
Abstract
The decision for a migratory animal to be site faithful in its non-breeding season has profound implications for migratory connectivity, resilience to winter habitat loss and population dynamics through carry-over effects on future breeding success and fitness. Knowledge of the temporal and spatial scale of site fidelity and dispersal is also central to accurate survival estimates. We established the observed spatial and temporal scale of site fidelity and the ability to detect small-scale dispersal within and between years for a wintering long-distance Palearctic migrant, the Whinchat Saxicola rubetra, by comparing predicted and observed detection rates within the study site. Across 2 years, 54 % of birds returned to the study site and all returning birds reoccupied the territories they used in the previous winter. Observed dispersal was very low despite the high probability of detecting any local dispersal, suggesting that return rates are indicative of true between-winter survival rates for this population. In any winter, 50 % of returning individuals had a previously occupied but now empty territory that was less than one territory-span away from the centre of their current territory; high site fidelity was therefore very unlikely to be because of limited territory availability. Over-winter residency time (defined by departure month) differed significantly across sites and with age, but did not determine the probability of whether a bird returned in the following year. This suggests the use of more than one wintering site for some individuals, rather than reduced over-winter survival. This study is one of the first to comprehensively document site fidelity at the territory scale in a Palearctic system, although less comprehensive studies or anecdotal evidence suggests that high winter site fidelity may be relatively common. Here we provide evidence for the serial residency hypothesis, where selection acts for individual migrants to have generalist habitat requirements, allowing them to survive in and remain site faithful to even relatively low-quality, but sufficient and familiar sites. Lower dispersal and higher site fidelity compared to that during breeding suggest that annual survival estimates are more accurate when measured on the wintering grounds. This study supports previous findings that wintering conditions do not limit Whinchat populations.
Zusammenfassung
Hohe Wintergebietstreue bei einem Langstreckenzieher und deren Bedeutung für Winterökologie und Überlebensraten
Entscheidet sich ein ziehender Organismus außerhalb der Brutzeit zur Ortstreue, so hat dies weitreichende Folgen für Zugkonnektivität, Toleranz gegenüber Lebensraumverlusten im Wintergebiet und - aufgrund von zeitverzögerten Effekten auf zukünftigen Bruterfolg und Fitness - für die Populationsdynamik. Wissen über das zeitliche und räumliche Ausmaß von Ortstreue und Abwanderung ist außerdem von entscheidender Bedeutung für genaue Schätzungen von Überlebensraten. Durch den Vergleich vorhergesagter und tatsächlicher Nachweisraten im Untersuchungsgebiet bestimmten wir das beobachtete räumliche und zeitliche Ausmaß der Ortstreue und der Nachweisbarkeit kleinräumiger Abwanderung sowohl innerhalb als auch zwischen verschiedenen Jahren bei einem überwinternden Langstreckenzieher, dem Braunkehlchen Saxicola rubetra. Über einen Zeitraum von zwei Jahren kehrten 54 % der Vögel ins Untersuchungsgebiet zurück und alle Rückkehrer besetzten wieder ihre Territorien vom vorigen Winter. Obwohl die Wahrscheinlichkeit, lokale Abwanderung zu registrieren, sehr hoch war, lag die beobachtete Abwanderung trotzdem sehr niedrig, was ein Zeichen dafür ist, dass die Rückkehrraten die tatsächlichen Überlebensraten von einem Winter zum nächsten für diese Population gut abbilden. In jedem der Winter befand sich bei 50 % der Rückkehrer ein vormals besetztes aber nun vakantes Revier in weniger als einer Territoriumsspanne Abstand vom Zentrum; die große Ortstreue wird daher höchstwahrscheinlich nicht durch begrenzte Revierverfügbarkeit bedingt. Die Überwinterungsdauer (definiert durch den Monat des Abzugs) variierte signifikant zwischen den Orten und mit dem Alter, war aber nicht bestimmend für die Wahrscheinlichkeit dafür, ob ein Vogel im Folgejahr zurückkehrt. Dies spricht eher dafür, dass manche Individuen mehrere Überwinterungsgebiete nutzen als für verringerte Winterüberlebensraten. Dies ist eine der ersten Studien zur umfassenden Dokumentation von Ortstreue auf Revierniveau in einem paläarktischen System, obgleich weniger übergreifende Studien sowie anekdotische Hinweise nahelegen, dass eine hohe Wintergebietstreue recht häufig sein könnte. Hier liefern wir Belege für die Serial Residency-Hypothese, bei der die Selektion bewirkt, dass individuelle Zugvögel Lebensraumgeneralisten sind, die in Gebieten von relativ schlechter Qualität überleben können und diesen treu bleiben, so lange es genug davon gibt und sie mit diesen vertraut sind. Eine geringere Abwanderung und höhere Ortstreue als zur Brutzeit spricht dafür, dass die jährlichen Überlebensraten genauer geschätzt werden können, wenn sie in den Überwinterungsgebieten bestimmt werden. Diese Untersuchung bestätigt ältere Befunde, die besagen, dass Braunkehlchenpopulationen nicht durch die Überwinterungsbedingungen begrenzt werden.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The non-breeding period is a significant part of the yearly cycle for migrant birds and wintering ecology has significant carry-over effects for many aspects of survival and reproduction (Baillie and Peach 1992; Sherry and Holmes 1996; Norris et al. 2004; Both et al. 2006; Newton 2006, 2010a; Pulido 2007; Studds et al. 2008; Reudink et al. 2009). A key decision for non-breeding migrants is whether to maintain a winter territory and whether to return to that same wintering site between years. Many migrants have been shown to be site faithful in the non-breeding season to some degree (McNeil 1982; Holmes and Sherry 1992; Sauvage et al. 1998; Marra 2000; Salewski et al. 2000; King and Hutchinson 2001; Cresswell 2014). The scale of this site fidelity has substantial consequences for population dynamics and migratory connectivity (Newton 2010a; Cresswell 2014), because how dependent a migrant is on specific wintering areas will determine how susceptible (or resilient) that species is to the loss or alteration of those wintering sites. Migrants that rely upon a larger number of wintering sites run the greater risk of being impacted by site loss or alteration, for example, because the chance that one of their wintering sites will be impacted by environmental change is greater when more sites are used [known as the ‘multiple jeopardy’ hypothesis (Newton 2004)].
The amount of suitable winter habitat that may be available and the ability to stay resident in a territory of poorer quality are likely to be greater for migrants with more generalist wintering requirements within their wintering habitats (Cresswell 2014). Migrant birds, particularly passerines wintering within Africa, are likely to be habitat generalists during winter and to be faithful to any site that promotes their survival because of the stochastic nature of site selection on a very large scale by birds in their first winter (the ‘serial residency hypothesis’: Cresswell 2014). If wintering sites are lost or change considerably, individuals with a low dependency on specific sites or those that can use a wider range of winter habitat conditions can avoid the high costs and unpredictability of moving long distances, and should have higher survival as a result (Warkentin and Hernandez 1996; Cresswell 2014).
At a finer spatial scale, maintaining a territory during the winter as opposed to being itinerant instils knowledge of local food resources and aids territory defence and predator avoidance (Brown and Long 2007; Förschler et al. 2010). Whether birds shift territories within winters or occupy a different territory upon return may suggest how important winter territory quality is for survival and future fitness. Within- or between-winter territory switching suggests competition for higher-quality territories (i.e. dominance-based territory occupancy) and that territory quality influences survival or future reproductive success (Marra and Holmes 2001). Wintering in suboptimal habitats has been shown to lower reproductive success for some migrants (Norris et al. 2004; Reudink et al. 2009), and this has implications at the population level if habitat loss forces a higher proportion of individuals into lower-quality habitats. Furthermore, if the degree of site fidelity differs with age or sex, any resulting differential survival may lead to skewed population dynamics and related population declines (Sherry and Holmes 1996; Marra 2000; Marra and Holmes 2001; Steifetten and Dale 2006).
The scale of site fidelity is also an essential component of survival estimates, because measuring the scale of site fidelity allows the estimation of “true” survival as opposed to “apparent survival”, the latter being a function of the scale and intensity of monitoring and is less valuable for population management (Anders and Marshall 2005; Gilroy et al. 2012; Ergon and Gardner 2013; Schaub and Royle 2013). If migrants exhibit some degree of wintering site fidelity, survival can always be estimated from return rates, at least to some extent (Sauvage et al. 1998; Salewski et al. 2000; King and Hutchinson 2001); yet determining true survival is problematic because the chance of resighting an individual depends on a combination of survival and dispersal plus the ability to detect returning individuals (Marshall et al. 2000, 2004; Anders and Marshall 2005; Schaub and Royle 2013). Distinguishing between survival and dispersal is challenging, especially when exploring population dynamics between years (Marshall et al. 2004; Ergon and Gardner 2013), and the ability to detect individuals is often a function of the size of the study area and resighting effort (Baker et al. 1995; Marshall et al. 2000, 2004). Consequently, survival estimates based on return rates often underestimate true survival (Gilroy et al. 2012; Ergon and Gardner 2013; Schaub and Royle 2013). Determining both where a species lies on the continuum between 100 % between-year survival and 100 % site fidelity and the degree of any dispersal are therefore fundamental to establishing the value of survival estimates calculated from return rates. This can be essential to understanding population dynamics. Winter site fidelity appears to be relatively common amongst migrants (Skilleter 1995; Sauvage et al. 1998; Salewski et al. 2000; Koronkiewicz et al. 2006; Barshep et al. 2012; Cresswell 2014), implying that true survival estimates can be obtained from return rates, but only if detection probability is high because return rates are the product of both apparent survival probability and detection probability. Studies of this detail during winter are lacking, however, especially for Palearctic migrants (Vickery et al. 2014). Furthermore, accurate between-winter return rates and the temporal scale of within-winter site fidelity are essential to understanding whether short-term residency at any one wintering site is due to mortality or dispersal to other areas.
In this study we aimed to establish the degree of site fidelity and quantify the ability to detect any dispersal during winter for a declining long-distance Palearctic migrant, the Whinchat Saxicola rubetra. This species exhibits varying degrees of fidelity in both the breeding and non-breeding seasons (Bastian 1992; Barshep et al. 2012; Shitikov et al. 2012). Some breeding populations have declined significantly over the past few decades, as illustrated by the 53.1 % decline in breeding abundance documented over the past 40 years (1970–2010) (BirdLife International 2004; Henderson et al. 2004; Henderson et al. 2015). Similar population trends have been shown for many long-distance Palearctic migrants, the causes of which are often poorly understood (Berthold et al. 1998; Sanderson et al. 2006; Thaxter et al. 2010; Vickery et al. 2014), and so establishing whether migrants are flexible on the wintering grounds and the degree of site fidelity has profound implications for the conservation of migrants. This is particularly relevant given the increasing pressure of anthropogenic habitat loss in Africa (Vickery et al. 2014).
We ask: (1) do individuals return to the same wintering area and do return rates differ with age and sex; (2) what is the power to detect returning individuals and within- and between-year dispersal over different spatial scales, and what opportunities are there for individuals to disperse; (3) what is the spatial scale of site fidelity and dispersal observed; and (4) what is the temporal scale of site fidelity and does this differ with age and sex?
Methods
Study site
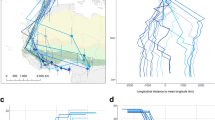
The study took place over three consecutive winters (hereby referred to chronologically as Winter 1, 2 and 3) from January 2011 to March 2014 on the Jos Plateau in the Guinea savannah zone of central Nigeria, West Africa (N09°53′, E08°59′, approximately 1,250 m altitude; Fig. 1). The region experiences wet and dry seasons, with the majority of the wintering period for migrants (early September–late April) within the dry season. Study sites were primarily open scrubland with varying degrees of degradation due to human habitation, arable farming and livestock grazing, the last two often increasing in intensity as the dry season progresses (Hulme and Cresswell 2012). Sites with high densities of Whinchats were chosen and are typical of wintering habitat for this species in the area. Three study sites were used in all winters, Sites A–C, and two Sites D and E were added at the end of the Winter 2 (Fig. 1). Birds were captured at two additional sites in Winter 3 (marked ‘x’ Fig. 1) but are only included in analyses exploring age and sex ratios across sites. Note that sites represent logistic areas, rather than biologically different distinct areas identified a priori: we therefore consider site effects as a potentially confounding nuisance variable (e.g. sampling effort may have varied across sites) rather than representing anything likely to have been correlated with Whinchat biology. Sites in any case were very close together and site boundaries only existed as defined by us during the study.

Map of the six study sites and their location within Nigeria in West Africa (filled circle). Sites A, B and C were used in all three winters; Sites D and E in Winters 2 and 3, and sites marked x used in Winter 3 only (see “Methods”). Note that sites exist for logistical reasons rather than representing biologically distinct areas, although they are considered because of variation in sampling time and effort with site
Study subjects
Birds were captured with spring traps and mist nets using conspecific playback and live bait. In Winter 1, birds were captured from late January to mid-February; in Winter 2, birds were captured from late September until mid-November, and from mid-February to early March. Additional birds were captured in Winter 3 but are excluded from analyses involving return rates and territory size because return rates were unknown and resightings were less precise in Winter 3 (see below). Each bird was uniquely colour ringed, sexed and aged as either first-winter or adult (Jenni and Winkler 2004), and biometric information (maximum wing cord, tarsus, mass) and moult score were recorded. The age and sex of birds caught did not differ between the study sites (Chi-squared tests on all birds captured across study: age: χ 2 = 2.7, df = 5, p = 0.75, n = 409; sex: χ 2 = 5.9, p = 0.32, n = 407). In total, 176 birds were captured in Winters 1 and 2, of which 154 were resighted at least once after capture.
Resighting individuals
Individuals were resighted throughout winters to determine (1) the location and size of territories (Winters 1 and 2), and (2) the degree of residency and site fidelity within and between winters (all years). In Winters 1 and 2, resighting efforts focussed on obtaining undisturbed resightings to gain accurate estimates of territory size and location and winter residency. In Winter 3, resightings were less precise because focus shifted to just relocating birds. Resightings were carried out from dawn (ca. 0600 h) until 1100 h and from 1600 h until dusk (ca. 1830 h). Because Whinchats typically perch on shrubs and other structures where they are easily visible and make frequent sallies to fly-catch and feed from the ground, birds were easily detected if they were present. For Winter 2 (for which we later determined departure month and resighting probability, see below), all territories were visited at least weekly from late September to mid-May (but often more frequently) until a bird was considered to have departed, as determined by not resighting an individual after four territory visits. Note that because territories were spread throughout the study area and were close together, we often continued to visit territories of departed birds (i.e. when visiting other territories). We are therefore confident that departure month could be accurately determined. Whinchats were located with binoculars (Swarovski SV 8 × 32) and colour combinations read using a spotting scope (Zeiss Diascope 65 mm with ×25 eyepiece). Locations of ringed individuals were recorded with a GPS device (Garmin GPSMAP 64). To enable unbiased estimates of the territory location, and to determine territory size and use, each sighting was scored for confidence regarding whether the bird was undisturbed before being sighted, and we recorded the date and time to the nearest minute of each resighting. A Whinchat could travel across its territory within seconds and the time between observations was found to have no influence on the spatial independence of resightings [general linear model (LM) of distance between resightings ~ observation time (controlling for individual): F 1,762 = 0.02, p = 0.90; 765 observations of 119 individuals]. Coordinates were converted to UTM units (Zone 32N) for analyses to give position in metres on the globe. Resightings were viewed and edited where necessary in ArcMAP 10.1 (ERSI 2012) and Garmin BaseCamp software (version 4.2.4; Garmin International Inc., Olathe, KS, USA).
Calculating territory size
The conventional method of determining territory or range size for animals is with minimum convex polygons (MCP), but this method requires many resightings for accurate descriptions of space use and is subject to several biases (Hansteen et al. 1997; Börger et al. 2006). Our aim was to determine the area a bird used over winter and the size and location of its territory, rather than exact territory boundaries. Therefore we determined an index of territory size from resightings scored as undisturbed for birds resighted in Winters 1 and 2. For each individual, the territory centre was determined by averaging the position of all resightings. The mean distance between each resighting and the territory centre was then calculated and doubled to give an estimate of territory diameter. This was used as a measure of territory size. Territory size was calculated for individuals with at least five undisturbed resightings from W inters 1 and 2. Five was chosen as a minimum number in order to reduce the influence of increasing resighting sample size on estimates of territory size (Börger et al. 2006), whilst still maintaining adequate sample sizes. Of the 35 and 67 birds resighted after capture in Winters 1 and 2, respectively, 33 and 39 individuals had at least five independent resightings, respectively. If the minimum number of resightings required to calculate territory size was increased to 10, the average territory diameter was altered by 4 m (6 %) and the results of all analyses including territory size were unchanged, suggesting that five resightings were adequate for analyses. We used an individual’s average territory diameter across years when its territory size was measured in multiple winters. Coordinates recorded with GPS devices have an error of approximately ±5 m (our device gave an accuracy of approximately ±2–3 m in Nigeria), which may have influenced our measures of territory size; however, we assume here that this error was similar across territories and should affect each individual measure of territory size similarly. We therefore did not adjust for GPS inaccuracy when calculating territory size.
Establishing the scale of site fidelity
To determine the degree of observed site fidelity for Winter 1 birds returning in Winter 2, we compared the distance between territory centres between years with territory size. Because territory size was not calculated in Winter 3 for birds returning from Winter 2 as a result of the less precise resighting methodology, we compared the distances between the mean central point of resighting locations in Winter 3 and the mean central point of all available resighting locations in Winter 2 to establish the degree of site fidelity in Winter 3. Some individuals that were ringed as first-winter birds in Winter 1 returned in both Winters 2 and 3. When calculating return rates, these birds were considered as first-winter birds between Winters 1 and 2 and as adults between Winters 2 and 3. When modelling predictors of return rates, any birds from Winter 1 that returned in multiple years were only considered in return rates between Winters 1 and 2 to avoid pseudoreplication. As with calculating territory size, we did not adjust for any GPS inaccuracy because we assume this error to be unbiased and similar across all territories and years.
Establishing resighting effort and detectability
Resighting effort was determined from 94 resighting visits to sites between 1 November 2012 and 24 April 2013 when resighting routes were recorded. Note that many more resighting visits were made to all territories both outside and within this period where effort was not recorded. Visits were either partial (less than 50 % of all territories at the site visited) or complete visits (at least 50 % of all territories visited). A territory was considered visited if the observer walked across at least 30 % of the known territory area. Across all sites, resighting effort was recorded for 33 partial and 25 complete site visits with 82 territories visited in total during this time.
The percentage probability of resighting a bird was calculated as the number of times a bird was seen out of the total visits to its territory for individuals that were resighted at least once during the above period. The probability of detecting an individual across years, should that individual disperse at increasing spatial scales, was estimated for all birds resighted in Winter 2 by calculating the proportion of their territory that would still be included in the study area in Winter 3 after dispersal. Territory diameter was used as territory size. For birds resighted in both Winters 1 and 2, territory diameter from Winter 2 was used. For birds where the territory location was known but territory diameter was not calculated as a result of an insufficient number of resightings (n = 52 birds), we used the mean territory diameter across all individuals (64 ± 1.8 m, range 11–106 m, n = 113). A GIS analysis was used to determine detection probability after dispersal [ArcMAP 10.1 (ERSI 2012)]. To do this, possible territory locations after dispersal were determined using a combination of the ‘buffer’ tool (to create new territories of multiples of the territory size for each individual) and the ‘union’ feature in the overlay analysis toolbox to calculate the location of territories after dispersal and the proportion still within the study area.
Within-winter residency
There was a negative relationship between capture date and residency time if individual residency time was calculated as the period from capture date to last resighting (b = −1.22, t 55 = −2.9, R 2 = 0.12, F 1,53 = 8.5, p < 0.01, n = 55: i.e. birds caught late in the year could only be resident for shorter periods). Furthermore, the probability of resighting a bird was not 100 % for one territory visit. This meant that neither capture date nor departure day could not be used as a biologically meaningful predictor of how long Whinchats chose to spend in a wintering territory. Because of this uncertainty in arrival times and relatively low resolution of measuring departure times, we investigated variation in residency within a winter simply by scoring the month of departure of birds, as determined by not resighting an individual after four territory visits. Departure month was then used as a response variable in a model with sex and age as predictors, and site included as a potential confounding variable, and their interactions. This method was adopted rather than the mark-recapture survival analysis initially attempted because resighting and capture effort varied across the three study winters. To determine whether shorter residency was due to mortality, return rates for the following winter were compared between individuals that left sites early (short residency) and those that remained for at least 50 % of the winter (long residency). Resightings over four winters suggest that Whinchats arrive in mid-September and depart until late April, and so we defined short residency as departure within September, October, November or December, and long residency as departure from 1st January onwards.
Sample sizes and statistical analyses
Birds were only included in the study if they had been resighted at least once after capture. Birds from Sites A–C are included in all analyses, whereas birds from Sites D and E were excluded from analyses involving territory size or winter residency because resighting locations recorded for these individuals were less accurate and none of these birds were ringed within the period required to calculate departure month (see below). One individual was excluded from analyses exploring the probability of detection because its territory was visited too infrequently. Birds which could not be confidently aged or sexed were excluded from models including age and sex as predictors. We modelled month of departure and residency time (short or long) within Winter 2 only, i.e. when resighting effort was highest, and only for birds captured either in Winter 2 before December, or birds returning from Winter 1 and resighted before December. To exclude any transient birds or any capture effects (e.g. relocation after capture), all of these individuals had been resighted at least 30 days after capture. The numbers of individuals included in this study are shown in Table 1, with test-specific sample sizes given within “Results”.
Aspects of wintering ecology were explored with general and generalised LMs carried out in R version 3.0.1 (R Development Core Team 2013) and RStudio version 0.98.507. Analysis of variance (ANOVA) was used to explore differences in resighting probability across sites. Logistic regression (GLM) was used for exploring predictors of return rates and the influence of residency period on the probability of return in the following winter. A general LM was used to explore predictors of departure month. Paired t tests were used to explore differences in territory size between years. Multiple regression model simplification was based on Akaike information criterion (AIC) (Bozdogan 1987) for dredge analyses (see below), where a value of ΔAICc ≥ 2 was used to identify distinct models. For models with fewer variables, we used an ANOVA comparison between models to remove variables that failed to add a significant contribution to a model. For models of the same or similar sample size, a ‘dredge’ analysis was used to identify the best, simplest models (Bartoń 2012): cases with missing values were removed from the data set as required for dredge analyses, and resulting best models were produced using complete data sets. When presenting top models from dredge analyses, we presented models within four AIC of the top model. Model fits were evaluated from diagnostic model plots and models were presented if assumptions were reasonably met (Crawley 2007). Data were checked for normality where necessary and for multicollinearity with variance inflation factors in the CARS package (Fox and Weisberg 2010). Mean values are presented as mean ± 1 standard error in all cases. A statistical significance level of p < 0.05 was chosen to reject null hypotheses.
Results
Return rates to the study site and influence of age and sex
Return rates were 58 % in Winter 2 and 52 % in Winter 3, averaging 54 % across the two winters. In no cases did a bird fail to return in its second winter but then return in its third winter. A bird’s age or sex did not influence the probability of resighting that individual in the following winter [logistic regression: model averaged parameters of top 11 models within four AIC of top model: age: z = 0.7, p = 0.47; sex: z = 0.04, p = 0.97; body size at time of capture, winter and site also included within main model (all terms including interaction terms between age, sex and site not significant); n = 88].
Power to detect dispersal and opportunity for dispersal
Probability of detection within winters
Across all individuals, the probability of resighting an individual that was seen at least once after ringing if its territory was visited once was 63 % (±2.4 %, range 11–100 %, n = 81). Each time a site was visited, 59 % of birds on average were detected out of all birds whose territories were visited (±3.3 %, range 0–100 % detected, n = 77). For the period for which resighting effort was known, on average each territory was visited nine times (±0.51, range 1–18, n = 81); note that further visits were made to territories where resighting effort was not recorded. When accounting for the probability of encountering a bird when its territory was visited, there was a probability of almost 100 % of detecting a resident individual during a study winter.
The probability of resighting an individual did not vary between sites (F 2 = 2.6, p = 0.08, n = 81) nor with the number of days into the study (F 1,78 = 0.5, p = 0.47, n = 81; Fig. 2a). Similarly, the proportions of individuals resighted each visit did not differ between sites (F 2,74 = 0.5, p = 0.63, n = 77) nor with number of days into the study (F 1,75 = 0.6, p = 0.44, n = 77; Fig. 2b).

a The percentage chance of resighting a bird if its territory was visited, and b the proportion of birds resighted out of the number of territories visited during a site visit, across the three study sites. There was no difference in the probability of seeing a bird, or in the proportion of birds detected, during visits across sites
Probability of detecting dispersal between years
The average territory diameter across all birds in both years was 64 m (±1.8 m, range 11–106 m, n = 113, Fig. 3). Across all birds, shifting one territory upon return reduced the probability of remaining in the study area by on average 7 %, and a returning bird could disperse up to eight territories from the previous winter before the chance of it remaining within the study area dropped below 50 % (n = 130; Table 2; Fig. 3). At least 80 % of the territory would remain in the study area for 25 % of all birds (n = 32) after shifting as many as six territories (Fig. 3). For birds resighted in Winter 2, a 500-m shift from the previous winter’s territory resulted in a 44 % probability of a bird staying in the study area. No birds would have been within the study area if they dispersed 10 km or more in the following year (Table 2).

The probability of detecting a returning bird within the study site in the following year if small-scale dispersal occurred from the territory location in the previous year (grey points, as determined by the proportion of the study area that could contain the territory after dispersal), and the maximum distance individuals were actually observed to move the following year (grey bars; n = 54). The shade of each point represents the frequency of individuals with that value, where darker shades represent higher frequencies, starting from black (furthest left data point). The mean proportion of the new territory within the study area should dispersal occur is also shown (solid black line) with upper and lower CIs (dashed lines)
Overall power of estimating larger-scale dispersal
Overall, 89 individuals were known to be resident in Winter 2. With observed return rates of 54 % and site fidelity at the territory level, 48 individuals returned to their territories in the following year. Of the remaining 41 individuals, assuming all survived, eight, four and two birds should have been found within the study area if they dispersed 0.5, 1 or 5 km, respectively (Fig. 4). If only 50 % (n = 20) of these individuals had survived and dispersed, four, two and one birds, respectively, should have been detected away from their territories.

The number of birds estimated to be within the study area if dispersal occurred of 0.5, 1 and 5 km upon return to the study site in the following winter assuming a return rate of 54 %, as a function of the total number of birds studied in a year. The actual number of individuals we studied (i.e. where territory diameter was measured) is also shown (filled diamond; n = 112)
Opportunity for local dispersal
Across all returning birds, the average distance between their territory centre in the first year and the centre of the nearest unoccupied territory in the following year (i.e. a territory that was occupied in the previous winter but now empty) was 107 m (±10.0 m, range 7–538 m, n = 76, Fig. 5). On average, each individual had at least one known unoccupied territory within 100 m, and three unoccupied territories within 200 m of the centre of the territory in the previous year (n = 80). For birds for which the location of the territory centre was accurately known in both years, 50 % of individuals had unoccupied territories that were closer to the territory centre than their average territory diameter (n = 19).

Map of study Site A showing the territories in both years of birds ringed in Winter 1 that returned in Winter 2, birds that did not return in Winter 2, and birds newly ringed in Winter 2, identified by age. Each circle represents a Whinchat territory in either Winter 1 or 2, the radius of which is equal to the individual’s index of territory size (m). Connecting lines join territories belonging to the same individual in Winters 1 and 2. Note that territories of birds ringed in Winter 2 were not necessarily unoccupied in Winter 1. Stippled areas show areas of thick scrub; areas with tree symbols show wooded patches. All territories of non-returning birds and territories occupied in Winter 2 by first-year birds represent territories available to birds returning from Winter 1. The inset figure shows the average territory size in each winter and the average distance between territories of a returning individual in Winters 1 and 2. Overall the figure illustrates that birds returned to the same territory used in the previous year despite many empty territories being available close by
Scale of spatial site fidelity and levels of dispersal detected
Winter 1 versus Winter 2
The average movement between territory centres in Winter 1 versus Winter 2 was 30 m (±4.0 m, range 6–58 m, n = 19; Fig. 5). The furthest movement was 58.3 m. For all individuals, any movement in the centre of the territory between years was less than the territory size for that individual in Winter 1 (average difference between movement across years and territory size in Winter 1 = −35.5 ± 3.4 m, range −61 to −2 m; Fig. 5), showing that individuals returned to the same territory in Winter 2 and that “movements” observed were likely due to the resolution at which data were collected (e.g. stochasticity from using a limited number of locations to estimate territory size index and GPS accuracy) rather than actual territory shifts.
Birds returning from Winter 1 used a similar sized territory in Winter 2 (paired t test: t = −0.07, df = 17, p = 0.95, n = 18; Fig. 5).
Winter 2 versus Winter 3
Across all individuals, birds were resighted on average 21 m from where they were seen in the previous year (±3.2 m, range 1–118 m, n = 54) and none were resighted further than 118 m from any one known location in the previous year. In terms of range size, the average difference between the maximum range in Winter 2 (as determined by the furthest distance between known locations) and the distance between the furthest resighting in Winter 3 from a known location in Winter 2 was 55 m (±5.5 m, range 2–165 m), equal to less than the width of one territory (Fig. 3).
Temporal scale of fidelity
Degree of residency and predictors of residency period
The minimum number of days a bird was known to be resident at a site (i.e. from capture until last resighting for birds ringed in Winter 2, and from first to last resighting for birds returning from the previous winter) was 31–180 days (mean 107 ± 4.9 days, n = 67). Departure month varied from November to March (12, 9, 28, 24, 27 % departing November, December, January, February and March, respectively, n = 67 birds). Departure month differed significantly across sites and with age, with birds spending less of the winter at Sites B and C relative to Site A, and departing later if they were first-winter birds (Table 3; Fig. 6).

Departure month (from September, where September = month 1) according to age (first-winter or adult) and site. Values are predicted from the model: LM of departure month ~ site + age (Table 3, n = 63). Departure month differed significantly between sites and with age
Relationships between residency period and probability of return
Ten of 14 birds (71 %) with short residency periods and 33 of 53 birds (62 %) with long residency periods returned in the following winter (n = 67). Return rates did not differ between birds with short or long residency periods when controlling for site (Table 4); i.e. birds which were resighted for only a short duration of the winter were just as likely to return in the following winter as birds that were resident for most of the winter. We also tested month of departure as a continuous variable to see if departure date was a proxy for within-winter survival (i.e. early departures were in fact more likely to be deaths), but departure month was not a significant predictor when included in the same model (β = −0.31 ± 0.46, z = −0.7, p = 0.50).
Discussion
Return rates, dispersal and the ability to estimate ‘true’ survival
The lack of local dispersal, high detection probability and low probability that large-scale dispersal occurred suggests that a 54 % return rate across the study closely mirrors true between-winter survival for Whinchats. Although we are not aware of another study documenting between-winter return rates or overwinter survival for Whinchats, our return rates in both years are higher or similar to return or survival rates documented for Whinchats on the breeding grounds (return rates of 11–47 %) (Schmidt and Hantge 1954; Bezzel and Stiel 1977; Bastian 1992) and apparent survival estimates of 48 % in adult males, 21 % in adult females and 17 % in juveniles in Switzerland (Müller et al. 2005), and 27 % in Russia (Shitikov et al. 2015). Our return rates are also generally similar or higher than those documented for other site-faithful Palearctic migrants [true survival rates of adult passerines are reported to be 30–60 % (Karr et al. 1990; Payevsky et al. 1997; Siriwardena et al. 1998)]: e.g. Great Reed Warbler Acrocephalus arundinaceus, return rate of 55 % (Bensch and Hasselquist 1991); Garden Warbler Sylvia borin, apparent survival estimates of 11–44 % (Shitikov et al. 2013); Yellow Wagtail Motacilla flava, apparent survival estimates of 20–100 and 53 % in Russia and Britain, respectively (Siriwardena et al. 1998; Shitikov et al. 2012); Barn Swallow Hirundo rustica, survival estimates of 36–42 % (Møller and Szép 2005; Robinson et al. 2008); Sand Martin Riparia riparia, survival estimates of 29–31 % (Cowley and Siriwardena 2005; Robinson et al. 2008); Common House Martin Delichon urbicum, survival estimate of 42 % (Robinson et al. 2008); Common Whitethroat Sylvia communis, survival estimates of 9–39 % (Shitikov et al. 2013); Reed Warbler Acrocephalus scirpaceus, survival estimates of 33–60 %; and also many Neotropical migrants (Desante et al. 1995). That winter return rates are higher than return rates documented in the breeding season is not surprising given the high site fidelity we documented and our ability to detect dispersal on the wintering grounds. If this is generally true across migrants, annual survival estimates from the wintering grounds are likely to be more accurate than those estimated on the breeding grounds because dispersal is frequently reported during the breeding season (Paradis et al. 1998; Newton 2010b; Shitikov et al. 2015). Consequently, our results suggest that studies aiming to accurately measure true annual survival for migrants should focus on winter return rates. Most importantly, high overwinter survival (suggested by no evidence of departure month or residency time influencing between-year survival) supports previous findings that the decline in breeding Whinchat populations is not a result of wintering conditions (Hulme and Cresswell 2012).
We are confident that our findings of extremely high site fidelity are a true representation of wintering behaviour because of our ability to detect any dispersal occurring on a number of scales. There was a high probability of encountering locally dispersing birds—at least 50 % of ‘missing’ individuals would have been detected if they had dispersed up to eight territories upon return, and nine individuals would have been detected if they moved 0.5 km—yet observed dispersal was extremely low. When we were able to compare territory locations between years, all individuals returned to the same territory they held in the previous winter and no bird was resighted more than 118 m from where it had been seen in the previous year. Therefore, if dispersal did occur, this was only on a larger scale (i.e. greater than 5 km). As we did not have the ability to detect these movements we cannot rule out that this population could comprise both highly resident and widely dispersing individuals. However, the similarity between observed return rates and those expected when comparing studies of other migrants and return rates on the breeding grounds (see above) plus extremely high site fidelity at the territory scale implies that any large-scale dispersal was unlikely.
Site fidelity and wintering strategy
To our knowledge, this is one of the first studies to document site fidelity for a Palearctic migrant on the wintering grounds and to further explore the influence of local dispersal on this. Our results support previous observations that Whinchats are site faithful and territorial (Barshep et al. 2012), but at a much finer scale than originally thought. Although a shift of on average 30 m occurred between years, territory centres were always within the boundaries of the old territories, and these shifts are likely an artefact of the resolution at which data were recorded and the inherent inaccuracy of GPS devices. Repeat occupancy of winter territories has been documented in several other migrants (Kricher and Davis 1986; Holmes et al. 1989; Kelsey 1989; Cuadrado 1992; Holmes and Sherry 1992; Dejaifve 1994; Skilleter 1995; Sauvage et al. 1998; Marra 2000; Salewski et al. 2000; Koronkiewicz et al. 2006), along with territoriality over winter (Kelsey 1989; Bates 1992; Brown et al. 2000; Latta and Faaborg 2001; Salewski et al. 2002; Rappole et al. 2003), indicating that high site fidelity may be relatively common across long-distance migrants.
Our findings suggest that the benefits of winter territoriality and site fidelity outweigh the benefits of competing for or moving to higher-quality territories during winter. High site fidelity most likely results in site familiarity and greater predictability of foraging opportunities, plus more efficient territory defence and predator avoidance, all of which will promote survival (Brown and Long 2007; Förschler et al. 2010). Moving during winter or not holding a territory, on the other hand, is often associated with reduced survival (Rappole et al. 1989; Winker 1998). Furthermore, the benefits of high site fidelity both within and between years may be greater during winter than during the breeding season because the potential benefits of dispersing to find a mate or a suitable nesting location are absent. Site fidelity on the breeding grounds is often lower than that observed during winter, even for species with high winter site fidelity (Herremans et al. 1995; Koronkiewicz et al. 2006; Tryjanowski et al. 2007). Evidence suggests that this may also be true for Whinchats (Bezzel and Stiel 1977; Bastian 1992; Shitikov et al. 2012), with territory fidelity often reported to be lower on the breeding grounds than what we document here for winter (Bastian 1992 and references therein; I. Henderson and J. Taylor, unpublished data). Most likely, the additional pressure of mate acquisition reduces any benefits of very high fidelity at the territory scale on the breeding grounds because both individuals would have to survive if either is to breed, and dispersal is also required to prevent inbreeding. Breeding site fidelity may therefore only benefit individuals if it occurs at a larger scale, such as fidelity to a larger breeding region. In short, there may be many reasons that force a breeding bird to move—death or movement of a partner, lack of nesting habitat, lack of resources to raise chicks, etc. (e.g. Beheler et al. 2003; Middleton et al. 2006)—while a wintering bird likely requires only a relatively low level of resources to ensure daily survival and therefore has fewer reasons to move but several reasons for remaining site faithful.
Evidence for a generalist wintering strategy and lack of dominance-based habitat occupancy
High site fidelity, especially within winters, should demand a generalist wintering strategy to prevent survival and future reproduction being significantly influenced by fine-scale variation in territory characteristics. As conditions often alter during winter (Blackburn and Cresswell 2015a), a wide range of conditions must be utilised and individuals must be flexible to changing conditions if a winter territory is to be maintained. For example, Salewski et al. (2002) hypothesised that winter territoriality in Pied Flycatchers Ficedula hypoleuca was a result of a more diverse and generalist foraging strategy, enabling individuals to reside in and defend small areas over the winter. Other migrant species have also been suggested to be winter generalists (Latta and Faaborg 2002).
Our results indicate a lack of dominance-based territory occupancy during winter, suggesting that individual Whinchats do not compete much, if at all, for territories of higher quality. Returning Whinchats always returned to the same winter territory, regardless of the proximity of an alternative territory, and neither residency nor wintering ecology differed with age and sex. This suggests that there may be little benefit from shifting small distances (i.e. to a nearby territory). That residency differs only with site, and not with demographics, suggests that larger movements outside of the study area (i.e. to a different wintering site) are likely due to reasons other than competition between birds, and for reasons that are not spatially independent. The degradation of a wintering site, for example, would likely affect all individuals and result in the spatial differences in residency that we found. The lack of small-scale movements suggests that (1) territories did not differ greatly regarding their influence on survival, or (2) that Whinchats have few specialist wintering requirements. Consequently, variation in winter territory habitat characteristics may have few carry-over effects on migration success and future survival, and a ‘good’ winter territory is simply one that promotes survival through the winter. For example, some Whinchats may be able to fatten for migration in a range of conditions, or may fatten for migration at other sites between leaving the winter territory and crossing the Sahara. We cannot disregard the theory that a lack of territory relocation within or between winters may be the result of historic wintering conditions (i.e. before anthropogenic degradation), and that birds are now simply unable or unwilling to relocate even under increasing levels of degradation (Bourn and Wint 1994). However, if this were the case we would expect low overwinter survival and reduced winter survival in more degraded habitats due to carry-over effects, but neither is the case for this population (Blackburn and Cresswell 2015a, b). In contrast to our results, dominance-based winter habitat occupancy has been documented for wintering Red-backed Shrikes Lanius collurio (Herremans 1997), Eastern Great Reed Warblers Acrocephalus orientalis (Nisbet and Medway 1972) and European Robins Erithacus rubetra (Catry et al. 2004), and for many Neotropical migrants, such as American Redstarts Setophaga ruticilla (Marra 2000; Marra and Holmes 2001), Prairie Warblers Setophaga discolour (Latta and Faaborg 2001), Hooded Warblers Setophaga citrina (Lynch et al. 1985; Morton 1990; Stutchbury 1994) and Black-throated Blue Warblers Setophaga caerulescens (Wunderle 1995), amongst others (Ornat and Greenberg 1990). In contrast to the winter, higher territory fidelity has been shown on the breeding grounds for adult Whinchats over first years and for males over females, along with higher return rates for adults over first years (Bastian 1992 and references therein).
Difference in residency times
Only whether birds had short or long residency times, and not the degree of territoriality (i.e. age or sex-specific variation in the degree of territory defence) varied across sites, as is seen also in wintering Pied Flycatchers (Salewski et al. 2002). Even though some Whinchats were resident for only short residency, these individuals were just as likely to return in the following winter, suggesting that some individuals may have multiple wintering sites. Multiple site use by individuals has been described for several migrants in both the Neotropical and Palearctic systems (Belda et al. 2007; McKinnon et al. 2013). Some migrants may move southward during the winter (Cresswell et al. 2009), possibly in response to changing conditions (Jones 1995). The early departure of these individuals may provide territories for winter floaters, the presence of which has been suggested by some studies (Winker 1998), principally because birds that disappear during winter are replaced by new individuals (Morton et al. 1987; Holmes et al. 1989; Brown and Long 2007). As already suggested, any reasons for relocating appear to be linked by site only, and not because of competition between individuals for territories of varying quality, suggesting that Whinchats may relocate if a wintering site becomes unsuitable over the winter. Observations made during this study suggest that whilst some empty territories were reoccupied, others remained empty for the remainder of the winter, even though they had been used previously. Further studies involving more intensive monitoring or tracking with geolocators would be needed to determine the extent of any movements. Because individuals do not appear to compete for territories and do not relocate upon return, even when given the opportunity, and likely have generalist wintering requirements, winter territories are probably plentiful, and therefore not all suitable territories are occupied.
In summary, we documented extremely high site fidelity by individuals both between and within winters, and a lack of dominance-based territory occupancy for a declining long-distance migrant. We provide strong evidence for a generalist wintering strategy and suggest that winter territory quality may have little influence on future survival and thus population declines. This study supports the serial residency hypothesis (Cresswell 2014), where selection acts for most migrants to have generalist habitat requirements, allowing them to survive in and remain faithful to even relatively low-quality, but adequate and familiar sites. As a result of lower dispersal and higher site fidelity during winter compared to breeding, the ability to accurately measure annual survival rates may be highest on the wintering grounds. Furthermore, generalist wintering requirements and the seemingly high availability of suitable wintering territories may provide some resilience to the on-going habitat degradation occurring throughout Africa (Bourn and Wint 1994).
References
Anders AD, Marshall MR (2005) Increasing the accuracy of productivity and survival estimates in assessing landbird population status. Conserv Biol 19:66–74. doi:10.1111/j.1523-1739.2005.00543.x
Baillie SR, Peach WJ (1992) Population limitation in Palearctic-African migrant passerines. Ibis 134:120–132. doi:10.1111/j.1474-919X.1992.tb04742.x
Baker M, Nur N, Geupel GR (1995) Correcting biased estimates of dispersal and survival due to limited study area: theory and an application using wrentits. Condor 97:663–674. doi:10.2307/1369175
Barshep Y, Ottosson U, Waldenstroem J, Hulme M, Svensson S (2012) Non-breeding ecology of the whinchat Saxicola rubetra in Nigeria. Ornis Svec 22:25–32
Bartoń K (2012) MuMIn: multi-model inference. R package version. 1
Bastian HV (1992) Breeding and natal dispersal of whinchats Saxicola rubetra. Ringing Migr 13:13–19. doi:10.1080/03078698.1992.9674010
Bates JM (1992) Winter territorial behavior of Gray Vireos. Wilson Bull 104:425–433
Beheler AS, Rhodes OE Jr, Weeks HP Jr, Moore F (2003) Breeding site and mate fidelity in Eastern phoebes (Sayornis phoebe) in Indiana. Auk 120:990–999. doi:10.1642/0004-8038(2003)120[0990:BSAMFI]2.0.CO;2
Belda EJ, Barba E, Monros JS (2007) Resident and transient dynamics, site fidelity and survival in wintering blackcaps Sylvia atricapilla: evidence from capture–recapture analyses. Ibis 149:396–404. doi:10.1111/j.1474-919X.2007.00657.x
Bensch S, Hasselquist D (1991) Territory infidelity in the polygynous great reed warbler Acrocephalus arundinaceus: the effect of variation in territory attractiveness. J Anim Ecol 60:857–871
Berthold P, Fiedler W, Schlenker R, Querner U (1998) 25-year study of the population development of central European songbirds: a general decline, most evident in long-distance migrants. Naturwissenschaften 85:350–353. doi:10.1007/s001140050514
Bezzel E, Stiel K (1977) The biology of the whinchat Saxicola rubetra in the Bavarian Alps. Anz Ornithol Ges Bayern 17:1–9
BirdLife International (2004) Birds in Europe: population estimates, trends and conservation status. BirdLife Conservation Series No. 12
Blackburn E, Cresswell W (2015a) Fine-scale habitat use during the non-breeding season suggests that winter habitat does not limit breeding populations of a declining long-distance Palearctic migrant. J Avian Biol. doi:10.1111/jav.00738
Blackburn E, Cresswell W (2015b) High overwinter and annual survival for a declining Palearctic migrant: evidence that wintering conditions may not limit migrant populations. Ibis (submitted)
Börger L et al (2006) Effects of sampling regime on the mean and variance of home range size estimates. J Anim Ecol 75:1393–1405. doi:10.1111/j.1365-2656.2006.01164.x
Both C et al (2006) Pied flycatchers Ficedula hypoleuca travelling from Africa to breed in Europe: differential effects of winter and migration conditions on breeding date. Ardea 94:511–525
Bourn D, Wint W (1994) Livestock, land use and agricultural intensification in sub-Saharan Africa. Pastoral Development Network Paper 37a. Overseas Development Institute (ODI), London
Bozdogan H (1987) Model selection and Akaike’s information criterion (AIC): the general theory and its analytical extensions. Psychometrika 52:345–370. doi:10.1007/BF02294361
Brown DR, Long JA (2007) What is a winter floater? Causes, consequences, and implications for habitat selection. Condor 109:548–565. doi:10.1650/8351.1
Brown DR, Stouffer PC, Strong CM (2000) Movement and territoriality of wintering hermit thrushes in Southeastern Louisiana. Wilson Bull 112:347–353. doi:10.1676/0043-5643(2000)112[0347:MATOWH]2.0.CO;2
Catry P, Campos A, Almada V, Cresswell W (2004) Winter segregation of migrant European robins Erithacus rubecula in relation to sex, age and size. J Avian Biol 35:204–209. doi:10.1111/j.0908-8857.2004.03266.x
Cowley E, Siriwardena GM (2005) Long-term variation in survival rates of sand martins Riparia riparia: dependence on breeding and wintering ground weather, age and sex, and their population consequences. Bird Study 52:237–251. doi:10.1080/00063650509461397
Crawley MJ (2007) The R book. Wiley, Chichester
Cresswell W (2014) Migratory connectivity of Palaearctic–African migratory birds and their responses to environmental change: the serial residency hypothesis. Ibis 156:493–510. doi:10.1111/ibi.12168
Cresswell W, Boyd M, Stevens M (2009) Movements of Palearctic and Afrotropical bird species during the dry season (November–February) within Nigeria. In: Harebottle DM, Craig AJFK, Anderson MD, Rakotomanana, Muchai H (eds) Proceedings of the 12th Pan African ornithological congress, 2008. 2008 edn. Animal Demography Unit, Cape Town, South Africa, pp 8–28
Cuadrado M (1992) Year to year recurrence and site-fidelity of blackcaps Sylvia atricapilla and robins Erithacus rubecula in a Mediterranean wintering area. Ringing Migr 13:36–42. doi:10.1080/03078698.1992.9674013
Dejaifve PA (1994) Ecology and behavior of a Palearctic migrant in Africa. The wintering of the whinchat Saxicola rubetra in Zaïre and its winter distribution in Africa. Rev Ecol Terre Vie 49:35–52
Desante DF, Burton KM, Saracco JF, Walker BL (1995) Productivity indices and survival rate estimates from MAPS, a continent-wide programme of constant-effort mist-netting in North America. J Appl Stat 22:935–948. doi:10.1080/02664769524720
Ergon T, Gardner B (2013) Separating mortality and emigration: modelling space use, dispersal and survival with robust-design spatial capture–recapture data. Methods Ecol Evol. doi:10.1111/2041-210X.12133
ERSI (2012) ArcGIS desktop: release 10, vol 2. Environmental Systems Research Institute, Redlands
Förschler MI, del Val E, Bairlein F (2010) Extraordinary high natal philopatry in a migratory passerine. J Ornithol 151:745–748. doi:10.1007/s10336-010-0495-y
Fox J, Weisberg HS (2010) An R companion to applied regression. Sage, California
Gilroy JJ, Virzi T, Boulton RL, Lockwood JL (2012) A new approach to the “apparent survival” problem: estimating true survival rates from mark–recapture studies. Ecology 93:1509–1516. doi:10.1890/12-0124.1
Hansteen TL, Andreassen HP, Ims RA (1997) Effects of spatiotemporal scale on autocorrelation and home range estimators. J Wildl Manag 61:280–290. doi:10.2307/3802583
Henderson IG, Fuller RJ, Conway GJ, Gough SJ (2004) Evidence for declines in populations of grassland-associated birds in marginal upland areas of Britain. Bird Study 51:12–19. doi:10.1080/00063650409461327
Henderson I, Calladine J, Massiminoa D, Taylor JA, Gillings S (2015) Evidence for contrasting causes of population change in two closely related, sympatric breeding species the whinchat Saxicola rubetra and stonechat Saxicola torquata in Britain. Bird Study 61:553–565. doi:10.1080/00063657.2014.962482
Herremans M (1997) Habitat segregation of male and female red-backed shrikes Lanius collurio and lesser grey shrikes Lanius minor in the Kalahari Basin, Botswana. J Avian Biol 28:240–248. doi:10.2307/3676975
Herremans M, Herremans-Tonnoeyr D, Borello WD (1995) Non-breeding site-fidelity of red-backed shrikes Lanius collurio in Botswana. Ostrich 66:145–147
Holmes RT, Sherry TW (1992) Site fidelity of migratory warblers in temperate breeding and Neotropical wintering areas: implications for population dynamics, habitat selection, and conservation. In: Hagan I JM, Johnston DW (eds) Ecology and conservation of Neotropical migrant landbirds. Smithsonian Institution Press, Washington DC, pp 563–575
Holmes RT, Sherry TW, Reitsma L (1989) Population structure, territoriality and overwinter survival of two migrant warbler species in Jamaica. Condor 91:545–561. doi:10.2307/1368105
Hulme MF, Cresswell W (2012) Density and behaviour of whinchats Saxicola rubetra on African farmland suggest that winter habitat conditions do not limit European breeding populations. Ibis 154:680–692. doi:10.1111/j.1474-919X.2012.01258.x
Jenni L, Winkler R (2004) Moult and ageing of European passerines. Academic, London
Jones PJ (1995) Migration strategies of Palearctic passerines in Africa. Isr J Zool 41:393–406. doi:10.1080/00212210.1995.10688809
Karr JR, Nichols JD, Klimkiewicz MK, Brawn JD (1990) Survival rates of birds of tropical and temperate forests: will the dogma survive? Am Nat 136:277–291. doi:10.1086/285098
Kelsey MG (1989) A comparison of the song and territorial behaviour of a long-distance migrant, the marsh warbler Acrocephalus palustris, in summer and winter. Ibis 131:403–414. doi:10.1111/j.1474-919X.1989.tb02788.x
King JMB, Hutchinson JMC (2001) Site fidelity and recurrence of some migrant bird species in The Gambia. Ringing Migr 20:292–302. doi:10.1080/03078698.2001.9674255
Koronkiewicz TJ, Sogge MK, Van Riper C III, Paxton EH (2006) Territoriality, site fidelity, and survivorship of willow flycatchers wintering in Costa Rica. Condor 108:558–570. doi:10.1650/0010-5422(2006)108[558:TSFASO]2.0.CO;2
Kricher JC, Davis WE Jr (1986) Returns and winter-site fidelity of North American migrants banded in Belize, Central America. J Field Ornithol 57:48–52. doi:10.2307/4513089
Latta SC, Faaborg J (2001) Winter site fidelity of prairie warblers in the Dominican Republic. Condor 103:455–468. doi:10.1650/0010-5422(2001)103[0455:WSFOPW]2.0.CO;2
Latta SC, Faaborg J (2002) Demographic and population responses of Cape May warblers wintering in multiple habitats. Ecology 83:2502–2515. doi:10.2307/3071810
Lynch JF, Morton ES, Van der Voort ME (1985) Habitat segregation between the sexes of wintering hooded warblers (Wilsonia citrina). Auk 102:714–721
Marra PP (2000) The role of behavioral dominance in structuring patterns of habitat occupancy in a migrant bird during the nonbreeding season. Behav Ecol 11:299–308. doi:10.1093/beheco/11.3.299
Marra PP, Holmes RT (2001) Consequences of dominance-mediated habitat segregation in American redstarts during the nonbreeding season. Auk 118:92–104. doi:10.1642/0004-8038(2001)118[0092:CODMHS]2.0.CO;2
Marshall MR, Wilson RR, Cooper RJ, Bonney R, Pashley DN, Cooper RJ, Niles L (2000) Estimating survival of Neotropical–Nearctic migratory birds: are they dead or just dispersed. In: Bonney R, Pashley DN, Cooper RJ, Niles L (eds) US Forest Service General Technical Report RMRS-P-16. US Department of Agriculture Forest Service, Ogden
Marshall MR, Diefenbach DR, Wood LA, Cooper RJ (2004) Annual survival estimation of migratory songbirds confounded by incomplete breeding site-fidelity: study designs that may help. Anim Biodivers Conserv 27:59–72
McKinnon EA, Fraser KC, Stutchbury BJM (2013) New discoveries in landbird migration using geolocators, and a flight plan for the future. Auk 130:211–222. doi:10.1525/auk.2013.12226
McNeil R (1982) Winter resident repeats and returns of austral and boreal migrant birds banded in Venezuela. J Field Ornithol 53:125–132
Middleton HA, Morrissey CA, Green DJ (2006) Breeding territory fidelity in a partial migrant, the American dipper Cinclus mexicanus. J Avian Biol 37:169–178. doi:10.1111/j.2006.0908-8857.03514.x
Møller AP, Szép T (2005) Rapid evolutionary change in a secondary sexual character linked to climatic change. J Evol Biol 18:481–495. doi:10.1111/j.1420-9101.2004.00807.x
Morton ES (1990) Habitat segregation by sex in the hooded warbler: experiments on proximate causation and discussion of its evolution. Am Nat 135:319–333. doi:10.1086/285048
Morton ES, Lynch JF, Young K, Mehlhop P (1987) Do male hooded warblers exclude females from nonbreeding territories in tropical forest? Auk 104:133–135
Müller M, Spaar R, Schifferli L, Jenni L (2005) Effects of changes in farming of subalpine meadows on a grassland bird, the whinchat (Saxicola rubetra). J Ornithol 146:14–23. doi:10.1007/s10336-004-0059-0
Newton I (2004) Population limitation in migrants. Ibis 146:197–226. doi:10.1111/j.1474-919X.2004.00293.x
Newton I (2006) Can conditions experienced during migration limit the population levels of birds? J Ornithol 147:146–166. doi:10.1007/s10336-006-0058-4
Newton I (2010a) Bird migration. Collins, London
Newton I (2010b) The migration ecology of birds. Academic, London
Nisbet ICT, Medway L (1972) Dispersion, population ecology and migration of eastern great reed warblers Acrocephalus orientalis wintering in Malaysia. Ibis 114:451–494. doi:10.1111/j.1474-919X.1972.tb00850.x
Norris DR, Marra PP, Kyser TK, Sherry TW, Ratcliffe LM (2004) Tropical winter habitat limits reproductive success on the temperate breeding grounds in a migratory bird. Proc R Soc Lond B 271:59–64. doi:10.1098/rspb.2003.2569
Ornat AL, Greenberg R (1990) Sexual segregation by habitat in migratory warblers in Quintana Roo, Mexico. Auk 107:539–543
Paradis E, Baillie SR, Sutherland WJ, Gregory RD (1998) Patterns of natal and breeding dispersal in birds. J Anim Ecol 67:518–536. doi:10.1046/j.1365-2656.1998.00215.x
Payevsky V, Vysotsky V, Yefremov V, Markovets M, Morozov Y, Shapoval A (1997) Sex-specific survival rates in birds. Zh Obshch Biol 58:5–20
Pulido F (2007) Phenotypic changes in spring arrival: evolution, phenotypic plasticity, effects of weather and condition. Clim Res 35:5–23. doi:10.3354/cr00711
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Rappole JH, Ramos MA, Winker K (1989) Wintering wood thrush movements and mortality in southern Veracruz. Auk 106:402–410. doi:10.1676/13-187.1
Rappole JH, King DI, Diez J (2003) Winter vs. breeding-habitat limitation for an endangered avian migrant. Ecol Appl 13:735–742. doi:10.1890/1051-0761(2003)013[0735:WVBLFA]2.0.CO;2
Reudink MW, Marra PP, Kyser TK, Boag PT, Langin KM, Ratcliffe LM (2009) Non-breeding season events influence sexual selection in a long-distance migratory bird. Proc R Soc Lond B 276:1619–1626. doi:10.1098/rspb.2008.1452
Robinson RA, Balmer DE, Marchant JH (2008) Survival rates of hirundines in relation to British and African rainfall. Ringing Migr 24:1–6. doi:10.1080/03078698.2008.9674375
Salewski V, Bairlein F, Leisler B (2000) Recurrence of some Palaearctic migrant passerine species in West Africa. Ringing Migr 20:29–30. doi:10.1080/03078698.2000.9674224
Salewski V, Bairlein F, Leisler B (2002) Different wintering strategies of two Palearctic migrants in West Africa—a consequence of foraging strategies? Ibis 144:85–93. doi:10.1046/j.0019-1019.2001.00007.x
Sanderson FJ, Donald PF, Pain DJ, Burfield IJ, Van Bommel FPJ (2006) Long-term population declines in Afro-Palearctic migrant birds. Biol Conserv 131:93–105. doi:10.1016/j.biocon.2006.02.008
Sauvage A, Rumsey S, Rodwell S (1998) Recurrence of Palaearctic birds in the lower Senegal river valley. Malimbus 20:33–53
Schaub M, Royle JA (2013) Estimating true instead of apparent survival using spatial Cormack–Jolly–Seber models. Methods Ecol Evol. doi:10.1111/2041-1210X.12134
Schmidt K, Hantge E (1954) Studien an einer farbig beringten Population des Braunkehlchens (Saxicola rubetra). J Ornithol 95:130–173. doi:10.1007/BF01951433
Sherry TW, Holmes RT (1996) Winter habitat quality, population limitation, and conservation of Neotropical–Nearctic migrant birds. Ecology 77:36–48. doi:10.2307/2265652
Shitikov D, Fedotova S, Gagieva V, Fedchuk D, Dubkova E, Vaytina T (2012) Breeding-site fidelity and dispersal in isolated populations of three migratory passerines. Ornis Fenn 89:53–62
Shitikov DA, Morozova MM, Yurchenko YA, Anashina AD (2013) Apparent survival rates of two Sylvia warbler species in northwestern Russia. Ringing Migr 28:16–20. doi:10.1080/03078698.2013.810858
Shitikov DA, Vaytina TM, Gagieva VA, Fedchuk DV (2015) Breeding success affects site fidelity in a whinchat Saxicola rubetra population in abandoned fields. Bird Study 62:96–105. doi:10.1080/00063657.2014.988120
Siriwardena GM, Baillie SR, Wilson JD (1998) Variation in the survival rates of some British passerines with respect to their population trends on farmland. Bird Study 45:276–292. doi:10.1080/00063659809461099
Skilleter M (1995) Winter site fidelity of redstart Phoenicurus phoenicurus in N. Nigeria. Malimbus 17:101–102
Steifetten Ø, Dale S (2006) Viability of an endangered population of ortolan buntings: the effect of a skewed operational sex ratio. Biol Conserv 132:88–97. doi:10.1016/j.biocon.2006.03.016
Studds CE, Kyser TK, Marra PP (2008) Natal dispersal driven by environmental conditions interacting across the annual cycle of a migratory songbird. Proc Natl Acad Sci 105:2929–2933. doi:10.1073/pnas.0710732105
Stutchbury BJ (1994) Competition for winter territories in a Neotropical migrant: the role of age, sex and color. Auk 111:63–69
Thaxter CB, Joys AC, Gregory RD, Baillie SR, Noble DG (2010) Hypotheses to explain patterns of population change among breeding bird species in England. Biol Conserv 143:2006–2019. doi:10.1016/j.biocon.2010.05.004
Tryjanowski P, Goławski A, Kuźniak S, Mokwa T, Antczak M (2007) Disperse or stay? Exceptionally high breeding-site infidelity in the red-backed shrike Lanius collurio. Ardea 95:316–320. doi:10.5253/078.095.0214
Vickery JA, Ewing SR, Smith KW, Pain DJ, Bairlein F, Škorpilová J, Gregory RD (2014) The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis 156:1–22. doi:10.1111/ibi.12118
Warkentin IG, Hernandez D (1996) The conservation implications of site fidelity: a case study involving Nearctic–Neotropical migrant songbirds wintering in a Costa Rican mangrove. Biol Conserv 77:143–150. doi:10.1016/0006-3207(95)00146-8
Winker K (1998) The concept of floater. Ornitol Neotrop 9:111–119
Wunderle JM Jr (1995) Population characteristics of black-throated blue warblers wintering in three sites on Puerto Rico. Auk 112:931–946
Acknowledgments
This research was funded by the Leventis Conservation Foundation. We thank the staff and associates of the A.P. Leventis Ornithological Research Institute who made this study possible and who assisted with data collection, and the referees who helped to improve this manuscript. This is paper number 92 from the A.P. Leventis Ornithological Research Institute. This study complies with the current ethics regulations of Nigeria and the A.P. Leventis Ornithological Research Institute.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by N. Chernetsov.
Rights and permissions
About this article

Cite this article
Blackburn, E., Cresswell, W. High winter site fidelity in a long-distance migrant: implications for wintering ecology and survival estimates. J Ornithol 157, 93–108 (2016). https://doi.org/10.1007/s10336-015-1252-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-015-1252-z