
Abstract
The small-bodied mouse lemurs of Madagascar (Microcebus) are capable of heterothermy (i.e., torpor or hibernation). The expression of these energy-saving strategies has been physiologically demonstrated in three species: M. berthae, the pygmy mouse lemur (daily torpor), M. murinus, the gray mouse lemur (daily torpor and hibernation), and M. griseorufus, the reddish-gray mouse lemur (daily, prolonged torpor and hibernation). Additional evidence, based on radiotracking and seasonal body mass changes, indicated that mouse lemur capabilities for heterothermy extended to M. lehilahytsara, the Goodman’s mouse lemur. In this study, we confirm the use of hibernation in Goodman’s mouse lemurs at a new location, a high-plateau forest fragment in Ankafobe, central Madagascar. Our evidence is based on sleeping site monitoring of radiocollared individuals and the retrieval of three mouse lemurs from inside a tree hole, all of which displayed a lethargic state. Though our data are preliminary and scant, we show that hibernation occurs in high-plateau mouse lemurs, and suggest that a buffered environment (i.e., tree holes instead of nests) may be crucial to avoiding potentially extreme ambient temperatures.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The mouse lemurs of Madagascar (genus Microcebus) are one of the most speciose groups of lemurs, with 24 species described thus far (Hotaling et al. 2016). They are, as a group, the most pervasive in Madagascar’s forests, and can be found even in degraded and fragmented habitats close to human habitation. The ability of mouse lemurs to survive erratic climate patterns and high resource seasonality in their habitats stems from their ecological flexibility. They are opportunistic omnivores and rely on a wide range of food resources, including gums, arthropods, reptiles, fruits, and flowers (Thorén et al. 2011). Mouse lemurs also display unusual physiological plasticity, one example being the use of daily/prolonged torpor or hibernation (i.e., heterothermy) by some individuals, populations, and species (Schülke and Ostner 2007; Schmid and Ganzhorn 2009; Kobbe et al. 2011).
Daily torpor (i.e., controlled reductions in body temperature and metabolic rates for periods of less than 24 h) has previously been physiologically demonstrated in two mouse lemur species, M. berthae (one of the smallest species, referred to as M. myoxinus in the original study) and M. murinus (the gray mouse lemur) at Kirindy, a dry deciduous forest in western Madagascar (Ortmann et al. 1997; Schmid 2001). Prolonged torpor and hibernation—controlled reductions in temperature and metabolic rates for up to a couple of days (in the former) or several weeks at a time (in the latter)—have been indisputably reported in two species: the gray mouse lemur from the littoral southeastern forests, and M. griseorufus (the reddish-gray mouse lemur) from spiny southern forests (Schmid and Ganzhorn 2009; Kobbe et al. 2011). Skin temperature profiles, used as a proxy for metabolic rates, showed that a subset of mouse lemurs were able to passively track the ambient temperature over periods of several days up to several weeks.
Indirect evidence of heterothermy, as ascertained by the lack of nocturnal sightings during active phases and/or observations of fat accumulation in captured individuals (i.e., tail fattening in preparation for prolonged torpor or hibernation), was reported for gray mouse lemurs in dry deciduous western forests and M. rufus (brown mouse lemurs) from montane rainforests (Atsalis 1999; Schmid 2000). Radiotracking observations and body-mass data from M. lehilahytsara (Goodman’s mouse lemurs, referred to as M. rufus in the original study) at Mantadia, an eastern rainforest, showed that a portion of the mouse lemur population remained inactive during the winter (Randrianambinina et al. 2003). Mouse lemurs that were heavier were more likely to experience seasonal torpor, confirming the association between body mass and hibernation. At Tsinjoarivo forest, a high-elevation rainforest in central-eastern Madagascar, several Goodman’s mouse lemurs captured at the end of the reproductive season (February–March) also showed significant tail fattening, so prolonged torpor/hibernation was suspected at this site (Blanco 2010). More recently, studies conducted under semi-natural conditions at the Zoo Zürich, Switzerland, showed that most Goodman’s mouse lemurs stopped visiting feeding stations during the winter and were, presumably, hibernating (Karanewsky et al. 2015).
Goodman’s mouse lemurs are considered highland specialists. This species is regularly exposed to cold temperatures during the winter/dry season, and is therefore the best candidate to employ hibernation in the wild (Radespiel et al. 2012). Torpor use can be more energetically profitable in colder habitats because it allows individuals to reduce body temperature, and consequently metabolic demands, to a greater degree.
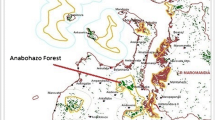
Here we present evidence of prolonged torpor in Goodman’s mouse lemurs at Ankafobe Forest, a tiny forest fragment enclaved in the high plateau of central Madagascar. Ankafobe seems particularly relevant to the study of torpor/hibernation in mouse lemurs because, unlike other lower-elevation rainforests harboring Goodman’s mouse lemurs (such as Mantadia), absolute temperature minima can drop significantly on occasion, increasing the risks of freezing. Thus, selection for buffered environments (e.g., sleeping sites and/or hibernacula) may be strong at this site. In this preliminary survey, we begin to answer basic ecological questions such as the prevalence of hibernation among mouse lemurs at Ankafobe and the preferences for particular hibernacula.
Methods
Study site
We used capture/mark/recapture techniques between February 5 and 13, 2016 at a forest fragment in Ankafobe, central Madagascar (S18°06′23.1″ E47°11′13.2″, 1472 m). Climate profiles from a nearby site (Ambohitantely) characterize it as subhumid, with the highest average temperatures observed during November and the lowest in August (Birkinshaw et al. 2009). At Ankafobe, temperature minima can drop below 0 °C on rare occasions (pers. obs.).
Sherman traps were set at ~1.5 m high along a single trail, in pairs, baited with pieces of fresh banana. Traps were set at 16:00 and checked at 5:00 the following morning. We set up to 29 Sherman traps daily for 9 nights, resulting in a total of 243 trap nights. Captured mouse lemurs were brought back to the campsite, marked with a microchip, weighed, measured, and fitted with small transmitters (1.5 g, Advance Telemetry Systems, Isanti, MN, USA). We employed telemetry to locate sleeping sites and monitor nocturnal activity. Between February 6 and March 10, 2016 we conducted sleeping site monitoring daily. During a second short expedition to the site, from May 16–20, 2016, we monitored sleeping sites four times a day to document sleeping site occupancy during day and night.
Study animals
A total of six mouse lemurs were captured and radiocollared in February. Two of the collars were retrieved from mouse lemurs at the beginning of March. During the May expedition, we detected signals from three individuals. We suspected hibernation expression in one of the radiocollared mouse lemurs based on: (a) clear evidence of body and tail fattening during the February capture, (b) absence of nocturnal activity for two nights during the May expedition, and (c) a change to a more thermally stable (buffered) sleeping site, a tree hole as opposed to a nest. In order to confirm hibernation, we proceeded to retrieve the radiocollared individual from the sleeping site location. Retrieval procedures are described below. A total of three individuals were found inside the tree hole. All three mouse lemurs were brought back to the campsite and weighed. Once fully aroused from hibernation, they were released at the capture location around 19:00.
Results
Captured mouse lemurs
Despite our small sample size, captured mouse lemurs displayed a range of body masses and tail fattening conditions. Table 1 shows body mass and tail base circumference measurements. The individual “Bet” was the heaviest, displaying significant tail fattening in early February (Fig. 1). Reproductive observations showed she was an adult female, though there was no evidence of recent lactation. A second adult female captured the same day displayed no tail fattening but swollen nipples, suggestive of active lactation. A third female, estimated to be around a year old, showed no evidence of reproductive activity and almost no tail fattening at the time of capture (Table 1).

Adult female “Bet,” showing tail fattening
Sleeping site monitoring
Between February 6 and March 10, we documented a minimum of 11 and a maximum of 19 sleeping site observations per mouse lemur (Table 2). During this period, all six radiocollared mouse lemurs occupied nest-like structures as sleeping sites (Table 3). During May 16–18, we observed that one of the three radiocollared individuals (the adult female) was using a tree hole, whereas the two remaining males were occupying nest-like structures (Table 4). The individual female occupying the tree hole remained at this site during all telemetry checks, whereas two individual males occupying nests were absent from their sleeping sites during at least a portion of the night, i.e., the active phase. In order to confirm that the tree hole was used as a hibernaculum, we proceeded to retrieve the individual from this location.
Retrieval procedure and description of the tree hole
The sleeping site/hibernaculum of individual “Bet” was identified as a cavity in an Aphloia theiformis tree, about 9 m high. We observed several tree holes along the trunk but the radio signal was strongest when an antenna was directed to one specific hole, located about 5 m high. The external aperture of the hole was about 4 cm wide and 5 cm long. Due to its small size, the hole had to be widened before we could manually retrieve the radiocollared individual. This procedure took about 40 min, during which no vocalizations were heard from within the tree cavity. The first visible individual was a mouse lemur wearing a collar (later identified as “Bet”). Upon removal of this mouse lemur, two additional individuals were found. All three individuals were unresponsive when removed from the tree hole and cold to the touch. Body temperature was too low to be detected by a standard digital thermometer. Inspection of the hole after the removal of the three individuals showed humus-like soil and Aphloia theiformis leaves (Fig. 2). The internal dimensions of the hole were approximately 42.5 cm (height) by 6.5 cm (depth). The enlarged hole was later boarded with wood to close up the entrance to approximate the original entrance size.

Hole inside tree used as a hibernaculum. Note the accumulation of humus-like soil and leaves
Discussion
Goodman’s mouse lemurs may be better characterized as highland specialists rather than rainforest mouse lemurs (Radespiel et al. 2012). By virtue of occupying higher-elevation habitats, Goodman’s mouse lemurs are consistently exposed to low ambient temperatures during the winter. Exposure to cold increases the energetic demands associated with maintaining a high body temperature (i.e., remaining normothermic); thus, high-plateau habitats should favor the expression of hibernation. Heterothermic responses can be predicted on the basis of body mass and habitat characteristics, such as temperature minima and degree of resource seasonality (Schülke and Ostner 2007); however, more ecophysiological studies are warranted to test these predictions. The paucity of ecophysiological data on mouse lemurs stems in part from the technical difficulties involved in fitting individuals with light radiocollars or devices (weighing less than 5% of their body mass) that can measure temperature and, simultaneously, provide a radio signal to detect the individual’s location. Thus far, with rare exceptions (Schmid 2001, for torpor use), researchers have mainly confirmed that mouse lemurs hibernate without pinpointing hibernation locations (Schmid and Ganzhorn 2009; Kobbe et al. 2011). Here, we have identified a potential hibernaculum but had to disturb the site in order to confirm that the subject was heterothermic.
Future investigations into torpor/hibernation in mouse lemurs should provide not only an increased understanding of their ecological flexibility and resilience to habitat degradation or fragmentation, but could also shed light on mouse lemur biogeographic history and the role that hibernation may have played in the diversification and radiation of this group across Madagascar. For example, torpor and/or hibernation strategies may have favored the dispersion of mouse lemurs across the highlands of Madagascar, which may have originally comprised a mosaic of scrublands, grasslands, and forest fragments like Ankafobe (Yoder et al. 2016). Moreover, studies of mouse lemur torpor/hibernation should be carried out in tandem with those on the dwarf lemurs (Cheirogaleus), a lemur group closely related to mouse lemurs. Dwarf lemurs occupy many of the same environments as do mouse lemurs, but they are obligatory hibernators, are larger in size (about ten times bigger), and are more ecologically susceptible to disturbance (i.e., they are primarily fruit eaters).
We observed intrapopulation plasticity in the expression of hibernation at Ankafobe. Males captured in February showed no evidence of tail fattening, and remained active in May. Among female mouse lemurs, there was a range of behaviors. Female “Ja” showed evidence of recent lactation and no fat deposits, whereas female “Bet” displayed tail fattening but no evidence of recent reproductive activity. This is consistent with the argument that reproduction seems to be incompatible with hibernation (or fattening to sustain hibernation). Variability in the expression of polyestry (more than one reproductive opportunity per season) and weaning profiles likely influences fattening capacity and eventually the timing of hibernation in females. Though resource availability is severely depressed during the winter time, the inactivity of a portion of the mouse lemur population may ameliorate intraspecific competition for the remainder of the active population.
Our observations highlight the incredible behavioral flexibility of mouse lemurs from the same population exposed to identical ecological/climatic conditions, but more detailed ecological studies could profitably address ecological flexibility in relation to population dynamics. Future studies should also endeavor to assess differential life-history strategies in populations of mouse lemurs under a variety of environmental conditions. Our findings are only the first tiny step towards those goals. At a minimum, with our study, we extend the previously known range of hibernation for mouse lemurs to one of the coldest habitats in Madagascar.
References
Atsalis S (1999) Diet of the brown mouse lemur (Microcebus rufus) in Ranomafana National Park, Madagascar. Int J Primatol 20:193–229
Birkinshaw C, Andrianjafy M, Rasolofonirina JJ (2009) Survival and growth of seedlings of 19 native tree and shrub species planted in degraded forest as part of a forest restoration project in Madagascar’s highlands. Madag Conserv Dev 4:128–131
Blanco MB (2010) Reproductive biology of mouse and dwarf lemurs of eastern Madagascar, with an emphasis on brown mouse lemurs (Microcebus rufus) at Ranomafana National Park, a southeastern rainforest. PhD thesis. University of Massachusetts, Amherst, p 141
Hotaling S, Foley ME, Lawrence NM, Bocanegra J, Blanco MB, Rasoloarison R, Kappeler PM, Barrett MA, Yoder AD, Weisrock DW (2016) Species discovery and validation in a cryptic radiation of endangered primates: coalescent-based species delimitation in Madagascar’s mouse lemurs. Mol Ecol 25:2029–2045. doi:10.1111/mec.13604
Karanewsky CJ, Bauert MR, Wright PC (2015) Effects of sex and age on heterothermy in Goodman’s mouse lemur (Microcebus lehilahytsara). Int J Primatol 36:987–998
Kobbe S, Ganzhorn JU, Dausmann KH (2011) Extreme individual flexibility of heterothermy in free-ranging Malagasy mouse lemurs (Microcebus griseorufus). J Comp Physiol B 181:165–173
Ortmann S, Heldmaier G, Schmid J, Ganzhorn JU (1997) Spontaneous daily torpor in Malagasy mouse lemurs. Naturwissenschaften 84:28–32
Radespiel U, Ratsimbazafy JH, Rasoloharijaona S, Raveloson H, Andriaholinirina N, Rakotondranvony R, Randrianarison RM, Randrianambinina B (2012) First indications of a highland specialist among mouse lemurs (Microcebus spp.) and evidence for a new mouse lemur species from eastern Madagascar. Primates 53:157–170
Randrianambinina B, Rakotondravony D, Radespiel U, Zimmermann E (2003) Seasonal changes in general activity, body mass and reproduction of two small nocturnal primates: a comparison of the golden brown mouse lemur (Microcebus revelobensis) in northwestern Madagascar and the brown mouse lemur (Microcebus rufus) in eastern Madagascar. Primates 44:321–331
Schmid J (2000) Torpor in the tropics: the case of the gray mouse lemur (Microcebus murinus). Basic Appl Ecol 1:133–139
Schmid J (2001) Daily torpor in free-ranging gray mouse lemurs (Microcebus murinus) in Madagascar. Int J Primatol 22:1021–1031
Schmid J, Ganzhorn JU (2009) Optional strategies for reduced metabolism in gray mouse lemurs. Naturwissenschaften 96:737–741
Schülke O, Ostner J (2007) Physiological ecology of cheirogaleid primates: a variation in hibernation and torpor. Acta Ethol 10:13–21
Thorén S, Quietzsch F, Schwochow D, Sehen L, Meusel C, Meares K, Radespiel U (2011) Seasonal changes in feeding ecology and activity patterns of two sympatric mouse lemur species, the gray mouse lemur (Microcebus murinus) and the golden-brown mouse lemur (M. ravelobensis), in northwestern Madagascar. Int J Primatol 32:566–586
Yoder AD, Campbell CR, Blanco MB, dos Reis M, Ganzhorn JU, Goodman SM, Hunnicutt KE, Larsen PA, Kappeler PM, Rasoloarison RM, Ralison JM, Swofford D, Weisrock DW (2016) Geogenetic patterns in mouse lemurs (genus Microcebus) reveal the ghosts of Madagascar’s forest past. Proc Nat Acad Sci 113:8049–8056
Acknowledgements
We would like to especially thank Chris Birkinshaw for assisting with plant identification, Jeannie Raharimampionona for her logistical help, and Lydia Greene for editing an earlier version of this paper. We are very appreciative of the comments/suggestions made by Dr. Ute Radespiel about a previous draft. Thanks also to the Associate Editor for comments that helped to improve the manuscript. Financial support for MBB was provided by the CI/Primate Action Fund. This is Duke Lemur Center publication number #1333.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Blanco, M.B., Andriantsalohimisantatra, A.A., Rivoharison, T.V. et al. Evidence of prolonged torpor in Goodman’s mouse lemurs at Ankafobe forest, central Madagascar. Primates 58, 31–37 (2017). https://doi.org/10.1007/s10329-016-0586-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-016-0586-3