
Abstract
Identifying the factors that determine local extinction of populations is crucial to ensure species conservation. Forest-dwelling primates are especially vulnerable to habitat fragmentation, although few studies have provided systematic evidence of local extinctions. Over an 11-year period, approximately 100 remnant populations of the endangered Coimbra Filho’s titi monkey (Callicebus coimbrai) have been found within the geographic range of the species in Bahia and Sergipe, Northeast Brazil. During the present study, extinction of 13 of these populations was recorded through intensive surveys. These extinctions were detected from evidence of intensive logging and clear-cutting, interviews with local residents and systematic searches of the sites where occurrence of the species had been confirmed in previous surveys. These local extinctions represent approximately 10 % of the known populations of C. coimbrai and up to 28.3 % of the area occupied by the species. Comparison of the vegetation structure in fragments where extinction was recorded and where the species still occurs indicated that sparser understorey may be a correlate of extinction, combined with the fact that extinctions occurred within fragments characterised by relatively high levels of anthropogenic disturbance. These findings reinforce the Endangered status of the species and the urgent need for intensification of conservation measures within the most impacted areas of the geographic distribution of C. coimbrai.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Worldwide, many species face considerable threats, and current extinction rates are comparable to those of the past five mass extinction events (Barnosky et al. 2011). Habitat reduction and fragmentation, habitat degradation, introduction of exotic species, overexploitation of species and global warming are among the main threats to biodiversity currently (Primack and Rodrigues 2001; Barnosky et al. 2011). A species is considered extinct when exhaustive surveys within its original range fail to locate any individuals. It can also be considered extinct in Nature when there are no free-ranging individuals left, even though some do survive in captivity (IUCN 2012). Local extinction is a less drastic scenario, where populations of a species are lost within some areas, but remain at other localities (Primack and Rodrigues 2001).
The principal threats to survival of primate populations are deforestation, habitat degradation and fragmentation, and hunting (Mittermeier et al. 2012). Deforestation is the primary driving force affecting primate survival, since it causes both loss of habitat and a reduction in the quality and continuity of remaining habitat. One of the consequences of this process is reduction in the size of populations (Arroyo-Rodríguez and Mandujano 2009), which leaves them increasingly vulnerable to stochastic processes, including loss of genetic variability, and increasing endogamy and/or vulnerability to catastrophic events (Shaffer 1981), thereby greatly increasing the probability of extinction of affected populations.
In fragmented landscapes, where primates may have occurred originally throughout most of the environment, studies can assess extinction through occupancy surveys. In such environments, a number of studies have found evidence of a systematic relationship between occurrence of primate populations and specific attributes of the fragments (Onderdonk and Chapman 2000; Arroyo-Rodríguez et al. 2007; Raboy et al. 2010), the surrounding matrix (Hoffman and O’Riain 2012) or the species itself (Onderdonk and Chapman 2000). Fragment size is the most important correlate of local extinction, although there is not always a straightforward relationship between fragment size and the probability of occurrence of a given species (e.g. Onderdonk and Chapman 2000; Mbora and Meikle 2004; Arroyo-Rodríguez and Mandujano 2006; Boyle and Smith 2010).
Forest structure may also be important; For example, human activities, such as logging and collection of firewood, affect the structure of the forest (Ganzhorn 1995) and can promote exclusion of some species [e.g. Plecturocebus moloch: Michalski and Peres 2005—formerly Callicebus moloch, see Byrne et al. (2016); Colobus angolensis palliatus: Anderson et al. 2007] or benefit others (e.g. three species of lemurs: Ganzhorn 1995). The main effect of forest structure on primate occurrence and density is the change in the relative availability of different resources (Ganzhorn 1995; Rode et al. 2006). Forest structure may also be related to the abundance of some plant groups, such as lianas (Parthasarathy et al. 2004), palms (Farris-Lopez et al. 2004) and bamboo (Palminteri and Peres 2012), which can affect primate presence and density (Palminteri and Peres 2012; Hilário 2013).
Little is known regarding the factors that influence the occurrence of titis (Callicebinae) in forest fragments. Michalski and Peres (2005) found that the occurrence of P. moloch populations in a fragmented landscape in the southern Amazon Basin was inversely related to selective logging, but not to factors such as fragment size, connectivity, isolation time, habitat quality or hunting pressure. In a second study, Ferrari et al. (2003) concluded that the occurrence and density of P. moloch in different fragments were affected primarily (and negatively) by the presence and density of the silvery marmoset, Mico argentatus. Clearly, then, even populations of the same species may respond to threats in different ways, emphasising the need to avoid generalisations (Marsh et al. 2003; Isaac and Cowlishaw 2004).
Coimbra Filho’s titi (Callicebus coimbrai) was described in 1999 (Kobayashi and Langguth 1999), and was almost immediately classified as Critically Endangered by the International Union for Conservation of Nature (IUCN). However, the increasing number of populations identified during subsequent surveys (Jerusalinsky et al. 2006; Sousa et al. 2008; Marques et al. 2013) led to reclassification of the species’ conservation status as Endangered (Veiga et al. 2008). This does not mean, however, that there is much less cause for concern, given that the vast majority of C. coimbrai populations are confined to small, isolated fragments of less than 100 ha, where they are extremely vulnerable to extinction (Jerusalinsky 2013). During recent surveys for identification of new C. coimbrai populations, evidence of local extinction was found at a number of sites located throughout the species’ range. In addition to providing evidence on these extinctions, the present study discusses its possible determinants as well as the implications of these findings for the conservation status of the species.
We predict that local extinction will be more likely in smaller fragments (Onderdonk and Chapman 2000; Mbora and Meikle 2004; Arroyo-Rodríguez and Mandujano 2006; Boyle and Smith 2010) and in forests with lower biomass. Lower biomass, which is an aspect of forest structure, should reflect lower resource availability (Peres 2008), leading to lower primate density and an increase in the probability of extinction (Oates et al. 1990; Rode et al. 2006; Anderson et al. 2007). As forest disturbance has already been implicated in local extinctions of P. moloch (Michalski and Peres 2005), and given that fruit production is correlated with humidity (van Schaik et al. 1993), we also predict that local extinctions of C. coimbrai may be related negatively to indicators of humidity and conservation (bromeliads and palms—Gentry and Dodson 1987; DeWalt et al. 2003) and positively to indicators of disturbance, i.e. bamboo (Tabarelli and Mantovani 2000).
Methods
Study area
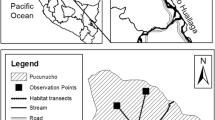
The present study focussed on the known geographic distribution of Coimbra Filho’s titi (Callicebus coimbrai), which coincides primarily with the Brazilian Atlantic Forest domain of the States of Sergipe and northern Bahia, as far south as the Paraguaçu River (Fig. 1). The forest cover of this region has been extensively devastated, with less than 11.5 % of the original habitat remaining (Fundação SOS Mata Atlântica and INPE 2013). Deforestation is ongoing in many areas.

Sites in Northeast Brazil (Bahia and Sergipe) at which populations of Callicebus coimbrai are present or were known to exist in the past. The extinction “polygon” is highlighted in the south-eastern portion of the map. The smaller polygon encompasses a group of nearby fragments in which recent extinction of C. coimbrai was confirmed in interviews. Red fragments: extinction of C. coimbrai confirmed by playback surveys (category ii); brown fragments: surveyed non-systematically with playback; purple fragments: extinction reported to have occurred more than 10 years prior to the interview (not included in analysis); pink fragments: recent extinctions reported in interviews (category i); blue fragments: extinction inferred from either complete suppression or intense degradation of forest (category iii); black fragments: with C. coimbrai in which vegetation was sampled for comparison with fragments in which extinction was confirmed through playback surveys; green fragments: occurrence of C. coimbrai reported in previous studies. Fragment numbers: 1 Mata da Cova; 2 Conceição; 3 Fazenda Itamarati; 4 Canabrava 3; 5 Usina Aliança/São Bento Inhatá; 6 Faz. Juerana; 7 Subaumirim; 8 Lontra; 9 Cachoeira II; 10 Faz. Gameleira; 11 Pov. Pereira; 12 Pov. Jangada; 13 Gameleira III; 14 Faz. Boa Nova (Map: Daniela P. B. Ruiz-Esparza) (color figure online)
The majority of the remnants of Atlantic Forest found within the geographic range of C. coimbrai are relatively small, isolated fragments, with a predominance of disturbed habitat (Hilário 2013; Jerusalinsky 2013; Marques et al. 2013). As the region is densely populated, most fragments are also located in close proximity to human settlements, ranging from urban centres to rural communities, which reflects ongoing anthropogenic pressures, such as hunting and exploitation of lumber, firewood and other forest resources. Local agriculture is dominated by cattle ranching and plantations of Eucalyptus, orange and sugarcane.
Collection of evidence on local extinction
The present study was based on data obtained during studies aimed at locating new populations of C. coimbrai through interviews and non-systematic playback surveys (Jerusalinsky 2013), and identifying correlates of the density of the species through systematic playback surveys and vegetation sampling within the known geographic distribution of the species (Hilário 2013). Following preliminary identification of potential sites based on analysis of satellite images and aerial photographs, localities throughout the whole study area were visited to confirm occurrence of titi populations in situ (Jerusalinsky 2013). We focussed sampling primarily on those fragments larger than 100 ha, since these are more likely to support populations of Callicebus coimbrai. Nevertheless, we also included smaller fragments whenever interviews pointed to occurrence of the species at these sites.
Between 2004 and 2012, we conducted 593 non-directed semi-structured interviews (Chizzotti 2006; Richardson et al. 2008) with local residents and other knowledgeable individuals, such as government employees. We used snowball sampling strategies to select specific informants (Davis and Wagner 2003; Browne 2005). Interviews are a tool that allows researchers to acquire information on occurrence of a species much more quickly than by surveying each locality, and are thus ideal for studies that encompass a wide geographic region. This method has been applied successfully in a number of previous studies (Coimbra-Filho et al. 1991; Ferreira et al. 2009) and is considered reliable, especially for species with conspicuous vocalisations, such as titis (Jerusalinsky et al. 2006; Marques et al. 2013). We considered the interviews to be reliable because we talked with informers known locally as people with extensive local environmental knowledge, i.e. “experts” (Davis and Wagner 2003).
Interviews began informally with questions regarding the largest fragments of forest within the local area. We subsequently enquired about the animals that occur in these forest fragments. Due to its conspicuous vocalisation, C. coimbrai was frequently one of the first species cited whenever titis occurred in the fragment. When informants referred to extinction of the species from specific sites, additional details were requested. These reports were considered reliable when the informant provided evidence of a detailed knowledge of the species, such as its morphological and ecological traits, and in particular, its characteristic vocalisation, as well as being able to identify the site of the most recent encounter (sighting or hearing), and the approximate date of this event. We also used a field guide with plates of different primates (Mittermeier et al. 2007) to certify the identification of the species present in the forest remnant indicated by the informant.
The evidence of extinction analysed in the present study falls into three categories: (i) reports of extinction occurring within the preceding 11 years (2002–2012), (ii) extinction of populations identified during previous surveys conducted between 2002 and 2012 (Jerusalinsky et al. 2006; Sousa et al. 2008; Marques et al. 2013) and (iii) extinction of populations identified during previous surveys resulting from clear-cutting or extreme habitat degradation (suppressed areas), or inferred by non-systematic surveys (Tables 1, 2, 3). We also obtained 11 reports of local extinction occurring prior to the present study period, i.e. more than 10 years prior to the interviews. However, these records were not included in the analysis because the discovery of most of the populations of C. coimbrai took place during the study period. Thus, including events that occurred before the study period would bias the calculation of an extinction rate.
As part of a wider study (Hilário 2013), in 2012, the four fragments in the second category were surveyed by playback using a digital recording of a typical titi duet (of the closely related Callicebus personatus, obtained from Emmons et al. 1997), which was broadcast through a 20-W megaphone. Two minutes of long calls were broadcast, followed by a 10-min interval to record the response of C. coimbrai, which was recorded whenever this species was heard or sighted. Standard surveys were conducted along linear transects, with playback conducted at 200-m intervals along the transect between 15 and 30 min after sunrise and 10:00 h. This standard procedure was also applied at 19 other sites where occurrence of C. coimbrai had been confirmed, providing data for the calculation of an index of probability of non-response to the playback, that is, the probability that members of the species were present in the fragment but did not respond to the playback. The number of playback points conducted each day varied from 8 to 13 (mean 10.8 ± 1.3), and the sampling was replicated over three days in each fragment. The mean number of responses per transect point did not deviate significantly from a normal distribution (Shapiro–Wilk W = 0.9642; p = 0.0898; N = 57), and this number was used to calculate the probability of obtaining no response during a day of survey at a site occupied by titis, based on a normal distribution, with the mean number of responses and variance equal to that of the data. This analysis was run in BioEstat 5.0 (Ayres et al. 2007).
Vegetation sampling and fragment size
A standardised survey of vegetation was also conducted at all sites surveyed by playback (except for one site at which extinction was confirmed, due to logistic problems) for identification of potential factors affecting the occurrence and density of C. coimbrai and other primates. The vegetation surveys were based on the point-centred quarter method (Cottam and Curtis 1956), with five points sampled at intervals of 50 m along randomly selected transects. The transects were separated by a distance of at least 300 m from one another. The number of transects established in each fragment varied depending on its size, ranging from four (in fragments <150 ha) to 23 (in fragments >1000 ha). At each point, the nearest tree with diameter at breast height (DBH) of at least 5 cm in each quadrant was identified, and the following data were collected: distance from the tree to the centre of the sampling point, height of the lowest branch, and DBH. The DBH was then converted into a basal area (BA) for each tree using the following formula: BA = π(DBH/2)2. We also assessed understorey density using a sighting target [adapted from Barlow et al. (2002) and McIntyre (1995)], consisting of a 20 cm × 20 cm red plastic card, which was hung around the neck of the principal investigator, who moved backwards away from the field assistant standing at the sampling point until no part of the card could be visualised. The distance between the sampling point and the final position of the target was measured with a surveyor’s tape.
Canopy openness was assessed using a 30 cm × 30 cm poly(vinyl chloride) (PVC) pipe frame divided into 100 squares of equal size using nylon lines. The frame was held at arm’s length above the head of the principal investigator at each sampling point, and the number of squares in which the sky was visible was estimated. These variables are related to forest biomass: areas with more trees (shorter distances to the centre of the sampling point), taller trees (and canopy), thicker trees, denser understorey and closed canopy all reflect higher biomass. Nevertheless, these variables are not necessarily positively correlated with biomass or with one another. Other data were also collected for each point: canopy height (m) and the abundance of bromeliads and bamboo (using standardised indices ranging from zero to three, with zero corresponding to absence; one, to when these plants were rare around the point; two, to when these plants were common; and three, to maximum abundance), lianas (number of stems within a radius of 3 m), and palms (number of individuals of height >2 m within a radius of 20 m). All of these variables were useful to assess forest structure and composition, which is related to the level of conservation/degradation of the forest and to resource availability (DeWalt et al. 2003; Boubli et al. 2010).
The size of each fragment was measured from satellite images using Global Mapper version 7.01 (Blue Marble Geographics 2005). In two cases, it was impossible to obtain adequate satellite images, so fragment size was obtained from Sousa et al. (2008).
Analysis of correlates of extinction
The Wilcoxon test was used to compare the scores recorded for vegetation variables and fragment size between the two groups of sites, i.e. those with C. coimbrai populations (19 fragments) and those without C. coimbrai populations (three fragments). As the number of sites in the latter group was small, and thus relatively vulnerable to error, 95 % confidence intervals were calculated for each of the variables that returned significant differences in the Wilcoxon test for comparison. All sites were mapped using ArcGIS 10 (ESRI 2011).
Results
Overall, evidence of local extinction or probable extinction of C. coimbrai populations over the 11 years between 2002 and 2012 was obtained for 13 fragments (Tables 1, 2, 3; Fig. 1). In addition, 11 reports of local extinction occurring prior to the present study period, i.e. more than 10 years prior to the interviews, were also obtained, although these records were not included in the analysis.
Four of the five sites at which extinction was reported in the interviews (category i) are located in the southern portion of the range of C. coimbrai, while the other is in the northern extreme (Fig. 1). Deforestation was cited as the principal cause of extinction of titi populations by all informants, followed by hunting, although C. coimbrai was not identified as a preferred game species.
The mean number of playback responses obtained per point per day of survey in the fragments where C. coimbrai was known to occur was 1.7 ± 0.7 (range 0.0–2.7). Based on these values, the probability of not obtaining a response assuming that titis were present in the four sites surveyed systematically ranged between 0.000177 and 0.0604, according to the number of days on which each site was surveyed, and the number of survey points (Table 4). The responsiveness of Callicebus species to playbacks and the probabilities presented in Table 4 make it highly unlikely that titis would have been present in these fragments and did not respond. Based on this analysis, the evidence of local extinction of C. coimbrai from the forest fragments at Lontra, Altamira I, Subaumirim and Cachoeira (category ii) was assumed to be conclusive.
Extinction of C. coimbrai was suspected at two other fragments—Pereira and Fazenda Gameleira (category iii)—which were not surveyed systematically during the present study. At Pereira, the playback was broadcast only twice on one afternoon, and at Fazenda Gameleira, it was played on a number of different occasions, at irregular intervals, between 09:00 and 11:30 h. At both sites, in addition to the lack of response to the playback, local residents invariably reported not knowing of the occurrence of the species in the fragment, which is atypical of sites in which titis occur, given the potent and characteristic vocalisation of these monkeys. At Fazenda Gameleira, evidence was also found of ongoing logging and habitat degradation.
At two sites (12 and 13), extinction of C. coimbrai was due to deforestation (category iii). At site 12 (Jangada), the original fragment, identified by Sousa et al. (2008), had been clear-cut, while at Gameleira III (site 13), intensive logging had left a sparsely treed landscape dominated by a grassy matrix.
All sites in categories ii and iii are located within the same general area in the southern portion of the range of C. coimbrai (Fig. 1). This region was denominated the extinction polygon and is characterised by a general lack of titi populations.
The eight fragments for which extinction of C. coimbrai was confirmed or inferred (categories ii and iii) varied in size from 60 to 3497 ha, with total area of 6008 ha (Tables 2, 3). One of the authors (Sidnei Santos, unpublished data) recorded the presence of C. coimbrai in all of these fragments between 2002 and 2004, confirming local extinction of the species within an 11-year timeframe. The five other sites (category i) had area of 3–300 ha (Table 1), with total area of 368 ha. This total of 13 fragments represents 9.9 % of the approximately 131 fragments in which the species is known or presumed to occur (Jerusalinsky et al. 2006; Sousa et al. 2008; Jerusalinsky 2013; Marques et al. 2013).
Statistical analysis of the habitat variables (Table 5) indicated that extinction of C. coimbrai is associated (inversely) with two factors: the density of the understorey and the occurrence of bamboo. The association with bamboo is relatively weak, however, and the values recorded at the sites from which titis were absent are subsumed in the 95 % confidence interval recorded for the sites at which the species was present (Table 6). In the case of understorey density, by contrast, only one of the extinction sites overlaps minimally with the upper limit of the confidence interval. This indicates that more open understorey may contribute to extinction of C. coimbrai.
Discussion
The present study confirmed local extinction of C. coimbrai at six sites and found strong evidence of extinction at two others (Pereira and Fazenda Gameleira), all concentrated within the southern portion of the species’ geographic distribution. Unconfirmed reports of extinction were also obtained at a further four sites in the south-western extreme of the range and another one in the northern extreme. If stochastic processes were the cause of the extinctions reported here, we would expect to observe an even distribution of these cases of extinction throughout the study area. The concentration of local extinctions within a relatively small portion of the species’ distribution is especially worrying, given that it suggests the role of specific local factors—presumably anthropogenic—rather than stochastic processes. The fragments that have been clear-cut are the most obvious example, although even where the forest has persisted, ongoing habitat degradation and in particular logging appear to be more intense than at other sites (pers. obs.). Michalski and Peres (2005) identified selective logging as a primary determinant of local extinction of P. moloch in the Amazon Basin.
However, the possible extinction of C. coimbrai at Lontra is contentious, given the large size of the fragment (3497 ha) and the relatively low levels of habitat disturbance, which is restricted mainly to the edges of the fragment. While sampling effort at this site was greater than that at the other sites (Table 4), it was highly disproportionate to the size of the fragment, especially if population density were very low and/or the local C. coimbrai population has a discontinuous distribution within the fragment. Most of the local residents at Lontra did not have any knowledge of titis whatsoever, although one informant did confirm that the species is found in specific portions of the fragment. This suggests a situation similar to that found at Fazenda Boa Nova (1591 ha), where a similar number of playbacks (34) evoked only five responses—corresponding to a response rate 3.9–16 times smaller than other sites—concentrated within a small portion of the fragment. If such low densities are a characteristic of the extinction polygon, this may have been an additional factor contributing to the local extinction of so many populations within this area (Fig. 1). It should be noted, however, that the likelihood that titis were present at Lontra but did not respond to the playback appears to be extremely low (Table 4), reinforcing the conclusion that the species has become extinct at this site. All the other fragments were surveyed more comprehensively, and the evidence on extinction appears to be more conclusive.
Recording the presence of a species is a more straightforward process than confirming its absence, given that only a single record is required. Confirming the absence of a species is more complex, given that the species may occur at the site but remain undetected. In this context, our systematic playback survey and calculation of the statistical probability of the presence of the species in a fragment is an approach that could be used by other researchers to provide more conclusive evidence of the absence of a species at a given locality.
Our hypothesis that forest characteristics that reflect lower biomass would predict local extinction was accepted, given that the fragments where these extinctions occurred had sparser understorey. The apparent preference of C. coimbrai for forests with relatively dense understorey may be related to the fact that most titi species, including C. coimbrai (Souza-Alves 2010), tend to prefer the middle and lower strata of the forest (Kinzey 1981; Carrillo-Bilbao et al. 2005; Bicca-Marques and Heymann, 2013). Hilário (2013) also found a correlation between the density of the understorey and the population density of C. coimbrai at the other (non-extinction) sites surveyed. Coincidentally, a relatively dense understorey also appeared to favour the density of common marmosets (Callithrix jacchus) at the same sites (Hilário and Ferrari 2015). No aspect of the abundance of any plant group was related to local extinction.
Contrary to our hypothesis, and despite the importance of fragment size for many other primates (e.g. Arroyo-Rodríguez and Mandujano 2006; Anderson et al. 2007; Boyle and Smith 2010), this variable did not appear to be a reliable predictor of extinction risk in C. coimbrai. Most of the extinctions reported here were recorded in fragments larger than 100 ha, whereas dozens of populations are known to occur in fragments of much smaller size (Jerusalinsky 2013). This is far from encouraging, however, and the fact that even relatively large fragments presented extinctions may also have important implications for other species. The general notion that larger areas host larger populations that are less vulnerable to local extinction (Shaffer 1981), supported by studies that found a relation between fragment size and primate occupancy, can wrongly lead to the idea that large fragments are a simple and efficient choice for conservation programs. The absence of C. coimbrai from Lontra, a protected area and one of the largest fragments in which the species was detected in previous surveys, indicates that conservation efforts should not focus exclusively on the largest fragments. There is a clear need, not only to protect large fragments, but also to evaluate whether these fragments meet the needs of the species in terms of forest structure and resource availability before implementing a conservation program. In a number of previous studies, fragment size also did not have a significant effect on patch occupancy by primate species, including P. moloch (Michalski and Peres 2005), Alouatta pigra (Estrada et al. 2002), Colobus guereza, Procolobus pennantii and Cercopithecus ascanius (Onderdonk and Chapman 2000), confirming that this may be a more general pattern.
Jerusalinsky (2013) estimated a total occupied area of 22,500 ha for C. coimbrai in 131 fragments, which would mean that the species became extinct in at least 12.8 % (2897 ha) of this area between 2002 and 2012. Assuming extinction at Lontra, this percentage would rise to 28.3 % (6376 ha). Optimistically, even if the species does still occur at Lontra and the total area occupied has been underestimated, it still seems reasonable to conclude that approximately 10 % of the total area and the total number of fragments occupied were lost over the 11 years of the study period. Either way, the evidence is far from encouraging with regard to the long-term prospects for the species. In addition, the concentration of extinctions within a central portion of the species’ geographic distribution may lead to the formation of two isolated nuclei of populations, accentuating the potential problems for metapopulation management.
The current Endangered status of C. coimbrai is derived from IUCN criteria B2ab(ii,iii,iv,v), C1, and C2a(i) (Veiga et al. 2008), which refer to an occupied area of less than 500 km2, severe habitat fragmentation, ongoing reduction in the area occupied, habitat quality, the number of populations, and fewer than 2500 mature individuals. The loss of occupied area and populations recorded in the present study is consistent with criteria B2ab(iv) and C1, respectively, and reinforces the classification of the species as Endangered, but falls short of the criteria for a Critically Endangered status. The trends observed here are nevertheless worrying, and reinforce the need for urgent actions (e.g. protection of fragments, maintenance of riparian forests as wildlife corridors, recovery of areas of illegal deforestation) to mediate the ongoing processes of habitat degradation and loss that continue to threaten the remaining populations of C. coimbrai. The findings of the present study also indicate that populations of other primate species may have been lost silently and encourage other conservationists to routinely revisit locations already surveyed to guarantee the persistence of populations at these sites.
References
Anderson J, Cowlishaw G, Rowcliffe JM (2007) Effects of forest fragmentation on the abundance of Colobus angolensis palliatus in Kenya’s Coastal Forests. Int J Primatol 28:637–655
Arroyo-Rodríguez V, Mandujano S (2006) Forest fragmentation modifies habitat quality for Alouatta palliata. Int J Primatol 27:1079–1096
Arroyo-Rodríguez V, Mandujano S (2009) Conceptualization and measurement of habitat fragmentation from the primates’ perspective. Int J Primatol 30:497–514
Arroyo-Rodríguez V, Mandujano S, Benítez-Malvido J, Cuende-Fanton C (2007) The influence of large tree density on howler monkey (Alouatta palliata mexicana) presence in very small rain forest fragments. Biotropica 39:760–766
Ayres M, Ayres Jr. M, Ayres DL, Santos AAL (2007) BioEstat 5.0
Barlow J, Haugaasen T, Peres CA (2002) Effects of ground fires on understorey bird assemblages in Amazonian forests. Biol Conserv 105(2):157–169
Barnosky AD, Matzke N, Tomiya S, Wogan GO, Swartz B, Quental TB, Marshall C, McGuire JL, Lindsey EL, Maguire KC, Mersey B, Ferrer EA (2011) Has the Earth’s sixth mass extinction already arrived? Nature 471:51–57
Bicca-Marques JC, Heymann EK (2013) Ecology and behavior of titi monkeys (genus Callicebus). In: Veiga LM, Barnett A, Ferrari SF, Norconk M (eds) Evolutionary biology and conservation of Titis, Sakis, and Uacaris. Cambridge University Press, Cambridge, pp 196–207
Blue Marble Geographics (2005) Global Mapper 7.01
Boubli JP, Couto-Santos FR, Mourthé IMC (2010) Quantitative assessment of habitat differences between northern and southern muriquis (Primates, Atelidae) in the Brazilian Atlantic Forest. Ecotropica 16:63–69
Boyle SA, Smith AT (2010) Can landscape and species characteristics predict primate presence in forest fragments in the Brazilian Amazon? Biol Conserv 143:1134–1143
Browne K (2005) Snowball sampling: using social networks to research non-heterosexual women. Int J Soc Res Methodol 8:47–60
Byrne H, Rylands AB, Carneiro JC, Lynch-Alfaro JW, Bertuol F, da Silva MN, Messias M, Groves CP, Mittermeier RA, Farias I, Hrbek T, Schneider H, Sampaio I, Boubli JP (2016) Phylogenetic relationships of the New World titi monkeys (Callicebus): first appraisal of taxonomy based on molecular evidence. Front Zool 13:1–25
Carrillo-Bilbao G, Di Fiore A, Fernández-Duque E (2005) Dieta, Forrajeo y Presupuesto de Tiempo en Cotoncillos (Callicebus discolor) del Parque Nacional Yasuní en la Amazonia Ecuatoriana. Neotrop Primates 13:7
Chizzotti A (2006) Pesquisa qualitativa em ciências humanas e sociais. Vozes, Petrópolis
Coimbra-Filho AF, Rylands AB, Pissinatti A, Santos IB (1991/1992) The distribution and conservation of the buff headed capuchin monkey, Cebus xanthosternos, in the Atlantic forest region of eastern Brazil. Primate Conserv 12–13:24–30
Cottam G, Curtis JT (1956) The use of distance measures in phytosociological sampling. Ecology 37:451–460
Davis A, Wagner JR (2003) Who knows? On the importance of identifying “experts” when researching local ecological knowledge. Hum Ecol 31:463–489
DeWalt SJ, Maliakal SK, Denslow JS (2003) Changes in vegetation structure and composition along a tropical forest chronosequence: implications for wildlife. Forest Ecol Manag 182:139–151
Emmons LH, Whitney BT, Ross DL (1997) Sounds of neotropical rainforest mammals: an audio field guide. Cornell Laboratory of Ornithology, Ithaca
ESRI (2011) ArcGIS 10
Estrada A, Mendoza A, Castellanos L, Pacheco R, Van Belle S, García Y, Muñoz D (2002) Population of the black howler monkey (Alouatta pigra) in a fragmented landscape in Palenque, Chiapas, Mexico. Am J Primatol 58:45–55
Farris-Lopez K, Denslow JS, Moser B, Passmore H (2004) Influence of a common palm, Oenocarpus mapora, on seedling establishment in a tropical moist forest in Panama. J Trop Ecol 20:429–438
Ferrari SF, Iwanaga S, Ravetta AL, Freitas FC, Sousa BAR, Souza LL, Costa CG, Coutinho PEG (2003) Dynamics of primate communities along the Santarem–Cuiaba highway in south-central Brazilian Amazonia. In: Marsh L (ed) Primates in fragments: ecology and conservation. Springer, New York, pp 123–144
Ferreira RG, Jerusalinsky L, Silva TCF, Fialho MS, Roque AA, Fernandes A, Arruda F (2009) On the occurrence of Cebus flavius (Schreber 1774) in the Caatinga, and the use of semi-arid environments by Cebus species in the Brazilian state of Rio Grande do Norte. Primates 50:357–362
Fundação SOS Mata Atlântica, INPE (2013) Atlas dos remanescentes florestais da Mata Atlântica—Período de 2011 a 2012. Fundação SOS Mata Atlântica/INPE, São Paulo
Ganzhorn J (1995) Low-level forest disturbance effects on primary production, leaf chemistry, and lemur populations. Ecology 76:2084–2096
Gentry AW, Dodson CH (1987) Diversity and biogeography of neotropical vascular epiphytes. Ann Mo Bot Gard 74:205–233
Hilário RR (2013) Determinantes ambientais da densidade de Callicebus coimbrai no nordeste brasileiro e implicações para a sua conservação, PhD thesis. Universidade Federal da Paraíba, João Pessoa
Hilário RR, Ferrari SF (2015) Dense understory and absence of capuchin monkeys (Sapajus xanthosternos) predict higher density of common marmosets (Callithrix jacchus) in the Brazilian Northeast. Am J Primatol 77:425–433
Hoffman TS, O’Riain MJ (2012) Landscape requirements of a primate population in a human-dominated environment. Front Zool 9:1–17
Isaac NJB, Cowlishaw G (2004) How species respond to multiple extinction threats. Proc R Soc Lond B Biol Sci 271:1135–1141
IUCN (2012) IUCN Red List categories and criteria: Version 3.1, 2nd edn. IUCN, Gland
Jerusalinsky L (2013) Distribuição geográfica e conservação de Callicebus coimbrai Kobayashi & Langguth, 1999 (Primates–Pitheciidae) na Mata Atlântica do nordeste do Brasil, PhD thesis. Universidade Federal da Paraíba, João Pessoa
Jerusalinsky L, Oliveira MM, Pereira RF, Santana V, Bastos PCR, Ferrari SF (2006) Preliminary evaluation of the conservation status of Callicebus coimbrai Kobayashi & Langguth, 1999 in the Brazilian State of Sergipe. Primate Conserv 21:25–32
Kinzey WG (1981) The titi monkeys, genus Callicebus. In: Coimbra-Filho A, Mittermeier RA (eds) Ecology and behavior of neotropical primates, vol 1. Academia Brasileira de Ciências, Rio de Janeiro, pp 241–276
Kobayashi S, Langguth A (1999) A new species of titi monkey, Callicebus Thomas, from north-eastern Brazil (Primates, Cebidae). Rev Bras Zool 16:531–551
Marques ELN, Jerusalinsky L, Rocha JCAG, Santos PM, Beltrão-Mendes R, Ferrari SF (2013) Primates, Pitheciidae, Callicebus coimbrai Kobayashi and Langguth 1999: new localities for an endangered titi monkey in eastern Sergipe, Brazil. Check List 9:696–699
Marsh L, Chapman C, Norconk MA, Ferrari SF, Gilbert KA, Bicca-Marques JC, Wallis J (2003) Fragmentation: specter of the future or the spirit of conservation. In: Marsh L (ed) Primates in fragments: ecology and conservation. Springer, New York, pp 381–398
Mbora DNM, Meikle DB (2004) Forest fragmentation and the distribution, abundance and conservation of the Tana river red colobus (Procolobus rufomitratus). Biol Conserv 118:67–77
McIntyre NE (1995) Effects of forest patch size on avian diversity. Landscape Ecol 10(2):85–99
Michalski F, Peres CA (2005) Anthropogenic determinants of primate and carnivore local extinctions in a fragmented forest landscape of southern Amazonia. Biol Conserv 124:383–396
Mittermeier RA, Coimbra-Filho AF, Kierulff MCM, Rylands AB, Mendes SL, Pissinatti A, Almeida LM (2007) Monkeys of the Atlantic forest of eastern Brazil: pocket identification guide. Conservation International, Arlington
Mittermeier RA, Rylands AB, Schwitzer C, Taylor LA, Chiozza F, Williamson EA (2012) Primates in Peril: the world’s 25 most endangered primates 2010–2012. IUCN/SSC Primate Specialist Group, International Primatological Society, Conservation International, Arlington
Oates JF, Whitesides GH, Davies AG, Waterman PG, Green SM, Dasilva GL, Mole S (1990) Determinants of variation in tropical forest primate biomass: new evidence from West Africa. Ecology 71:328–343
Onderdonk D, Chapman C (2000) Coping with forest fragmentation: the primates of Kibale National Park, Uganda. Int J Primatol 21:587–611
Palminteri S, Peres CA (2012) Habitat selection and use of space by bald-faced sakis (Pithecia irrorata) in southwestern Amazonia: lessons from a multiyear, multigroup study. Int J Primatol 33:401–417
Parthasarathy N, Muthuramkumar S, Reddy MS (2004) Patterns of liana diversity in tropical evergreen forests of peninsular India. Forest Ecol Manag 190:15–31
Peres CA (2008) Soil fertility and arboreal mammal biomass in tropical forests. In: Carson W, Schnitzer S (eds) Tropical forest community ecology. Wiley-Blackwell, New York, pp 349–364
Primack R, Rodrigues E (2001) Biologia da conservação. Planta, Londrina
Raboy BE, Neves LG, Zeigler S, Saraiva NA, Cardoso N, Santos GR, Ballou JD, Leimgruber P (2010) Strength of habitat and landscape metrics in predicting golden-headed lion tamarin presence or absence in forest patches in southern Bahia, Brazil. Biotropica 42:388–397
Richardson RJ, Peres JAS, Wanderley JCV, Correia LM, Peres MHM (2008) Pesquisa Social: métodos e técnicas, 3rd edn. Atlas, São Paulo
Rode KD, Chapman CA, Mcdowell LR, Stickler C (2006) Nutritional correlates of population density across habitats and logging intensities in redtail monkeys (Cercopithecus ascanius). Biotropica 38:625–634
Shaffer ML (1981) Minimum population sizes for species conservation. Bioscience 31:131–134
Sousa MC, Santos SS, Valente MCM (2008) Distribuição e variação na pelagem de Callicebus coimbrai (Primates, Pitheciidae) nos estados de Sergipe e Bahia, Brasil. Neotrop Primates 15:54–59
Souza-Alves JP (2010) Ecologia alimentar de um grupo de Guigó-de-Coimbra-Filho (Callicebus coimbrai Kobayashi & Langguth, 1999): perspectivas para a conservação da espécie na paisagem fragmentada do sul de Sergipe, Master’s thesis. Universidade Federal de Sergipe, São Cristóvão
Tabarelli M, Mantovani W (2000) Gap-phase regeneration in a tropical montane forest: the effects of gap structure and bamboo species. Plant Ecol 148:149–155
Veiga LM, Sousa MC, Jerusalinsky L, Ferrari SF, Oliveira MM, Santos SSD, Valente, MCM, Printes RC (2008) Callicebus coimbrai. The IUCN red list of threatened species. http://www.iucnredlist.org/details/39954/0. Accessed 28 June 2016
van Schaik CP, Terborgh JW, Wright SJ (1993) The phenology of tropical forests: adaptive significance and consequences for primary consumers. Ann Rev Ecol Evol 24:353–377
Acknowledgements
We are grateful to CAPES for graduate stipends to R.R.H. and R.B.-M., Idea Wild, and Conselho Nacional de Pesquisa (CNPq processes: 476064/2008–2 and 303994/2011–8). We thank CNPq for a research fellowship to R.B.-M. (503372/2014-5), and CAPES for a postdoctoral stipend to S.F.F. (process 99999.001536/2015-00). We also thank Mohamed bin Zayed Species Conservation Fund (process: 12055114), Primate Action Fund (1001257) and Primate Conservation Inc. (1158), for financial support to R.B.-M.; and Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) and Secretaria de Estado do Meio Ambiente e dos Recursos Hídricos (SEMARH/Sergipe) for field research support. We would like to thank Daniela Ruiz-Esparza, Copener and the owners of the properties visited during this study. We also thank the editor and two anonymous reviewers for their valuable comments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Statement on the welfare of animals
All applicable international, national and/or institutional guidelines for the care and use of animals were followed.
About this article

Cite this article
Hilário, R.R., Jerusalinsky, L., Santos, S. et al. A primate at risk in Northeast Brazil: local extinctions of Coimbra Filho’s titi (Callicebus coimbrai). Primates 58, 343–352 (2017). https://doi.org/10.1007/s10329-017-0599-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-017-0599-6