stem numbers and basal area more than 1.5-fold). In the medium-gray zone, two stages exceed the other one more than 1.5-fold. In the light-gray zone, a single stage exceeds both other stages more than 1.5-fold. The size of the circles gives the size of the total index score I Total of the plot (small: < 0.9, medium: 0.9–1.3, large: > 1.3), indicating the total density of stages
Discussion
Plot number and plot size requirements in old-growth forest studies
Plot numbers and plot size determine the information quality available when studying forest stand structure. In the long-term absence of stand-level disturbances, primary forests typically develop toward horizontally and vertically diverse ‘old-growth’ structures (Franklin et al. 2002, Bauhus et al., 2009). In forests with high structural heterogeneity the coefficient of variation of many structural characteristics tends to increase with decreasing plot size (Král et al. 2010a). This inflates the minimum number of sample plots required for reliably estimating the spatial variability of the stand structure at small plot sizes. According to the approach given by Král et al. (2010a), our 118 plots well exceed the minimum plot number needed for reliably estimating total basal area (n = 27) and total tree number (n = 49).
Plot size depends on the type of information sought in the study. A meaningful spatial scale in the study of beech forest dynamics may be defined by gap size (Drößler and Meyer 2006). Spies and Franklin (1996) also use the term ‘shifting-gap phase’ for their last phase in forest developmental succession. Most canopy gaps in beech old-growth forests result from the death of single trees or small groups of trees (Bottero et al. 2011; Drößler and Lüpke 2005; Kenderes et al. 2009; Kucbel et al. 2010; Tabaku and Meyer 1999; Zeibig et al. 2005) with a size of < 100 m2 being most frequent, while stand-replacing disturbances (e.g., Hobi et al. 2015; Kucbel et al. 2012) seem to be rare in European beech forests. Analyzing spatial tree distribution in the Kyjov virgin beech forest, Drößler et al. (2016) found gap size patterns to be reflected in stand structure throughout the entire forest development cycle. Single trees were the most frequent ‘group size’ suggesting to collect information with a spatial grain size of a single tree in studies on beech forest dynamics. Since sample plots must be larger than this minimum grain size to conclude on stand structure and structural interactions, plot dimensions should exceed the area covered by a single tree, i.e., approximately 150 m2 for a beech canopy tree (Meyer, 1999). When studying the vertical stratification of canopies, even larger plots should be picked. We followed Lombardi et al. (2015) who assumed a plot size of 500 m2 to be adequate for quantifying old-growth structural indicators in European temperate forests.
Quantifying forest structure with the Development Stage Index I DS
The introduced Development Stage Index I DS is based on the grouping of trees into three diameter classes that are then referred to as ontogenetic development phases. Goff and West (1975) also divided the life cycle in old-growth forests of shade-tolerant North American tree species into three distinct phases: an understory phase of slow growth and high mortality, a vigorous canopy phase of relatively rapid growth and low mortality, and an older canopy phase of reduced growth and increased mortality. These authors associate the transitions between these phases with the benchmarks in the inclination of the polynomial curve fitted to the tree diameter–density distribution, which has a rotated sigmoidal shape. This curve reflects the initial exponential decline in stem numbers (understory phase) that turns into a subsequent plateau with relatively stable stem numbers (vigorous canopy phase) and is then followed by an increasing decline at high dbh values (senescent overstory phase). The diameter–density distributions in our three stands also fit to this curve type, as has also been reported from other beech-dominated virgin forests (e.g., Alessandrini et al. 2011; Westphal et al. 2006). This suggested to adopt the criteria formulated by Goff and West (1975) for distinguishing between the three ontogenetic phases in our study. We used this dbh-based classification also for standing dead trees, assuming that mortality is to a considerable extent caused by community-level processes characteristic of the considered life history stage (Holzwarth et al. 2013).
One may argue that describing the forest development cycle by distinguishing only three stages, as proposed here, must represent an over-simplification of forest structure and a loss of information compared to other approaches which used more stages (e.g., disintegration, gap, regeneration and plenter phases or stages; Král et al. 2016; Tabaku, 2000). However, the results obtained from our analysis with the I DS scores clearly demonstrate that the areal mingling of only three development stages results in complex patterns (e.g., Fig. 3). These patterns correspond to a diversity of forest structures and thus facilitate the interpretation of structural dynamics of primeval forests, in particular when applied in repeated surveys.
The calculation of I DS scores requires referencing to tree density (N) and basal area (BA) data of plots which are dominated mostly or exclusively by that development stage. Since the Terminal stage forms extended patches only in old-growth forests, our approach is applicable only to forest stands with a long continuity of natural development, but not to stands with higher human impact. Yet, transferring I DS reference values derived from virgin forests to other forests of the same species and site characteristics seems feasible. This is indicated by the only moderate variation in N max values (found in the Optimum and Terminal stages) and BA max values (found in the Initial and Optimum stages) of a development stage among the three Slovakian beech forests. Considerable variation in N max and BA max values was only observed in plots, where either stem number (as found in N maxIni ) or dbh was exceptionally large (as seen in BA maxTer ). Recorded differences in the stem density and basal area maxima likely are caused by the sampling design with plot numbers probably still being too small to reliably detect the N and BA maxima in these forests, as the whole-stand means of tree density and basal area were not different between the three forests. To account for this possible shortcoming, we averaged over the N max and BA max values of the three forests, when calculating the stem density and basal area reference.
I DS is a mixed index which expresses the effects of both tree abundance and tree size on the spatial extension of a development stage in the forest. This is important as the cover of the Initial stage is mostly determined by the number of trees (N), while it is BA that largely determines cover in the Optimum stage, when stand thinning is completed. In the Terminal stage, dying trees reduce stem density, a process which typically is not compensated by the basal area growth of the remaining trees; thus, N achieves a larger influence on cover again. Combining both measures ensures that I DS can be used to estimate the extension of all three development stages. Highly significant negative correlations between the I DS scores of the three stages are an expression of the fact that the different demographic populations of beech in a stand compete for canopy space.
As expected, the I DS index was sensitive to the choice of dbh class ranges in our run with altered dbh thresholds. If a diameter class is narrowed, this class represents a shorter period in the forest cycle and the forest area assignable to the corresponding forest development stage must decrease. From diameter frequency plots of beech old-growth forests, it appears that the chosen 40 and 70 cm class limits are justified because they roughly correspond to demographic benchmarks in the life cycle of a beech tree. The nearly equal abundance found for the three development stages in the pooled plots of the three Slovakian virgin forests also underpins this judgment. Certainly, other tree species may require defining different dbh thresholds. While plot size alters the size of I DS and I Total values in the plots, it appears to be less influential on the calculated proportion of the development stages at the stand level. This indicates that the I DS index is applicable to different plot sizes and thus to different types of forest inventory data, while the proportion of forest area assigned to the three stages is largely unaffected. In contrast, the development stage classification approach proposed by Tabaku (2000) is highly sensitive to plot size: Raising plot size from 156.25 to 500 m2 caused large changes in the proportion of phases identified in the stands we studied (Table 3 in supplementary material), as well as in the Ukrainian beech virgin forest of Uholka (compare Peck et al. 2015 and Zenner et al. 2016). The greatest changes were visible in the large extension of the ‘plenter phase,’ which is a surrogate for high spatial heterogeneity.
Distribution and abundance of development stages
We found roughly similar frequencies (about 30–36%) of the Initial, Optimum and Terminal stages across our 118 plots, irrespective of the calculation approach used (the proportion of the stage-specific I DS in I Total versus the proportion of plots dominated by a certain stage). Under the assumption of a landscape-scale quasi-equilibrium state of the studied virgin forests, the relatively balanced distribution of indices evidences similar spatial extensions of the three development stages in the forest suggesting that the dbh classes were well chosen. Since growth and development of shade-tolerant trees are often retarded in the understory, the extension of a forest development stage, however, is not a good indicator for the duration of a given stage in a tree’s life cycle. In fact, dendroecological data indicate that beech trees are often spending much longer time spans in the Initial stage than in the other two stages (Trotsiuk et al. 2012).
Different development stage definitions in other classification approaches can lead to largely deviating results. For example, the approach of Tabaku (Drößler and Meyer 2006; Tabaku 2000; Zenner et al. 2016) applied to primeval beech forests results in a much larger proportion of phases corresponding to our Terminal stage (nearly 50%), while only about 17 and 12% were assigned to phases related to our Initial and Optimum stages, respectively. This is caused by a focus on attributes with association to late developmental stages (deadwood and large trees), while information on other structural elements such as lower canopy strata is not considered. A considerable proportion of the plots had been characterized as the ‘plenter phase,’ which has no direct equivalent in our classification system and has been criticized for being a synonym for small-scale spatial heterogeneity without logical order in the forest dynamics cycle (Winter and Brambach 2011). According to this approach, the abundance of development stages is highly unbalanced in all six beech primeval forests, to which it was applied (see Zenner et al. 2016). On our plots, we see the same tendency when the approach of Tabaku is used (supplementary Table 3). This suggests that all investigated beech primeval forests in eastern-central and southeastern Europe should shift in the next decades from the terminal and decay phases to early development phases at large parts of their area. We assume that this is a consequence of methodology rather than an overarching ecological phenomenon of beech forest dynamics in our time. Král et al. (2010b) proposed another classification approach, which yielded more balanced proportions of the stages, but the results varied considerably during the study period of 30 years (Král et al. 2014).
Analysis of the I DS data in our study shows that all three stages coexist in the plots of 500 m2 size, demonstrating high spatial heterogeneity in the three Slovakian virgin forests. This seems to contrast with reports of other authors on the occurrence of relatively homogeneous patches in European old-growth forests (e.g., Müller 1929 and Leibundgut 1982, 1993). A possible explanation is that these authors studied forests in which past large-scale disturbances created extensive forest patches with cohort-like structure. We rarely encountered the exclusive presence of one single development stage on larger areas, as has been described by Korpel (1995) for Slovakian virgin forests. In fact, only very few of our 118 plots reached high proportions (> 70% of I Total ) for any of the three stages, or alternatively low proportions (< 10%) for the Initial or Optimum stages. The high evenness of the three stages in the plots (> 0.84 in all stands) is another expression of the high small-scale heterogeneity, which has been addressed in earlier old-growth forest studies by introducing the term ‘plenter structure’ (Tabaku 2000). The standard deviation of I DS in Table 6, which varied from 43 to 77% of the mean in the three forests, expresses differences between the forests in terms of the degree of structural heterogeneity. Deviation from equality in the abundance of Initial, Optimal and Terminal stage plots [as in the Havešová (HA) and Stužica (ST) forests] may indicate past disturbance events such as windstorms that may affect a forest on the stand scale (e.g., Nagel et al. 2014).
From the ternary plots, different stand-level structural patterns become visible for the three studied primeval forests. These patterns likely are providing information on past disturbance regimes and environmental heterogeneity at the stand level, but in the absence of repeated inventory data, any interpretation must remain speculative. However, a remarkable outcome is that many study plots are localized relatively close to the center of the ternary plot in all three forests, suggesting that the degree of small-scale mixing of the three stages is indeed high with frequent stage overlap within a plot. The graphical presentation of canopy structural heterogeneity in a ternary plot is a promising tool, which allows visualizing the complex mixing of life history stages in temperate old-growth forests and enables interpreting the underlying dynamic processes. Application to repeated inventory data might help in understanding structural self-organization processes in the canopy, as they become visible in changing study plot positions over time in the ternary plot.
Deadwood as an old-growth forest attribute
Deadwood adds largely to the conservation value of old-growth forests due to its importance for xylobiontic organisms and cavity-nesting birds (e.g., Begehold et al. 2015; Müller and Bütler 2010; Winter and Brambach 2011). It is also a valuable structural attribute in assessments of the old-growth character of forests (e.g., Bauhus et al. 2009) and a decisive structural feature in the definition of development phases and stages (e.g., Tabaku 2000). The I DS index associates deadwood objects with the different forest development stages through diameter, assigning thin deadwood to the Initial stage and thick dead logs to the Terminal stage. High deadwood numbers and volumes increase the I DS score. This approach has not been used before, since deadwood then partly loses its indicator value for forest development phases or stages. We share the opinion on the important role of deadwood as a characteristic structural attribute of old-growth forests and its high value in conservation matters. However, tree death is in many cases related to processes characteristic of the specific life history stage and thus the diameter class of the tree (Holzwarth et al. 2013). Furthermore, in forest dynamics, the death of a tree in the first place implies a release of growing space and the associated availability of resources (light, water and nutrients) to the surrounding trees. The still important role of deadwood in the I DS may become visible in particular, when the index is applied to repeated inventory data, because deadwood decay gradually decreases the I DS score of the respective stage. For example, if a storm kills all Terminal stage trees in a plot, where this stage was dominant, 10 years later the I Ter score will decrease considerably by the decline in N Ter and BA Ter , while other individuals that survived will profit from the released growing space and react with increased growth. Thus, the I DS score of the respective stages will increase. The consideration of deadwood in the calculation of the score causes the index to react only slowly to structural changes caused by tree mortality. Therefore, the I DS score is best applied in long-term observational studies. Yet, deadwood in its role as a structural memory of the past can improve the understanding of interactions among the different stages and may help in tracking stage transitions in the forest that happened in past decades.
Incorporating deadwood abundance in the index allows searching for structural inter-dependencies by means of correlation analysis. Earlier studies have frequently reported a negative correlation between deadwood volume and live wood mass (e.g., Král et al. 2010a; Holeksa et al. 2009). The I DS score, however, incorporates deadwood volume (through BA d ) and the number of deadwood objects (through N d ), both of which may change independently across development stages. We observed a positive relation between I Ini and N d , probably reflecting the outcome of self-thinning processes in this stage. Dead trees in the Initial stage are of small size and do only marginally influence BA d and V d . As the Initial stage usually follows the Terminal stage with gradual stand decay (or large-scale disturbance) and typically is lasting much longer than the period of deadwood decomposition, deadwood volume and numbers often vary largely (see also Král et al. 2010b and Tabaku 2000). While deadwood amount passed through a minimum in the Optimum stage with typically lowest mortality (Korpel 1995), a significant negative relation between the number of dead logs (N d ) and the I Opt score did not exist in our data (negative trends in the HA and KY stands, but positive relation in ST with the presence of fir).
In our dbh-based definition, the Terminal stage covers a time span from ‘growing old’ to ‘replacement by young trees,’ resembling the definition of the ‘breakdown stage’ proposed by Král et al. (2010b). While our definition is not necessarily related to the presence of dead trees, other classification approaches define distinct disintegration phases or decay stages (Jaworski and Podlaski 2007; Korpel 1995; Tabaku 2000), which may result in higher deadwood amounts in this phase. While we found only a nonsignificant tendency toward higher deadwood amounts with higher I Ter scores, this index showed a significant positive relation to deadwood volume (r = 0.25, p = 0.006), when only coarse woody debris of low to medium decay (> 20 cm dbh, decay class ≤ 3) was considered. This substrate is of higher value for xylobionts (Schuck et al. 2004). For I Ini and I Opt , such a relation was not found, which indicates that the over-mature trees of the Terminal stage indeed are largely determining the deadwood amount.
I DS as a proxy for further stand structural characteristics
From the correlation analyses, it is evident that the I DS scores, and likewise the stages´ proportions, can provide further information on stand structure in old-growth forest plots (see Table 5). As expected, higher I Ini scores generally stand for higher stem densities also in plots that are dominated by the respective other development stages, while a higher I Ter score indicates smaller overall tree densities. Higher I Opt and I Ter scores stand for higher cumulative basal areas in the plot, while higher I Ini values relate to smaller basal areas. The same is true for the volume of above-ground wood biomass. Interestingly, a higher frequency of microhabitats (such as cavities in the stem and bark injury) is indicated not only by higher I Ter scores, but also by higher I Opt values. A similar abundance of microhabitats in the Optimum and Terminal stages (see Table 2 in supplementary material) may suggest that microhabitats relevant for xylobionts are created in virgin beech forests well before the trees are reaching over-mature size. One explanation is the generally higher stem density in the Optimum than Terminal stage, which may outweigh the lower specific frequency of microhabitat occurrence. Another explanation could be that falling dead trees are damaging younger, vital neighbors, which is prevented in managed forests. Since we registered only the density of microhabitats, but did not assess their quality, it may, however, be that the Terminal stage with its very old and large trees does possess habitats of greater value for xylobionts than do exist in the Optimum stage.
High I Ini scores are also indicators of a reduced density of tree saplings > 1.5 m height, probably because they are suppressed by a dense cover of young trees. Thus, in these montane virgin beech forests, regeneration is highest in the Optimum and Terminal stages, but is largely suppressed in the Initial stage, which allows the next beech generation to develop only after the trees of the Initial stage have grown tall. As the I DS scores of the three stages correlate positively with the I Total score (r: 0.22–0.49), the latter is also associated with stand structural characteristics that are strongly correlated with one or more I DS scores.
Conclusions
The proposed I DS index can be viewed as an important step toward the goal to describe and analyze the complex mosaic structure of temperate old-growth forests with objective and quantitative measures. The index bases on two easily measured variables, which serve as suitable proxies for quantifying the spatial extension of life history stages from the Initial to the Terminal stage. The index seems to be relatively robust against variation in plot size, but is sensitive to altered classification schemes of the development stages. The I DS score allows interpreting stand development from the plot to the landscape scale. On the plot level, I DS scores provide information on the mixing of tree populations of different demographic positions, allowing conclusions on vertical structure and its spatial variation in the forest and thus on the character of the disturbance regime. At the stand and landscape scales, mean I DS values can give hints on past major disturbances, visible through deviation from equilibrium conditions. Displaying I DS scores in ternary graphical plots allows visualizing spatial variation in the mixing of development stages. This approach can also facilitate the comparison of different forests in terms of canopy structure and function. Further, when the ternary plot is applied to repeated inventory data, dynamic changes in stand structure can be analyzed. Finally, the improved empirical database generated by introducing the I DS score may enable more rigorous hypothesis testing in forest dynamics research. In future studies, the I DS index should be applied to other beech old-growth forests, repeated inventory data and further structurally different forest types. This may require modifying the index by altering the dbh thresholds, including other or additional structural variables and extending the set of stem density and basal area data, which are needed as a reference.
References
Alessandrini A, Biondi F, Di Filippo A, Ziaco E, Piovesan G (2011) Tree size distribution at increasing spatial scales converges to the rotated sigmoid curve in two old-growth beech stands of the Italian Apennines. For Ecol Manag 262:1950–1962
Assmann E (1961) Waldertragskunde. BLV Verlagsgesellschaft, München
Bauhus J, Puettmann K, Messier C (2009) Silviculture for old-growth attributes. For Ecol Manag 258:525–537
Begehold H, Rzanny M, Flade M (2015) Forest development phases as an integrating tool to describe preferences of breeding birds in lowland beech forests. J Ornithol 156:19–29
Bohn U, Neuhäusl R, Gollub R, Hettwer C, Neuhäuslova Z, Schlüter H, Weber H (2003) Karte der natürlichen Vegetation Europas. Teil 1: Erläuterungstext. Landwirtschaftsverlag, Münster
Bottero A, Garbarino M, Dukić V, Govedar Z, Lingua E, Nagel TA, Motta R (2011) Gap-phase dynamics in the old-growth forest of Lom, Bosnia and Herzegovina. Silva Fenn 45(5):875–887
Christensen M, Emborg J, Nielsen AB (2007) The forest cycle of Suserup Skov—revisited and revised. Ecol Bull 52:33–42
Commarmot B, Bachofen H, Bundziak Y, Bürgi A, Ramp B, Shparyk Y, Sukhariuk D, Viter R, Zingg A (2005) Structures of virgin and managed beech forests in Uholka (Ukraine) and Sihlwald (Switzerland): a comparative study. For Snow Landsc Res 79:45–56
Commarmot B, Brändli UB, Hamor F, Lavnyy V (2013) Inventory of the largest virgin beech forest of Europe. A Swiss-Ukrainian scientific adventure. Birmensdorf, Swiss Federal Research Institute WSL; Lviv, Ukrainian National Forestry University, Rakhiv, Carpathian Biosphere Reserve
Drößler L, Lüpke B (2005) Canopy gaps in two virgin beech forest reserves in Slovakia. J For Sci 51:446–457
Drößler L, Meyer P (2006) Waldentwicklungsphasen in zwei Buchen-Urwaldreservaten in der Slowakei. Forstarchiv 77:155–161
Drößler L, Feldmann E, Glatthorn J, Annighöfer P, Kucbel S, Tabaku V (2016) What happens after the gap?—Size distributions of patches with homogeneously sized trees in natural and managed beech forests in Europe. Open J For 6:177–190
Emborg J, Christensen M, Heilmann-Clausen J (2000) The structural dynamics of Suserop Skov, a near natural temperate deciduous forest in Denmark. For Ecol Manag 126:173–179
Franklin JF, Spies TA, Van Pelt R, Carey AB, Thornburgh DA, Berg DR, Lindenmayer DB, Harmon ME, Keeton WS, Shaw DC, Bible K, Chen J (2002) Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For Ecol Manag 155:399–423
Goff FG, West D (1975) Canopy-understory interaction effects on forest population structure. For Sci 21:98–108
Grassi G, Minotta G, Giannini R, Bagnaresi U (2003) The structural dynamics of managed uneven-aged conifer stands in the Italian eastern Alps. For Ecol Manag 185:225–237
Hobi ML, Ginzler C, Commarmot B, Bugmann H (2015) Gap pattern of the largest primeval beech forest of Europe revealed by remote-sensing. Ecosphere 6:1–15
Holeksa J, Saniga M, Szwagrzyk J, Czerniak M, Staszyńska K, Kapusta P (2009) A giant tree stand in the West Carpathians—an exception or a relic formerly widespread mountain European forests? For Ecol Manag 257:1577–1585
Holzwarth F, Kahl A, Bauhus J, Wirth C (2013) Many ways to die—partitioning tree mortality dynamics in a near-natural mixed deciduous forest. J Ecol 101:220–230
Hunter ML Jr (1990) Wildlife, forests and forestry: principles of managing forests for biological diversity. Prentice-Hall, Englewood Cliffs
Jaworski A, Podlaski R (2007) Structure and dynamics of selected stands of primeval character in the Pieniny National Park. Dendrobiology 58:25–42
Kenderes K, Král K, Vrška T, Standovar T (2009) Natural gap dynamics in a Central European mixed beech-spruce-fir old-growth forest. Ecoscience 16(1):39–47
Korpel Š (1995) Die Urwälder der Westkarpaten. Gustav Fischer Verlag, Stuttgart
Král K, Janík D, Vrška T, Adam D, Hort L, Unar P, Šamonil P (2010a) Local variability of stand structural features in beech dominated natural forests of Central Europe: implications for sampling. For Ecol Manag 260:2196–2203
Král K, Vrška T, Hort L, Adam D, Šamonil P (2010b) Developmental phases in a temperate natural spruce-fir-beech forest: determination by a supervised classification method. Eur J For Res 129:339–351
Král K, Valtera M, Janík D, Šamonil P, Vrška T (2014) Spatial variability of general stand characteristics in central European beech-dominated natural stands—effects of scale. For Ecol Manag 328:353–364
Král K, Shue J, Vrška T, Gonzalez-Akre EB, Parker GG, McShea WJ, McMahon SM (2016) Fine-scale patch mosaic of developmental stages in Northeast American secondary temperate forests: the European perspective. Eur J For Res 135:981–996
Kucbel S, Jaloviar P, Saniga M, Vencurik J, Klimaš V (2010) Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians. Eur J For Res 129:249–259
Kucbel S, Saniga M, Jaloviar P, Vencurik J (2012) Stand structure and temporal variability in old-growth beech-dominated forests of the northwestern Carpathians: a 40-years perspective. For Ecol Manag 264:125–133
Leibundgut H (1982) Europäische Urwälder der Bergstufe. Paul Haupt, Bern
Leibundgut H (1993) Europäische Urwälder—Wegweiser zur naturnahen Waldwirtschaft. Paul Haupt, Bern
Lombardi F, Cherubini P, Lasserre B, Tognetti R, Marchetti M (2008) Tree rings used to assess time since death of deadwood of different decay classes in beech and silver fir forests in central Apennines (Molise, Italy). Can J For Res 38:821–833
Lombardi F, Marchetti M, Corona P, Merlini P, Chirici G, Tognetti R, Burrascano S, Alivernini A, Puletti N (2015) Quantifying the effect of sampling plot size on the estimation of structural indicators in old-growth forest stands. For Ecol Manag 346:89–97
Meyer P (1999) Bestimmung der Waldentwicklungsphasen und der Texturdiversität in Naturwäldern. Allg Forst-u J-Ztg 170:203–211
Meyer P, Ackermann J, Balcar P, Boddenberg J, Detsch R, Förster B, Fuchs H, Hoffmann B, Keitel W, Kölbel M, Köthke C, Koss H, Unkrig J, Weber J, Willig J (2001) Untersuchungen der Waldstruktur und ihrer Dynamik in Naturwaldreservaten. Arbeitskreis Naturwälder. Bund-Länder-Arbeitsgemeinschaft Forsteinrichtung. IHW-Verlag, Eching
Müller KM (1929) Aufbau, Wuchs und Verjüngung der südosteuropäischen Urwälder. Schaper, Hannover
Müller J, Bütler R (2010) A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. Eur J For Res 129:981–992
Müller-Using S, Bartsch N (2009) Decay dynamic of coarse and fine woody debris of (Fagus sylvatica L.) forest in Central Germany. Eur J For Res 128:287–296
Nagel TA, Svoboda M (2008) Gap disturbance regime in an old-growth Fagus–Abies forest in the Dinaric Mountains, Bosnia-Herzegovina. Can J For Res 38:2728–2737
Nagel TA, Svoboda M, Kobal M (2014) Disturbance, life history traits, and dynamics in an old-growth forest landscape of southeastern Europe. Ecol Appl 24(4):663–679
Neumann M (1979) Bestandesstruktur und Entwicklungsdynamik im Urwald Rothwald/NÖ und im Urwald Čorkova Uvala/Kroatien. PhD thesis, Univ. f. Bodenkultur, Wien
Oliver CD, Larson BC (1996) Forest stand dynamics. Wiley, New York
Paluch JG (2007) The spatial pattern of a natural European beech (Fagus sylvatica L.)—silver fir (Abies alba Mill.) forest: a patch mosaic perspective. For Ecol Manag 253:161–170
Peck JE, Commarmot B, Hobi ML, Zenner EK (2015) Should reference conditions be drawn from a single 10 ha plot? Assessing representativeness in a 10,000 ha old-growth European beech forest. Restor Ecol 23:927–935
Peterken GF (1996) Natural woodland ecology and conservation in northern temperate regions. Cambridge University Press, Cambridge
Peters R (1997) Beech forests. Geobotany 24. Springer, Berlin
Petráš R, Pajtík J (1991) Sústava Česko-slovenských objemových tabuliek drevín. Lesnícky Časopis 37:49–56
Pretzsch H (2009) Forest dynamics, growth and yield. Springer, Berlin
Přívětivý T, Janík D, Unar P, Adam D, Král K, Vrška T (2016) How do environmental conditions affect the deadwood decomposition of European beech (Fagus sylvatica L.)? For Ecol Manag 381:177–187
Remmert H (1991) The mosaic-cycle concept of ecosystems—an overview. In: Remmert H (ed) The mosaic-cycle concept of ecosystems. Springer, Berlin, pp 1–21
Röhrig E, Bartsch N, Lüpke B (2006) Waldbau auf ökologischer Grundlage. Ulmer, Stuttgart
Schuck A, Meyer P, Menke N, Lier M, Lindner M (2004) Forest biodiversity indicator: deadwood—a proposed approach towards operationalising the MCPFE indicator. In: Marchetti M (ed) Monitoring and indicators of forest biodiversity in Europe—from ideas to operationality. European Forest Institute, EFI PROC. 51, pp 49–77
Spies TA, Franklin JF (1996) The diversity and maintenance of old-growth forests. In: Szaro RC, Johnson DW (eds) Biodiversity in managed landscapes: theory and practice. Oxford Universisty Press, New York, pp 296–314
Tabaku V (2000) Struktur von Buchen-Urwäldern in Albanien im Vergleich mit deutschen Buchen-Naturwaldreservaten und-Wirtschaftswäldern. PhD thesis, Cuvillier Verlag, Göttingen
Tabaku V, Meyer P (1999) Lückenmuster albanischer und mitteleuropäischer Buchenwälder unterschiedlicher Nutzungsintensität. Forstarchiv 70:87–97
Trotsiuk V, Hobi ML, Commarmot B (2012) Age structure and disturbance dynamics of the relic virgin beech forest Uholka (Ukrainian Carpathians). For Ecol Manag 265:181–190
Watt AS (1947) Pattern and process in the plant community. J Ecol 35:1–22
Westphal C, Tremer N, Oheimb G, Hansen J, Gadow K, Härdtle W (2006) Is the reverse J-shaped diameter distribution universally applicable in European virgin beech forests? For Ecol Manag 223:75–83
Winter S, Brambach F (2011) Determination of a common forest life cycle assessment method for biodiversity evaluation. For Ecol Manag 262:2120–2132
Zeibig A, Diaci J, Wagner S (2005) Gap disturbance patterns of a Fagus sylvatica virgin forest remnant in the mountain vegetation belt of Slovenia. For Snow Landsc Res 79:69–80
Zenner EK, Peck JE, Hobi ML, Commarmot B (2014) The dynamics of structure across scale in a primaeval European beech stand. Forestry 88:180–189
Zenner EK, Peck JE, Hobi ML, Commarmot B (2016) Validation of a classification protocol: meeting the prospect requirement and ensuring distinctiveness when assigning forest development phases. Appl Veg Sci 19:541–552
Acknowledgements
The support by the Stemmler Foundation is gratefully acknowledged. We are also grateful to the Poloniny National Park authority, the local forest administrations and the Ministry of Defence of the Slovak Republic for the permits to conduct the study and for technical support during the fieldwork. For organizational and technical support we also like to thank Viliam Pichler and his working group at the Technical University of Zvolen. Many thanks for assistance in the field to Matthias Steckel. We thank two anonymous reviewers for highly useful comments and suggestions on the manuscript.
Funding
This study was supported by the Stemmler Foundation, a member of the Stifterverband für die Deutsche Wissenschaft, Essen, Germany (Grant Number T206/23493/2012).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Claus Bässler.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Feldmann, E., Glatthorn, J., Hauck, M. et al. A novel empirical approach for determining the extension of forest development stages in temperate old-growth forests. Eur J Forest Res 137, 321–335 (2018). https://doi.org/10.1007/s10342-018-1105-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-018-1105-4