
Abstract
Wedelia trilobata (L.) Hitchc., an ornamental groundcover plant introduced to areas around the world from Central America, has become invasive in many regions. To increase understanding of its geographic distribution and potential extent of spread, two presence-only niche-based modeling approaches (Maxent and GARP) were employed to create models based on occurrence records from its: (1) native range only and (2) full range (native and invasive). Models were then projected globally to identify areas vulnerable to W. trilobata invasion. W. trilobata prefers hot and humid environments and can occur in areas with different environmental conditions than experienced in its native range. Based on native and full occurrence points, GARP and Maxent models produced consistent distributional maps of W. trilobata, although Maxent model results were more conservative. When used to estimate the global invasive distribution of the species, both modeling approaches projected the species to occur in Africa. The GARP full model succeeded in predicting the known occurrences in Australia, while the other models failed to identify favorable habitats in this region. Given the rapid spread of W. trilobata and the serious risk of this species poses to local ecosystems, practical strategies to prevent the establishment and expansion of this species should be sought.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A large proportion of the world’s introduced ornamental plants have become invasive in areas where they were purposely introduced. The intentional import of species for horticultural, landscape, or agricultural purposes contributes most to the presence of alien floras in many regions. For instance, 52 % of naturalized alien plant species in Europe were introduced for ornamental or horticultural purposes (Lambdon et al. 2008), and 82 % of invasive woody plants in the United States were used by the landscaping sector (Reichard and White 2001). In Germany, 50 % of the alien flora consist of deliberately introduced species, and more than half of these are ornamentals (Kühn and Klotz 2003). The deleterious effects of invasive ornamental plants on economies and biodiversity of natural areas have raised serious concerns in recent years.
Wedelia trilobata (L.) Hitchc. [syn. Sphagneticola trilobata (L.) Pruski] (creeping oxeye) is a creeping, mat-forming perennial herb native to Central America that has invaded many areas in the tropics and subtropics after its introduction as an ornamental groundcover (Hossain and Hassan 2005; Thaman 1999). The International Union for Conservation of Nature (IUCN) lists W. trilobata in its 100 of the world’s worst invasive alien species (Lowe et al. 2000), and the Florida Exotic Plant Pest Council classifies it as a category II invader (FLEPPC 2014). W. trilobata is reported to dominate agricultural lands, roadsides, abandoned urban lands and other disturbed sites. It also naturalizes and becomes invasive along streams, canals, borders of mangroves and coastal strands (Thaman 1999). It spreads vegetatively, and once established forms a dense ground cover that crowds out or prevents other plants from regenerating including native species (Csurhes and Edwards 1998; Wagner et al. 1990). Knowledge about the potential geographic distribution of this aggressive invader can provide better guidance as to which regions are at risk and mobilize planning to prevent its colonization. Numerous surveys and reports detail areas where W. trilobata was introduced and subsequently colonized. These data provide valuable insight into the environmental conditions that favor its establishment and spread in new areas (Batianoff and Franks 1997; Koheil 2000; McConnell and Muniappan 1991). However, more detailed information (e.g. predictive ability of distribution models, the effect of species occurrence dataset on range estimates, etc.) is needed to improve our understanding of the spatial distribution and potential range expansion of W. trilobata into novel regions.
Ecological niche modeling techniques establish mathematical functions that link potential predictor variables to available occurrence information of a species. These models have been developed and widely used to predict the likely distribution of introduced species based on climatic and edaphic constraints (Elith et al. 2006; Guisan and Thuiller 2005). Recent work has also focused on assessing the underlying assumptions, inherent simplifications, critical limitations and the reliability of these modeling techniques (Rodda et al. 2011; Sinclair et al. 2010; Terribile and Diniz-Filho 2010). Maximum entropy (Maxent; Phillips et al. 2006) and genetic algorithm for rule set production (GARP; Stockwell and Noble 1992), two of the most commonly used presence data-only niche-based modeling methods, have been used for predicting spatial distributions of various species at different scales (e.g. Larson et al. 2010). The two algorithms differ in their rationales and procedures, leading to arguable results regarding their recent performance. For instance, Maxent was more successful than GARP in extrapolating results to new regions (Heikkinen et al. 2012), and showed better predictive accuracy than GARP and other methods (Elith et al. 2006; Hernandez et al. 2006). Townsend Peterson et al. (2007) demonstrated that GARP was more successful in predicting species distributions in broad, unsampled regions than Maxent, as evidenced by the percentage of area a target species was predicted to be present in. When predicting invasion ranges, one important concern is the information (native, invaded or full ranges) used for developing the niche-based models. Models based on data from the native range assume the same environmental factors determine the distribution of the species in the invaded range. However, this assumption may not adequately reflect the distribution of an invasive species in a novel region (Estrada-Peña et al. 2007). Combining native and introduced distribution records in models may be most insightful (Welk 2004), but may not consistently improve model projections (Thompson et al. 2011). Nonetheless, the extent to which the occurrence records represent the environmental space occupied by a target species should be considered when generating these models.
As one of the most commonly introduced ornamental plants, W. trilobata has spread across vast regions of the world. It is becoming widely naturalized in South Africa, the southeastern USA (e.g. Florida and Louisiana), tropical Asia (e.g. Indonesia, Papua New Guinea, and southern China), northern and eastern Australia and many Pacific islands (e.g. American Samoa, the Cook Islands, and Fiji), where climate characteristics may differ from those encountered in the native range. Hence, this species is an ideal candidate for investigating variability in predictions using different models and range occurrence data sets. We developed models for W. trilobata using the Maxent and GARP modeling techniques based on native and full range (including native and invaded ranges) occurrence records. The objectives of the research were to (1) test the predictive capability of the two modeling methods, (2) determine the most effective method for anticipating the species’ global potential distribution, and (3) assess the contribution of the various environmental profiles to model prediction.
Materials and methods
Species occurrence data
Presence records of W. trilobata were obtained from the following online herbaria databases: the global biodiversity information facility (GBIF; http://www.gbif.org), Tropicos (http://www.tropicos.org/), Pacific Island ecosystems at risk (PIER) and the University of South Florida Herbarium (http://www.plantatlas.usf.edu/isb/herbarium.htm), as well as Chinese Virtual Herbarium databases, China Species Information System and National Standard Integration of Species Information for teaching and resources sharing platform. Results from other published scientific research literature and reports of field surveys were also included. Records collected during 1950–2000 were used for model building. The dataset was checked in the DIVA-GIS software (Hijmans et al. 2002) and records with obvious geocoding errors were discarded. Duplicate records were removed manually. The resulting dataset was then overlaid with a 10 km grid and one record was randomly selected from each cell, thereby 861 documented global presence records were obtained for constructing the models. Of these, 528 records were collected from the native ranges of W. trilobata, i.e. Mexico, Central America (Costa Rica, Panama, Nicaragua, Honduras, Belize, and Guatemala), Caribbean and northern South America (Guyana, Venezuela, Brazil, Bolivia, Colombia, Ecuador and Peru). The remaining 333 records were collected from invaded ranges (North America, Australia and New Zealand, Western Europe, Eastern and Southeast Asia), including 190 occurrence points from China. W. trilobata was reported present in West Africa (Liogier and Martorell 2000), however, documented records and museum specimens from the area are very limited and therefore no occurrence record from West Africa was used in the study.
Selection of environmental variables
Climate information derived from the WorldClim 1.4 database at a spatial resolution of 5 arc-min was used to develop the species distribution model (Hijmans et al. 2005). The input data were selected from a total of 48 climate variables that describe monthly total precipitation and mean, minimum and maximum monthly temperature, along with 19 other bioclimatic variables collected from 1950 to 2000. To reduce high collinearity and minimize model overfitting, pairwise correlation analyses were performed first. Variables showing a correlation higher than 0.90 were considered redundant, whilst low-correlation variables were employed to construct the niche models. The subsets of temperature and precipitation redundant variables were then used to perform a principal component analysis (PCA) separately. For each of the PCA factors, variables with the highest (>0.9) factor loadings, measurements for the correlations between the original variables and the factor axes, were selected. This resulted in a final environmental dataset containing 31 temperature and 12 precipitation variables. Grid layers of these variables were clipped with ArcGIS 10.0 (ESRI, Redlands, California, USA) in the assumed native range and the entire invaded range, including China, to make projections in these regions.
Environmental space comparison
Principal component analysis on selected environmental variables was performed to compare the position of occurrences from the native ranges and invaded range in China in multi-dimensional space, following Broennimann et al. (2007). The convex hulls encompassing 95 % of occurrence clouds in the two ranges were identified visually in the PCA bi-plots. Significance differences between the two ranges were further assessed by a between-class analysis and 99 Monte Carlo randomization tests (Romesburg 1985). Differences in the mean values of environmental variables from the native ranges and invaded range in China were tested on individual variables, using Welch’s t test and Levene’s test for homogeneity of variance.
Model development
Maxent and GARP methods were applied to define W. trilobata climatic exigencies as a function of related environmental variables. The modeling process consisted of three steps: (1) predictive models were built separately based on point occurrence data from: (a) native range only and (b) full range (native and invasive) using Maxent and GARP approaches; (2) all models were projected into the assumed native range and China for evaluation of invasion distributions. Environmental variations between the native ranges and invaded range in China were investigated; and (3) models were then projected at a global scale to identify areas putatively susceptible to W. trilobata invasion.
Maxent
Maxent is a machine learning algorithm based on the maximum entropy principle. It assesses the probability distribution of a species by estimating the probability distribution of maximum entropy (Phillips and Dudík 2008). In this study, Maxent software version 3.3.3 k (Princeton University; Princeton, NJ, USA) was used to predict the probability of W. trilobata occurrence and to map its potential spatial distribution. Models were trained using a randomly selected 75 % of occurrence records and then tested on the remaining 25 %. For each training partition, 10 replicates were produced and results averaged. Models were run with default features for the convergence threshold (10−5) and a maximum of 500 iterations. Background points were randomly selected within the area enclosed by a minimum-sized convex polygon that contains all native range or invaded range records. Hinge feature types were used to make simpler and more succinct approximations of the true species response to the environment (Phillips 2008), and a jackknife procedure was used to limit the number of environmental layers to only those layers that affected the distribution of the species. A fade by clamping function was used to remove predicted areas where clamping occurred. The outputs were generated in the form of continuous cumulative probabilities ranging from 0 to 100, with higher values indicating higher probabilities of the areas being suitable habitats for W. trilobata (Elith et al. 2006).
GARP
The GARP model predicts environmental niches of species by identifying non-random relationships between environmental characteristics of known presence localities compared with the entire study region (Mau-Crimmins et al. 2006). In our study, the Desktop GARP application version 1.16 was used as a second approach for predicting the distribution of W. trilobata. The same subsets to build and validate Maxent models were used in the GARP model for comparing the two methods. For each model run, the best-subsets selection procedure (Anderson et al. 2003) was used with a convergence limit of 0.01, maximum number of iterations of 1,000, and 20 runs per model. Based on omission and commission errors (Anderson et al. 2003), the best 10 models were identified and summed to create a final output grid of model agreement, ranging from 0 (areas where none of the 10 models predicted the presence of W. trilobata) to 10 (areas where all of the models predicted the presence of W. trilobata). The rule sets from these 10 models were projected onto the environmental layers for the specified area to generate predictions of W. trilobata distribution.
Model evaluation
A re-sampling function in DIVA-GIS was applied to generate five randomly generated sub-datasets by bootstrapping from the original occurrence record dataset. Each sub-dataset comprised 75 % of the species records as training data. Additionally, pseudo-absence points were randomly generated with an equal number of occurrence records for each study range in each sub-dataset. This approach enabled statistical evaluation of replicate model running in DIVA-GIS to objectively compare the modeling performance of distinct algorithms (Giovanelli et al. 2010). The minimum presence threshold was selected to generate a binary map depicting the predicted area for each model. This threshold equals the minimum model prediction area whilst maintains zero omission error in the training data set (Pearson et al. 2007), and has been used for presence-only models, especially for Maxent and GARP methods producing different outputs with very different frequency distributions (De Meyer et al. 2010).
Null models were created and used to test the significance of both Maxent and GARP models (Raes and ter Steege 2007). For each prediction model of W. trilobata, 99 null-distributions of random points in the study area were generated. The number of random points per distribution was equal to the actual number of presence points. Models were run in Maxent by relating the null-distributions to the environmental layers for calculation of the average AUC. The AUC value of each randomly generated model was then compared with the AUC of the models generated using the actual distribution data in a one-tailed test (α = 0.05). If the AUC of W. trilobata models was significantly higher than the AUC of randomly generated models, the species distribution model performs significantly better than expected by chance (Raes and ter Steege 2007). Model performances were evaluated using an adaptation of the ROC curve approach by plotting omission on an inverse scale (sensitivity) along the y-axis versus proportion of area predicted present (an estimator of 1-specificity) on the x-axis (Peterson et al. 2008; Phillips et al. 2006).
Results
Importance of environmental variables
There were 12 and 15 variables with contributions greater than 1 % identified to be important in creating native and full model fit for W. trilobata, respectively (Table 1). Of these environmental variables, average monthly precipitation in September (prec9) and June (prec6) contributed most for the native model (30.82 %) and full model (33.33 %), respectively. Elevation (alt), average monthly precipitation in October (prec10), prec6, average maximum temperature in November (tmax11) and average minimum temperature in April (tmin4) were important variables for both models. When observing the individual importance of each variable to the model, the variables that contributed the most alone (i.e. running the model with that variable only) were prec9, prec10 and average maximum temperature in June (tmin6) for the native model. Annual precipitation (bio12), precipitation of wettest quarter (bio16) and prec6 were the three variables that contribute most alone to the full model. All these variables had generally limited redundant information (model gain varied considerably when the variable was excluded, see Table 1) and therefore their contributions failed to show large differences when projected at a global scale. For the native model, average monthly maximum temperature in December (tmin12) and average precipitation in January (prec1) had the least redundant information and were the variables that impacted model performance the most if excluded. Similarly, precipitation seasonality (bio15) was the most influential variable to the full model.
Environmental conditions across native and invasive ranges
Principal components analysis (PCA) on environmental layers for the occurrences of W. trilobata performed successfully in niche comparisons of environmental space. The first two axes of the PCA were primarily associated with temperature and moisture and accounted for about 62.6 % of the total variation in environmental niches (Fig. 1). The niche centroids of native and invasive populations differed (i.e. between class inertia ratio of 23 %, P < 0.01). Five thermal and nine moisture variables highly correlated with the first two components were compared in the native range and invaded range in China. Introduced populations of W. trilobata could establish in invaded areas with more variable and lower winter monthly temperatures (e.g. tmean1, tmean2, tmean12), minimum temperature of the coldest month (bio6), and mean temperature of the driest (bio9) and coldest quarter (bio11) than native populations (Fig. 2). In China, this species also occurred in habitats where annual precipitation (bio12), precipitation in the wettest (bio16), average monthly precipitation in October (prec10) and November (prec11) were significantly lower and less variable than for the native ranges, despite the greater precipitation in the warmest quarters (bio18).

Principal component analysis (PCA) for environmental variables associated with occurrence datasets of W. trilobata in the native range (red cross) and China (black filled circle). The ellipses and convex hulls indicate the prevalence (95 and 100 % of sites included) of environments in the study ranges, respectively


The most important thermal variables (a–i) and moisture variables (j–n) for W. trilobata across localities sampled from the native range and China. In box plots, the minimum and maximum values are indicated as short horizontal bars; the 1st and 3rd quartiles are indicated as filled circles and asterisks respectively; the median and means are shown as a broad horizontal bar and unfilled square, respectively. Mean values in the native range were statistically higher than those in China (P < 0.05)
W. trilobata distribution in the native range
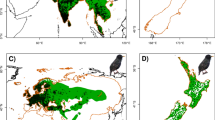
Predictions from the GARP and Maxent models based on separate random testing occurrences were significantly (P < 0.05) better than random expectations. The prediction of environmental suitability between native regions and invaded regions in China varied widely across the models (Table 2; Fig. 3). The GARP full model best predicted known distributions in native regions. The model indicated that most regions of Central America, including Guatemala, Belize, Nicaragua, Honduras, Costa Rica, Panama, and the southwestern and southeastern border areas of Mexico were suitable for W. trilobata and encompassed 74.58 % of its natural range. High environmental suitability was detected for over half of Central and South America. GARP full model results also suggest that W. trilobata may spread further into interior areas of several countries including Brazil, Bolivia, and Venezuela from border regions where the species is present. The GARP native model predicted an extensive convergent range of spread in Brazil and Mexico. Predicted suitable habitats were restricted in Central American regions and sizable contiguous areas of Venezuela, Columbia, and Ecuador bordering northwestern Brazil (Fig. 3).


Occurrence records of W. trilobata collected in a native range; f China; k global range. Predicted geographic distribution of W. trilobata using GARP models in native range (b, c), China (g, h) and global range (l, m) based on native and full range occurrences respectively were provided, results using Maxent models in native range (d, e), China (i, j) and global range (n, o) based on native and full range occurrences were also included
The Maxent model predictions differed substantially from the GARP model predictions (Fig. 3). Compared with GARP models, the Maxent native models predicted a narrower distribution of W. trilobata that may extend from the southern border areas of Mexico to Central America, and connecting with northwestern regions of South America. This species may also be less likely to invade large areas of interior Brazil. The Maxent native model also suggested that suitable environmental conditions may occur in some small areas located in southeastern Mexico, northeastern borders of Belize, Honduras, Costa Rica and Panama. Suitable habitats may also span into western border areas of Columbia. The Maxent full model identified only 0.02 % of the natural range of W. trilobata as suitable habitat, largely in the western border regions of Columbia (Table 2; Fig. 3).
W. trilobata distribution in China
The GARP full model performed well in predicting high environmental suitability in most parts of coastal southern China, including Hong Kong and the tip of Taiwan (Fig. 3). Regions of southeastern Zhejiang, Jiangxi, Hunan, southern Guizhou, Yunnan bordering on Fujian, Guangdong and Guangxi were also predicted to be environmentally suitable for W. trilobata. Although not currently colonized, the contiguous areas between southern Chongqin and southeastern Sichuan were potentially suitable for establishment of this invasive species. Compared with the full model, the GARP native model predicted the occurrence of W. trilobata in more regions along the coast of southeast China and detected suitable areas from these regions. However, the model failed to identify suitable regions extending to the North. The potential suitable area estimated by the GARP full model was 3.5 times that produced by the native model.
The Maxent full model predicted most known areas invaded by W. trilobata in China (Fig. 3). However, the model predicted small and scattered suitable areas consistent with the GARP full model, except for the Guizhou and Yunnan regions. The Maxent native model predicted extensive areas of China invaded by W. trilobata including northeastern regions. However, much smaller areas were identified by the Maxent models as potentially suitable for W. trilobata, confined mostly in the eastern and western regions of Hainan and Taiwan.
Global distribution of W. trilobata
Occurrences of W. trilobata in China were used to assess the predictive ability of the models to detect suitable areas for the species globally. Both GARP and Maxent model predictions were significantly better than expected under null models (P < 0.0001), indicating a reliable ability of the two approaches to predict the potential distribution of W. trilobata worldwide. Based on the full occurrence dataset, the two models performed better than models based only on native range points (Fig. 3). The GARP full model performed better at lower omission values while performance of the Maxent full model was better at middle and high level omission values (Fig. 4).

Accumulation of predictive ability against proportion of area (China) predicted using GARP and Maxent models. Filled and unfilled squares represent GARP and Maxent full models, respectively. Filled and unfilled circles represent GARP and Maxent native models, respectively
Global projections of the GARP and Maxent models indicated three geographically separate regions, namely the northern part of South America, mid-western Africa, and south and southeast Asia. The predicted distributional maps of W. trilobata were generally consistent with current occurrence information from its native and non-native ranges. All the models identified various ranges vulnerable to colonization by W. trilobata in mid-western Africa, located around the Equatorial rain forest belt and East African coastal regions. The GARP full model predicted high suitability areas farther removed from the coast and areas extending more broadly southwards. This model also predicted high suitability in tropical South America and some bordering countries of Central and North America including Panama, Guatemala, Cuba, Dominican Republic and Mexico, a large southern part of Africa including Namibia, Angola, Zambia, Tanzania, Zaire, Congo, Uganda and West Tropical Africa (Guinea; Sierra Leone). Also predicted to have high suitability areas were northeastern Australia and southeastern Asia (especially Thailand, Cambodia, Vietnam, Malaysia and Indonesia), and some Pacific Island countries and territories (e.g. Fiji, Papua New Guinea, Federated States of Micronesia, New Caledonia). The main region where the GARP full model indicated broader potential distributional areas than the other models was in northeastern Australia. Compared with the full model, the GARP native model predicted distinct smaller range areas, especially in tropical South America. The model identified limited high suitability areas near the southwestern Equatorial rain forest belt, while detecting potential distribution areas north of the Equator (Fig. 3). The Maxent model projections yielded similar but more conservative predictions.
Discussion
Despite not including occurrence records from Africa for model generation, distributional ranges were successfully detected using the GARP and Maxent approaches. Observations and reports indicate that W. trilobata has been cultivated and become naturalized in parts of Africa including Benin, Congo Democratic Republic, Guinea, Mayotte, Sierra Leone, South Africa, and Zimbabwe (Hyde et al. 2014). In Zimbabwe, introduced W. trilobata plants have escaped from gardens and formed thick ground covers along the borders of plantations, roadsides and other disturbed areas (Henderson 2001). This species was observed thriving in high rainfall areas in the Eastern highlands and also in urban areas like Harare, which is characterized by a humid-subtropical mild summer climate. This climate is usually found in the highlands of some tropical countries (Köppen–Geiger classification: Cwb) (Kottek et al. 2006) including Mexico, Peru, and Bolivia. These climatic conditions are similar to conditions in the native range of W. trilobata and made it possible for this species to establish in Africa. Both the GARP and Maxent models make successful use of widely available environmental data to identify suitable regions for the potential establishment of a species. These areas of suitability cannot be derived solely from the original occurrence dataset. Such regions of model ‘over-prediction’ having the potential to identify unknown distributional areas and unknown species have been demonstrated (e.g. Raxworthy et al. 2003). Additional research should focus on finding more effective ways (e.g. improve the transferability of the model by occurrence thinning or predictor selection) for projecting and interpreting the potential distribution of W. trilobata.
Performance of models
Using the GARP and Maxent models and based on native and full occurrence data points in this study resulted in a consistent distributional map for W. trilobata, although Maxent model findings were more conservative. The two modeling methods provided reliable predictions of the geographical extent of W. trilobata invasion using historical museum records which tend to be biased because they do not represent true absence values or guarantee that all areas have been adequately sampled (e.g. Anderson et al. 2003; Stockwell and Peterson 2003). When projected geographically to estimate the global distributional potential of the species, the GARP full model succeeded in predicting the known occurrences in Australia, while the Maxent models failed to identify the favorable habitats along the edges of invaded areas (e.g. Australia, Caribbean). This finding suggests that the GARP model based on the full range occurrences points may be able to better cope with spatial bias than Maxent models. As demonstrated, the GARP model does not respond to coordinate spacing, and might perform better in predicting distributions from incomplete coordinate sets (Costa and Schlupp 2010). The greater predictive capability of GARP models has been reported in other studies (Terribile and Diniz-Filho 2010; Townsend Peterson et al. 2007).
Despite most known occurrences falling within predicted areas, models based solely on native occurrences records were less effective in discriminating relative climate suitability. These native models also failed to capture habitats occupied by W. trilobata in Australia. One possible reason is that W. trilobata may occupy an environmental niche that is environmentally different from regions in its native range. Significant differences of the most important environmental variables between native and introduced W. trilobata ranges and results of principal components analysis on environmental layers indicates that this species may occupy distinct environmental niches in its invaded ranges. These differences and the ability of some plant species to rapidly evolve to adapt to a changing climate (i.e. environment) makes it especially challenging to predict the future geographic extent of plant species with models using only native range data (Clements and DiTommaso 2011). In environmental space, the model identifies neither the occupied niche nor the fundamental niche (Hutchinson 1957) since the target species were unlikely to be at equilibrium and occurrence records will not completely reflect the full range of environmental conditions occupied by the species. Therefore, it would be more insightful to combine data from both the native and invaded ranges (e.g. Welk 2004). Because of potential niche shifts in a species such as W. trilobata, models based on data from both the invaded and native ranges may be more appropriate and insightful in determining its geographic range. As demonstrated in our study, models built on the full dataset can effectively predict the future spread of the species in areas that can be potentially invaded (Broennimann and Guisan 2008).
Potential distributions and environments
Both GARP and Maxent results indicated that W. trilobata can occur in areas with different environmental conditions than experienced in its native range. Using the updated Köppen–Geiger climate classification system (Kottek et al. 2006), most of the suitable areas susceptible to invasion by W. trilobata identified by these models fell within the Aw (tropical wet and dry or savanna climate) climate class. Other suitable regions prone to invasion included the Cfa or Cwa (the humid subtropical climate) classifications. The GARP full model also detected high suitability for invasion to some areas in the Af (tropical rainforest climate) and Am (tropical monsoon climate) climate classifications. These results confirm that W. trilobata, a tropical species prefers hot and humid environments. For example, W. trilobata colonized regions with temperate/mesothermal climates with dry winters (i.e. warm average temp. >10 °C, cold average temp. >0 °C, dry winters). Annual precipitation of suitable regions is typically high, although it does not have to be evenly distributed throughout the year. A good example of this suitability preference is southern China, a region generally classified as Cwa based on the Köppen climate classification system, and where W. trilobata is present year-round. In this region, average annual precipitation totals 1,652 mm with rainfall received mostly in the summer months when plant needs are greatest. Approximately 32.7 and 42.3 % of precipitation falls in warmest and wettest quarter of the year, respectively (Fig. 2).
The predicted distribution of W. trilobata provides evidence of the ability of this species to survive under a wide range of environmental conditions across numerous geographical regions. In its native range, W. trilobata occurs in habitats with tropical wet and warm winters. Outside of its native range as in China for example, W. trilobata has expanded into areas that have a wider range of monthly mean temperatures and narrower range of annual precipitation. These findings combined with plant traits that allow this species to adapt to a wide range of environmental conditions including vegetative propagation (Wu et al. 2005), high photosynthetic activity (Song et al. 2010), allelopathic potential (e.g. Nie et al. 2004), and high energy-use efficiency (Song et al. 2009) are the main factors that may explain the success of W. trilobata to invade novel regions of the world. The models used in this study predicted regions of the world that are environmentally suitable for W. trilobata colonization, but they do not identify regions that will necessarily be invaded. The major reason for this difference is that our models did not account for barriers to seed dispersal, local soil or management factors, biotic interactions, and the capacity of species to adapt to new environments (Thuiller et al. 2008). Therefore, a comprehensive assessment of the invasion risk posed by this species indifferent regions of the world would require a more in depth understanding of the way that these barriers affect the population dynamics of the species (Thuiller et al. 2005).
Threat of W. trilobata invasion
Given the rapid spread of W. trilobata and serious risk this species may pose to resident ecosystems, efforts to limit its introduction and establishment in regions outside of its native range should be fully considered. For instance, W. trilobata was first introduced to South China in the 1970s as ornamental and has since become a major invasive weed. This plant is gaining increased popularity because it is an excellent ground cover and reduces soil erosion (Wu et al. 2005). Many Chinese farmers also believe that W. trilobata can be used in traditional medicine (Tsai et al. 2009). The GARP full model predictions showed that W. trilobata has the potential to expand its range along southeastern coastal areas and is able to extend into interior regions of eastern and southwestern China. More than 7 % of the study area was suitable for W. trilobata colonization under current environmental conditions. With development of a national horticultural industry and the fast growing ornamental trade sector in China, it is very likely that W. trilobata will quickly naturalize and invade regions of the country that are projected to experience increased temperatures in the coming decades. The deleterious effect of W. trilobata on native plant populations and biodiversity has raised concern in regions of China at higher risk of infestations because effective control strategies are not available in these areas. As a way forward, it is essential to identify and monitor areas most vulnerable to invasion by this aggressive plant, since early detection and rapid response will be crucial for minimizing economic losses and ecological damage. Practical management strategies are also needed in other regions of the world vulnerable to invasion by W. trilobata to meet conservation and biodiversity goals.
References
Anderson RP, Lew D, Peterson AT (2003) Evaluating predictive models of species’ distributions: criteria for selecting optimal models. Ecol Model 162:211–232
Batianoff GN, Franks AJ (1997) Invasion of sandy beachfronts by ornamental plant species in Queensland. Plant Prot Q 12:180–186
Broennimann O, Guisan A (2008) Predicting current and future biological invasions: both native and invaded ranges matter. Biol Lett 4:585–589
Broennimann O, Treier UA, Müller-Schärer H, Thuiller W, Peterson AT, Guisan A (2007) Evidence of climatic niche shift during biological invasion. Ecol Lett 10:701–709
Clements DR, DiTommaso A (2011) Climate change and weed adaptation: can evolution of invasive plants lead to greater range expansion than forecasted? Weed Res 51:227–240
Costa GC, Schlupp I (2010) Biogeography of the Amazon molly: ecological niche and range limits of an asexual hybrid species. Global Ecol Biogeogr 19:442–451
Csurhes S, Edwards R (1998) National weeds program: potential environmental weeds in Australia—candidate species for preventative control. National Parks and Wildlife Biodiversity Group, Environment Australia, Canberra
De Meyer M, Robertson MP, Mansell MW, Ekesi S, Tsuruta K, Mwaiko W, Vayssières JF, Peterson AT (2010) Ecological niche and potential geographic distribution of the invasive fruit fly Bactrocera invadens (Diptera, Tephritidae). B Entomol Res 100:35–48
Elith J et al (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Estrada-Peña A, Pegram RG, Barré N, Venzal J (2007) Using invaded range data to model the climate suitability for Amblyomma variegatum (Acari: Ixodidae) in the New World. Exp Appl Acarol 41:203–214
FLEPPC (2014) Florida exotic pest plant council invasive plant lists. http://www.fleppc.org/list/list.htm. Accessed 5 Feb 2015
Giovanelli JGR, de Siqueira MF, Haddad CFB, Alexandrino J (2010) Modeling a spatially restricted distribution in the neotropics: how the size of calibration area affects the performance of five presence-only methods. Ecol Model 221:215–224
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8:993–1009
Heikkinen RK, Marmion M, Luoto M (2012) Does the interpolation accuracy of species distribution models come at the expense of transferability? Ecography 35:276–288
Henderson L (2001) Alien weeds and invasive plants. Plant Protection Research Institute Handbook No 12. Plant Protection Research Institute, Agricultural Research Council, South Africa. pp 58
Hernandez PA, Graham CH, Master LL, Albert DL (2006) The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 29:773–785
Hijmans R, Guarino L, Mathur P, Jarvis A (2002) DIVA-GIS: geographic information system for biodiversity research manual. International Potato Center, Lima
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978
Hossain ABME, Hassan MA (2005) Wedelia trilobata (l.) AS Hitchc. (Asteraceae)—a new record for Bangladesh. Bangl J of Plant Taxon 12:63–65
Hutchinson GE (1957) Concluding remarks. Cold Spring Harb Sym 22:415–427
Hyde M, Wursten B, Ballings P, Palgrave MC (2014) Flora of Zimbabwe: species information: Sphagneticola trilobata. http://www.zimbabweflora.co.zw/speciesdata/species.php?species_id=160400. Accessed 10 Nov 2014
Koheil MA (2000) Study of the essential oil of the flower-heads of Wedelia trilobata (L.) Hitch. Al-Azhar J Pharm Sci 26:288–293
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World Map of the Köppen–Geiger climate classification updated. MetZe 15:259–263
Kühn I, Klotz S (2003) The alien flora of Germany-basics from a new German database. In: LE Child, Brock JH, Brundu G, Prach K, Pyšek P, Wade PM, Williamson M (eds) Plant invasions: ecological threats and management solutions. Backhuys, Leiden, pp 89–100
Lambdon et al (2008) Alien flora of Europe: species diversity, temporal trends, geographical patterns and research needs. Preslia 80:101–149
Larson ER, Olden JD, Usio N (2010) Decoupled conservatism of Grinnellian and Eltonian niches in an invasive arthropod. Ecosphere 1:art16
Liogier AH, Martorell LF (2000) Flora of puerto rico and adjacent islands: a systematic synopsis. Ed. de la Univ
Lowe S, Browne M, Boudjelas S, Poorter MD (2000) 100 of the World’s worst invasive alien species: a selection from the global invasive species database. The IUCN Invasive Species Specialist Group (ISSG), pp 12
Mau-Crimmins TM, Schussman HR, Geiger EL (2006) Can the invaded range of a species be predicted sufficiently using only native-range data? Ecol Model 193:736–746
McConnell J, Muniappan R (1991) Introduced ornamental plants that have become weeds on Guam. Micronesia (Suppl 3):47–49
Nie C, Luo S, Zeng R, Mo M, Li H, Lin C (2004) Allelopathic potential of Wedelia trilobata L.: effects on germination, growth and physiological parameters of rice. Acta Agron Sin 30:942–946
Pearson RG, Raxworthy CJ, Nakamura M, Townsend Peterson A (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117
Peterson AT, Papeş M, Soberón J (2008) Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol Model 213:63–72
Phillips SJ (2008) Transferability, sample selection bias and background data in presence-only modelling: a response to Peterson et al. (2007). Ecography 31:272–278
Phillips SJ, Dudík M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Raes N, ter Steege H (2007) A null-model for significance testing of presence-only species distribution models. Ecography 30:727–736
Raxworthy CJ, Martinez-Meyer E, Horning N, Nussbaum RA, Schneider GE, Ortega-Huerta MA, Townsend Peterson A (2003) Predicting distributions of known and unknown reptile species in Madagascar. Nature 426:837–841
Reichard SH, White P (2001) Horticulture as a pathway of invasive plant introductions in the United States: most invasive plants have been introduced for horticultural use by nurseries, botanical gardens, and individuals. Bioscience 51:103–113
Rodda GH, Jarnevich CS, Reed RN (2011) Challenges in identifying sites climatically matched to the native ranges of animal invaders. PLoS One 6:e14670
Romesburg HC (1985) Exploring, confirming, and randomization tests. Comput Geosci 11:19–37
Sinclair SJ, White MD, Newell GR (2010) How useful are species distribution models for managing biodiversity under future climates. Ecol Soc 15:8
Song L, Wu J, Li C, Li F, Peng S, Chen B (2009) Different responses of invasive and native species to elevated CO2 concentration. Acta Oecol 35:128–135
Song L, Chow WS, Sun L, Li C, Peng C (2010) Acclimation of photosystem II to high temperature in two Wedelia species from different geographical origins: implications for biological invasions upon global warming. J Exp Bot 61:4087–4096
Stockwell DRB, Noble IR (1992) Induction of sets of rules from animal distribution data: a robust and informative method of data analysis. Math Comput Simulat 33:385–390
Stockwell D, Peterson AT (2003) Comparison of resolution of methods used in mapping biodiversity patterns from point-occurrence data. Ecol Indic 3:213–221
Terribile LC, Diniz-Filho JAF (2010) How many studies are necessary to compare niche-based models for geographic distributions? Inductive reasoning may fail at the end. Braz J Biol 70:263–269
Thaman RR (1999) Wedelia trilobata: daisy invader of the Pacific Islands. IAS Tech Rep 99. Institute of Applied Science, University of the South Pacific, Suva, Fiji
Thompson GD, Robertson MP, Webber BL, Richardson DM, Le Roux JJ, Wilson JRU (2011) Predicting the subspecific identity of invasive species using distribution models: Acacia saligna as an example. Divers Distrib 17:1001–1014
Thuiller W, Richardson DM, PyŠEk P, Midgley GF, Hughes GO, Rouget M (2005) Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Glob Change Biol 11:2234–2250
Thuiller W et al (2008) Predicting global change impacts on plant species’ distributions: future challenges. Perspect Plant Ecol Evol Syst 9:137–152
Townsend Peterson A, Papeş M, Eaton M (2007) Transferability and model evaluation in ecological niche modeling: a comparison of GARP and Maxent. Ecography 30:550–560
Tsai CH, Lin FM, Yang YC, Lee MT, Cha TL, Wu GJ, Hsieh SC, Hsiao PW (2009) Herbal extract of Wedelia chinensis attenuates androgen receptor activity and orthotopic growth of prostate cancer in nude mice. Clin Cancer Res 15:5435–5444
Wagner WL, Herbst DR, Sohmer SH (1990) Manual of the flowering plants of Hawai’i. University of Hawaii Press and Bishop Museum Press, Honolulu
Welk E (2004) Constraints in range predictions of invasive plant species due to non-equilibrium distribution patterns: purple loosestrife (Lythrum salicaria) in North America. Ecol Model 179:551–567
Wu YQ, Hu YJ, Chen JN (2005) Reproductive characteristics of alien plant Wedelia trilobata. Acta Sci Nat Univ Sunyatseni 44:93–96
Acknowledgments
We are grateful to the Key Laboratory of Ecological Agriculture of the Ministry of Agriculture of China and the Key Laboratory of Agroecology and Rural Environment of Guangdong Regular Higher Education Institutions, South China Agricultural University for fostering research on this topic. This work was supported by the National Natural Science Foundation of China (No. U1131006, 31300405), the Doctoral Fund of the Ministry of Education of China (No. 20124404110009). We thank Rongkui Han and anonymous reviewers for providing helpful comments on earlier versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Qin, Z., Zhang, Je., DiTommaso, A. et al. Predicting invasions of Wedelia trilobata (L.) Hitchc. with Maxent and GARP models. J Plant Res 128, 763–775 (2015). https://doi.org/10.1007/s10265-015-0738-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-015-0738-3