
Abstract
Studies of food preferences in captive primates have so far mainly been restricted to frugivorous species. It was therefore the aim of the present study to assess the occurrence of spontaneous food preferences in a mainly folivorous primate, the captive Southern brown howler monkey, and to analyze whether these preferences correlate with nutrient composition. Using a two-alternative choice test, we presented ten male and five female adult Alouatta guariba clamitans with all possible binary combinations of ten types of food that are part of their diet in captivity and recorded their choice behavior. We found the howler monkeys to display the following rank order of preference: banana > mango > watermelon > papaya > beetroot > apple > pear > orange > cucumber > tomato. This preference ranking significantly and positively correlated with the total carbohydrate content and with the sucrose content of the food items. We also found significant positive correlations between the food preference ranking and the content of the minerals copper and magnesium. Male and female howler monkeys did not differ significantly in their food preference rankings. These results suggest this howler monkeys under human care are not opportunistic, but selective feeders with regard to maximizing their net gain of energy as only the content of carbohydrates, but not the contents of total energy, proteins, or lipids significantly correlated with the displayed food preferences. Thus, the food preferences of this primate are similar to those reported in several species of frugivorous primates tested with cultivated fruits and vegetables.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Most primates are known to feed on a wide variety of foods of plant origin (Hohmann 2009). However, numerous field studies have shown that primates do not feed at random on available plants but are highly selective in their food choices and aim at maximizing their intake of metabolic energy and critical nutrients (Chivers 1998; Raubenheimer et al. 2015). The food choices of primates and other mammals are thought to be steered by two main determinants: the nutritional and/or toxic content of the consumed plant part (Barton and Whiten 1994; Windley et al. 2022) and its relative temporal and spatial distribution (Leighton 1993; Trapanese et al. 2019). Plant secondary metabolites such as alkaloids, phenolics, or tannins, which inhibit the digestion of polysaccharides and proteins or are even toxic, are produced by plants to deter herbivores (Iason et al. 2012). Accordingly, high concentrations of such compounds have been reported to correlate negatively with food choices in primates (Belovsky and Schmitz 1994; Glander 1982; Windley et al. 2022).
In contrast, food choices of primates should be expected to correlate positively with high concentrations of nutritionally valuable compounds which provide metabolic energy such as carbohydrates, proteins, and lipids, or of certain critical micronutrients such as minerals or vitamins (Lambert and Rothman 2015; Simpson et al. 2004). However, only a few studies in nonhuman primates so far have reported such positive correlations between food choice and the contents of specific nutrients (e.g., Conklin-Brittain et al. 1998; Felton et al. 2009; Milton 1998; Raubenheimer et al. 2015; Rothman et al. 2011). This may be due to the fact that plants usually comprise a complex mixture of both aversive non-palatable or toxic compounds and attractive beneficial nutrients. Thus, it is difficult to decide whether plants consumed by primates may be preferred because of their content of certain attractive nutrients or because of their low concentration of aversive secondary metabolites (Glander 1982; Goyal et al. 2012).
One way to circumvent this problem and to gain first insight into which nutrients may positively affect food selection is to present animals with plants that contain only low and presumably negligible concentrations of secondary metabolites and to assess whether their food choices correlate positively with certain nutrients. This approach may be helpful to increase our understanding of optimal foraging which so far has been applied with only limited success to herbivores (Stephens et al. 2008) and may aid in the formulation of appropriate diets for primates ex-situ (National Research Council 2003). Cultivated fruits and vegetables comply with this notion as they have been selectively bred to contain only low amounts of plant secondary metabolites (Paliyath et al. 2008) and their nutrient content is well documented (Food Standards Agency 2002). Previous studies using the approach to allow captive primates to choose between cultivated fruits and vegetables found that some primate species such as white-handed gibbons (Hylobates lar), ring-tailed lemurs (Lemur catta), and pigtail macaques (Macaca nemestrina) showed significant positive correlations between their food preferences and carbohydrate content (Hansell et al. 2020; Jildmalm et al. 2008; Laska 2001). They are therefore considered as selective feeders with regard to their preferred source of metabolic energy. Other primate species such as squirrel monkeys (Saimiri sciureus), spider monkeys (Ateles geoffroyi), and white-faced sakis (Pithecia pithecia), in contrast, showed significant positive correlations between their food preferences and total energy content, irrespective of whether the metabolic energy originated from carbohydrates, proteins, or lipids (Laska et al. 2000; Laska 2001; Martins et al. 2023). Accordingly, they are considered as opportunistic feeders with regard to their preferred source of metabolic energy. Additionally, most of the above-mentioned studies reported significant positive correlations between a species’ food preferences and the content of certain minerals. Such correlations may indicate that the diet fed to captive primates may lack one or the other critical micronutrient and are therefore useful to prevent the development of health problems due to malnutrition (Schwitzer and Kaumanns 2003).
All primate species tested so far for their food preferences using cultivated fruits and vegetables are considered as frugivores. As far as we can tell, no study to date adopted this approach with a folivorous primate species. The present study therefore aims to fill this gap in our knowledge.
The Southern brown howler monkey (Alouatta guariba clamitans) is a folivorous platyrrhine whose diet comprises more than 400 species of plants (Chaves and Bicca-Marques 2013). Depending on study site, the proportion of leaves in the diet of the brown howler monkey has been found to range between 62% (Agostini et al. 2010) and 81% (Martins 2008), with two meta-analyses reporting mean values across studies of 63% (Chaves and Bicca-Marques 2013) and 64% leaves (Dias and Rangel-Negrin 2015), respectively. However, brown howler monkeys also include between 8% (Martins 2008) and 24% (Agostini et al. 2010) fruits into their diet, depending on season (Santos et al. 2013). Although leaves, in contrast to fruits, are considered as a low-quality diet, it is commonly agreed that brown howler monkeys obtain the bulk of their metabolizable energy from leaves and not from fruits (Garber et al. 2015). As brown howler monkeys, similar to other arboreal platyrrhines, have only rarely been observed to drink from open water sources, it is likely that they mainly meet their water requirements via their food (Dias and Rangel-Negrin 2015). Reports that drinking in brown howler monkeys is negatively correlated with fruit consumption and precipitation suggest that they may at least partly meet their water requirements during the dry season by exploiting the water contained in the pulp of fleshy fruits (Miranda et al. 2005; Steinmetz 2001). Although not very common, brown howler monkeys have also been reported to engage in soil eating and in using mineral licks (Ferrari et al. 2008; Pebsworth et al. 2019). Geophagy is thought to serve different functions including supplementation of minerals, detoxification of plant secondary metabolites, and treatment of gastrointestinal ailments (Krishnamani and Mahaney 2000).
It was therefore the aim of the present study to assess food preferences in a group of captive Southern brown howler monkeys for a variety of cultivated fruits and vegetables, and to analyze whether these preferences correlate with the abundance of macronutrients such as carbohydrates, proteins, lipids, dietary fiber and water, as well as micronutrients such as minerals and vitamins. We hypothesized that captive Southern brown howler monkeys (1) should display clear preferences for specific types of food and that these preferences are based on their nutrient composition, (2) should prefer food items high in total energy content, irrespective of the source of metabolic energy, and (3) should show a negative correlation between their food preferences and water content of the food items.
Materials and methods
Animals
Ten male and five female adult Southern brown howler monkeys (Alouatta guariba clamitans) participated in this study. They were maintained in the scientific breeding center of the Biological Research Center of Indaial (CEPESBI), located in the municipality of Indaial, Santa Catarina, Brazil. The animals were housed in enclosures measuring 3 × 5 × 2.6 m (width × length × height), with covered containment measuring 3 × 1.5 × 2.6 m. The animals were exposed to the ambient climate, with sliding doors that allowed for social interaction. The enclosures were provided with enrichment such as logs, hoses, ropes, and heated boxes. The daily tests described below took place three hours before feeding. The animals were fed six meals per day, of which three were composed of various fruits and three of freshly cut branches bearing leaves. The animals did not have ad libitum access to water, but rather met their water requirements from juicy fruits. The animals were not on a food deprivation schedule and their participation in the food preference tests described below was completely voluntary.
Procedure
We used a two-alternative choice test (Laska et al. 2000) to assess food preferences in the howler monkeys. Twice a day, before feeding, we presented the animals with pairs of food items on a cutting board, spaced 15 cm apart, and recorded their choice behavior. As soon as an animal had decided for one of the two simultaneously presented food items by taking it, we removed the cutting board to prevent the animal from taking the other food item (Fig. 1). Care was taken to present food items of approximately equal size (3 × 2 × 1 cm) to minimize the risk that apparent differences in the size of the food items affected the animals’ food choices. We tested the howler monkeys singly in order to avoid the animal´s behavior being affected by competition or distraction.

Animals that participated in the study, choosing between the offered food items
The howler monkeys were presented with all 45 possible binary combinations of ten types of food for a total of ten times per animal. In each session an animal voluntarily placed itself on a horizontal perch that was attached in parallel to the mesh that separated the animal from the experimenter. The mesh was wide enough to allow the howler monkeys to fit their hand through and grab hold of a food item. Each animal performed one session per day with two trials per session.
The ten different types of food were banana (Musa paradisiaca), apple (Malus pumila), papaya (Carica papaya), pear (Pyrus sp.), beetroot (Beta vulgaris), tomato (Solanum lycopersicum), cucumber (Cucumis sativus), mango (Mangifera indica), watermelon (Citrullus lanatus) and orange (Citrus x sinensis).
The rationale for choosing these types of food was that: (a) all of them were part of the diet of the howler monkeys under human care and, therefore, familiar to the animals and readily consumed when presented singly, (b) information on the macro- and micronutrient contents of these fruits was available from the literature (Food Standards Agency 2002), allowing us to assess possible correlations between food preferences and nutrient content, and (c) they differed markedly from each other in their content of certain macro- or micronutrients.
The contents of total energy, carbohydrates, proteins, lipids, dietary fiber, sodium, calcium, folate (= vitamin B9), ascorbic acid (= vitamin C), and carotene, for example, differed by up to a factor of 10, 15, 6, 3, 22, 66, 12, 150, 12, and 45, respectively, between the types of food used here. To minimize the inevitable variation in the nutrient content of a given type of food, we took care to always present pieces of a given type of food using the same variety or cultivar and with the same degree of ripeness.
Ethical note
The experiments reported here complied with the American Society of Primatologists' Principles for the Ethical Treatment of Primates, and with current Brazilian, Mexican, and Swedish laws. They were approved by the Committee on Ethics in the Use of Animals/CEUA – FURB (nº 011/18).
Data analysis
We recorded a total of 6750 choices (45 binary combinations × 10 presentations per animal × 15 animals), and we established food preference rankings using the following criteria (Hansell et al. 2020):
Criterion 1 (individual level): We built the sum total of choices for each of the ten types of food across all binary combinations for each individual animal. The theoretical maximum score for any type of food with this criterion was 90 (9 combinations × 10 presentations per animal × 1 animal).
Criterion 2 (group level): This criterion adopts the same procedure of building the sum total of choices as for criterion 1, although, here, we collapsed the data for all 15 animals. Thus, the theoretical maximum score for any type of food with this criterion was 1350 (9 combinations × 10 presentations per animal × 15 animals).
We performed two-tailed binomial tests using the sum total of choices for each member of a given binary combination of food items to assess significant preferences both at the individual level and at the group level (p < 0.05). We evaluated correlations between the food preference rankings and the contents of nutrients by calculating Spearman rank-order correlation coefficients rs, which we tested for significance by computing z-scores. We used the same test to assess whether the food preference rankings of the 15 howler monkeys correlated with each other, and whether the food preference rankings of the males and the females correlated with each other. We performed all statistical tests with Bonferroni corrections for multiple testing.
Results
Food preferences
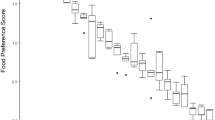
With 35 of the 45 binary combinations of food items that we presented to the animals, the howler monkeys as a group displayed a statistically significant preference for one of the options (two-tailed binomial test, p < 0.05) (Table 1). Banana was clearly the most attractive food item tested and, accordingly, was significantly preferred over all nine other food items (p < 0.05). The high attractiveness of banana is further illustrated by the fact that 81.1% of all possible choices were in favor of this food item (Table 2). Mango and watermelon were significantly preferred over six of the other food items and 68.2 and 67.6%, respectively, of all possible choices were in favor of these two fruits. Tomato and cucumber, on the other hand, were the least attractive food items and were never significantly preferred over any of the other nine food items. Accordingly, only 22.4 and 25.4%, respectively, of all possible choices were in favor of these two food items.
Rankings derived from the food preferences
As a group, the howler monkeys displayed the following rank order of preference: banana > mango > watermelon > papaya > beetroot > apple > pear > orange > cucumber > tomato (Table 2).
All 15 individual howler monkeys displayed similar rankings of preference for the ten food items. The food preference rankings were also similar between the ten males and the five females and, accordingly, correlated significantly with each other (Spearman, rs = 0.67, p < 0.05).
Food preference rankings and nutritional content
The food preference rankings displayed by the Southern brown howler monkeys correlated significantly (p < 0.05) with total carbohydrate content and with sucrose content of the food items (Table 3). Additionally, we found significant positive correlations between the food preference ranking and the nutritional content of the food items with the mineral’s magnesium and copper. Further, we found non-significant trends (0.05 < p < 0.1) for a positive correlation between the food preference ranking and the content of riboflavin (= vitamin B2) and zinc, and for a negative correlation with water content and sodium content of the food items.
Discussion
The results of the present study demonstrate that captive Southern brown howler monkeys display marked food preferences in a two-alternative choice test using cultivated fruits and vegetables. Further, the results show that these preferences significantly correlated positively with carbohydrate and sucrose content of the food items used as well as with the contents of the minerals copper and magnesium.
Our finding that the Southern brown howler monkeys clearly preferred certain types of food over others and that these preferences correlated with their content of certain nutrients is in line with our first hypothesis. According to the optimal foraging theory, natural selection should favor individuals that succeed in maximizing their intake of energy and critical nutrients and this should be reflected in their food preferences (Stephens et al. 2008). Thus, it should not be surprising that the brown howler monkeys displayed clear preferences in their food choices, similar to other nonhuman primate species tested previously and adopting the same approach as used in the present study (Hansell et al. 2020; Jildmalm et al. 2008; Laska et al. 2000; Laska 2001; Martins et al. 2023). However, our finding of nutrient-based food preferences in captive primates is not trivial if we consider that several factors driving food selection under natural conditions such as seasonal variation in the availability, spatial distribution, and quality of foods (Hemingway and Bynum 2005; Santos et al. 2013) are much weaker, if present at all, in animals under human care. Therefore, our finding supports the notion that assessing food preferences using cultivated fruits and vegetables may give us first insight into which nutrients affect food choices positively and whether they may reflect evolutionary adaptations to the dietary specialization of a species. Future studies comparing food preferences between primates under human care and their conspecifics in the wild should allow us to better understand the roles of plant secondary compounds whose abundance is largely reduced in cultivated fruits and vegetables for food selection. Comparative studies including captive and free-ranging animals of a given primate species should also allow us to gain insight into which factors may determine whether an animal´s food choices are opportunistic or selective in order to obtain the amount of metabolic energy it needs and in how far findings from studies with captive animals are consistent with findings from studies with animals in the wild.
Our finding that male and female brown howler monkeys did not differ significantly in their food preference rankings (Table 2) is remarkable considering that the nutrient requirements of male and female mammals may differ considerably, depending on hormonal and reproductive status (Key and Ross 1999). However, none of the female howler monkeys of the present study were pregnant or lactating during our data collection which might explain this lack of significant sex differences. It should be interesting to assess whether the food preferences of captive female brown howler monkeys for cultivated fruits and vegetables may change during the period of pregnancy and lactation—as has been reported for the diet of howler monkeys in the wild (Serio-Silva et al. 1999).
Macronutrients
Our finding that the Southern brown howler monkeys displayed a significant positive correlation between their food preference ranking and carbohydrate (p = 0.03) and sucrose content (p = 0.011) of the food items, but not with total energy content (p = 0.16), is in contrast to our second hypothesis. This hypothesis was based on the notion that leaves are a low-quality diet which should favor opportunistic food selection with regard to maximizing the intake of metabolic energy, irrespective of whether this energy stems from carbohydrates, proteins, or lipids. Some studies even suggest an important role for both proteins (Ganzhorn et al. 2017) and lipids (Righini et al. 2017) for the food selection in folivorous primates, including members of the genus Alouatta. However, our results suggest that captive brown howler monkeys are selective rather than opportunistic feeders with regard to carbohydrates as their main source of metabolic energy—at least with the food items employed in the present study.
This is interesting as three other platyrrhine primate species, squirrel monkeys, spider monkeys, and white-faces sakis have all been reported to show significant positive correlations between their food preferences and total energy content, but not with carbohydrate content (Laska et al. 2000; Laska 2001; Martins et al. 2023). Other primate species such as white-handed gibbons, ring-tailed lemurs, and pigtail macaques, in contrast, showed significant positive correlations between their food preferences and carbohydrate content, but not with total energy content, and thus behaved similar to the brown howler monkeys of the present study (Jildmalm et al. 2008; Hansell et al. 2020; Laska 2001). However, all of these above-mentioned species are frugivores rather than folivores. Our findings therefore support the notion that being a selective feeder concerning the source of metabolic energy is not only found in frugivorous primates, but also in a folivorous primate species. Future studies should assess if other folivorous primate species may adopt the strategy of being an opportunistic rather than a selective feeder with regard to meeting their energy requirements.
Further, it would be interesting to assess whether the marked preferences of the brown howler monkeys for food items containing high amounts of soluble carbohydrates are based on their taste property of sweetness or on their physiological property of being an easily metabolizable source of energy. Whereas a variety of nonhuman primate species has been studied with regard to their sweet-taste sensitivity for food-associated carbohydrates (e.g., Laska et al. 1996; Norlén et al. 2019; Wielbass et al. 2015), corresponding data for any member of the genus Alouatta, including the brown howler monkey, are so far lacking.
Our finding that the brown howler monkeys showed a negative correlation between their food preferences and water content of the food items which just fell short of statistical significance (p = 0.07) is in line with our third hypothesis. It is interesting to note that the animals of the present study did not have ad libitum access to water and thus had to meet their water requirements via their food. Nevertheless, our results suggest that the water content of the fruits and vegetables employed here was high enough so that the howler monkeys did not have to make trade-offs between the needs to meet their water and their energy requirements, respectively, but could afford to focus on the latter. This notion is supported by the fact that the kidneys of brown howler monkeys have a high capacity to reabsorb water (Peçanha et al. 2023). Previous studies showed that spider monkeys (Laska et al. 2000), squirrel monkeys (Laska 2001), and white-faced sakis (Martins et al. 2023), all displayed a significant negative correlation with water content whereas pigtail macaques, white-handed gibbons, and ring-tailed lemurs (Hansell et al. 2020; Jildmalm et al. 2008; Laska 2001) did not. Here, too, future studies should assess whether other folivorous primate species may differ from the brown howler monkeys in their food preferences as a function of water content.
Micronutrients
The brown howler monkeys of the present study displayed significant positive correlations between their food preference ranking and the nutritional content of the food items with the minerals copper (p = 0.011) and magnesium (p = 0.03). Additionally, they showed non-significant trends for a positive correlation between the food preference ranking and the content of riboflavin (p = 0.09) and zinc (p = 0.08), and for a negative correlation with sodium content (p = 0.051) of the food items.
Numerous studies found that animals are able to adapt their feeding patterns by selecting foods containing high amounts of certain micronutrients such as minerals or vitamins to counteract the shortage of a given micronutrient in their current diet (Simpson et al. 2004). Minerals such as sodium, copper, zinc, and iron are usually only found at low concentrations in the leaves and fruits consumed by primates and may thus be limiting factors in primate diets (Rothman et al. 2014). To prevent mineral deficiencies, a wide variety of primates in the wild have been reported to perform geophagy and to use mineral licks (Krishnamani and Mahaney 2000) and both behaviors have also been reported in free-ranging brown howler monkeys (Ferrari et al. 2008; Pebsworth et al. 2019).
Based on the present data, we cannot decide whether our findings may indicate a lack of the above-mentioned minerals in the diet provided to our captive animals or whether the contents of these micronutrients simply correlated with the contents of another nutrient. The latter case would suggest that the corresponding preference for the mineral may just be a byproduct of a preference for this other nutrient. Indeed, the contents of copper and of riboflavin, respectively, in the food items used in the present study correlated significantly with the content of total carbohydrates (rs = 0.80, p = 0.005 with copper; rs = 0.85, p = 0.002 with riboflavin). However, no significant correlations were found between total carbohydrate content and the contents of magnesium, zinc, and sodium (p > 0.10 with all three correlations) in the food items used here.
Nevertheless, it is interesting to note that two other platyrrhine primate’s species, squirrel monkeys and spider monkeys, also displayed significant positive correlations between their food preferences and copper content of the food items used in corresponding tests (Laska 2001; Laska et al. 2000). Copper deficiency in nonhuman primates has been associated with osteoporosis, cardiovascular disease, and poor immune response (Lopez de Romana et al. 2011), and zinc deficiency is known to elicit alopecia and dermal lesions (Crissey and Pribyl 2000). Similarly, an insufficient supply of magnesium has been associated with aortic lipid deposition and thus with cardiovascular disease in platyrrhines (Vitale et al. 1963). The diet of platyrrhine primates under human care should therefore ensure a sufficient supply of these trace minerals. Food preference tests using cultivated fruits and vegetables with known amounts of critical micronutrients provide a readily applicable means to perform such screening.
Folivory, food preferences, and nutrient composition
Leaves are commonly considered as a low-quality diet (Hohmann 2009; Milton 1993). Although leaves may contain higher concentrations of proteins compared to fruits, their contents of soluble and thus easily metabolizable carbohydrates and their total energy content per mass unit is usually much lower compared to fruits (Aristizabal et al. 2017). Accordingly, a leaf-based diet requires morphological and physiological adaptations to properly exploit the nutrients contained in leaves. The dentition of Southern brown howler monkeys shows the typical morphological adaptations for folivory with incisors that are markedly smaller than those of frugivorous primates and high-crested molars instead of the low-relief molars that are characteristic for frugivores (Youlatos et al. 2015). Like all members of the genus Alouatta, brown howler monkeys are hindgut fermenters and thus rely on symbiotic bacteria to digest non-soluble and structural carbohydrates such as cellulose (Garber et al. 2015). As a consequence, howler monkeys have a slow digesta passage compared to frugivores which, interestingly, does not change when they are feeding on fruits (Espinosa-Gomez et al. 2013).
All of these evolutionary adaptations to folivory are in line with the numerous studies reporting that howler monkeys, including the Southern brown howler monkey, rely on leaves as their main source of metabolic energy (Dias and Rangel-Negrin 2015). Rosenberger et al. (2011) suggested that folivory in platyrrhine primates may be a derived trait, with frugivory being the ancestral trait. This might, at least partly, explain why the brown howler monkeys of the present study displayed food preferences that correlated positively with carbohydrate content of the food items used, just as several species of frugivorous primates did in corresponding tests using cultivated fruits and vegetables (Hansell et al. 2020; Jildmalm et al. 2008; Laska 2001). Thus, our finding might represent an example of Liem’s paradox. Liem's paradox refers to the observation that species with seemingly specialized phenotypes can sometimes behave as ecological generalists (Liem 1980). Accordingly, the morphological and physiological adaptations of the brown howler monkeys to efficiently feed on a leaf-based diet does not seem to compromise their ability to also exploit carbohydrates.
We are aware that the cultivated fruits and vegetables presented to the brown howler monkeys in the present study differ markedly in their nutritional composition from the diet of their conspecifics in the wild. The same is true for the diet of most primates kept under human care. Cultivated fruits, for example, have been selectively bred to appeal to human tastes and are therefore higher in soluble carbohydrates and lower in dietary fiber and plant secondary metabolites compared to the non-cultivated fruits that primates co-evolved with and feed on in the wild (Milton 2000). Therefore, we can, of course, not exclude the possibility that the food selection of brown howler monkeys in the wild may be governed by other macro- or micronutrients than found here. Rather, it is quite likely that the contents of plant secondary metabolites found in the non-cultivated plants that the howler monkeys feed on in the wild may have a marked impact on their food selection, probably by eliciting more or less pronounced avoidance responses. In this context, it should be emphasized that food choices are probably only rarely determined by one type of nutrient of feeding deterrent only but by a combination of both. Future studies should therefore consider the ratios between attractive nutrients and aversive plant secondary metabolites for food selection in primates. Nevertheless, our approach to assess food preferences for cultivated fruits and vegetables that are, on purpose, low in usually aversive compounds such as alkaloids, phenolics, or tannins allows us to draw first conclusions as to which nutrients may affect food choices of primates in a positive manner.
In conclusion, we found that captive brown howler monkeys display marked food preferences in a two-alternative choice test using cultivated fruits and vegetables and that these preferences significantly correlated positively with carbohydrate and sucrose content of the food items used.
References
Agostini I, Holzmann I, Di Bitetti MS (2010) Are howler monkey species ecologically equivalent? Trophic niche overlap in syntopic Alouatta guariba clamitans and Alouatta caraya. Am J Primatol 72:173–186. https://doi.org/10.1002/ajp.20775
Aristizabal JF, Rothman JM, Garcia-Feria LM, Serio-Silva JC (2017) Contrasting time-based and weight-based estimates of protein and energy intake of black howler monkeys (Alouatta pigra). Am J Primatol 79:1–8. https://doi.org/10.1002/ajp.22611
Barton RA, Whiten A (1994) Reducing complex diets to simple rules: Food selection by olive baboons. Behav Ecol Sociobiol 35:283–293. https://doi.org/10.1007/BF00170709
Belovsky GE, Schmitz OJ (1994) Plant defenses and optimal foraging by mammalian herbivores. J Mamm 75:816–832. https://doi.org/10.2307/1382464
Chaves OM, Bicca-Marques JC (2013) Dietary flexibility of the brown howler monkey throughout its geographic distribution. Am J Primatol 75:16–20. https://doi.org/10.1002/ajp.22075
Chivers DJ (1998) Measuring food intake in wild animals: Primates. Proc Nutr Soc 57:321–332. https://doi.org/10.1079/PNS19980047
Conklin-Brittain ML, Wrangham R, Hunt KD (1998) Dietary response of chimpanzees and cercopithecines to seasonal variation in fruit abundance: II. Macronutrients Int J Primatol 19:971–998. https://doi.org/10.1023/A:1020370119096
Crissey S, Pribyl L (2000) A review of nutritional deficiencies and toxicities in captive New World primates. Int Zoo Yb 37:355–360. https://doi.org/10.1111/j.1748-1090.2000.tb00741.x
Dias PAD, Rangel-Negrin A (2015) Diets of howler monkeys. In: Kowalewski MM, Garber PA, Cortes-Ortiz L, Urbani B, Youlatos D (eds) Howler monkeys: behavior, ecology, and conservation. Springer, New York, pp 21–56
Espinosa-Gomez F, Gomez-Rosales S, Wallis IR, Canales-Espinosa D, Hernández-Salazar L (2013) Digestive strategies and food choice in mantled howler monkeys Alouatta palliata Mexicana: bases of their dietary flexibility. J Comp Phyiol B 183:1089–1100. https://doi.org/10.1007/s00360-013-0769-9
Felton AM, Felton A, Raubenheimer D, Simpson SJ, Foley WJ, Wood JT, Wallis IR, Lindenmayer DB (2009) Protein content of diets dictates the daily energy intake of a free-ranging primate. Behav Ecol 20:685–690. https://doi.org/10.1093/beheco/arp021
Ferrari SF, Veiga LM, Urbani B (2008) Geophagy in New World monkeys (Platyrrhini): ecological and geographic patterns. Folia Primatol 79:402–415. https://doi.org/10.1159/000141901
Food Standards Agency (2002) McCance and Widdowson’s The composition of foods, 6th edn. Royal Society of Chemistry, Cambridge
Ganzhorn JU, Arrigo-Nelson SJ, Carrai V, Chalise MK, Donati G, Droescher I, Eppley TM, Irwin MT, Koch F, Koenig A, Kowalewski MM, Mowry CB, Patel ER, Pichon C, Ralison J, Reisdorff C, Simmen B, Stalenberg E, Starrs D, Terboven J, Wright PC, Foley WJ (2017) The importance of protein in leaf selection of folivorous primates. Am J Primatol 79:e22550. https://doi.org/10.1002/ajp.22550
Garber PA, Righini N, Kowalewski MM (2015) Evidence of alternative dietary syndromes and nutritional goals in the genus Alouatta. In: Kowalewski MM, Garber PA, Cortes-Ortiz L, Urbani B, Youlatos D (eds) Howler monkeys: behavior, ecology, and conservation. Springer, New York, pp 85–109
Glander KE (1982) The impact of plant secondary compounds on primate feeding behavior. Am J Phys Anthropol 25:1–18
Goyal S, Lambert C, Cluzet S, Mérillon JM, Ramawat KG (2012) Secondary metabolites and plant defense. In: Mérillon JM, Ramawat KG (eds) Plant defense: biological control. Springer, New York, pp 109–138
Hansell M, Åsberg A, Laska M (2020) Food preferences and nutrient composition in zoo-housed ring-tailed lemurs Lemur catta. Physiol Behav 226:113125. https://doi.org/10.1016/j.physbeh.2020.113125
Hemingway CA, Bynum N (2005) The influence of seasonality on primate diet and foraging. In: Brockman DK, van Schaik CP (eds) Seasonality in primates. Cambridge University Press, Cambridge, pp 57–104
Hohmann G (2009) The diet of non-human primates: frugivory, food processing, and food sharing. In: Hublin JJ, Richards MP (eds) The evolution of hominin diets. Springer, New York, pp 1–14
Iason GR, Dicke M, Hartley SE (2012) The ecology of plant secondary metabolites. Cambridge University Press, Cambridge, UK
Jildmalm R, Amundin M, Laska M (2008) Food preferences and nutrient composition in captive white-handed gibbons, Hylobates lar. Int J Primatol 29:1535–1547. https://doi.org/10.1007/s10764-008-9314-1
Key C, Ross C (1999) Sex differences in energy expenditure in non-human primates. Proc R Soc Lond B 266:2479–2485. https://doi.org/10.10998/rspb.1999.0949
Krishnamani R, Mahaney WC (2000) Geophagy among primates: adaptive significance and ecological consequences. Anim Behav 59:899–915. https://doi.org/10.1006/anbe.1999.1376
Lambert JE, Rothman JE (2015) Fallback foods, optimal diets, and nutritional targets: primate responses to varying food availability and quality. Ann Rev Anthropol 44:493–512. https://doi.org/10.1146/annurev-anthro-102313-025928
Laska M (2001) Comparison of food preferences and nutrient composition in captive squirrel monkeys, Saimiri sciureus, and pigtail macaques, Macaca nemestrina. Physiol Behav 73:111–120. https://doi.org/10.1016/s0031-9384(01)00439-5
Laska M, Carrera Sanchez E, Rodriguez Rivera JA, Rodriguez Luna E (1996) Gustatory thresholds for food-associated sugars in the spider monkey (Ateles geoffroyi). Am J Primatol 39:189–193. https://doi.org/10.1002/(SICI)1098-2345(1996)39:3%3c189::AID-AJP4%3e3.0.CO;2-V
Laska M, Hernández-Salazar LT, Rodriguez Luna E (2000) Food preferences and nutrient composition in captive spider monkeys, Ateles geoffroyi. Int J Primatol 21:671–683. https://doi.org/10.1023/A:1005517421510
Leighton M (1993) Modeling dietary selectivity by Bornean orangutans: evidence for integration of multiple criteria in fruit selection. Int J Primatol 14:257–311. https://doi.org/10.1007/BF02192635
Liem KF (1980) Adaptive significance of intra- and interspecific differences in the feeding repertoires of cichlid fishes. Amer Zool 20:295–314. https://doi.org/10.1093/icb/20.1.295
Lopez de Romana D, Olivares M, Uauy R, Araya M (2011) Risks and benefits of copper in light of new insights of copper homeostasis. J Trace Elem Med Biol 25:3–13. https://doi.org/10.1016/j.jtemb.2010.11.004
Martins MM (2008) Fruit diet of Alouatta guariba and Brachyteles arachnoides in Southeastern Brazil: comparison of fruit type, color, and seed size. Primates 49:1–8. https://doi.org/10.1007/s10329-007-0050-5
Martins VA, Magnusson N, Laska M (2023) Go for lipids! Food preferences and nutrient composition in zoo-housed white-faced sakis, Pithecia pithecia. Int J Primatol 44:341–356. https://doi.org/10.1007/s10764-022-00344-5
Milton K (1993) Diet and primate evolution. Sci Amer 269:86–93
Milton K (1998) Physiological ecology of howlers (Alouatta): energetic and digestive considerations and comparison with the Colobinae. Int J Primatol 19:513–547. https://doi.org/10.1023/A:1020364523213
Milton K (2000) Back to basics: why foods of wild primates have relevance for modern human health. Nutrition 16:481–483. https://doi.org/10.1016/s0899-9007(00)00293-8
Miranda JMD, Moro-Rios RF, Bernardi IP, Passos FC (2005) Formas não usuais para a obtenção de água por Alouatta guariba clamitans em ambiente de floresta com Araucária no sul do Brasil. Neotrop Primates 13:21–23. https://doi.org/10.1896/1413-4705.13.2.21
National Research Council (2003) Nutrient requirements of nonhuman primates, 2nd, revised. The National Academies Press, Washington
Norlén E, Sjöström D, Hjelm M, Hård T, Laska M (2019) Taste responsiveness of Western chimpanzees (Pan troglodytes verus) to five food-associated saccharides. Primates 60:29–39. https://doi.org/10.1007/s10329-018-0697-0
Paliyath G, Murr DP, Handa AK, Lurie S (2008) Postharvest biology and technology of fruits, vegetables, and flowers. Wiley, New York
Pebsworth PA, Huffman MA, Lambert JE, Young SL (2019) Geophagy among nonhuman primates: a systematic review of current knowledge and suggestions for future directions. Am J Phys Anthropol 168:164–194. https://doi.org/10.1002/ajpa.23724
Peçanha SV, Estruc TM, Carvalho RBJ, Santos-Sousa CA, Souza-Junior P, Abidu-Figueiredo M (2023) Morphometry and skeletopy of kidneys and renal vessels in Alouatta guraiba clamitans (Primates: Atelidae): case reports. Braz J Vet Med 45:e005222
Raubenheimer D, Machovsky-Capuska GE, Chapman CA, Rothman JM (2015) Geometry of nutrition in field studies: an illustration using wild primates. Oecologia 177:223–234. https://doi.org/10.1007/s00442-014-3142-0
Righini N, Garber PA, Rothman JM (2017) The effects of plant nutritional chemistry on food selection of Mexican black howler monkeys (Alouatta pigra): the role of lipids. Am J Primatol 79:e22524. https://doi.org/10.1002/ajp.22524
Rosenberger AL, Halenar L, Cooke SB (2011) The making of platyrrhine semifolivores: models for the evolution of folivory in primates. Anat Rec 294:2112–2130
Rothman JM, Raubenheimer D, Bryer MAH, Takahashi M, Gilbert CC (2014) Nutritional contributions of insects to primate diets: implications for primate evolution. J Hum Evol 71:59–69. https://doi.org/10.1016/j.jhevol.2014.02.016
Rothman JM, Raubenheimer D, Chapman CA (2011) Nutritional geometry: gorillas prioritize non-protein energy while consuming surplus protein. Biol Lett 7:847–849. https://doi.org/10.1098/rsbl.2011.0321
Santos GASD, Bianchini E, Reis NR (2013) Seasonal variation of consumption of the species used as fruit source by brown howler monkeys (Alouatta clamitans) in southern Brazil. Biota Neotrop 13:148–153. https://doi.org/10.1590/S1676-06032013000300019
Schwitzer C, Kaumanns W (2003) Foraging patterns of free-ranging and captive primates – implications for captive feeding regimes. In: Fidgett A, Clauss M, Ganslosser U, Hatt JM, Nijboer J (Eds) Zoo Animal Nutrition II. Filander, Fürth, pp 247–265.
Serio-Silva JC, Hernandez-Salazar LT, Rico-Gray V (1999) Nutritional composition of the diet of Alouatta palliata mexicana females in different reproductive states. Zoo Biol 18:507–513. https://doi.org/10.1002/(SICI)1098-2361(1999)18:6%3c507::AID-ZOO5%3e3.0.C0;2-R
Simpson SJ, Sibly RM, Lee KP, Behmer ST, Raubenheimer D (2004) Optimal foraging when regulating intake of multiple nutrients. Anim Behav 68:1299–1311. https://doi.org/10.1016/j.anbehav.2004.03.003
Steinmetz S (2001) Drinking by howler monkeys (Alouatta fusca) and its seasonality at the Intervales State Park, Sao Paulo, Brazil. Neotrop Primates 9:111–112
Stephens DW, Brown JS, Ydenberg RC (2008) Foraging behavior and ecology. University of Chicago Press, Chicago
Trapanese C, Meunier H, Masi S (2019) What, where and when: spatial foraging decisions in primates. Biol Rev 94:483–502. https://doi.org/10.1111/brv.12462
Vitale JJ, Velez H, Guzman C, Correa P (1963) Magnesium deficiency in the Cebus monkey. Circulation Res 12:642–650
Wielbass A, Amundin M, Laska M (2015) Gustatory responsiveness of black-and-white ruffed lemurs (Varecia variegata variegata) to food-associated sugars. Int J Primatol 36:460–472. https://doi.org/10.1007/s10764-015-9835-3
Windley HR, Starrs D, Stalenberg E, Rothman JM, Ganzhorn JU, Foley WJ (2022) Plant secondary metabolites and primate food choices: a meta-analysis and future directions. Am J Primatol 2022:e23397. https://doi.org/10.1002/ajp.23397
Youlatos D, Couette S, Halenar LB (2015) Morphology of howler monkeys: a review and quantitative analyses. In: Kowalewski MM, Garber PA, Cortes-Ortiz L, Urbani B, Youlatos D (eds) Howler monkeys: behavior, ecology, and conservation. Springer, New York, pp 133–176
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Silveira, P., Valler, Í.W., Hirano, Z.M.B. et al. Food preferences and nutrient composition in captive Southern brown howler monkeys, Alouatta guariba clamitans. Primates 65, 115–124 (2024). https://doi.org/10.1007/s10329-023-01109-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-023-01109-1