
Abstract
Weak oscillating magnetic fields (OMF) in the radiofrequency range are known to disrupt the orientation of birds. However, until now, it has not been experimentally verified that the sensitivity to OMF is a characteristic feature specifically of the magnetic compass and OMF does not influence the celestial compass system as well. Here we studied if OMF affected the star compass of a long-distance migrant, the Garden Warbler. The birds were tested under the natural starry sky under two different conditions: in the natural magnetic field (NMF) and in radiofrequency OMF with the amplitude 20nT and frequency 1.41 MHz (matching the Larmor frequency of a freestanding electron spin in the local NMF of 50,400 nT). This amplitude is about ten times higher than the sensitivity threshold to OMF shown for this species in previous studies. Our experimental results clearly demonstrated that OMF did not influence the celestial (star) compass system: with access to the starry sky garden warblers showed migratory orientation appropriate for autumn migratory season both in the NMF and in the OMF. Thus, the OMF effect is pertinent to the magnetic compass system, not to the avian orientation in general.
Zusammenfassung
Ein oszillierendes Magnetfeld stört die Orientierung eines Zugvogels in Gegenwart von Gestirnen nicht.
Es ist bekannt, dass schwache oszillierende Magnetfelder (OMF) im Hochfrequenzbereich die Orientierung von Vögeln stören können. Bisher konnte jedoch nicht experimentell nachgewiesen werden, dass die Empfindlichkeit gegenüber OMF ein spezifisches Merkmal des magnetischen Kompasses ist und OMF nicht auch das Himmelskompasssystem beeinflussen. Wir untersuchten, ob OMF den Sternenkompass eines Langstreckenziehers, der Gartengrasmücke, beeinflusst. Die Vögel wurden unter dem natürlichen Sternenhimmel unter zwei verschiedenen Bedingungen getestet: im natürlichen Magnetfeld (NMF) und im hochfrequenten OMF mit einer Amplitude von 20nT und einer Frequenz von 1,41 MHz (was der Larmor-Frequenz eines freistehenden Elektronenspins im lokalen NMF von 50.400 nT entspricht). Diese Amplitude ist etwa zehnmal höher als die Empfindlichkeitsschwelle für OMF, die in früheren Studien für diese Vogelart ermittelt wurde. Unsere experimentellen Ergebnisse zeigen eindeutig, dass OMF das himmlische (Sternen-)Kompasssystem nicht beeinflussen: Bei Zugang zum Sternenhimmel zeigten Gartengrasmücken sowohl im NMF als auch im OMF eine der Herbstzugzeit entsprechende Zugorientierung. Der OMF-Effekt spielt also eine Rolle im magnetische Kompasssystem, nicht aber für die Orientierung der Vögel im Allgemeinen.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Migratory birds may use different orientation cues during their seasonal migrations such as the stars (Sauer 1957; Emlen 1967, 1970), the sun’s position and patterns of skylight polarization (Kramer 1957; Able 1982; Schmidt-Koenig 1990) and the geomagnetic field (Wiltschko and Wiltschko 1972, 2019). Both the celestial compass system (stars) and the magnetic compass system are important for many passerine species migrating at night (Wiltschko and Wiltschko 2015; Mouritsen 2018). Different species of avian migrants may rely on different compass systems to a varying degree, and their integration rules and hierarchy vary (Chernetsov 2015; Pakhomov and Chernetsov 2020; Pakhomov et al. 2022). The information obtained from these compass systems seems to be redundant, but in some circumstances may be very useful, for example, during cloudy weather, when nocturnal migrants can rely only on their magnetic compass.
Weak oscillating magnetic fields (OMF) in the radiofrequency range (from 0.4 to 85 MHz as reported so far) are known to disrupt the orientation of birds (Ritz et al. 2004, 2009; Thalau et al. 2005; Engels et al. 2014; Schwarze et al 2016; Kavokin et al. 2014; Pakhomov et al. 2017; Kobylkov et al. 2019; Leberecht et al. 2022, 2023). These experiments showed that birds could not choose the correct direction either in monochromatic, single-frequency OMF (Ritz et al. 2004, 2009; Thalau et al. 2005; Pakhomov et al. 2017) or when subjected to broadband magnetic noise (Ritz et al. 2004; Engels et al. 2014; Schwarze et al. 2016; Leberecht et al. 2022). All these experiments on the effect of OMF on the ability of birds to choose the correct direction during their seasonal migrations were carried out without access to astronomic orientation cues, therefore the OMF effect is considered to influence only the magnetic compass system.
Though the OMF effect has been experimentally demonstrated by several research groups, its biophysical origin as well as its possible role in migration ecology remain not quite clear. Indeed, it finds a qualitative explanation in the model of the light-dependent magnetic compass of birds, based on cryptochrome molecules in the eyes (Ritz et al. 2000), due to the electron paramagnetic resonance in photoexcited radical-pairs (Timmel and Hore 1996; Ritz et al. 2004; Hore and Mouritsen 2016). However, the predictions of this model are in considerable quantitative disagreement with the experimental results. In the experiments, birds were disoriented by OMFs with the amplitudes of 2–3 orders of magnitude lower than theoretically estimated (Kavokin 2009, 2017; Hiscock et al. 2017). On top of that, OMFs applied locally to the eyes of Garden Warblers did not disorient them, while the OMF of similar amplitude applied to the whole body of the bird caused clear disorientation (Bojarinova et al. 2020). Kirschvink (2014) suggested the existence of a separate receptor, specifically sensitive to OMFs, that blocks the sensory pathway of the magnetic compass system to avoid navigational errors during solar storms. Another possible role of such a receptor would be warning of atmospheric perturbations (tropical thunderstorms) on the migration route (Granger et al. 2022). No plausible biophysical mechanism of this hypothetic OMF receptor has been proposed so far.
In view of the difficulties in understanding the OMF effect, outlined above, the question arises whether or not this effect is pertinent to the magnetic compass system only. Indeed, if, for example, the OMF is perceived by the bird as a warning of a close thunderstorm, it might block the bird’s program of directional migration flight to prevent it from flying in adverse atmospheric conditions. In that case, one would expect that in experimental conditions the OMF would disrupt not only magnetic orientation but also the orientation by celestial objects like stars. However, until now, it has not been experimentally verified that the sensitivity to OMF is a characteristic feature specifically of the magnetic compass and OMF does not influence the celestial compass system as well.
In this work, we study whether OMF affected the star compass of a long-distance migrant, the Garden Warbler (Sylvia borin). The effect of single-frequency OMF on the magnetic compass of this species was studied in detail (Kavokin et al. 2014; Pakhomov et al. 2017). Garden Warblers were not able to show the seasonally appropriate orientation if subjected to the 1.4 MHz OMF (the Larmor frequency of the free electron spin in the geomagnetic field at the experimental location in Rybachy, Eastern Baltic) as weak as 1 nT (Pakhomov et al. 2017). The sensitivity threshold for this species, obtained from the value of the average length of the orientation vector, is somewhat higher, between 2 and 3 nT.
Materials and methods
Study site and bird keeping
We performed orientation tests in autumn 2022 with first-year Garden Warblers captured during their autumn migration on the Courish Spit (Kaliningrad region, Russia; 55°09′ N, 20°52′ E). Experimental birds (n = 23) were captured by mist nets on August 11–August 18 and were kept in an outdoor aviary in individual cages. They experienced the natural photoperiod, the natural geomagnetic field and had access to astronomic orientation cues (the sun, sunset polarization patterns and stars) during their time in captivity. They were provided with food (mealworms Tenebrio molitor, a homemade mix of mashed boiled eggs with grated carrots, Padovan complete feed for insect-eating birds, berries of the Black Elderberry (Sambucus nigra)) and water ad libitum. The aviary was equipped with online infrared video cameras, and only those birds that showed migratory restlessness on a given night were selected for experiments.
Experimental conditions and setup
The experiments were performed outdoors on wooden tables placed in the clearing of reeds on the coast of the Courish Lagoon. Migratory orientation of Garden Warblers was studied in plastic Emlen funnels (Emlen and Emlen 1966) (top diameter 350 mm, bottom diameter 100 mm, slope 45°).
The birds were tested under the natural starry sky in two different conditions: in the natural magnetic field (NMF) and in radiofrequency OMF with the amplitude 20 nT and frequency 1.41 MHz (matching the Larmor frequency of a freestanding electrons spin in the local NMF of 50,400 nT). This amplitude is about ten times higher than the sensitivity threshold to OMF shown for this species in the previous studies (Kavokin et al. 2014; Pakhomov et al. 2017).
For this experiment, we used the equipment employed in our previous studies of the orientation of birds, including the Garden Warblers, in the OMF (Kavokin et al. 2014; Pakhomov et al. 2017; Bojarinova et al. 2023). To apply OMF, we used 16 two-loop coils 75 cm in diameter, fed by radiofrequency current from 16 custom made broadband amplifiers. There were 4 coils on each table, each fed from its own amplifier. The amplifiers were mounted in aluminium boxes (4 in each box), placed under each of the 4 experimental tables and received 1.41 MHz sinusoidal input signals through coaxial cables from 4 output channels of 2 function generators (Rigol DG4162) placed in the cabin 15 m apart along the straight line from the center of a clearing in reeds, where four wooden tables were built. This ensured that the acoustic noise from the generators was not discernible over the background of natural noises. This assembly could provide the OMF with an amplitude of up to 200 nT at the center of each coil at the frequency of 1.41 MHz. One side of the coil was raised above the table top to increase the angle between OMF and NMF, which as a result was approximately 30°. Emlen funnels were placed on tables, one in the center of each coil. The parameters of the OMF were controlled before and after each test. To this end, we used a 25 cm loop antenna, connected to a digital storage oscilloscope Tektronix TPS 2012B (Tektronix Inc., USA).
To measure detailed spectra of the applied OMF and of the background electromagnetic noise, we used Rigol DSA 815 spectrum analyzer (9 kHz–1.5 GHz) with one of 3 different antennas. OMF and magnetic noise spectra were measured using a Schwarzbeck Mess-Electronic FMZB 1512 active loop antenna (in the range 350 kHz–20 MHz) and a custom-made broadband active loop antenna (5 MHz–150 MHz). The electric component of the electromagnetic noise was measured using a custom-made passive 0.5 m dipole antenna. The spectra taken with the spectrum analyzer bandwidth of 100 kHz are shown in Fig. 1. The broadband magnetic noise did not exceed 10–2 pT/Hz1/2, while the noise of the vertical component of the electric field did not exceed 10–4 mV/(m Hz1/2). Series of sharp peaks in the ranges of 10–20 MHz and 80–120 MHz are due to radio broadcasting stations. The radio transmission signals have predominant vertical polarization of the electric component and horizontal polarization of the magnetic component; the spectra of these components are plotted in Fig. 1. The background broadband noise is not polarized. Switching on the OMF produced no detectable effect on the electromagnetic noise spectra above 10 MHz; spectra with the operating OMF equipment for this frequency range are not shown to avoid duplication. The magnetic noise spectrum up to 10 MHz with the OMF equipment switched on is shown by the dashed green line. The visible width of the 1.41 MHz OMF peak is determined by the spectrum analyzer bandwidth; no sidebands are discernible over the noise. The OMF spectrum with higher frequency resolution (spectrum analyser bandwidth 100 Hz) is shown in the inset to Fig. 1. The estimated overall amplitude of the ambient magnetic noise, including radio transmission signals, comprises about 10–3 of the amplitude of the applied OMF.

Spectra of horizontal magnetic (upper curve, dark blue) and vertical electric (lower curve, red) ambient fields at the experimental site. The dashed green line shows the spectrum of the OMF in the centre of one of the coils. All the spectra are taken in the “average” mode with the spectrum analyzer bandwidth 100 kHz. Inset: spectra of the OMF (black) and of background magnetic noise (grey) around 1.41 MHz with 100 Hz resolution (color figure online)
Orientation tests and data analysis
Orientation tests were carried out in 2022 within a period of 13 days (August 25–September 6). The tests started not earlier than the beginning of astronomical twilight on clear starry nights (usually ≥ 90%). Experimental birds were transferred from their cages to the Emlen funnels in individual opaque textile bags. The directionality of the birds’ activity was recorded as scratches left by their claws as they hopped in the funnels on a polyethylene film covered with a dried mixture of whiting and glue. Each test lasted for 45–50 min. After 2 nights of tests, the birds had a rest for 1–3 days. Each bird was tested in every trial at least twice (except two birds that were tested in one of the trials only once) (Table 1). From three to four researchers (JB, DS, AF, A. Prokshina, and AP) independently determined each bird’s mean direction from the distribution of scratches. The mean of the directions determined by the observers was recorded as the orientation result. In most cases, the mean direction could be very precisely identified using the simple visual estimation method (Mouritsen and Larsen 1998). If at least two observers considered the scratches to be randomly distributed, the bird was considered to be not oriented in the given test. If a pattern of scratches was not clear, scratches in each of 36 × 10° sectors were counted. Then we used circular statistics software to assess the directionality based on the numbers of scratches. If the number of scratches was less than 40, the bird was considered to be inactive in a given test. To minimize observer bias, blinded method was used when the data were analysed. The final decision was made by AP who was not aware of which experimental condition was realised in each specific test.
The total number of tests in a trial was 55 (NMF) and 54 (OMF). The number of tests evaluated as non-oriented was the same for both trials (20%, Table1); birds that showed no activity were exceptional (two tests in NMF and one test in OMF). We included the results of all birds that were tested at least one time in experimental conditions and showed at least one sufficiently active result (for details, see Table 1). For group direction analysis, to avoid pseudoreplication, we used the mean direction of each bird obtained during all days.
Statistical tests were performed with ORIANA (Kovach Computing Services, v. 4.0). The differences in mean orientation direction between experimental groups were tested by the nonparametric Mardia-Watson-Wheeler (MWW) test.
Results
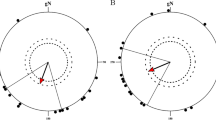
Garden Warblers tested in the NMF under the starry sky showed the appropriate migratory direction (α = 232°, n = 23, r = 0.70, P < 0.001, 95% CI = 212–252°; Fig. 2a). The mean direction of birds was similar to the mean autumn migratory direction of the same species, according to recoveries of birds ringed on the Courish Spit (α = 213°, n = 14, r = 0.96, p < < 0.001, 95% CI = 205–222°; Bolshakov et al. 2001; and unpublished data of the Biological Station Rybachy). A similar mean direction was obtained under the same conditions in previous experiments in Garden Warblers tested in Emlen funnels (α = 218°, n = 38, r = 0.57, p < 0.01, 95% CI = 194–242°; Pakhomov and Chernetsov 2014).

Orientation of Garden Warblers under clear starry sky: a in the natural geomagnetic field (NMF); b in 20 nT OMF. Dots show mean directions of individual birds in each experimental condition. The arrows show the second-order mean of the group of birds in each condition. The inner and outer dashed circles indicate the 5% and 1% significance levels of the Rayleigh test, respectively. The radial lines indicate the 95% CI
Then the same birds were tested under the clear starry sky but in radiofrequency OMF with the amplitude 20nT and frequency 1.41 MHz. In this trial, Garden Warblers also showed the appropriate migratory direction (α = 210°, n = 22, r = 0.69, P < 0.001, 95% CI = 189–230°; Fig. 2b). The two distributions (both under the starry sky, but in the NMF and OMF conditions) did not differ according to the MWW test: W = 0.58, p = 0.75.
The directional preferences of most birds did not change between the two trials. The angular within-individual differences between the two conditions (OMF under the starry sky and NMF under the starry sky) were unimodally distributed and its mean direction (α = 347°, r = 0.41, p = 0.02; Fig. 3) was not significantly different from 0° (95% CI = 334–52°).

Distribution of the angular difference in the headings of individuals between in the 20 nT OMF and the NMF. Dots show the directions shown by individual birds in the 20 nT OMF in respect to the direction shown by the same bird in the NMF. The angle of 0° would mean that both directions are identical. Arrows show the second-order mean of the sample. The inner and outer dashed circles indicate the 5% and 1% significance levels of the Rayleigh test, respectively; radial lines indicate 95% CI
Discussion
Our experimental results clearly demonstrated that OMF did not influence the celestial (star) compass system. When Garden Warblers with access to starry sky were subjected to OMF with an amplitude about ten times higher than the sensitivity threshold to OMF shown for this species earlier (Kavokin et al. 2014; Pakhomov et al. 2017), they nevertheless showed appropriate seasonal migratory orientation. Since the present study was performed at exactly the same location, by the same team of researchers and under the same conditions (including the OMF frequency spectrum, equipment, time of the year and of the day), this conclusion indeed seems to be robust.
OMFs as well as broadband radio frequency electromagnetic noise of anthropogenic origin have been shown to disrupt magnetic orientation behaviour in different vertebrates including birds (Malkemper et al 2015; Landler et al. 2015; Ritz et al. 2004, 2009; Thalau et al. 2005; Engels et al. 2014; Schwarze et al 2016; Pakhomov et al. 2017; Kobylkov et al. 2019; Leberecht et al. 2022, 2023; Bojarinova et al. 2023). That is why experiments on the magnetic compass in birds are encumbered near cities and/or in the neighborhood of universities where irregular and uncontrollable anthropogenic time-dependent electromagnetic fields are omnipresent (Engels et al. 2014). Natural radio frequency electromagnetic noise might also have the potential to disturb magnetic orientation behavior, specifically solar RF and atmospheric RF electromagnetic noise (Bianco et al. 2019; Granger et al. 2022). Some researchers hold on the idea that the sensitivity to OMF is related to the mechanism of magnetoreception itself (Ritz et al. 2000, but see Kavokin 2009), others suggest that there may be an additional sensory system aimed at the detection of the environmental magnetic noise (Kirschvink 2014; Bojarinova et al. 2023). We would like to stress that our experimental results do not allow us to make a choice among different models of the OMF effect. Our experiments just show that the single-frequency OMF effect, whatever its origin is—either direct suppression of the magnetic compass functionality, or reaction of an “alarm” system to potentially adverse magnetic conditions – is pertinent to the magnetic orientation system, not to the avian navigation system in general. At least the star compass is not affected by the single-frequency OMF.
This fact suggests that the coexistence of both compass systems (stars and magnetic compasses) is crucial for nocturnal migrants and not as redundant as it might seem at first glance. The magnetic compass is indeed very useful during cloudy weather. Birds can rely on it and do not stop their migration when they have no celestial cues such as stars. However, in cases when the magnetic compass could fail, such as during geomagnetic perturbations or while crossing magnetic anomalies, the star compass becomes more important and helps birds to maintain the direction of their migratory flight.
Having in mind the discussion on the relative efficiency of single-frequency and broadband OMFs in suppressing the magnetic orientation of birds (Engels et al. 2014; Hiscock et al. 2017; Pakhomov et al. 2017; Schwarze et al. 2016; Leberecht et al. 2022, 2023), it would make sense to stage a similar study using a broadband OMF (preferably with the spectrum similar to that of environmental magnetic noise, either natural or induced by human activity). However, it is unlikely that the effects of single-frequency and broadband OMFs are mediated by different sensory mechanisms. Our model species, the Garden Warbler, repeatedly showed disruption of the magnetic compass under weak single-frequency OMFs, while demonstrating no reaction to the much stronger OMF when tested under the starry sky. We consider this a strong argument in favour of the hypothesis that all the OMF effects are directly related to the magnetic compass system of birds.
Data availability
The data analyzed in this study are available in Table 1.
References
Able KP (1982) Skylight polarization patterns at dusk influence migratory orientation in birds. Nature 299:550–551. https://doi.org/10.1038/299550a0
Bianco G, Ilieva M, Åkesson S (2019) Magnetic storms disrupt nocturnal migratory activity in songbirds. Biol Lett 15:20180918. https://doi.org/10.1098/rsbl.2018.0918
Bojarinova J, Kavokin K, Pakhomov A, Cherbunin R, Anashina A, Erokhina M, Ershova M, Chernetsov N (2020) Magnetic compass of Garden Warblers is not affected by an oscillating magnetic field applied to their eyes. Sci Rep 10:3473. https://doi.org/10.1038/s41598-020-60383-x
Bojarinova J, Kavokin K, Cherbunin R, Sannikov D, Fedorishcheva A, Pakhomov A, Chernetsov N (2023) Sensitivity threshold of avian magnetic compass to oscillating magnetic field is species-specific. Behav Ecol Sociobiol 77:6. https://doi.org/10.1007/s00265-022-03282-7
Bolshakov CV, Shapoval A, Zelenova N (2001) Results of bird ringing by Biological Station «Rybachy» on the Courish Spit: long-distance recoveries of birds ringed in 1956–1997. Avian Ecol Behav Suppl 1:1–126
Chernetsov N (2015) Avian compass systems: do all migratory species possess all three? J Avian Biol 46:342–343. https://doi.org/10.1111/jav.00593
Emlen ST (1967) Migratory orientation in the Indigo bunting, Passerina cyanea. Part I: evidence for use of celestial cues. Auk 84:309–342. https://doi.org/10.2307/4083084
Emlen ST (1970) Celestial rotation: its importance in the development of migratory orientation. Science 170:1198–1201. https://doi.org/10.1126/science.170.3963.1198
Emlen ST, Emlen JT (1966) A technique for recording orientation of captive birds. Auk 83:361–367. https://doi.org/10.2307/4083048
Engels S, Schneider NL, Lefeldt N, Hein CM, Zapka M, Michalik A, Elbers D, Kittel A, Hore PJ, Mouritsen H (2014) Anthropogenic electromagnetic noise disrupts magnetic compass orientation in a migratory bird. Nature 509:353–356. https://doi.org/10.1038/nature13290
Granger J, Cummer SA, Lohmann KJ, Johnsen S (2022) Environmental sources of radio frequency noise: potential impacts on magnetoreception. J Comp Physiol A 208:83–95. https://doi.org/10.1007/s00359-021-01516-z
Hiscock HG, Mouritsen H, Manolopoulos DE, Hore PJ (2017) Disruption of magnetic compass orientation in migratory birds by radiofrequency electromagnetic fields. Biophys J 113:1475–1484. https://doi.org/10.1016/j.bpj.2017.07.031
Hore PJ, Mouritsen H (2016) The radical-pair mechanism of magnetoreception. Annu Rev Biophys 45:299–344. https://doi.org/10.1146/annurev-biophys-032116-094545
Kavokin K (2009) The puzzle of magnetic resonance effect on the magnetic compass of migratory birds. Bioelectromagnetics 30:402–410. https://doi.org/10.1002/bem.20485
Kavokin K (2017) Can a hybrid chemical-ferromagnetic model of the avian compass explain its outstanding sensitivity to magnetic noise? PLoS ONE 12:e0173887. https://doi.org/10.1371/journal.pone.0173887
Kavokin K, Chernetsov N, Pakhomov A, Bojarinova J, Kobylkov D, Namozov B (2014) Magnetic orientation of garden warblers (Sylvia borin) under 1.4 MHz radiofrequency magnetic field. J R Soc Interface 11:20140451. https://doi.org/10.1098/rsif.2014.0451
Kirschvink JL (2014) Sensory biology: radio waves zap the biomagnetic compass. Nature 509:296–297. https://doi.org/10.1038/nature13334
Kobylkov D, Wynn J, Winklhofer M, Chetverikova R, Xu J, Hiscock H, Hore PJ, Mouritsen H (2019) Electromagnetic 0.1–100kHz noise does not disrupt orientation in a night-migrating songbird implying a spin coherence lifetime of less than 10µs. J R Soc Interface 16:20190716. https://doi.org/10.1098/rsif.2019.0716
Kramer G (1957) Experiments on bird orientation and their interpretation. Ibis 99:196–227. https://doi.org/10.1111/j.1474-919X.1957.tb01947.x
Landler L, Painter MS, Youmans PW, Hopkins WA, Phillips JB (2015) Spontaneous magnetic alignment by yearling snapping turtles: rapid association of radio frequency dependent pattern of magnetic input with novel surroundings. PLoS ONE 10(5):e0124728. https://doi.org/10.1371/journal.pone.0124728
Leberecht B, Kobylkov D, Karwinkel T et al (2022) Broadband 75–85 MHz radiofrequency fields disrupt magnetic compass orientation in night-migratory songbirds consistent with a flavin-based radical pair magnetoreceptor. J Comp Physiol A 208:97–106. https://doi.org/10.1007/s00359-021-01537-8
Leberecht B, Wong SY, Satish B et al (2023) Upper bound for broadband radiofrequency field disruption of magnetic compass orientation in night-migratory songbirds. PNAS 120(28):e2301153120. https://doi.org/10.1073/pnas.2301153120
Malkemper EP, Eder SHK, Begall S, Phillips JB, Winklhofer M, Hart V, Burda H (2015) Magnetoreception in the wood mouse (Apodemus sylvaticus): infuence of weak frequency-modulated radio frequency fields. Sci Rep 5:9917. https://doi.org/10.1038/srep09917
Mouritsen H (2018) Long-distance navigation and magnetoreception in migratory animals. Nature 558:50–59. https://doi.org/10.1038/s41586-018-0176-1
Mouritsen H, Larsen ON (1998) Evaluating alternative methods for the analysis of Emlen funnel data. In: Mouritsen H (ed) Compasses and orientational strategies of night migrating passerine birds. Center of Sound Communication, Institute of Biology, Odense University, Odense, pp 115–124
Pakhomov A, Chernetsov N (2014) Early evening activity of migratory garden warbler Sylvia borin: compass calibration activity? J Ornithol 155:621–630. https://doi.org/10.1007/s10336-014-1044-x
Pakhomov A, Chernetsov N (2020) A hierarchy of compass systems in migratory birds. Biol Comm 65:262–276. https://doi.org/10.21638/spbu03.2020.306
Pakhomov A, Bojarinova J, Cherbunin R, Chetverikova R, Grigoryev PS, Kavokin K, Kobylkov D, Lubkovskaja R, Chernetsov N (2017) Very weak oscillating magnetic field disrupts the magnetic compass of songbird migrants. J R Soc Interface 14:20170364. https://doi.org/10.1098/rsif.2017.0364
Pakhomov A, Prokshina A, Cellarius F, Mouritsen H, Chernetsov N (2022) Access to the sky near the horizon and stars does not play a crucial role in compass calibration of European songbird migrants. J Exp Biol 225:jeb243631. https://doi.org/10.1242/jeb.243631
Ritz T, Adem S, Schulten K (2000) A model for photoreceptor-based magnetoreception in birds. Biophys 78:707–718. https://doi.org/10.1016/S0006-3495(00)76629-X
Ritz T, Thalau P, Phillips JB, Wiltschko R, Wiltschko W (2004) Resonance effects indicate a radical pair mechanism for avian magnetic compass. Nature 429:177–180. https://doi.org/10.1038/nature02534
Ritz T, Wiltschko R, Hore PJ, Rodgers CT, Stapput K, Thalau P, Timmel CR, Wiltschko W (2009) Magnetic compass of birds is based on a molecule with optimal directional sensitivity. Biophys J 96:3451–3457. https://doi.org/10.1016/j.bpj.2008.11.072
Sauer F (1957) Die Sternenorientierung nächtlich ziehender Grasmücken (Sylvia atricapilla, borin und curruca). Z Tierpsychol 14:20–70. https://doi.org/10.1111/j.1439-0310.1957.tb00525.x
Schmidt-Koenig K (1990) The sun compass. Experientia 46:336–342. https://doi.org/10.1007/BF01952166
Schwarze S, Schneider NL, Reichl T, Dreyer D, Lefeldt N, Engels S, Baker N, Hore PJ, Mouritsen H (2016) Weak broadband electromagnetic fields are more disruptive to magnetic compass orientation in a night-migratory songbird (Erithacus rubecula) than strong narrow-band fields. Front Behav Neurosci 10:55. https://doi.org/10.3389/fnbeh.2016.00055
Thalau P, Ritz T, Stapput K, Wiltschko R, Wiltschko W (2005) Magnetic compass orientation of migratory birds in the presence of a 1.315MHz oscillating field. Naturwissenschaften 92:86–90. https://doi.org/10.1007/s00114-004-0595-8
Timmel CR, Hore PJ (1996) Oscillating magnetic field effects on the yields of radical pair reactions. Chem Phys Lett 257:401–408. https://doi.org/10.1016/0009-2614(96)00466-6
Wiltschko W, Wiltschko R (1972) Magnetic compass of European robins. Science 176:62–64. https://doi.org/10.1126/science.176.4030.62
Wiltschko R, Wiltschko W (2015) Avian navigation: a combination of innate and learned mechanisms. Adv Study Behav 47:229–310. https://doi.org/10.1016/bs.asb.2014.12.002
Wiltschko R, Wiltschko W (2019) Magnetoreception in birds. J R Soc Interface 16:20190295. https://doi.org/10.1098/rsif.2019.0295
Acknowledgements
We are grateful to Anisia Prokshina for assistance in the experiments. The authors acknowledge support from the Russian Science Foundation (grant 21-14-00158). We cordially thank Prof. Henrik Mouritsen and two anonymous reviewers for their helpful comments on an earlier version of the manuscript.
Funding
Russian Science Foundation, 21-14-00158, Nikita Chernetsov.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing or financial interests.
Ethical approval
All animal procedures (in this case, capture of the birds and simple, non-invasive, behavioural experiments) were approved by the appropriate authorities: Permit 24/2018-06 by Kaliningrad Regional Agency for Protection, Reproduction, and Use of Animal World and Forests; and Permit #2–3/2022 (dated 24/02/2022) by the Bioethics Committee of Sechenov Institute of Evolutionary Physiology and Biochemistry RAS. All experiments were performed in accordance with relevant guidelines and regulations. The birds were released back into the wild after all experiments had been completed.
Additional information
Communicated by H. Mouritsen.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bojarinova, J., Kavokin, K., Fedorishcheva, A. et al. Oscillating magnetic field does not disrupt orientation in the presence of stellar cues in an avian migrant. J Ornithol 165, 347–354 (2024). https://doi.org/10.1007/s10336-023-02129-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-023-02129-w