
Abstract
The effectiveness of surveys of breeding birds varies due to multiple factors, with the primary being imperfect detection, which is particularly severe for elusive species. For example, the territory mapping method requires surveying an area multiple times a season to compensate for missing individuals during single surveys. Novel methods require much less effort in the field and include estimation of both detection probability and abundance corrected for individuals that went undetected. The aim of this study was to check if point counts and model-based results provide estimates similar to the ones from the territory mapping method. We studied the abundance of two forest birds—Goldcrest Regulus regulus and Firecrest R. ignicapilla—on three permanent census plots in the Białowieża Forest (E Poland). We compared abundance estimates resulting from the territory mapping method in its ‘standard’ (~ 10 visits) and intensive (~ 20 visits) approaches. We also performed point counts at the same plots using distance sampling methodology and hierarchical models in an attempt to get unbiased estimates by correcting for imperfect detection. We found that the standard territory mapping method produces much lower abundances than model-based estimates, which was particularly evident for the more numerous Firecrest. At the same time, results from point counts were more consistent with numbers from the intensive territory mapping. Our findings suggest that applying point counts and distance sampling models meet modern standards by considering various effects in abundance, availability and detection processes along with providing uncertainty of their estimates. We assume that our results might be applicable to other elusive species.
Zusammenfassung
Bestandsschätzungen anhand von Punktzählungen und Revierkartierungen: Vergleich verschiedener Ansätze für zwei Regulus-Arten
Die Effektivität von Brutvogelerhebungen variiert aufgrund mehrerer Faktoren, wobei der wichtigste Faktor die unzureichende Erfassung (d. h. die eingeschränkte Wahrnehmung durch den Menschen) ist, die besonders bei der Erfassung schwer erfassbarer Arten eine Rolle spielt. Die Methode der Revierkartierung erfordert beispielsweise, dass ein Gebiet mehrmals pro Saison begangen wird, um dem Problem entgegenzuwirken, dass Individuen bei einzelnen Erhebungen übersehen werden. Neuartige Methoden erfordern einen wesentlich geringeren Aufwand im Feld und umfassen sowohl eine Abschätzung der Entdeckungswahrscheinlichkeit als auch des Bestandes, der um nicht entdeckte Individuen korrigiert wurde. Ziel dieser Studie war es, zu überprüfen, ob Punktzählungen und modellbasierte Ergebnisse ähnliche Schätzungen liefern wie die Methode der Revierkartierung. Dazu untersuchten wir den Bestand von zwei Waldvogelarten – des Wintergoldhähnchens Regulus regulus und des Sommergoldhähnchens R. ignicapilla – auf drei festgelegte Zählflächen im Białowieża-Urwald (Ostpolen). Wir verglichen die Bestandsschätzungen, die auf Revierkartierungen in einer „standardisierter “ (~ 10 Begehungen) und einer intensivierten (~ 20 Begehungen) Form basieren. Außerdem haben wir auf denselben Flächen Punktzählungen durchgeführt, wobei wir die „Distance-Sampling-Methode “ und hierarchische Modelle verwendet haben, um über die Korrektur unzureichender Erfassungen unverfälschte Schätzungen zu erhalten. Wir fanden heraus, dass die standardisierte Revierkartierung zu wesentlich geringeren Bestandszahlen führt als modellbasierte Schätzungen, was besonders beim in größerer Anzahl vorhandenen Sommergoldhähnchen deutlich wurde. Gleichzeitig stimmten die Ergebnisse der Punktzählungen besser mit den Zahlen der intensiven Revierkartierung überein. Unsere Ergebnisse deuten darauf hin, dass die Anwendung von Punktzählungen und hierarchischem „Distance-Sampling “ modernen Standards entspricht, indem sie verschiedene Effekte in Bezug auf Bestand, Verwendbarkeit und Erfassungsprozesse berücksichtigt und zugehörige Schätzungsungenauigkeiten liefert. Wir gehen davon aus, dass unsere Ergebnisse auch auf andere schwer erfassbare Arten übertragbar sind.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Knowledge of wildlife abundance is a central part of ecology and forms the basis of applied protection and management (Ralph et al. 1993; Kéry and Royle 2016). In birds, apart from infrequent, easy-to-detect species, abundance estimation is far from trivial in wild populations, mostly because some individuals go undetected during field surveys. To avoid problems with imperfect detection, study plots or observation points are usually visited repeatedly during the breeding season to get the best available estimates of abundance. Older, once commonly used methods to deal with this issue in territorial species include repeated surveys coupled with mapping observations, known as the territory mapping (or spot-mapping) protocol (Tomiałojć 1980), nest search or estimators based on capture-mark-recapture approaches (Bibby et al. 2000; Gregory et al. 2004), but they all are costly in terms of effort. The combined territory mapping method, with 8–10 surveys per breeding season, was developed to handle this: successive surveys result in new detections or new contemporary records (individuals missing from previous surveys), so that the cumulative number of territories increases, as assumed, until all are detected and can be delimited. Multiplying surveys allows also to exclude non-territorial or transient birds (e.g. detected only early in the season) (Gnielka 1990). Therefore, this method is considered to be effective in assessing bird abundance, particularly if nest searches or behavioural cues (alarm calls or adults carrying food for offspring) are taken into account in the combined version (Tomiałojć 1980). Despite this, underestimations have been documented for several passerine species (Best 1975; Tomiałojć and Lontkowski 1989; Walankiewicz et al. 1997; Tomiałojć 2004). Whether they occur or not depends on the criteria applied to delimit territories. For example, in its standard version, as used in the Białowieża Forest for 49 years now, for most of the species at least three, clustered (close to each other) records over nine surveys are required to determine a territory in the absence of contemporary records (simultaneously singing males) (Wesołowski et al. 2022). It is easy to imagine, however, that species with quiet songs or low song rates or both might be detected just once or twice, despite holding permanent territories; in effect, with conservative criteria, abundance gets underestimated with such single or double records, that are merged with neighbouring territories or ignored (as in the case of insufficient information to delimit a territory). On the other hand, some migrating species can sing during stopovers which can lead to overestimating abundance (Gnielka 1992; Flade 1994). The number of territories is easy to determine if based on contemporary records, but this is only applicable to relatively numerous and vocally active species with loud songs, while, again, for more elusive species which sing infrequently and quietly, such records are rare and result in underestimations. Unfortunately, combined territory mapping has been tested for very few species: for some, the method appeared to provide reliable estimates, like Eurasian Wren Troglodytes troglodytes, Wood Warbler Phylloscopus sibilatrix or Common Chiffchaff P. collybita (Tomiałojć 1980). For others, it could not compensate for territories that could not be delimited due to non-detections of individuals in the field, which results in underestimation. Tests performed for Collared Flycatcher Ficedula albicollis (Walankiewicz et al. 1997), Song Thrush Turdus philomelos (Tomiałojć and Lontkowski 1989; Lõhmus 2022), Hawfinch Coccothraustes coccothraustes (Tomiałojć 2004), Blackbird T. merula and Mistle Thrush T. viscivorus (Lõhmus 2022) indicate underestimations of 15–35%. It shows that the territory mapping method might not be as accurate for some species and then the estimates reflect the minimal state rather than true abundance.
In recent decades, novel methods have been developed to help solve the issues of underestimation despite extensive fieldwork and to reduce the effort required with territory mapping methods. They promote time efficiency in the field, include estimation of both detection probability and abundance corrected for imperfect detection with hierarchical models and have become very popular in recent years (Kéry and Royle 2016). The simplest of these models, such as the binomial N-mixture model (Royle 2004), relies on assuming population closeness over the study course and can perform well if this assumption, along with other ones, are met (Bötsch et al. 2020; Neubauer et al. 2022). Despite this, numerous studies reported their sensitivity to violations of assumptions (Link et al. 2018). They are also known for not being particularly well suited to point counts, one of the commonest field protocols used in bird counts, primarily due to the so-called “area issue” (Kéry and Royle 2016; Neubauer and Sikora 2020). This can be solved by including a temporary emigration parameter in the generalised binomial N-mixture model version, which proved suitable for point counts (Chandler et al. 2011). An alternative is to use distance sampling protocols, where distances to detected birds allow the estimation of detection functions, describing the decline of detection probability with distance between the observer and birds (Buckland et al. 2015). Current models also aim to properly address sources of variation in both ecological and observation processes that generate data to yield valid results (Joseph et al. 2009; Kéry & Schaub 2012).
Here, we compared abundance estimates obtained from combined territory mapping with a novel, time-efficient approach based on point counts and hierarchical models for two elusive forest bird species: Firecrest Regulus ignicapilla and Goldcrest R. regulus. Males of these species sing infrequently (pers. obs.) and quietly, which makes them hard to detect and leads to underestimating their true abundance when territory mapping methods are applied. Moreover, finding the nests of these secretive species hardly ever happens without special searches, which makes it nearly impossible to improve estimates this way unless much increased effort is undertaken. The abundance of such elusive species can be underestimated by ‘standard’ methods, so studies on improving methods to estimate their abundance are needed. In the case of these two species, knowing their true abundance is particularly important to increase the reliability of existing (including national) population estimates and due to the contrasting population trends they exhibit over the last decades in Europe. Across the continent, Firecrest increases in numbers and expands its range towards north-east (https://www.pecbms.info/), while the population size of the more boreal Goldcrest is in decline in Central Europe. Goldcrest population decreases due to its association with Norway Spruce Picea abies, which is declining because of climate change (Treml et al. 2022) and its range is believed to retreat northward (Dyderski et al. 2018). These trends in both Regulus species are also clear in Poland: over the last 2 decades, the numbers of Firecrest have spectacularly increased by 5% per year, whereas Goldcrest numbers have declined by 1% per year (Chodkiewicz et al. 2019). Nearly identical trends were observed over the last ~ 50 years (1975–2019) in the Białowieża National Park (Wesołowski et al. 2022), where our research project was conducted.
The aim of this study was to apply novel methods of abundance estimation based on point counts and hierarchical models and compare the results with estimates from the territory mapping method. We also attempted to compare estimates from ‘standard’ territory mapping approach (9 surveys per plot within a season) with the intensive one (~ 20 surveys). We conducted territory mapping surveys on three permanent census plots in the Białowieża National Park (E Poland), monitored since 1975 (Wesołowski et al. 2022) along with point counts performed on the same plots using distance sampling protocols and hierarchical models to estimate density (or, equivalently, abundance), the latter since 2021. We hypothesised that (1) the territory mapping method in the ‘standard’ approach underestimates the number of territories compared to the intensive one due to expected, frequent nondetections and (2) the intensive mapping method can yield similar abundance estimates to those obtained from point counts analysed using methods that account for imperfect detection.
Methods
Study area
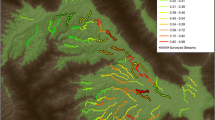
The study was conducted in the Białowieża Forest, primaeval temperate forest (~ 1600 km2) at the Polish-Belarussian border. This forest complex is divided between Poland (western part, around 45% of the area) and Belarus. Around 6000 ha on the Polish side forms the Strict Reserve belonging to the Białowieża National Park (BNP hereafter), protected since the late Middle Ages as a royal hunting area (Samojlik et al. 2013) and with no forest management since 1920s, when the reserve was established. BNP is a mostly deciduous woodland area, with continuous forest cover lasting for nearly 12,000 years and, currently, still a large share of high naturalness, pre-silvicultural stands (Jaroszewicz et al. 2019). Norway spruce Picea abies is frequent in all the forest types (Faliński 1986). Our study took place on three out of seven census plots which were established in 1975 in the Strict Reserve (Tomiałojć et al. 1977, 1984, Fig. 1). Two of our plots were lime-hornbeam forest types (plots C and W, 48 and 50 ha) and one riverine (plot K, 33.5 ha). On the remaining four plots (L—riverine, 25 ha, M—lime-hornbeam, 30 ha, NE and NW—coniferous forests, both 25 ha) we also performed point counts. For the most recent description of the forest habitats on the plots, see Wesołowski et al. (2022).

Study plots (with observation points embedded within them, black dots) in the Strict Reserve of the Białowieża National Park. Point counts were performed on all seven plots, while territory mapping on areas enveloped with red lines (entire plots or parts of the plots). The results from C, K and W plots, where territory mapping in two approaches was done were used in this study, while standard territory mapping only was performed on the NW plot
Study species
Firecrest is a common species breeding in central-western Europe and likewise in the whole area of Poland, where the population is estimated at around 258,000–539,000 breeding pairs (Chodkiewicz et al. 2019). The Goldcrest’s breeding range is the western Palearctic, from middle to upper temperate and boreal forests (Cramp 1998). In Poland, it occurs frequently in coniferous forests with population estimated at 522,000–811,000 breeding pairs (Chodkiewicz et al. 2019). These species exhibit opposite abundance trends in Poland. The Firecrest numbers have spectacularly increased nearly five-fold since 2000 while the Goldcrest abundance decreased by about one-third over this time (Chodkiewicz et al. 2019). In the BNP, the numbers of these two species were even more contrasting: in 2019, Firecrest numbers were 10 times higher than in 1975 whereas the abundance of Goldcrest was nearly 3 times lower than in 1975 (Wesołowski et al. 2022), so that the former outnumbers the latter currently (Fig. 2). Similar trends are visible across the whole Europe: for Firecrest the ten-year trend is + 12% and for Goldcrest it is –17% (https://www.pecbms.info/).

Annual changes in the total number of delimited territories of Firecrest (dark green circles) and Goldcrest (pale green triangles) at the seven monitoring plots in Białowieża National Park, Poland, 1975–2019. Trends visualised with loess curves. Data are from standard territory mapping (see Wesołowski et al. 2022) and given the results reported in the current paper, these absolute numbers can be underestimated, but should show trends properly
In both species, egg-laying in the first clutch depends on the geographical location and may start as early as in March. However, it usually occurs at the end of April into early May. They build nests as a three-layered cup of moss, lichens, feathers and hairs. Clutch size varies from nine to eleven in Goldcrest and from six to thirteen in Firecrest (Snow and Perrins 1998), and for both species second broods are common, but no information was available to us on whether the pairs use the same territory for the second brood or switch them. Second clutches commence in June–July. In Goldcrest, the female incubates eggs for 16–19 days to hatching, chicks leave the nest after 17 to 22 days. In Firecrest, it is 14–16 days and 8–10 days, respectively. In both species, both parents feed chicks and fledged young (Niethammer et al. 1991, Snow and Perrins 1998).
Firecrest and Goldcrest are characterised by quiet songs (80.4 dB, Winiarska et al. 2023) and low singing activity—they sing infrequently, making them elusive and hard to detect. The peak season of vocal (singing) activity in Poland falls in May for both species. Firecrest can be heard between early March and early October, and Goldcrest from late January to late September (https://www.ornitho.pl).
Field methods
Territory mapping method
On four plots (C, K, NW and W, see Fig. 1) we applied combined territory mapping with nine daytime surveys between early April and the second half of June, spaced by 8–11 days, as used in the Białowieża censuses since 1975 (Wesołowski et al. 2022, hereafter the standard approach). During the surveys, observers moved slowly across the plots and recorded all seen or heard singing birds on the maps (1:1000 scale), with additional details (i.e. pair seen, birds with nest material, alarm calls or feeding young) noted when available. We also paid attention to simultaneous records (i.e. two or more simultaneously singing males heard), particularly helpful in delimiting boundaries of ‘paper territories’. The plots are permanently marked with well-visible white stripes with black alphanumeric codes written on them and placed on tree trunks 1.8–2 m above the ground in grid nodes (grid 50 × 50 m), which facilitates the assessment of (heard or seen) birds locations and observers’ spatial orientation. Surveys started shortly after sunrise and lasted for 3–5 h, resulting in an average effort of 1.5–2 h per 10 ha of forest. We made additional surveys in between standard ones to approximately double the number of surveys performed (hereafter intensive approach). During these additional surveys, only Goldcrest and Firecrest were mapped. Mapping was done on either entire plots (K, 33.5 ha and NW, 25 ha) or on their monitored parts (C, 24 ha, W, 25.5 ha, see Fig. 1). On the NW plot birds were mapped only with the ‘standard’ approach.
Data processing and estimation of the number of territories
For both species and each of the three plots, all observations from field maps were redrawn to final single-sheet maps: one including results of nine surveys (standard approach) and another one with observations from all 19−21 surveys (intensive approach). At a minimum, three registrations in a cluster were required to delimit a territory in the absence of contemporary records (Wesołowski et al. 2022). After every single survey, starting from the 4th survey until the last one, separate estimates of the number of territories were produced for both species and both approaches. Therefore, six estimates for the standard approach (after 4th, 5th, 6th, 7th, 8th and 9th survey) and 16–17 estimates for the intensive approach (separate estimates after each between 4 and 19th–21st survey) were available. Each editor (JB, MCh, GN) prepared final maps for a single plot and estimates for both species under both (standard and intensive) approaches. All estimates were consulted among the editors to produce a consensus one, following criteria applied to delimit the number of territories (Tomiałojć 1980).
Point counts
We performed point counts with the distance sampling approach based on "point transects", in which the observer carried out short counts in the predefined points placed within the plots. The observer recorded any bird detected from the point, in distance bins. The observer was stationary and recorded distances to birds heard or seen around in three predefined distance bins (0–50, 51–100, and 101–150 m). On each plot, 10–14 points per plot (75 points in total) were surveyed five times during the breeding season, between 31 March and 3 June 2021, with intervals of around 15 days. Each survey consisted of 5-min-long counts repeated twice per survey and conducted immediately one after another to form data collected following the robust design protocol (5 surveys × 2 counts). In this study, only aural records were included—i.e. males detected by singing—to estimate abundance. We applied conservative criteria during counts and prepared data for modelling with only contemporary records—two or more males recorded at once—interpreted as referring to the observed number of males being larger than one to exclude the chances of multiple counting the same males (so-called false positive).
Abundance models
We estimated abundance from point counts using the hierarchical distance sampling (HDS) models (Chandler et al. 2011; Sillett et al. 2012; Kéry and Royle 2016). HDS model is a temporary emigration distance sampling model, and, apart from estimating abundance (λ) and detection probability (p), allows for inference about availability probability (ϕ) as well. Estimated detection functions g(x) describe the decline of detection with distance between the birds and the observer which allows us to estimate effective detection radius and area effectively covered with aural observations.
We fitted HDS models with three different detection functions: halfnormal, hazard and exponential (skipping the uniform one, which does not make any sense in these species, where detectability rapidly declines with distance). For Firecrest, abundance was treated as plot-dependent (a factor with seven levels) since we were interested in plot-specific estimates of density. This had to be simplified for Goldcrest due to the small number of observations: we used forest type factor with three levels (coniferous for two coniferous plots, a separate level for the K plot alone with a sufficient number of observations, and all the remaining plots pooled) in the abundance part. The detection probability was modelled as constant or observer-dependent (a factor with three levels, representing three observers). The availability probability was either set constant (no temporal variation over the season) or date-dependent, with the date being the days numbered since April 1. In total twelve HDS models were fitted per species. Model fitting was done with the gdistsamp() function using the R unmarked package (Fiske and Chandler 2011; Kellner et al. 2023) in R 4.1.2 (R Core Team 2022).
Fitted models were ranked according to AIC (Burnham and Anderson 2002) and model rankings produced with the modSel() function applied to a fitList() object in unmarked.
Since point counts with temporary emigration density are estimated as D = λ × ϕ / area, we used parametric bootstrapping of the top-supported models for each species (AIC weights of 0.77 and 0.74 for Goldcrest and Firecrest, respectively) to get distributions of densities computed as λ × ϕ. This procedure was repeated 100 times. To estimate abundance, densities per hectare were multiplied by plot areas. From these distributions of abundance, means and 95% quantile confidence intervals (CI) are presented.
We assessed the goodness of fit of the top supported models for each species by parametric bootstrap again with the parboot() function in unmarked to generate ‘perfect’ datasets under the models and using the chi-square discrepancy measure č (computed as the sum of (observed-expected)2/expected) for GoF assessment. Ĉ statistics were 1.35 (95% CI 0.77–1.99) and 1.28 (95% CI 0.97–1.50) for Goldcrest and Firecrest, respectively, indicating little overdispersion and an acceptable model fit.
Results
Territory mapping
For both species, there were 2–3 times more observations in the intensive territory mapping approach compared to the standard one (Table 1). The abundance obtained by the territory mapping method under the intensive approach was also around two times higher than under the standard one (Table 2). On the three studied plots (C, K and W) estimated abundance of Firecrest was from 5 to 6 per plot in the standard approach and from 9 to 14 per plot in the intensive approach. In turn, Goldcrest abundance varied from 1 to 2 per plot in the standard approach and from 1 to 4 per plot in the intensive approach. The abundance estimated after each single survey (starting from 4th one to the last one) increased with the cumulative number of surveys with no evident asymptotic value, particularly for Firecrest (Fig. 3), except for Goldcrest on plots W and C where adding subsequent visits did not change the estimate (Fig. 3C, D).

The increase of abundances estimate of Firecrest and Goldcrest with standard and intensive territory mapping method for the three plots in Białowieża National Park, Poland, spring 2021. Each symbol stands for an estimate produced after observations from consecutive surveys were redrawn onto species-and-plot-specific maps, starting from the 4th one—there were thus six estimates in the standard, and 16–17 in the intensive approach (see Methods). Blue triangles—plot K, green circles—plot C, brown diamonds–plot W (see Fig. 1). Top row—Firecrest, bottom row—Goldcrest. Left side (A, C)—standard approach, right side (B, D)—intensive approach. Trends shown with loess curves with polygons denoting 95% CI intervals
Point counts
Point counts yielded 154 observations of singing Firecrest males and 37 Goldcrest males. 133 Firecrests (86% of all) were recorded in the distance band closest to the observer (0–50 m), 21 (14%) in the second band (51–100 m) and none in the third band (101–150 m). For Goldcrest, respective numbers were 33 (89%), 4 (11%) and 0.
For both species, HDS models with observer-specific hazard detection functions were preferred (Table S1). Detection probability rapidly declined at a 40–50 m distance from the observer (Fig. 4), with a marked difference between GN and both remaining observers (JB and MCh). Effective radii were thus the shortest for GN (Firecrest, mean effective radius 42.9 m, 95% CI 34.0–53.6 m, Goldcrest, 47.6 m, CI 33.6–58.5 m) and much bigger for JB (Firecrest, effective radii: 65.0 m, CI 60.9–68.4 m, Goldcrest, 68.1 m, CI 54.2–74.8 m) and MCh (Firecrest, effective radii: 69.4 m, CI 65.8–73.8 m; Goldcrest, 64.0 m, CI 51.1–73.4 m). This translated to two-threefold differences among observers in areas effectively surveyed: for Firecrest, the areas were 0.59 ha (CI 0.36–0.90 ha), 1.33 ha (CI 1.17–1.50 ha) and 1.51 ha (1.36–1.71 ha) for GN, JB and MCh, respectively. The values for Goldcrest were 0.73 ha (CI 0.36–1.08 ha), 1.47 ha (CI 0.92–1.76 ha) and 1.30 ha (CI 0.82–1.69 ha) for the same three observers. Abundance submodels had plot and forest-type effects for Firecrest and Goldcrest, respectively. For the latter species, availability probability moderately and insignificantly declined with date from c 0.95 to c 0.50 (β = – 0.05, P = 0.16, Fig. 5, Table S2), while the top-supported model for Firecrest missed this effect in availability, which was constant across the season at about 0.95 (Table S2).

A Among-observer variation in detection functions from the top-supported models for Firecrest (left) and Goldcrest (right) for the three observers (top row). B Areas effectively surveyed (bottom row), indicated with observer-specific colours. A For detection functions, each thin line is a function computed from coefficients based on a single bootstrap replicate, bold lines show per-observer means. Effective radii are marked with small arrows on (A). B Areas effectively surveyed around observation points (black dot in the centre) by each observer are the colour-filled circles. A 100 m long radius (outer circle) is shown by horizontal lines inside the circles and the inner circle has a 50 m radius. The evident, worse precision for the Goldcrest is due to ~ 4 times less observations at the point counts (37 singing males observed vs 154 males of Firecrest)

The decline of availability probability with date under the top-supported HDS model for the Goldcrest. Thin lines show this relationship for individual 100 bootstrap resamples, and bold line is the mean. Polygon with dashed lines is the 95% confidence interval for the mean
On the three plots, where all three methods were applied, abundance estimated from point counts and HDS models was higher than numbers obtained by the standard territory mapping and similar to the estimates from the intensive approach (Table 2). This was evident for Firecrest and less clear for Goldcrest. Overall, the top-supported HDS models produced estimates highly correlated (r = 0.754, P = 0.016) with abundance from intensive territory mapping: the latter fell within 95% CI for the model-based estimates in five out of six cases (Fig. 6).

Relationship between abundance estimates from the intensive territory mapping (x axis) and point counts (y axis; points with error bars showing 95% CI for individual estimates) for Firecrest (dark green) and Goldcrest (light green). Bold line with polygon (95% CI) is a linear regression model, thin red line depicts a 1:1 relationship (perfect correlation)
Discussion
One of the main findings of this study is that abundance estimates from the territory mapping method depended on field effort—increasing the number (or density) of surveys twofold produces numbers that are about twice as high. This means that the numbers of both studied species are most likely underestimated with the standard approach of the territory mapping (8–10 surveys per season). At the same time, the intensive approach produced estimates which were consistent with results from point counts and HDS models.
The territory mapping method has been tested only for a few species (see Introduction) but for most others its accuracy remains unknown. Best (1975) indicates that results may also depend on many factors, like observation conditions or interpretational bias. The result may depend also on the number of visits (Svensson 1979), i.e. increasing the number of surveys may increase the number of detected individuals. Almost half a century ago, Tomiałojć (1980) tested the accuracy of a combined version of the mapping method by increasing the number of visits up to twenty. This test included twelve species, of which only four (Eurasian Treecreeper Certhia familiaris, Wren, Blackcap Sylvia atricapilla and Dunnock Prunella modularis) had the same abundance after increasing the number of controls. In the case of Goldcrest, the abundance after twenty visits increased by 25% compared to the standard ten, whereas for Firecrest the abundance increased from zero to one pair.
In our study, we increased the number of visits from nine to about twenty. Whether this—or a smaller—number of surveys is sufficient to get the actual numbers of Regulus species remains unclear. However, out of six species-plot comparisons under intensive mapping, in just two—for Goldcrest numbers on plots W and C—the estimates of the number of territories (one and three, respectively) changed just once and for most of the season remained the same, which means that adding more surveys did not result in increasing abundance estimates (Fig. 3D). In contrast, estimates for Firecrest—a much more numerous species—for all three plots tended to increase more or less continuously with intensive mapping (Fig. 3B). Nevertheless, knowledge of the actual number of territories can be unavailable for highly elusive birds, such as the species studied here, for which finding nests is rare (Thompson 2004). This indicates that the actual abundance of these species may be even higher than the numbers from the intensive territory mapping and/or point counts and HDS models.
The main reason for the underestimation is that songs of these species are perceived as quiet by the human ear. Their song amplitude is around 80.4 dB (Winiarska et al. 2023), which is not much lower than, e.g. European Robin Erithacus rubecula or Wood Warbler (both around 82 dB), but Regulus species sing at a high (5–8, mean ~ 7 kHz) frequency (Winiarska et al. 2023). Humans do not perceive sound intensity level linearly: a sound with the same intensity, but higher frequency, is heard as quieter. Another issue is the faster attenuation of high-frequency sounds, i.e. they can be heard from a shorter distance not because they are quiet, but because of their high frequency. Moreover, these species sing rarely (author’s pers. obs.) which leads to few detections (most of the time they stay silent and are simply missed in the field) and even fewer observations of simultaneously singing males over the whole season. All this results in underestimations with the conservative criteria of the territory mapping.
The way to improve estimates from the territory mapping method (i.e. allow the possibility that numbers are higher than set with conservative criteria) for Regulus, and perhaps other species with similar singing characteristics would be to relax the criteria used to draw and enumerate territories. When staying conservative, as applied in this work, and in all the previous papers based on territory mapping (Wesołowski et al. 2006, 2010, 2015, 2022) at least three records are required to define a ‘cluster’ and resulting ‘paper territory’, if there are no simultaneous records, which naturally separate neighbouring territories. For example, if few clusters are delimited, and there exist single records, that are apparently too distant to refer to any of the territories already delimited (bearing in mind that ‘too distant’ can still be a matter of debate, see below), this is ignored. Still, however, these might represent neighbouring territories—provided that a species sings rarely and quietly it is perfectly possible, that only one or two detections (or even none) in an existing territory emerge over the season just by chance. With these criteria relaxed—i.e. fewer records required to delimit the territory—the estimated number of territories increased as shown by Gottschalk and Huettmann (2011). Another problematic issue is the ‘distance’ criterion: how large a distance must be to treat a record as referring to another territory. This is a partly subjective and probably unavoidable decision, even if based on biological grounds. One never knows in advance what are the true numbers and density, but this affects average distances between records of birds from neighbouring territories. It appears, therefore, that both the assumptions used to delimit clusters and the distance criterion, might both be subjective and too conservative. One possible solution to this would be to produce a min–max range estimate of the abundance from territory mapping, where the lower bound would represent the ‘well-founded’ territories drawn according to the conservative criteria (the minimal number) and the upper would include next, probable territories delimited with relaxed criteria (e.g. one or two instead of three detections, distant from evident clusters).
The “true abundance” issue of an avian population stems from the dynamic nature and openness of (almost) any wild population. Abundance changes within a season (Gnielka 1992) and thus a single, ‘per-season’ number given as ‘abundance’ is in most cases an obvious simplification. In fact, even populations of strictly sedentary and territorial species are open to some degree, due to deaths, emigrations and immigrations (Paul and Roth 1983). Therefore, one should define what abundance is, in a population of interest and the context of the study, and perhaps also restrict the time window to which the estimate refers (see e.g. Neubauer et al. 2022 for the Marsh Tit Poecile palustris example). Both Regulus species have a long breeding period and probably frequently raise second broods during spring (Haftorn 1978). Also, second broods with a pair bond already established may result in reduced singing activity during the season. Given that birds can leave the territory occupied to raise their first brood and establish new ones, errors in ‘abundance’ defined this way can arise by any estimation method.
A possible solution to cope with the underestimation problem is increasing the number of visits during territory mapping, but this is extremely time consuming. We estimated the time and effort needed to complete counts in the field under the three methods. The point counts consumed ~ 55 h, whereas the territory mapping method took ~ 172 and ~ 296 h in standard and intensive approaches, respectively. An alternative way is to use novel methods to estimate abundance, like point counts and the relevant models (as applied here), which is a time-effective option. We found that the detectability strikingly declining with the distance from the observer is of significance for both Regulus species, which is consistent with Gottschalk and Huettmann’s (2011) findings (they showed that the effective detection radius of Goldcrest was 34 m, while that of Firecrest was 29 m). In our study, we found significant differences in the detection functions between observers (GN vs JB and MCh). Therefore, typical assumptions of a fixed radius to convert abundances derived from point counts into a density estimate are likely violated by observer-specific detection distances, which can be accounted for when using distance sampling. For this reason, it seems to be a wise solution to use the distance sampling methodology in this case. Other approaches (except for the spatial capture-recapture methods, Borchers 2012) assume some unknown radius survey around the observer, which means that the area effectively surveyed is also unknown.
Concluding remarks
To sum up, for elusive species abundances estimated with the standard territory mapping might not give absolute numbers as it was long believed, but underestimations, which is due to both the secretive behaviour of the species and conservative criteria of the method. Hence, they represent indices of abundance rather, than being close to true numbers. There are easier ways to get indices than effort- and time-consuming territory mapping method. Field tests are still missing for most species, while the general rules and assumptions of the method do not always hold. More reliable results require increasing the number of visits, or applying novel methods as much less time is needed to complete point counts than to perform intensive territory mapping. The modern methods and modelling as we used here provide estimates along with their uncertainty (unlike territory mapping which produces a single number) what can make more sense, given that avian populations are (mostly) open and abundance can change over the season. If one really wishes to do territory mapping, relaxing conservative criteria can be a good solution to cope with underestimations.
Data availability
Data are available on request from the corresponding author.
References
Best LB (1975) Interpretational errors in the “mapping method” as a census technique. Auk 92:452–460. https://doi.org/10.2307/4084601
Bibby CJ, Burgess ND, Hill DA (2000) Bird census techniques. Academic Press, London
Borchers DL (2012) A non-technical overview of spatially explicit capture-recapture models. J Ornithol 152:435–444
Bötsch Y, Jenni L, Kéry M (2020) Field evaluation of abundance estimates under binomial and multinomial N-mixture models. Ibis 162:902–910. https://doi.org/10.1111/ibi.12802
Buckland ST, Rexstad EA, Marques TA, Oedekoven CS (2015) Distance sampling: methods and applications. Springer. https://doi.org/10.1007/978-3-319-19219-2
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer-Verlag, New York
Chandler RB, Royle JA, King DI (2011) Inference about density and temporary emigration in unmarked populations. Ecology 92:1429–1435. https://doi.org/10.1890/10-2433.1
Chodkiewicz T, Chylarecki P, Sikora A, Wardecki Ł, Bobrek R, Neubauer G, Marchowski D, Dmoch A, Kuczyński L (2019) Raport z wdrażania art. 12 Dyrektywy Ptasiej w Polsce w latach 2013–2018: stan, zmiany, zagrożenia. Biuletyn Monitoringu Przyrody 20:1–80
Cramp S (1998) Cramp’s the complete birds of the Western Palearctic. Oxford Univ Press, Oxford
Dyderski MK, Paź S, Frelich LE, Jagodziński AM (2018) How much does climate change threaten European forest tree species distributions? Global Change Biol 24:1150–1163. https://doi.org/10.1111/gcb.13925
Faliński JB (1986) Vegetation dynamics in temperate lowland primeval forests. Springer, Netherlands. https://doi.org/10.1007/978-94-009-4806-8
Fiske I, Chandler R (2011) unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Soft 43:1-23. https://doi.org/10.18637/jss.v043.i10
Flade M (1994) Die Brutvogelgemeinschaften Mittel- und Norddeutschlands. Grundlagen für den Gebrauch vogelkundlicher Daten in der Landschaftsplanung. IHW-Verlag, Eching.
Gnielka R (1990) Anleitung Zur Brutvogelkartierung. Apus 7:145–239
Gnielka R (1992) Möglichkeiten und Grenzen der Revierkartierungsmethode. Vogelwelt 113:231–240
Gottschalk TK, Huettmann F (2011) Comparison of distance sampling and territory mapping methods for birds in four different habitats. J Ornithol 152:421–429. https://doi.org/10.1007/s10336-010-0601-1
Gregory RD, Gibbsons DW, Donald PF (2004) Bird census and survey techniques. In: Sutherland WJ, Newton I, Green R (eds) Bird ecology and conservation: a handbook of techniques. Oxford Univ. Press, New York, pp 17–54
Haftorn S (1978) Cooperation between the male and female Goldcrest Regulus regulus when rearing overlapping double broods. Ornis Scand 9:124. https://doi.org/10.2307/3675873
Jaroszewicz B, Cholewińska O, Gutowski JM, Samojlik T, Zimny M, Latałowa M (2019) Białowieża Forest—a relic of the high naturalness of european forests. Forests 10:849. https://doi.org/10.3390/f10100849
Joseph LN, Elkin C, Martin TG, Possingham HP (2009) Modeling abundance using N-mixture models: the importance of considering ecological mechanisms. Ecol Appl 19:631–642. https://doi.org/10.1890/07-2107.1
Kellner KF, Smith AD, Royle JA, Kéry M, Belant JL, Chandler RB (2023) The unmarked R package: twelve years of advances in occurrence and abundance modelling in ecology. Methods Ecol Evol. https://doi.org/10.1111/2041-210X.14123
Kéry M, Royle JA (2016) Applied hierarchical modeling in ecology. In: Analysis of distribution, abundance and species richness in R and BUGS. Vol. 1. Prelude and static models. Academic Press, London
Kéry M, Schaub M (2012) Bayesian population analysis using WinBUGS: a hierarchical perspective. Academic Press, London
Link WA, Schofield MR, Barker RJ, Sauer JR (2018) On the robustness of N-mixture models. Ecology 99:1547–1551. https://doi.org/10.1002/ecy.2362
Lõhmus A (2022) Absolute densities of breeding birds in Estonian forests: a synthesis. Acta Ornithol 57:29–47. https://doi.org/10.3161/00016454AO2022.57.1.003
Neubauer G, Sikora A (2020) Abundance estimation from point counts when replication is spatially intensive but temporally limited: comparing binomial N-mixture and hierarchical distance sampling models. Ornis Fenn 97:131–148
Neubauer G, Wolska A, Rowiński P, Wesołowski T (2022) N -mixture models estimate abundance reliably: A field test on Marsh Tit using time-for-space substitution. Ornithol Appl 124:duab054. https://doi.org/10.1093/ornithapp/duab054
Niethammer G, Bauer KM, Glutz von Blotzheim UN, Bezzel E (1991) Handbuch der Vögel Mitteleuropas: Passeriformes. Aula Verlag, Wiesbaden, Sylviidae
Paul JT, Roth RR (1983) Accuracy of a version of the spot-mapping census method. J Field Ornithol 54:42–49
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF (1993) Handbook of field methods for monitoring landbirds. Gen Tech Re PSW GTR-144. Albany, CA: Pacific Southwest Research Station, Forest Service, US Department of Agriculture. https://doi.org/10.2737/PSW-GTR-144
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org
Royle JA (2004) N–mixture models for estimating population size from spatially replicated counts. Biometrics 60:108–115. https://doi.org/10.1111/j.0006-341X.2004.00142.x
Samojlik T, Rotherham ID, Jędrzejewska B (2013) The cultural landscape of royal hunting gardens from the fifteenth to the eighteenth century in Białowieża Primeval Forest. In: Roterham ID (ed) Cultural severance and the environment. Springer, pp 191–204
Sillett TS, Chandler RB, Royle JA, Kéry M, Morrison SA (2012) Hierarchical distance-sampling models to estimate population size and habitat-specific abundance of an island endemic. Ecol Appl 22:1997–2006. https://doi.org/10.1890/11-1400.1
Snow DW, Perrins CM (1998) The birds of the Western Palearctic. Oxford University Press
Svensson SE (1979) Census efficiency and number of visits to a study plot when estimating bird densities by the territory mapping method. J Appl Ecol 16:61. https://doi.org/10.2307/2402728
Thompson WL (2004) Sampling rare or elusive species: concepts, designs, and techniques for estimating population parameters. Island Press
Tomiałojć L (1980) The combined version of the mapping method. In: Oelke H (ed) Bird census work and nature conservation. Dachverband Deutscher Avifaunisten, Göttingen, pp 92–106
Tomiałojć L (2004) Accuracy of the mapping technique for a dense breeding population of the Hawfinch Coccothraustes coccothraustes in a deciduous forest. Acta Ornithol 39:67–74
Tomiałojć L, Lontkowski J (1989) A technique for censusing territorial Song Thrushes Turdus philomelos. Ann Zool Fenn 26:235–243
Tomiałojć L, Walankiewicz W, Wesołowski T (1977) Methods and preliminary results of the bird census work in primeval forest of Białowieża National Park. Pol Ecol Stud 3:215–223
Tomiałojć L, Wesołowski T, Walankiewicz W (1984) Breeding bird community of a primaeval temperate forest (Bialowieza National Park, Poland). Acta Ornithol 20:241–310
Treml V, Mašek J, Tumajer J, Rydval M, Čada V, Ledvinka O, Svoboda M (2022) Trends in climatically driven extreme growth reductions of Picea abies and Pinus sylvestris in Central Europe. Global Change Biol 28:557–570. https://doi.org/10.1111/gcb.15922
Walankiewicz W, Czeszczewik D, Mitrus C, Szymura A (1997) How the territory mapping technique reflects yearly fluctuations in the Collared Flycatcher Ficedula albicollis numbers. Acta Ornithol 32:201–207
Wesołowski T, Rowiński P, Mitrus C, Czeszczewik D (2006) Breeding bird community of a primeval temperate forest (Białowieża National Park, Poland) at the beginning of the 21st century. Acta Ornithol 41:55–70
Wesołowski T, Mitrus C, Czeszczewik D, Rowiński P (2010) Breeding bird dynamics in a primeval temperate forest over 35 years: variation and stability in a changing world. Acta Ornithol 45:209–232
Wesołowski T, Czeszczewik D, Hebda G, Maziarz M, Mitrus C, Rowiński P (2015) 40 years of breeding bird community dynamics in a primeval temperate forest (Białowieża National Park, Poland). Acta Ornithol 50:95–120
Wesołowski T, Czeszczewik D, Hebda G, Maziarz M, Mitrus C, Rowiński P, Neubauer G (2022) Long-term changes in breeding bird community of a primeval temperate forest: 45 years of censuses in the Białowieża National Park (Poland). Acta Ornithol 57:71–100. https://doi.org/10.3161/00016454AO2022.57.1.005
Winiarska DM, Szymański P, Osiejuk TS (2023) Detection ranges of forest bird vocalisations: guidelines for passive acoustic monitoring. https://doi.org/10.21203/rs.3.rs-2996497/v1
Acknowledgements
We are very thankful to Tomasz Wesołowski, Cezary Mitrus, Dorota Czeszczewik and Fabian Przepióra who participated in territory mapping surveys. We thank Daniel O’Connell for improving our English. We are grateful to Thomas Gottschalk and two anonymous reviewers for their helpful comments on the earlier version of this paper.
Funding
This study was supported by the University of Wrocław.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study design, data collection in the field and material preparation. Data analysis was performed by Grzegorz Neubauer and Julia Barczyk. Julia Barczyk wrote the first draft of the manuscript and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
No conflict of interest.
Ethical approval
The research was conducted in compliance with the permissions from the Białowieża National Park.
Additional information
Communicated by T. Gottschalk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Barczyk, J., Cholewa, M. & Neubauer, G. Abundance estimation from point counts and territory mapping: comparing different approaches for two Regulus species. J Ornithol 165, 793–804 (2024). https://doi.org/10.1007/s10336-024-02151-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-024-02151-6