
Abstract
Despite the potential utility of a biogeographical approach to understanding the naturalization of exotic species, studies using this approach are scarce. Eucalyptus globulus is an economically important Australian tree species that has become naturalized in a number of countries where it was introduced. Portugal is an ideal territory to study the naturalization of E. globulus owing to: a long introduction history, the antipodal location compared to Australia and the large cultivated area. Wildling density was assessed in 116 E. globulus plantations in central Portugal through 213 transects established along plantation borders. Boosted regression trees were used to model the influence of plantation-scale variables. Results from this survey were compared with data obtained in plantations from seven Australian regions, where a similar sampling protocol had been used. In Portugal, wildlings were more abundant in plantations that were: located in moist aspects, coppiced, with older tree stems and corresponding to intermediate site growth indexes. The overall density (127 plants ha−1) was 14.9 times higher than in the Australian estate, but this ratio was reduced to 3.1 in a more comparable subset of unburnt, first rotation plantations. A generalized linear model fitted using a dataset combining the two surveys showed that country influenced wildling density, together with plantation rotation and stem age. These results provide insights into the naturalization of a widely cultivated tree species, pointing to a fundamental role of the introduction history, possibly acting along with the biogeographical characteristics of the introduced range.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Human-mediated transportation of organisms between different geographical regions is an ancient process. This movement has often been bolstered by human interests in particular species (Almeida and Freitas 2001; Wilson et al. 2009). At the global level, introduced species were used in 29% of afforestation and in 36% of reforestation programmes, between 2003 and 2007 (FAO 2010). Human activity resulted in the naturalization (sensu Richardson et al. 2000) of at least 3.9% of all known vascular plants outside their original territories (van Kleunen et al. 2015). The naturalization of these species is part of the invasion continuum, and although not all naturalized species become invasive, all invasive species first become naturalized (Richardson and Pyšek 2012).
Progression along this continuum involves several phases (Williamson 2006; Pyšek et al. 2009) and Blackburn et al. (2011) highlight that naturalization is essentially a multi-generational population process. Drivers of naturalization are context dependent and may interact (Lozon and MacIsaac 1997; Richardson and Pyšek 2006; Essl et al. 2011). Features related to introduction history, like residence time and propagule pressure, are strong determinants of naturalization potential (Lockwood et al. 2005; Pyšek and Jarošík 2005, Pyšek et al. 2009; Simberloff 2009). Other important factors include the susceptibility of the recipient community to invasion, and environmental and anthropic factors (Catford et al. 2009; Blackburn et al. 2011; Procheş et al. 2012; Richardson and Pyšek 2012).
The particular conditions created in cultivated areas can either promote or prevent reproduction and seedling establishment from introduced plants (Richardson and Rejmánek 2011). The purpose of the introduction, for example agriculture or forestry, often guides the selection of provenances for cultivation, aiming at high survival, growth, productivity and/or reproductive output in the target habitat (Richardson 1998; Thuiller et al. 2006; Rejmánek and Richardson 2011; Dodet and Collet 2012). Cultivated forests can induce a high propagule pressure through the accumulation of large amounts of seed which may be released into the environment, especially after harvesting of reproductively mature plantations (Wilson et al. 2009; Rejmánek and Richardson 2011). Moreover, cultivation inherently protects and maintains founder populations from which new foci can be continuously recruited (Mack 2000; Lockwood et al. 2005; Wilson et al. 2009). Concurrently, the disturbance associated with cultivation often fosters the establishment of exotic plants (Lozon and MacIsaac 1997). However, management operations intended to improve crop conditions, for instance removing crop competitors or reducing fire risk, may kill or at least injure offspring of cultivated individuals (e.g. Águas et al. 2014). Even harvest can have detrimental side effects on recruited plants, depending on the harvesting techniques (McIver and Starr 2000; Gonçalves et al. 2004).
Pinus and Eucalyptus are the most important genera used as exotics in forestry (Rejmánek and Richardson 2011). Eucalyptus globulus Labill. is probably the most widely cultivated eucalypt in temperate regions of the world (Jacobs 1979; Rejmánek and Richardson 2011). This species is native to south-eastern Australia, but it has been planted in many other world regions particularly in late twentieth century to supply the pulp and paper industry (Potts et al. 2004). In 2008, the global area of E. globulus was estimated to be around 2.3 million ha (Rejmánek and Richardson 2011).
The naturalization of E. globulus outside its native range has been reported by several sources (e.g. Sanz-Elorza et al. 2004; Cal-IPC 2006). The drivers of wildling establishment close to plantations were studied in seven planting zones in Australia (mostly outside the native range) by Larcombe et al. (2013), and in Portugal by Catry et al. (2015) and by Fernandes et al. (2016). These studies found important regional-scale variables that influence wildling establishment, including temperature seasonality and rainfall. The only significant plantation-scale variable identified was the age of stems, in the Australian study. Another study by Águas et al. (2014) assessed the role of some plantation-scale variables in Portugal, but it was only restricted to burnt areas. This assessment was not targeted to professionally managed plantations and therefore included a wide range of E. globulus-dominated stand types. All three Portuguese studies (Águas et al. 2014; Catry et al. 2015; Fernandes et al. 2016) considered that poor or lack of management may create more favourable conditions for wildling establishment. Therefore, it is expected that in industrial plantations where standard silvicultural practices involve more intensive management, wildling recruitment should be lower and more comparable with similar plantations in other parts of the world. Larcombe et al. (2013) found that E. globulus in industrial plantations in Australia was a low-risk invader, given the overall low density of wildlings detected (8 plants ha−1) within a 20-m strip along the stand edge. The higher densities found in Portugal (102 plants ha−1 inside; 67 plants ha−1 outside) by Fernandes et al. (2016) within the first 15 m on either side of the border of managed plantations may be related to stand characteristics but also to the very different ecological context, compared to Australia where the genus Eucalyptus is native. These biogeographical differences could arise through a reduced competitive environment (e.g. Xiao et al. 2016) or through enemy release (e.g. herbivores or pathogens) in the introduced range (Keane and Crawley 2002). However, despite the importance of a biogeographical approach to better understand the mechanisms that enable the higher success of some exotic species in their introduced ranges, studies using this approach are scarce (Hierro et al. 2005; Parker et al. 2013). Therefore, wider comparative studies are required for better understanding the ecology of plant naturalization and invasion. In the specific case of E. globulus, Portugal is an ideal territory for such an assessment since it has a long introduction history, it is one of the farthest regions from the native territory, and it has about 1/3 of the global area of plantations of this species (Rejmánek and Richardson 2011; ICNF 2013). The work by Fernandes et al. (2016) provides an interesting basis for such assessment, but it was not designed with that purpose, and does not address important plantation-scale variables such as stem age, rotation or site index.
The present work aims at filling some of the aforementioned knowledge gaps, by studying the influence of plantation-scale drivers on the abundance of E. globulus wildlings in Portugal. The study adopted a sampling methodology very similar to Larcombe et al. (2013) in Australia, allowing comparison of the results from the two antipodal regions. Therefore, this study specifically aims to: (1) assess the plantation-scale drivers of wildling establishment in a representative sample of E. globulus plantations in Portugal; (2) compare the results obtained in Portugal with data previously obtained in plantations from Australia, managed under a similar silvicultural regime; (3) use this comparison to provide insights into the naturalization process of one of the world’s most widely cultivated tree species.
Methods
Sampling
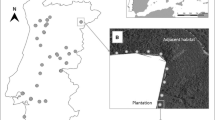
The sampling was undertaken in the Lower Tagus valley in central Portugal (Fig. 1) a region representative of much of the area managed intensively for pulpwood production in central-southern Portugal. The latitude of sampling ranged from 39°32′02′′ to 38°47′04′′ (83 km) and longitude ranged from −7°46′52′′ to −9°00′08′′ (106 km). This region has relatively homogeneous climate, topography and soils. Most of the sampled plantations were located on Podzols (66%) or Cambissols (29%) and a few (5%) on Lithosols (Cardoso et al. 1971). The studied region includes hilly areas and river flats with altitudes ranging between 30 and 307 m above sea level (according to GPS registrations during field work). Climate is Mediterranean with clear seasonality of temperature and precipitation, corresponding to type Csa (temperate with dry or hot summer) of the Köppen classification (AEMET and IM 2011). The region has a considerable area occupied by eucalypt plantations. The inspection of land cover maps (IGP 2010) within 10 km buffers centred on each sampling unit showed that E. globulus plantations correspond to 32% of forest occupation and to 15% of the total surface in these buffers (Fig. 1). According to our knowledge of the area, this indicator of planting intensity can be considered representative of the whole sampled region.

Location of sampled transects in Portugal. The E. globulus distribution map was based on land cover cartography (IGP 2010)
All sampled E. globulus plantations were owed by Altri Florestal SA (hereafter Altri), one of the two major Portuguese pulp companies. Altri manages an overall E. globulus estate of about 63,500 ha, roughly corresponding to 8% of the total area of eucalypt-dominated plantations in Portugal and to 40% of the total surface managed by pulp companies (CELPA 2015). Altri’s plantations are usually coppiced every 10–14 years, over 2–3 rotations. Sampling was planned to include a balanced number of plantations from all rotations.
The field work protocol was based on Larcombe et al. (2013), in order to compare wildling densities between Portuguese and Australian plantations. Our sampling units were transects established along plantation borders, aimed at assessing the number of wildlings per surface unit. A plantation border was defined as a virtual line that crossed the boles of neighbour border trees. Border segments were eligible for sampling as long as they were: neighbouring a different land use; located at least 50 m away from another eucalypt plantation at reproductive stage; as well as homogeneous in terms of aspect and slope. More than one transect could be sampled along the border of each plantation. Given that high densities of wildlings were frequent, plant counts were performed using plots (sub-samples) set along each transect, and not along the whole transect as in Larcombe et al. (2013). Plots were squares (20 × 20 m) straddling the plantation border, divided in two halves. One half-plot (10 × 20 m) was established inside the plantation, and the other half was established outside. The first plot of each transect was established 50 m after the starting point, and the remaining were established every 200 m farther along the transect. The geographical coordinates of plot centres, start point and end point of each transect were registered with a handheld GPS. The numbers of plants inside and outside the plantation were recorded separately. Wildlings were distinguished from resprouts or planted individuals by checking their position and their size relatively to the planted trees, and by checking the presence of a lignotuber. Conservative criteria were used, so that only plants with strong evidence of being wildlings were retained. In total, 514 plots were sampled along 213 transects established in 116 E. globulus plantations. The average number of transects per plantation was 1.84 ± 0.12 (min = 1, med = 1, max = 8). The average number of plots per transect was 2.41 ± 0.10 (min = 1, med = 2, max = 10).
Variables and datasets
Seven plantation-scale variables were used to model the abundance (density) of wildlings (Table 1) in the sampled region. These variables included information provided by Altri on: the developmental stage of the plantation (age, rotation), site index (see below), understory management and topography. Since tree age (corresponding to stump age in coppiced plantations) is correlated with the number of rotations, we only retained the latter for further analysis (variable ROTATION) and time-since-last coppice/planting was used as the age of tree stem (variable STEMAGE). Site index in E. globulus plantations is the height of dominant trees at 10 years. The original values (min = 9.0 m, median = 18.8 m, max = 22.5 m) were reclassified according to three categories (variable SIcat) with similar frequencies: low (<18 m); medium (18 m ≤ SIcat < 20 m); and high (≥20 m). The topographical information provided by Altri was originally obtained from 1:25,000 cartography produced by the Portuguese Army Geographical Institute (IGeoE). Altitude (variable ALTITUDE) was simply divided in two classes: low (≤100 m; a minimum of 30 m) and high (>100 m; a maximum of 307 m). Slope (variable SLOPE) was divided in three classes: flat (0%); gentle (≤15%); and moderate (>15%). This latter class included 12 transects with slope >30%. Topographical aspect (variable ASPECT) was defined by two classes: wet (North and West) and dry (South, East). Flat slopes were discarded from this variable. Management information was considered relevant due to the potential disturbance caused to wildling establishment. Management information (years 2009–2014) was also checked with local managers and field observations. Two types of management operations (disturbances from an ecological point of view) were considered: mechanical cutting (tractor with rotary mower) and tillage (tractor with disc harrow). For analysis purposes, only the last disturbance was retained for each transect, as it was considered the most relevant to explain wildling density. Therefore, management information was condensed in one single variable LD—Last Disturbance (None, Cut or Till). The “None” level includes transects in plantations where no management operations had been registered over the considered period. The average time-since-last intervention was 2.0 years for cutting and 0.7 years for tillage.
Data analysis
Collinearity among explanatory variables was checked by computing their VIF—variance inflation factors (function corvif from the R package AED; Zuur et al. 2009). VIF values were all below 1.5, so all variables were retained in further procedures. Wildling density (plants ha−1) was used as a dependent variable in a model developed using Boosted regression trees—BRT (Elith et al. 2008) fitted with a Poisson distribution (R package dismo; Hijmans et al. 2016). BRT modelling is a machine-learning technique that is able to automatically identify and model interactions, it is flexible in handling different types of data and predictors, and it has high predictive power compared to other methods (Elith et al. 2008). BRT combine two basic algorithms: regression trees and boosting. Regression trees are built through successive binary splits of the response variable; each split results in two groups as homogeneous as possible, according to a simple rule, based on the most influential variable (De’ath and Fabricius 2000). Boosting is a method that combines many simple models (in this case many regression trees) which allows an improved predictive performance (Elith et al. 2008). The relative importance of each explanatory variable is based on how often that variable is selected for splitting.
Given the recurrent disturbance of the outer half-plot, owing to the existence of roads and firebreaks close to the plantation border, modelling included only the inner half allowing obtaining more clear effects from the variables of interest. An initial model included the number of plots in each transect as a covariate to enable the detection of sampling bias. Modelling procedures followed the recommendations from Elith et al. (2008) and Elith and Leathwick (2016). The model for inside density was performed using: 2500 trees obtained from cross-validation; a bag fraction of 0.5 (proportion of data to be randomly selected at each step); a learning rate of 0.001 (the contribution of each tree to the growing model); a step size of 50 (initial number of trees); and a tree complexity of 5 (the number of interactions within a single tree). Model performance was assessed through the percentage of explained deviance. Differences in wildling density between the inner and the outer half plots were assessed using a Wilcoxon test.
In a separate analysis, data from Portuguese stands were pooled with those from plantations of seven planting zones (hereafter “regions”) in Australia, studied by Larcombe et al. (2013). Similarly to this latter work, wildling density was considered within the whole plot (inside and outside the plantation). In order to assess the relative influence of the country factor, a generalized linear model (GLM) with a negative binomial error distribution and a log link (in order to account for overdispersion) was fitted to the wildling density data. Variables Country (two levels: Portugal and Australia), Stem age (continuous variable) and Rotation (two levels: first rotation and coppiced plantations), were used as explanatory variables from the joint dataset combining the results of the Portuguese and the Australian surveys. In this analysis we discarded burnt transects because this variable was unevenly distributed, being relevant only in one Australian region (Gippsland). This model was simplified using a backwards stepwise procedure where single nonsignificant terms were removed and the subsequent simplified model was compared with the previous model using Chi-square tests. Higher-order interaction terms were removed first, followed by lower order terms, starting with those with the highest p value. Diagnostic plots of residuals versus fitted values and QQ-plots were used to check, respectively, for heteroscedasticity and residual normality.
Wilding density in Portugal was compared separately to each Australian region. This pairwise approach was used because variation in wildling density, as well as vast distances between regions, meant an average wildling value for Australia could be misleading. Mann–Whitney tests were used to perform these comparisons, and the same procedure was used to test differences in stem age. All transects from each region were included to perform these pairwise comparisons. No p value adjustment was introduced in the results of the multiple tests (e.g. Rothman 1990). All analyses were performed using R software (R Core Team 2015).
Results
Wildling establishment in Portugal
Overall, 2604 wildlings were counted across the 213 transects. Wildling density varied widely, showing a strongly right-skewed distribution, with 38% of zeros. Mean wildling density was 128.4 ± 24.0 plants ha−1 (min = 0, med = 25, max = 4000). Inside densities had a mean value of 180.8 ± 27.5 plants ha−1 (min = 0, med = 25, max = 2950). Outside densities showed higher data dispersion with a mean value of 76.1 ± 31.4 plants ha−1 (min = 0, med = 0, max = 6375). According to the Wilcoxon test, inside wildling densities were significantly higher than those outside (p < 0.001). However, higher densities were found further than 10 m from the plantation edge (data not shown); plants were from a range of different sizes, with about half of the individuals above 1.3 m height (data not shown).
The BRT model explained 43% of deviance and showed that the most influential variables explaining wildling density inside plantations were: ASPECT, ROTATION, STEMAGE and SIcat (Fig. 2). These variables accounted for 80% of importance in the model. The remaining three variables were all below 10% in importance. An initial checking run including the number of plots per transect showed that this covariate had no particular effect on the model results (an importance of only 6.3% with no particular trend). The BRT model showed that wildlings are more abundant in moist aspects, in second and third rotations, in plantations with older tree stems and in intermediate site index categories. The results from the remaining variables were not conclusive, since the fitted function values were relatively close among factor levels.

Partial dependence plots produced by the BRT model showing the relationships between the seven studied variables and wildling density. See Table 1 for variables description
A comparison with Australia
The characterization of the surveys performed in Portugal and in the seven Australian regions studied by Larcombe et al. (2013) is presented in Table 2. Although the length of surveyed plantation border was similar between Portugal and some Australian regions, the number of transects was in all cases much higher in the former. A considerable difference was also noticed: in the proportion of transects in coppiced plantations (much higher in Portugal); in the mean age of stems (lower in Portugal; all p < 0.001); and in the proportion of transects in replanted plantations. According to Altri registrations, plantations in Portugal were nearly all (>95%) replanted in sites previously occupied by E. globulus plantations. It must be noticed that the concept of second rotation adopted in Larcombe et al. (2013) included both the coppiced plantations and the replanted plantations. Here, the concept of second and third rotation refers only to plantations that were coppiced once and twice, respectively. Plantations surveyed in only one Australian region (Gippsland) were considered to be within the local native range of E. globulus, and this was also the region with the highest number of transects in burnt plantations. The overall wildling density (total number of wildlings divided by the total sampled surface) in Portugal was 6.4 times higher than the highest overall density obtained in Australia (in the Manjimup region) and 14.9 times higher than in the Australian regions altogether. In a subset of unburnt first rotation plantations, this ratio was reduced to 3.1.
The generalized linear model (Table 3) shows the importance of Country in explaining wildling density, within the ensemble of the three explanatory variables. The model coefficients and respective significance levels for Country and Rotation were similar and both considerably higher than the coefficient of Stem age. The GLM showed that wildling density is higher in Portugal, in coppiced plantations and in plantations with older stems (Fig. 3). Interactions among variables were not significant and were not included in the final model. The model fitted with these three variables was able to explain 22.2% of deviance. When comparing the wildling densities found in Portugal against each Australian region separately, the Mann–Whitney tests showed that wildling densities in Portugal were significantly higher than in all Australian regions except Portland, but the detected differences varied widely in their significance (Fig. 4).

Relationships between stem age and wildling density (plants ha−1) in a first rotation unburnt plantations and in b coppiced plantations, surveyed in Portugal and Australia, according to a GLM fitted using a database combining data from this study and from Larcombe et al. (2013). Dashed thin lines correspond to 95% confidence intervals. Note the different scales of the y-axis in each graph

Wildling density (plants ha−1; mean ± SE) in Portugal and in seven Australian regions using data from this study and from Larcombe et al. (2013). GIP stands for Gippsland, GRAMPS stands for Grampians, and TAS stands for Tasmania. Asterisks represent the result of pairwise comparisons through Mann–Whitney tests, between Portugal and each Australian region (* p < 0.05, ** p < 0.01, *** p < 0.001; ns nonsignificant)
Discussion
Plantation-scale drivers of wildling establishment
We found that plantation-scale factors explained a significant component of the variation in wildling density along the border of Portuguese E. globulus plantations. Topographical aspect was the most important driver of wildling density. Wildling density was higher on moist than on dry aspects. The wildling preference for moist/cool aspects might be related with edaphoclimatic constraints. The Portuguese study region is located on the transition zone between Atlantic and Mediterranean climates. Mean temperature in the vast majority of sampled sites in this region is close to or above the maximum found in the planting zones of E. globulus in Australia (Larcombe et al. 2013). More than half of the studied area in Portugal has annual precipitation close to the low limit for its natural distribution (Kirkpatrick 1975; Jacobs 1979; Turnbull and Pryor 1984), and precipitation is clearly seasonal with a dry season of four months. This contrasts with the preference for reduced seasonality in precipitation (Larcombe et al. 2013), especially if annual precipitation is close to its low limit (Kirkpatrick 1975). Therefore, drought (especially during summer) may limit wildling establishment in this region. This is in agreement with several references that identify either drought or frost as the main limiting factors for E. globulus development in different regions of Portugal (Almeida et al. 1994; Ribeiro and Tomé 2000; Alves et al. 2012). In addition, drought can be aggravated in areas where soils have little water holding capacity, such as podzols that are the commonest soil type in the southern part of the sampled region. Actually, in regions where climate induces reduced water availability, E. globulus persists only where mitigating factors exist such as southern aspects (in southern hemisphere), along streams (Kirkpatrick 1975) and in deep moist soils (Jacobs 1979). Moreover, González-Muñoz et al. (2011) showed that this species is more successful under moderate light and high water availability conditions, normally associated with deeper soils. In Portugal, these conditions are more frequently found in northern and western aspects (Marques and Mora 1992; Boer et al. 1996).
There may be several reasons why wildings were more abundant in coppiced than in first rotation plantations. Firstly, coppiced plants have well-developed root systems and are likely to reach maximum reproductive capacity sooner than seed-grown first rotation plants. Secondly, site preparation following harvesting involves ploughing, and may result in fewer residual seed than in coppiced plantations where slash (canopy branches) carrying unopened capsules is left on the soil surface. Moreover, slash may act as a buffer against drought and predation, helping seeds that dehisce from the slash to germinate and establish (Fagg 2001). In line with this, Skolmen and Ledig (1990) note that prolific germination occurs after logging E. globulus, resulting in abundant seedling recruitment. In addition, seed might be released from capsules on the ground over an extended period. For example, Santos et al. (2015) found that capsules on the ground, presumably aged 0–6 years, may contain significant proportions of viable seeds. Although we could not assess the effect of previously existing plantations, because nearly all plantations in the Portuguese study region had been replanted, these mechanisms are equally valid in this case and contribute to explain the role of residence time in the naturalization of woody plants (Pysek et al. 2009).
Wildlings were also more frequent in stands with older stems, as was found in Australian plantations (Larcombe et al. 2013). This pattern could be related with the production and the release of seeds from trees. E. globulus starts seed production in the wild approximately seven years after germination or crown destruction (Kirkpatrick 1975), while, in fertilized plantations, trees can produce seeds at three-five years old (Turnbull and Pryor 1984; Barbour et al. 2008b).
Wildlings were more abundant in plantations with site index between 18 and 20 than in better or worse sites. It would appear as a logical preposition that wildlings would have more resources to thrive in the best sites. However, the sensitivity of E. globulus to asymmetrical intraspecific competition (Tomé et al. 1994) may reverse this preposition. Actually, under asymmetrical competition, the better suited the conditions are for E. globulus growth, the stronger this competition is and the earlier it begins (Tomé et al. 1994). In cases of very strong and/or prolonged asymmetrical competition among eucalypts, the smaller plants may not be able to thrive (Florence 1996). Hence, the low density of wildlings in high site index plantations may be a result of suppressive effects from the planted trees. There is also the possibility of a trade-off between growth and reproduction, with plants allocating more resources to growth in the highly competitive environments of eucalypt plantations located at the best sites (Suitor et al. 2010). Meanwhile, in the plantations with the lowest site indexes, wildlings might have faced difficulties to thrive owing to more adverse abiotic conditions, like drought or nutrient availability.
The results for the remaining three studied variables were not conclusive, given the low importance attained in the variable selection process for building the regression trees. Nonetheless, it is worth mentioning the unexpected lack of importance of the management variable (LD) and the very small differences between the different levels in the partial dependence plot. This result may have several explanations. Firstly, it may be partially explained by uncertainty in the management data. It is possible that not all recorded management operations have directly affected the relatively small area surveyed by each individual transect plot. For instance, the cutting and tillage operations were restricted to inter-rows, leaving the space between trees within the rows undisturbed and available for wildling establishment. In some cases, operations were not continuous (covering one inter-row in every two) potentially also contributing to the obtained results. Finally, as sampled sites are owned by a pulp company, the absence of cutting and tillage operations in some sites was likely due to low density of wildlings and other understorey vegetation, which would not require mechanical control. Although our results apparently contradict those found by Águas et al. (2014) in central and northern Portugal, the difference may be due to the different methods used to assess management, as well as to different sample characteristics. Additionally, the registration of management operations in Águas et al. (2014) was restricted to direct field evidence in areas that had been completely burnt and in many cases submitted to salvage logging operations and replanting. In such cases, the effect of tillage can be more effective in reducing wildling density, as it normally affects the whole surface. Moreover, the so-called unmanaged sites sampled by Águas et al. (2014) were more likely to be abandoned plantations as opposed to the ones sampled in the present study.
Nonetheless, disturbance factors definitely played a role in the resulting wilding density. In our study, wildling densities were generally higher inside the plantation than outside, similarly to the results obtained by Larcombe et al. (2013) and by Fernandes et al. (2016). The type and the frequency of disturbance inside plantations can be significantly different from what happens outside. Plantations in the study region were usually delimited by dirt roads and firebreaks. Besides disturbance caused by traffic and trampling, these areas are exposed to maintenance operations, mainly for fire prevention. This contrasts with the less recurrent disturbance regime inside the plantations, therefore, influencing the abundance of naturally regenerated eucalypt plants in either side of the plantation border. The existence of higher wildling densities in undisturbed areas beyond 10 m from the plantation edge (unpublished data), somehow, confirms this explanation. Another possible reason contributing for this difference may be related with a potentially more intense seed rain inside than outside the plantation.
Establishment success compared to Australian plantations
The overall density of naturally regenerated plants found in the Portuguese E. globulus plantations was substantially higher than in the comparable Australian study by Larcombe et al. (2013). On the one hand, Portugal has a longer history of E. globulus domestication and plantation establishment than Australia (Doughty 2000). On the other hand, eucalypt species have evolved over millions of years on the Australian continent (Thornhill et al. 2015). The comparison between the two studies thus involves different biogeographical contexts and different introduction histories in each specific region, including different sources of germplasm, planting intensity and residence time. While the observed difference is consistent with expectations of naturalization and invasion syndromes such as release from competition (Xiao et al. 2016) or enemies (Keane and Crawley 2002), there are numerous other factors which are confounded in the comparison between the countries, and decoupling these effects is a major challenge.
Part of the observed country differences could be explained by differences in plantation-scale factors related with introduction history, including the proportion of coppiced stands and the proportion of replanted stands. Both these factors are associated with a longer residence time and a higher propagule pressure, therefore, potentially contributing to the greater wildling density observed in Portugal. While the effect of replanting could not be tested, coppicing was proven to increase wilding density as discussed above. However, even after accounting for coppicing, we still found a large effect of country in the GLM analysis, suggesting other factors may be contributing to higher establishment in Portugal. These may include other non-studied plantation-scale factors but also other more general biogeographical features that determine the ecological context in each of the two countries.
Genetic factors may partly explain these differences. It has been shown that the Portuguese landrace had its origin predominantly in southern and south-eastern Tasmania (Lopez et al. 2001; Freeman et al. 2007), while Australian plantations are likely to have a larger component of their germplasm from the three Victorian subraces, including the Strzelecki Ranges subrace (Potts et al. 2014). It has been reported that this subrace has lower fecundity and produces smaller seeds than those from southern and south-eastern Tasmania (McGowen et al. 2004b). Bigger heavier seeds produce seedlings with faster initial growth and may improve establishment success (Simberloff 2009). Thus, differences in germplasm may result in Portuguese plantations producing more seed that is better able to establish than the Australian plantations, possibly contributing to differences in wildling densities between the two countries. This higher reproductive output of Portuguese plantations has potentially been compounded by a longer residence time, when compared to Australia. This is in agreement with Donaldson et al. (2014): reproductive output of exotic species increases wildling densities over time, and this effect gradually overwhelms the effects of other factors over the course of residence time.
It is also possible that natural and/or artificial selection has increased the fecundity and/or establishment capacity of the Portuguese germplasm relative to the Australian plantations. Eucalyptus globulus was introduced in Portugal in the 1850s where it was extensively planted from the mid-twentieth century (Radich 1994) and now corresponds to 26% (around 0.8 million ha) of the forested area (ICNF 2013). In contrast, almost all Australian plantations of E. globulus were established after 1996 and today cover around 0.5 million ha (Gavran and Parsons 2011; Gavran 2015), predominantly outside the natural range of the species (Barbour et al. 2008b; Larcombe et al. 2013). The short history of Australian plantations means that there has been limited opportunity for adaptation. Conversely, the Portuguese E. globulus landrace has been influenced by bottlenecks and genetic drift (Borralho et al. 2007), and by both natural (Almeida 1993; Freeman et al. 2007) and artificial (Lopez et al. 2001; Potts et al. 2004) selection which may have resulted in better adaptation of E. globulus to the plantation environment (Almeida 1993). There is certainly evidence that after introduction, natural selection has acted to improve E. globulus adaption to frost in Portugal (Almeida 1993) and to drought in Spain (Turnbull and Pryor 1984; Toro et al. 1998). While artificial selection has mainly focused on improving growth and wood quality traits (Borralho et al. 1992, 2007), flowering precocity and fecundity may have also been enhanced (Borralho et al. 1992; L. Leal, pers. com.). There are two sources of evidence, suggesting that the selection of mother-trees may have increased seed production in Portuguese plantations: (i) 37.6% of trees at plantation borders have capsules in our Portuguese study region (unpublished data; 25 plantations; stem age: 3–10 years), while in the Australian plantation estate, reproductive output of border trees ranges from 7.8% (age 3–10 years) to 29% (age 8–12 years) (Barbour et al. 2008a; Larcombe et al. 2014); and (ii) we observed small wildlings which were reproductive in the juvenile phase in the Portuguese survey, but these were never observed in the Australian survey (M. Larcombe pers. com.). Thus, better adaptation to the plantation environment in Portugal, including frost and drought resistance, may have increased establishment potential, while artificial selection of traits that improve seed production may have increased net fecundity. These signals of adaptation may contribute to increasing reports of naturalization (e.g. Silva and Marchante 2012; Silva and Tomé 2016) since large areas of plantations were established in the mid-twentieth century. Concomitantly, adaptation may also help explaining the differences found in wildling densities between Portugal and Australia. In addition, the synergy between longer residence time and larger reproductive output in Portugal than in Australia may strengthen this idea (Donaldson et al. 2014), no matter the origin of fecundity differences between Portuguese and Australian plantations (i.e. introduced germplasm or evolution).
It is unlikely that differences in the abiotic environment would favour seedling establishment in Portuguese compared to Australian plantations. Larcombe et al. (2013) found that establishment was more likely in conditions that were similar to the native range of E. globulus. Therefore, if abiotic factors are important, it would be expected that abiotic conditions in Portugal would be more similar to the species native range than are the Australian planting regions. However, the climate in the studied region in Portugal is relatively distinct from both the native region in Australia and the Australian plantation regions. In central, Portugal precipitation tends to be less and seasonality is clearly stronger than in those Australian regions (Kirkpatrick 1975; Larcombe et al. 2013), and these conditions are compounded by major differences in soil types. In fact, other regions in Portugal, where wildlings are even more abundant than in the region assessed in this study, have climatic conditions that resemble more the native region (Catry et al. 2015). Hence, abiotic conditions do not seem to be the main factor explaining the high wildling success observed in many sites in the current study.
On the contrary, biotic interactions may have contributed both to higher reproductive success and increased establishment potential in Portuguese plantation environments. These include: release from competition, enemy release and build-up of mutualistic relationships. Given the industrial setting of both study regions, it is unlikely that release from competition would drive large differences in establishment. One possible interaction that could influence reproductive success is release from competition for pollinators. Australian plantations may experience competition for pollinator resources with neighbouring native eucalypt species, and such pollinator competition may be absent from Portugal. On the other hand, despite E. globulus being exotic in the Iberian Peninsula, there are reports of bird (Calviño-Cancela and Neumann 2015) and bee (Apis mellifera) pollination, (H. Águas, pers. com.; Feás et al. 2010) and no obvious pollinator limitation. Indeed, Moncur et al. (1995) showed that the introduced A. mellifera improves seed production and maintains outcrossing of E. globulus in Tasmania. Flowers which are insufficiently fertilized abort (Suitor et al. 2008) and, while higher proportion of trees with capsules in Portuguese plantations most likely reflects selection for fecundity, the possibility of more effective pollination services than in the Australian plantations cannot be dismissed. However, pollinators of E. globulus are generalists in its native range (Hingston et al. 2004), and having related plant species nearby may actually increase pollinator services in Australia relative to Portugal where the pollinator community is likely to be less diverse. Conversely, the high density of plantations in Portugal (namely in the studied region) compared to the Australian estate may significantly facilitate pollination and fruit production.
The enemy release hypothesis may be more relevant to the current study than release from competition. Strauss (2001) reports that Australian eucalypt plantations acquire more pathogens and pests from neighbouring native forests than plantations of species exotic to Australia. In contrast, in Portugal E. globulus had no major pests or diseases for more than 100 years, with few local pests or pathogens adapting to eucalypts (Goes 1977). Only recently, has E. globulus been significantly attacked by species of phytophagous insects and fungi, most of which were inadvertently imported from Australia (Branco 2007). In Portugal, local browsing species do not feed on E. globulus (Catry et al. 2007), and its juvenile leaves are unpalatable to stock (Jacobs 1955). This is in marked contrast to the susceptibility of E. globulus juveniles to marsupial browsing in Australian plantations (Borzak et al. 2015). In fact, key biotic interactions that could contribute to the success of wildlings in Portugal are escape from pre- and post-dispersal seed predators. In Australia, wasps of the genus Megastigmus are significant predators of seed held in capsules in the canopy of E. globulus (McGowen et al. 2004a). Seed harvesting ants are also known to deplete dispersed eucalypt seeds and limit plant establishment in Australia (Ashton 1979) contrasting with recent findings in Portugal, showing that a large proportion of post-dispersed E. globulus seeds remains unharvested (unpublished data).
The build-up of mutualistic relationships may also be an important factor in the different recruitment profiles between the countries. As discussed above, site-level residence time is far greater in Portugal where multiple plantings and rotations of E. globulus have occurred on most sites. However, in Australia the plantations assessed were typically on sites never previously planted with E. globulus. This difference raises the possibility that plantations in Portugal have developed better mutualistic interactions with other organisms that promote wilding establishment, than Australian plantations where they have had less time to develop. Positive soil conditioning, where the presence of a species changes the biological, structural and physio-chemical properties of a soil so that establishment of that species is facilitated, is a prominent theory in the invasion/naturalization literature (Suding et al. 2013). There is evidence that eucalypt plantations do positively condition soils (Orozco-Aceves et al. 2015), and Marchante et al. (2001) found frequent ectomycorrhizal associations between several native fungi and E. globulus in Portugal. Ectomycorrhizal fungi are potentially a biotic component of soil conditioning, so it is possible that the build-up of mycorrhizal communities at the site level over time contributes to establishment of wildlings. It has previously been suggested that these associations may be crucial for exotic establishment of eucalypt seedlings (Rejmánek and Richardson 2011). It is also conceivable that these relationships could be especially important in our study region where poor soils and water stress are frequent. However, E. globulus plantations in Australia are likely to share mycorrhizal species, pest enemies and pollinators with neighbouring native species (Strauss 2001), meaning that Australian plantations may come pre-primed with mutualists. Therefore, in the current contrast the effect of mutualistic relationships is unclear but cannot be dismissed.
Conclusion
This work is the first to assess plantation-scale drivers of wildling establishment in industrial E. globulus plantations in Portugal and to compare wildling densities with those in regions within the native range of the Eucalyptus genus. Plantation-scale drivers seem to explain a considerable amount of variation in wildling density in the studied Portuguese region. Aspect was the most important variable, leading to the conclusion that water stress is an important factor limiting wildling establishment. The significant effect of stand rotation and stem age, also highlight the role of propagule pressure. At present, the observed wildling densities, with plants of different size, some of them showing reproductive capacity, strongly confirm the naturalization status of the species in Portugal. Meanwhile, the observed densities at some plantation borders may require control, as they increase fire hazard and may become new propagule sources. Indeed, the location of observed wildlings outside the plantation boundary points to a potentially more relevant issue—their escape to other land uses. Where there is such a concern, it has been suggested that this risk may be minimized with suitable management of plantation boundaries (Calviño-Cancela and Rubido-Bará 2013; Catry et al. 2015; Fernandes et al. 2016). Comparisons between Portugal and several Australian regions showed that wildling density in Portuguese plantations was considerably higher. Identifying the precise differences that are causing variation in wildling recruitment between two regions so far apart is virtually an impossible task. Nonetheless, it is very likely that factors related with introduction history (namely residence time) should partly explain the observed differences, possibly along with the biogeographical context of the two regions. In fact, biogeographical factors may be at least as important as introduction history, but further studies are needed to shed additional light on the main naturalization drivers of E. globulus.
References
AEMET, IM, (2011) Iberian climate atlas. Agencia Estatal de Meteorología and Instituto de Meteorologia, Madrid
Águas A, Ferreira A, Maia P, Fernandes PM, Roxo L, Keizer J, Silva JS, Rego FC, Moreira F (2014) Natural establishment of Eucalyptus globulus Labill. in burnt stands in Portugal. For Ecol Manag 323:47–56. doi:10.1016/j.foreco.2014.03.012
Almeida MH (1993) Estudo da variabilidade geografica em Eucalyptus globulus Labill., Universidade Técnica de Lisboa, Lisboa
Almeida JD, Freitas H (2001) The exotic and invasive flora of Portugal. Bot Complut 25:317–327
Almeida MH, Chaves MM, Silva JC (1994) Cold acclimation in eucalypt hybrids. Tree Physiol 14:921–932
Alves AM, Pereira JS, Correia AV (2012) Silvicultura—a gestão dos ecossistemas florestais. Fundação Calouste Gulbenkian, Lisboa
Ashton D (1979) Seed harvesting by ants in forests of Eucalyptus regnans F. Muell. in central Victoria. Aust J Ecol 4:265–277
Barbour RC, Crawford AC, Henson M, Lee DJ, Potts BM, Shepherd M (2008a) The risk of pollen-mediated gene flow from exotic Corymbia plantations into native Corymbia populations in Australia. For Ecol Manag 256:1–19. doi:10.1016/j.foreco.2008.03.028
Barbour RC, Otahal Y, Vaillancourt RE, Potts BM (2008b) Assessing the risk of pollen-mediated gene flow from exotic Eucalyptus globulus plantations into native eucalypt populations of Australia. Biol Conserv 141:896–907
Blackburn TM, Pysek P, Bacher S, Carlton JT, Duncan RP, Vc Jaroaík, Wilson JRU, Richardson DM (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26:333–339
Boer M, Del Barrio G, Puigdefábres J (1996) Mapping soil depth classes in dry Mediterranean areas using terrain attributes derived from a digital elevation model. Geoderma 72:99–118
Borralho N, Cotterill P, Kanowski P (1992) Genetic parameters and gains expected from selection for dry weight in Eucalyptus globulus ssp. globulus in Portugal. For Sci 38:80–94
Borralho NMG, Almeida MH, Potts BM (2007) O melhoramento do eucalipto em Portugal. In: Alves AM, Pereira JS, Silva JMN (eds) O eucaliptal em Portugal: impactes anbientais e investigalção científica. ISA Press, Lisboa, pp 61–110
Borzak CL, O’Reilly-Wapstra JM, Potts BM (2015) Direct and indirect effects of marsupial browsing on a foundation tree species. Oikos 124:515–524
Branco M (2007) Agentes patogénicos do eucalipto em Portugal. In: Alves AM, Pereira JS, Silva JMN (eds) O eucaliptal em Portugal: impactes ambientais e investigação científica. ISA Press, Lisboa, pp 255–282
Cal-IPC (2006) California Invasive Plant Inventory. California Invasive Plant Council. www.cal-ipc.org. Accessed 10 June 2011 Cal-IPC Publication 2006–02
Calviño-Cancela M, Neumann M (2015) Ecological integration of eucalypts in Europe: interactions with flower-visiting birds. For Ecol Manag 358:174–179. doi:10.1016/j.foreco.2015.09.011
Calviño-Cancela M, Rubido-Bará M (2013) Invasive potential of Eucalyptus globulus: seed dispersal, seedling recruitment and survival in habitats surrounding plantations. For Ecol Manag 305:129–137
Cardoso JC, Bessa MT, Marado MB (1971) Carta de Solos 1:1,000,000. Serviço de Reconhecimento e Ordenamento Agrário, Lisboa
Catford JA, Jansson R, Nilsson C (2009) Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers Distrib 15:22–40
Catry F, Bugalho M, Silva JS (2007) Recuperação da Floresta após o fogo. O caso da Tapada Nacional de Mafra. Centro de Ecologia Aplicada Prof. Baeta Neves/ISA, Lisbon
Catry FX, Moreira F, Deus E, Silva JS, Águas A (2015) Assessing the extent and the environmental drivers of Eucalyptus globulus wildling establishment in Portugal: results from a countrywide survey. Biol Invasions 17:3163–3181. doi:10.1007/s10530-015-0943-y
CELPA (2015) Boletim Estatístico 2014—Indústria Papeleira Portuguesa CELPA—Associação da Indústria Papeleira, Lisbon
R Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
De’ath G, Fabricius KE (2000) Classification and regression trees: a powerful yet simple technique for ecological data analysis. Ecology 81:3178–3192
Dodet M, Collet C (2012) When should exotic forest plantation tree species be considered as an invasive threat and how should we treat them? Biol Invasions 14:1765–1778. doi:10.1007/s10530-012-0202-4
Donaldson JE, Hui C, Richardson DM, Robertson MP, Webber BL, Wilson JR (2014) Invasion trajectory of alien trees: the role of introduction pathway and planting history. Glob Chang Biol 20:1527–1537
Doughty RW (2000) The Eucalyptus: a natural and commercial history of the gum tree. The Johns Hopkins University Press, Baltimore
Elith J, Leathwick J (2016) Boosted Regression Trees for ecological modeling. https://cran.r-project.org/web/packages/dismo/vignettes/brt.pdf
Elith J, Leathwick JR, Hastie T (2008) A working guide to boosted regression trees. J Anim Ecol 77:802–813
Essl F, Mang T, Dullinger S, Moser D, Hulme PE (2011) Macroecological drivers of alien conifer naturalizations worldwide. Ecography 34:1076–1084
Fagg P (2001) Eucalypt Sowing and Seedfall. Native Forest Silviculture Guideline No. 8. Department of Natural Resources and Environment, Victoria
FAO (2010) Global forest resources assessment 2010. FAO Forestry Paper. Food and Agriculture Organization of the United Nations, Rome
Feás X, Pires J, Estevinho ML, Iglesias A, De Araujo JPP (2010) Palynological and physicochemical data characterisation of honeys produced in the Entre-Douro e Minho region of Portugal. Int J Food Sci Tech 45:1255–1262
Fernandes P, Antunes C, Pinho P, Máguas C, Correia O (2016) Natural regeneration of Pinus pinaster and Eucalyptus globulus from plantation into adjacent natural habitats. For Ecol Manag 378:91–102. doi:10.1016/j.foreco.2016.07.027
Florence RG (1996) Ecology and silviculture of eucalypt forests. CSIRO, Collingwood
Freeman JS, Marques CM, Carocha V, Borralho N, Potts BM, Vaillancourt RE (2007) Origins and diversity of the Portuguese Landrace of Eucalyptus globulus. Ann For Sci 64:639–647
Gavran M (2015) Australian plantation statistics 2015 update. Australian Bureau of Agricultural and Resource Economics and Sciences, Canberra
Gavran M, Parsons M (2011) Australian plantation statistics 2011. Australian Bureau of Agricultural and Resource Economics and Sciences, Canberra
Goes E (1977) Os eucaliptos—ecologia, cultura produção e rentabilidade. Portucel, Lisboa
Gonçalves JLDM, Stape JL, Laclau J-P, Smethurst P, Gava JL (2004) Silvicultural effects on the productivity and wood quality of Eucalypt plantations. For Ecol Manag 193:45–61
González-Muñoz N, Castro-Díez P, Fierro-Brunnenmeister N (2011) Establishment success of coexisting native and exotic trees under an experimental gradient of irradiance and soil moisture. Environ Manag 48:764–773. doi:10.1007/s00267-011-9731-3
Hierro JL, Maron JL, Callaway RM (2005) A biogeographical approach to plant invasions: the importance of studying exotics in their introduced and native range. J Ecol 93:5–15
Hijmans RJ, Phillips S, Leathwick J, Elith J (2016) dismo: Species Distribution Modeling. R package version 1.0–15
Hingston A, Potts B, McQuillan P (2004) Pollination services provided by various size classes of flower visitors to Eucalyptus globulus ssp. globulus (Myrtaceae). Aust J Bot 52:353–369
ICNF (2013) IFN6 – Áreas dos usos do solo e das espécies florestais de Portugal continental. Resultados preliminares. Instituto da Conservação da Natureza e das Florestas, Lisboa
IGP (2010) Land-cover cartography for continental Portugal from 2007 (Carta de uso e ocupação do solo de Portugal Continental para 2007 (COS 2007)). Portuguese Geographical Institute (IGP), Lisbon
Jacobs MR (1955) Growth habits of the eucalypts. Forest and Timber Bureau, Canberra
Jacobs (1979) Eucalypts for planting. FAO Forestry Series, vol 11. FAO, Rome
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170
Kirkpatrick JB (1975) Natural distribution of Eucalyptus globulus Labill. Aust Geogr 13:22–35
Larcombe MJ, Silva JS, Vaillancourt RE, Potts BM (2013) Assessing the invasive potential of Eucalyptus globulus in Australia: quantification of wildling establishment from plantations. Biol Invasions 15:2763–2781. doi:10.1007/s10530-013-0492-1
Larcombe MJ, Barbour RC, Vaillancourt RE, Potts BM (2014) Assessing the risk of exotic gene flow from Eucalyptus globulus plantations to native E. ovata forests. For Ecol Manag 312:193–202
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228
Lopez G, Potts B, Dutkowski G, Rodriguez Traverso J (2001) Quantitative genetics of Eucalyptus globulus: affinities of land race and native stand localities. Silvae Genet 50:244–252
Lozon JD, MacIsaac HJ (1997) Biological invasions: are they dependent on disturbance? Environ Rev 5:131–144
Mack RN (2000) Cultivation fosters plant naturalization by reducing environmental stochasticity. Biol Invasions 2:111–122
Marchante E, Madeira M, Freitas H (2001) Avaliação da colonização ectomicorrízica em plantações experimentais de Eucalyptus globulus Labill. Revista das Ciências Agrárias 24:205–212
Marques MA, Mora E (1992) The influence of aspect on runoff and soil loss in a Mediterranean burnt forest (Spain). CATENA 19:333–344
McGowen M, Potts B, Vaillancourt R, Gore P, Williams D A genetic basis to the destruction of Eucalyptus globulus seed by wasps from the genus Megastigmus. In: Borralho NMG, Pereira JS, Marques C, Coutinho J, Madeira M, Tomé M (eds) Eucalyptus in a Changing World, Aveiro, Portugal, 11–15 October 2004a. IUFRO Conference. RAIZ, Instituto Investigação de Floresta e Papel, Aveiro, Portugal, pp 187–188
McGowen MH, Potts BM, Vaillancourt RE, Gore P, Williams DR, Pilbeam DJ The genetic control of sexual reproduction in Eucalyptus globulus. In: Borralho NMG, Pereira JS, Marques C, Coutinho J, Madeira M, Tomé M (eds) Eucalyptus in a Changing World, Aveiro, Portugal, 11–15 October 2004b. IUFRO Conference. RAIZ, Instituto Investigação de Floresta e Papel, Aveiro, Portugal, pp 104–108
McIver JD, Starr L (2000) Environmental effects of post-fire logging: literature review and annotated bibliography. General technical report. United States Department of Agriculture, Forest Service, Pacific Northwest Research Station, Portland
Moncur MW, Mitchell A, Fripp Y, Kleinschmidt GJ (1995) The role of honey bees (Apis mellifera) in eucalypt and acacia seed production areas. Commonw For Rev 74:350–354
Orozco-Aceves M, Standish RJ, Tibbett M (2015) Long-term conditioning of soil by plantation eucalypts and pines does not affect growth of the native jarrah tree. For Ecol Manag 338:92–99
Parker JD, Torchin ME, Hufbauer RA, Lemoine NP, Alba C, Blumenthal DM, Bossdorf O, Byers JE, Dunn AM, Heckman RW (2013) Do invasive species perform better in their new ranges? Ecology 94:985–994
Potts BM, Vaillancourt RE, Jordan GJ, Dutkowski GW, Costa e Silva J, McKinnon GE, Steane DA, Volker PW, Lopez GA, Apiolaza LA, Li Y, Marques C, Borralho NMG Exploration of the Eucalyptus globulus gene pool. In: Borralho NMG, Pereira JS, Marques C, Coutinho J, Madeira M, Tomé M (eds) Eucalyptus in a changing world, Aveiro, Portugal, 11–15 October 2004. IUFRO Conference. RAIZ, Instituto Investigação de Floresta e Papel, Aveiro, Portugal, pp 46–61
Potts B, Hamilton M, Pilbeam D (2014) Genetic improvement of temperate eucalypts in Australia. In: Ipinza R, Barros AS, Gutiérrez CB, Borralho N (eds) Mejoramiento Genético de Eucaliptos de en Chile. INFOR Instituto Forestal, pp 411–443
Procheş Ş, Wilson JR, Richardson DM, Rejmánek M (2012) Native and naturalized range size in Pinus: relative importance of biogeography, introduction effort and species traits. Glob Ecol Biogeogr 21:513–523
Pyšek P, Jarošík V (2005) Residence time determines the distribution of alien plants. In: Inderjit S (ed) Invasive plants: ecological and agricultural aspects. Birkhäuser Verlag, Basel, pp 77–96
Pysek P, Křivánek M, Jarošik V (2009) Planting intensity, residence time, and species traits determine invasion success of alien woody species. Ecology 90:2734–2744
Radich MC (1994) Uma exótica em Portugal. Ler História 25:11–26
Rejmánek M, Richardson D (2011) Eucalypts. In: Simberloff D, Rejmánek M (eds) Encyclopedia of biological invasions. University of California Press, Berkeley, pp 203–209
Ribeiro F, Tomé M (2000) Climatic classification of Portugal based on digitised climatic maps [in Portuguese]. Rev Cienc Agrar 23:39–50
Richardson DM (1998) Forestry trees as invasive aliens. Conserv Biol 12:18–26
Richardson DM, Pyšek P (2006) Plant invasions: merging the concepts of species invasiveness and community invasibility. Prog Phys Geogr 30:409–431
Richardson DM, Pyšek P (2012) Naturalization of introduced plants: ecological drivers of biogeographical patterns. New Phytol 196:383–396
Richardson DM, Rejmánek M (2011) Trees and shrubs as invasive alien species—a global review. Divers Distrib 17:788–809
Richardson DM, Pyšek P, Rejmánek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107
Rothman KJ (1990) No adjustments are needed for multiple comparisons. Epidemiology 1:43–46
Santos P, Matias H, Deus E, Águas A, Silva JS (2015) Fire effects on capsules and encapsulated seeds from Eucalyptus globulus in Portugal. Plant Ecol 216:1611–1621. doi:10.1007/s11258-015-0544-y
Sanz-Elorza M, Dana-Sánchez E, Sobrino-Vesperinas E (eds) (2004) Atlas de las Plantas Alóctonas Invasoras en España. Dirección General para la Biodiversidad, Madrid
Silva JS, Marchante H (2012) Post-fire management of exotic forests. In: Moreira F, Heras Jdl, Corona P, Arianoutsou M (eds) Post-Fire Management and Restoration of Southern European Forests. Springer, Dordrecht, pp 223–255
Silva JS, Tomé M (2016) Tasmanian blue gum in Portugal—opportunities and risks of a widely cultivated species. In: Krumm F, Vítková L (eds) Introduced tree species in European forests: opportunities and challenges. European Forest Institute, Freiburg, pp 352–361
Simberloff D (2009) The role of propagule pressure in biological invasions. Annu Rev Ecol Evol Syst 40:81–102
Skolmen RG, Ledig T (1990) Eucalyptus globulus Labill. Bluegum eucalyptus. In: Burns R, Honkala B (eds) Silvics of North America, vol 2. Agriculture Handbook vol 654. United States Department of Agriculture, Forest Service, Washington, D. C., pp 299–304
Strauss SY (2001) Benefits and risks of biotic exchange between Eucalyptus plantations and native Australian forests. Aust Ecol 26:447–457
Suding KN, Stanley Harpole W, Fukami T, Kulmatiski A, MacDougall AS, Stein C, Putten WH (2013) Consequences of plant–soil feedbacks in invasion. J Ecol 101:298–308
Suitor S, Potts BM, Brown PH, Gracie A, Gore PL (2008) Post-pollination capsule development in Eucalyptus globulus seed orchards. Aust J Bot 56:51–58
Suitor S, Potts BM, Brown PH, Gracie A, Rix K, Gore PL (2010) The impact of flower density and irrigation on capsule and seed set in Eucalyptus globulus seed orchards. New For 39:117–127
Thornhill AH, Ho SY, Külheim C, Crisp MD (2015) Interpreting the modern distribution of Myrtaceae using a dated molecular phylogeny. Mol Phylogen Evol 93:29–43
Thuiller W, Richardson DM, Rouget M, Proches S, Wilson JR (2006) Interactions between environment, species traits, and human uses describe patterns of plant invasions. Ecology 87:1755–1769
Tomé M, Tomé J, Araújo MC, Pereira J (1994) Intraspecific competition in irrigated and fertilized eucalypt plantations. For Ecol Manag 69:211–218
Toro M, Silió L, Rodriguez M, Soria F, Toval G Genetic analysis of survival to drought in Eucalyptus globulus in Spain. In: Proceedings of the 6th World Congress on Genetics Applied to Livestock Production, Armidale, NSW, Australia, 1998. pp 499–502
Turnbull JW, Pryor LD (1984) Choice of species and seed sources. In: Hillis WE, Brown AG (eds) Eucalypts for wood production. CSIRO Academic Press, Melbourne, pp 6–65
van Kleunen M, Dawson W, Essl F, Pergl J, Winter M, Weber E, Kreft H, Weigelt P, Kartesz J, Nishino M, Antonova LA, Barcelona JF, Cabezas FJ, Cardenas D, Cardenas-Toro J, Castano N, Chacon E, Chatelain C, Ebel AL, Figueiredo E, Fuentes N, Groom QJ, Henderson L, Inderjit Kupriyanov A, Masciadri S, Meerman J, Morozova O, Moser D, Nickrent DL, Patzelt A, Pelser PB, Baptiste MP, Poopath M, Schulze M, Seebens H, W-S Shu, Thomas J, Velayos M, Wieringa JJ, Pysek P (2015) Global exchange and accumulation of non-native plants. Nature. doi:10.1038/nature14910
Williamson M (2006) Explaining and predicting the success of invading species at different stages of invasion. Biol Invasions 8:1561–1568
Wilson JR, Dormontt EE, Prentis PJ, Lowe AJ, Richardson DM (2009) Something in the way you move: dispersal pathways affect invasion success. Trends Ecol Evol 24:136–144. doi:10.1016/j.tree.2008.10.007
Xiao S, Callaway RM, Graebner R, Hierro JL, Montesinos D (2016) Modeling the relative importance of ecological factors in exotic invasion: the origin of competitors matters, but disturbance in the non-native range tips the balance. Ecol Model 335:39–47
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
This research was developed in the scope of the “WildGum” project (FCT PTDC/AGR-FOR/2471/2012) funded by Fundação para a Ciência e a Tecnologia (FCT). A. A. and E. D. were supported by a PhD scholarship from FCT (SFRH/BD/76899/2011; PB/BD/113936/2015). A. A.’s and J. S. S.’s travels to Australia were funded by TRANZFOR programme from the European Commission—Marie Curie Actions. M.J.L’s and B.M.P’s travel to Portugal was funded by the "WildGum" project and a Maxwell Ralf Jacobs Scholarship Grant to M.J.L. We thank to Altri Florestal SA for providing access to their plantation estate and to data on plantation characteristics and management history, as well as for providing facilities for field work. Special thanks to Luis Ferreira for help with Altri’s database.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Christian Ammer.
Rights and permissions
About this article

Cite this article
Águas, A., Larcombe, M.J., Matias, H. et al. Understanding the naturalization of Eucalyptus globulus in Portugal: a comparison with Australian plantations. Eur J Forest Res 136, 433–446 (2017). https://doi.org/10.1007/s10342-017-1043-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-017-1043-6