
Abstract
Altered mitochondrial DNA (mtDNA) is the most common denominator to numerous metabolic diseases. The present study sought to investigate the correlation between mtDNA content in lymphocytes and associated clinical risk factors for impaired fasting glucose (IFG). We included 23 healthy control and 42 IFG participants in this cross-sectional study. The measurements of mtDNA content in lymphocytes and pro-inflammatory markers derived from both normal and diseased individuals were quantified. Spearman partial correlation and multivariate statistical analyses were employed to evaluate the association between mtDNA content and other metabolic covariates in IFG. Reduced mtDNA content was observed in the IFG group with microvascular complications than those without complications. The IFG patients with lowest median of mtDNA content had considerably elevated hyperglycemia, insulin resistance and inflammation. The adjusted partial correlation analysis showed that mtDNA content was positively correlated with HDL-cholesterol and IL-10 (P < 0.005 for all). Further, multiple linear regression analyses verified that reduced mtDNA content in lymphocytes was independently associated with HOMA-IR (β = 0.027, P = 0.003), HbA1c (β = 0.652, P = 0.002), HDL-cholesterol (β = − 1.056, P = 0.021), IL-6 (β = 0.423, P = 0.002), IL-10 (β = − 1.234, P = 0.043) and TNF-α (β = 0.542, P < 0.001) after adjustment for confounding factors. Our data show that reduced mtDNA content in lymphocytes was associated with insulin resistance and inflammation in individuals with IFG.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Diabetes is a global healthcare problem affecting human population worldwide. According to International Diabetes Federation (IDF), 415 million people have been diagnosed with diabetes in 2015 and the number is projected to reach 642 million in 2040. Of this global burden, it is estimated that about 90% of diabetes patients are suffered from type 2 diabetes (T2D) with a greater degree of insulin resistance. Current classifications of individuals at risk of developing T2D are mainly based on the established factors such as blood fasting glucose, HbA1c, body mass index (BMI) and age [1]. Notably, recent estimation showed that up to 70% of individuals with prediabetes state progressively develop diabetes in years [2]. On account of health risks, the transitions from the early symptoms of insulin resistance including impaired fasting glucose (IFG) and impaired glucose tolerance (IGT) that precedes T2D may take several years. However, the identification and diagnosis of prediabetes individuals is not commonly employed in the clinical practice.
The proposed mechanisms underpinning the development of these metabolic dysregulations are embedded in the complex networks of genetics and environmental factors. A number of considerable reports have revealed that impaired mitochondrial homeostasis contributes to a broad range of T2D progressions [3]. Several defects in mitochondrial integrity have been found in offspring of type 2 diabetic patients, proving the degree of heritability [4]. Depletion of mitochondrial (mtDNA) content in pancreatic islet [5] and skeletal muscle tissues [6] of rodent and human participants with T2D, respectively, has been reported. In this regard, multiple studies demonstrated that diminished mitochondrial functions in peripheral blood mononuclear cells (PBMCs) and endothelial cells from diabetes patients were coupled with higher level of reactive oxygen species (ROS) [7, 8]. The activations of inflammatory response by multiple stimuli in both lymphocytes [9] and monocytes [10] were mainly modulated by mitochondrial hyperpolarization.
Dysregulated lymphocytes metabolism is among the key contributors to an enhanced susceptibility to infection in a poorly controlled hyperglycemia and early stage of diabetic states [11]. The leukocytes–macrophage lineage could be of specific importance as these peripheral cells play an important role in the chronic low-grade inflammation of insulin resistance and T2D patients. Diminished mitochondrial function of lymphocytes with activation in the cascade of NF-κB inflammatory signaling in type 2 diabetes was observed [11]. Nevertheless, the exact mechanism underlying these detrimental events is still a matter of debate. In light of the ongoing epidemic, future investigation designed to further explore the relationship between mitochondrial function and other metabolic parameters of diabetes-associated complications is of great interest.
Mitochondrial dysfunction is usually attributed by alteration in mtDNA copy numbers, morphology and severe defects in mitochondrial genes and proteins, which are ultimately leading to defective oxidative phosphorylation [12, 13]. In recent years, mtDNA content was extensively used as a surrogate marker of mitochondrial function in a number of peripheral cells and biofluids for studying metabolic disease progressions and mechanisms [13]. Though previous studies have investigated the roles of mtDNA content in T2D and obesity, none have explored the role of mtDNA content in lymphocytes as a candidate factor for prediabetes state such as IFG. Considering these, the aim of the present study was to investigate the association between mtDNA content assessed by quantification of mtDNA copy number in lymphocytes and other clinical risk factors including the circulating level of inflammatory cytokines of IFG patients.
Materials and methods
Study design and participants recruitment
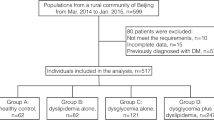
A total of 93 adult participants with newly diagnosed IFG or healthy control were recruited from outpatient clinics of the University Malaya Medical Centre (UMMC), Malaysia, between February 2015 and June 2015. After a subsequent routine health checkup, a total of 65 eligible individuals were included in the study. The participants were divided into two groups: normal healthy group (n = 23) and IFG group (n = 42). The classification and diagnosis of IFG participants with hyperglycemia was based on the American Diabetes Association (ADA) criteria [14], a fasting plasma glucose (FPG) level from 5.6 to 6.9 mmol/L. All diseased participants have not received any hypoglycemic drugs, insulin and diet control. Participants taking statin were also excluded. Patients with a history of malignancy, infectious, liver disease and acute illness were omitted in the study. Healthy control was defined as FPG level from 3.9 to 5.5 mmol/L. Healthy control participants recruited in the study were taking no antioxidant supplementation and had no clinical history of metabolic complications. The study protocol was approved by the institutional medical ethics review board of the UMMC (Reference Number: 201495.95) in compliance with the Declaration of Helsinki of the World Medical Association. All participants underwent the informed consent process.
Collection of blood and urine sample
The collection of 20 mL venous blood samples from all participants following 10–12 h of an overnight fasting was achieved in tubes containing sodium fluoride and ethylenediaminetetraacetic acid (EDTA), EDTA with and without no anticoagulants. Blood samples were collected in the early morning. The samples were routinely centrifuged at 2500×g for 15 min at 4 °C to separate plasma from red blood cells within 1 h of extraction. Plasma samples were aliquoted and immediately stored at − 80 °C freezer until further use. Urine samples were collected from first overnight morning and further processed for biochemical analyses.
Clinical and biochemical assessment
Anthropometric measurements were conducted using a standard method [15]. All participants were invited to come on a prescheduled morning at diabetic clinics after an overnight fasting (> 8 h). The diagnosis of nephropathy for IFG participants was based on the urinary albumin/creatinine ratio in a random urine sample of more than 30 mg/g of creatinine. Funduscopic examination was carried out to diagnose patients with retinopathy. Diseased participants with polyneuropathy were identified based on the sensitivity to light/touch pressure perception using a monofilament [16]. Plasma glucose and HbA1c levels were determined by using the glucose oxidase method and high-performance liquid chromatography (VARIANT™ II and D-10TM Systems, Bio-Rad, USA), respectively. Serum total cholesterol, triglycerides, high-density lipoprotein (HDL) and low-density lipoprotein (LDL) were analyzed using a biochemical auto-analyzer (Roche COBAS INTEGRA 700; Roche Diagnostics, Indianapolis, Indiana). Liver function tests including aspartate transaminase (AST), alanine transaminase (ALT) and gamma-glutamyl transpeptidase (-GGT) were quantified using an auto-analyzer (ARCHITECT c16000 System, Abbott Laboratories, IL, USA). Other clinical variables including height, body weight, BMI and blood pressure (BP) were measured accordingly [15]. A 24-h urine sample was collected for the measurement of creatinine and urinary albumin. Serum fasting insulin concentrations were determined using commercially available enzyme-linked immunosorbent assay kits (R&D Systems) following manufacturer’s instructions. The homeostasis model assessment of insulin resistance (HOMA-IR) was computed using a formula (fasting insulin [µU/mL] × fasting glucose [mmol/L]/22.5).
Measurement of pro-inflammatory cytokines
The quantification of pro-inflammatory cytokines level in the blood plasma was assessed by commercial ELISA kits for human IL-1, IL-6, IL-10 and TNF-α (Trevigen, Helgerman, Gaithersburg, USA) according to the manufacturer’s instruction. The intra-assay and inter-assay coefficients of variation were less than 9 and 10%, respectively. The measurement of these circulating cytokines was performed in triplicate. We observed no significant cross-reactivity and interference in the assays.
Isolation of lymphocytes
The lymphocytes isolation was achieved by using density gradient centrifugation method [17] in Gradisol L (Aqua-Medica, Poland) at 1200×g for 20 min at room temperature. Afterward, the lymphocytes were washed twice with 0.9% NaCl solution and spun at 500×g for 10 min. The resulting lymphocytes suspension was collected and subsequently washed with PBS. The cells were further diluted with PBS to obtain a density of 106 cells/ml. The concentration of lymphocytes was counted using hemocytometer. After isolation, the lymphocytes were stored at −20 °C for further use.
Measurement of mtDNA content
Total genomic DNA was extracted using a commercialized kit (QIAamp DNA Blood Midi Kit; Qiagen, Hilden, Germany) following the manufacturer’s guidelines. The kit was designed to extract both nuclear and mitochondrial DNA (mtDNA) from lymphocytes suspension. The concentration of purified DNA was assessed using a NanoDrop 2000 spectrophotometer Thermo Scientific, Wilmington, DE, USA). We utilized real-time duplex quantitative PCR to quantify the relative mtDNA content, using the same primers that were utilized for the mitochondrial ND1 gene (ND1-F and ND1-R) and the single-copy nuclear gene human globulin (HGB-1 and HGB-2). In brief, the relative quantification of mtDNA content was performed utilizing two pairs of primer sequences as follows: ND1 forward (ND1-F), 5′-CCCTAAAACCCGCCACATCT-3′; ND1 reverse (ND1-R), 5′-GAGCGATGGTGAGAGCTAAGGT-3′; HGB forward (HGB-1), 5′-GTGCACCTGACTCCTGAGGAGA-3′; HGB reverse (HGB-2), 5′-CCTTGATACCAACCTGCCCAG-3′. In the first step, the relative ratio of mtDNA copy number to HGB copy number was computed for each sample from standard curve. The second step involved the normalization of the ratio for each sample used in the study to a calibrator DNA, thus standardizing between different runs and total quantification of relative mtDNA content. A genomic DNA sample from the healthy control was served as a positive control (calibrator DNA), and standard curve was measured for each run to compare results from different independent assays. It was observed that the value of scaling factor varied from 0.83 to 1.05 across different runs. Further, 8 ng of genomic DNA was added into PCR mixtures containing the primers (10 nM each) and 1 × SYBR green mastermix (TaKaRa, Dalian, China) with a total volume of 20 µL. The amplification processes of thermal cycling conditions for both primer pairs were based on the following conditions; 95 °C for 30 s, followed by 35 cycles of 94 °C for 30 s, 58 °C for 30 s and 72 °C for 50 s with signal acquisition. One reference DNA sample was diluted with a threefold increment per dilution to generate a 5-point standard curve. The R2 for each standard curve was found to be > 0.95, with acceptable SD values set at 0.25 (for the Ct values).
Statistical analyses
We used the SPSS software (IBM, Armonk, NY, version 19.0) for database management and statistical analyses. All data are expressed as mean ± standard deviation (SD) for parametric variables. Distribution of data variable was verified by applying the Kolmogorov–Smirnov test. The median and interquartile ranges (IQR) were further used for nonparametric variables. Chi-square test was employed to evaluate the comparison between groups of categorical data (percentage). The means statistical differences between groups were compared and evaluated using the Mann–Whitney test or analysis of variance (ANOVA). To evaluate the relationship between mtDNA content and some clinical risk factors, the IFG group was stratified according to the median level of mtDNA content. A partial correlation analysis after adjustment by age and sexes was used to assess the inter-relationship between variables. Multiple linear regression was executed to normalize the result for the measurement of mtDNA content in lymphocytes. The association between mtDNA content and clinical risk factors in IFG groups was examined by using the multiple logistic regression analyses. P values (< 0.05) were assumed statistically significant.
Results
Clinical characteristics of the participants
We evaluated 65 participants (42 participants with IFG and 23 controls). The baseline characteristics of the study population are summarized in Table 1. Participants in IFG group were older than those of the control group (P < 0.05). Sexes, total cholesterol, LDL-cholesterol, albumin, creatinine, ALT and AST were not significantly differed with the control group. Compared to the normal group, participants with IFG exhibited higher level of hypertension, BMI, fasting glucose, fasting insulin, HbA1c, systolic blood pressure, HOMA-IR, triglycerides, IL-1, IL-6 and TNF-α. However, the levels of HDL-cholesterol and IL-10 were significantly higher in the control group than those of the IFG group. Overall, 22 IFG patients had microvascular complications, 49 patients had kidney disease, 7 patients showed neuropathy and 6 patients presented retinopathy.
Low mtDNA content in lymphocytes was linked to IFG, microvascular complications and high BP
To verify our hypothesis, we compared the level of mtDNA content in lymphocytes with study groups, sexes, microvascular complications and BP. Participants with IFG displayed a reduced trend of mtDNA content (774.78 ± 32.1) compared to the control group (1195.42 ± 63.2) (Fig. 1a, P < 0.05). As shown in Fig. 1b, no statistically significant difference for sexes, men (670.42 ± 32.1) and women (687. 28 ± 27.6), was observed on the level of mtDNA content in lymphocytes among groups (P > 0.05). In addition, the difference among those of the IFG group in the presence and absence of microvascular complications was further evaluated. We observed that those of the IFG participants with microvascular complications were exhibited (Fig. 1c, P < 0.05) with low level of mtDNA content (677.053 ± 36.1) than those of IFG patients without microvascular complications (978.297 ± 65). As depicted in Fig. 1d, participants with high level of BP (systolic) were suffered with reduced mtDNA content (674.43 ± 53, P < 0.05) in lymphocytes compared to the normal BP group (914.11 ± 27). This suggested that the level of mtDNA content in lymphocytes was able to distinguish the groups based on this metabolic parameter.

The level of mtDNA content in lymphocytes among study groups (control and IFG) (a), sexes (male and female) (b), microvascular complications (absence and presence) (c) and BP (normal and high) (d). Normal BP (systolic BP < 130 mm Hg; diastolic BP < 85 mm Hg) and high BP (systolic BP ≥ 130 mm Hg; diastolic BP ≥ 85 mm). Data were expressed as mean ± SD. For continuous variable, differences among groups were computed by ANOVA. **P < 0.05 compared with control group, **P < 0.01 compared to those without microvascular complications and normal BP
Stratification of median mtDNA content in lymphocytes from IFG patients to clinical and inflammatory markers
Our study also revealed that the level of mtDNA content in lymphocytes can be utilized to stratify the IFG group according to the median range. We stratified the IFG group into higher (n = 17) and lowest median (n = 25) of mtDNA content. Based on Table 2, IFG patients with lowest median of mtDNA content in lymphocytes (< 850 per cell) had considerably increased level of fasting glucose, HbA1c and HOMA-IR, signifying the severity of these metabolic complications in the IFG group with lowest median of mtDNA content. However, the IFG group with higher median of mtDNA content was associated with reduced level of fasting glucose, HbA1c and HOMA-IR compared to those group of lowest median of mtDNA content. Moreover, we further explored the correlation between both groups on the relative level of pro-inflammatory cytokines in the IFG participants. We observed that those of the IFG group with reduced mtDNA content in lymphocytes were significantly exhibited with increased level of IL-6 and TNF-α and decreased level of IL-10 than those of IFG group with higher median of mtDNA content. Nevertheless, we found no significant difference in the level of IL-1 in the median range of mtDNA content among the IFG group (Table 2).
The level of mtDNA content in lymphocytes was positively associated with HDL-cholesterol and IL-10
To explore further correlation between mtDNA content in lymphocytes and other clinical risk factors, we performed the Spearman and partial correlation analyses using a number of confounding factors as independent variables. It was demonstrated that mtDNA content was negatively correlated (P < 0.05) with hypertension, BMI, fasting glucose, fasting insulin, HbA1c, HOMA-IR, LDL-cholesterol, triglycerides, IL-6 and TNF-α after adjusted by age and sexes (Table 3). Importantly, we observed the positive relationship (P < 0.05) between mtDNA content in lymphocytes, HDL-cholesterol and IL-10 level. However, no significant different (P > 0.05) was detected in the partial correlation analyses after controlling for age and sexes in the levels of total cholesterol, urinary albumin, ALT, AST and IL-10.
Multiple logistic and linear regression analyses
To obtain clear insight on the role of mtDNA content in lymphocytes in IFG, we computed the multiple logistic regression analyses. Reduced mtDNA content was significantly correlated with IFG after normalizing for several confounding factors including age, sexes, ALT, AST, triglycerides, total cholesterol, LDL-cholesterol, HDL-cholesterol, IL-6, IL-10 and TNF-α (P < 0.001 for all) (Table 4). These results signified that mtDNA content in lymphocytes was linked to IFG in couple with these clinical risk factors and pro-inflammatory cytokines. To functionally validate our calculation on the association between mtDNA content in lymphocytes and several covariates, further multiple analyses using multiple linear regressions were used. It has been shown that mtDNA content in lymphocytes was independently associated with HOMA-IR, HbA1c, HDL-cholesterol, IL-6, IL-10 and TNF-α (P < 0.001 for all) (Table 5).
Discussion
Several studies to date have looked at the association between mtDNA copy numbers and other clinical risk factors from individuals with metabolic syndromes [13]. It is now becoming clear that many metabolic diseases with early distinct clinical symptoms are regulated by mitochondrial signaling in platelets and leukocytes to signify their signals of bioenergetics dysfunction [18]. In this context, studies are essentially needed to clarify the correlation between mtDNA content and other risk factors in the early onset of prediabetes individuals. In view of these considerations, the importance of this study was to provide such understanding on the role of dysregulated mitochondrial metabolism on associated metabolic parameters in prediabetes. In the present study, we observed that decreased level of mtDNA content in lymphocytes derived from IFG individuals was significantly allied to insulin resistance and inflammation after adjustment for several confounding factors. Therefore, we speculated that modulation of mtDNA content in lymphocytes may provide a potential therapeutic strategy to counteract inflammation in the development of prediabetes and early onset of insulin resistance and T2D. Our data indicate that altered mtDNA content in lymphocytes gives rise to inflammation and insulin resistance, intrinsically leading to early stage of diabetes progression in the long run.
Mitochondrial dysfunction and oxidative stress are among the common denominators contributing to the pathogenesis of prediabetes states such as insulin resistance and IFG, as previously reported [19]. Several metabolic dysregulations in prediabetes are also coupled with several microvascular complications including retinopathy, nephropathy and neuropathy [20,21,22]. In this regard, a number of experimental and epidemiological studies have looked into the metabolic role of mtDNA level in predicting the risks for diabetes complications. In a cross-sectional community-based study, higher mtDNA copy number was associated with decreased prevalence of microalbuminuria [23]. Another report revealed that peripheral blood mtDNA content in T2D individuals with diabetic nephropathy was substantially altered compared to those without nephropathy, proposing that oxidative stress might play important roles in the development of this diabetic complication [24]. Moreover, mtDNA damage in rat retina was found to provide subsequent increase in the risk factors associated with diabetic retinopathy in streptozotocin-diabetic rats [25]. The compromised mitochondrial electron transport chain systems with severe loss in metabolic memory are among the consequences of mtDNA damage in development of diabetes complications. Similarly, another study showed that mtDNA damage in both isolated retina endothelial cells from rats and human contributes to a significant alteration in mitochondrial biogenesis with accumulation of superoxide radicals [26]. Furthermore, mtDNA damage in peripheral neuron with oxidative stress has been proposed as a unifying mechanism for diabetic neuropathy [27, 28]. Such oxidative damage in mitochondrial genome with irregular mitochondrial trafficking in neurons may predispose to increased risk of neurodegeneration in diabetes [29]. Consistently, the current study showed that IFG patients with microvascular complications have decreased mtDNA content in lymphocytes compared to those without complications. Our data suggest that mtDNA content in lymphocytes may be a useful indicator of microvascular complications in IFG participants.
It has been demonstrated that decreased mtDNA content in adipose tissue was partly mediated by hyperinsulinemia and insulin resistance [30]. Several reports indicated the positive association between reduced mtDNA in peripheral blood monocytes and early phase of T2D complications [31]. Decreased mtDNA copy number was negatively associated with the fasting glucose level, diastolic BP and waist–hip circumference ratio of insulin resistance and T2D patients [32]. Sustained damage in mtDNA content may negatively affects various complications in diabetic retinopathy, and these deleterious effects on mitochondrial genomes can be attenuated by over-expression of manganese superoxide dismutase [33]. Other studies also found that altered mtDNA copies of leukocytes were intangibly linked to glucose dysregulation, leading to hyperglycemia and diabetes incidence [32, 34]. Consistent with these, several lines of evidence suggest the relationship between mitochondrial genome mutation and hypertension in a Chinese population [35, 36]. By contrast to the previous studies, another report revealed that reduced peripheral blood mtDNA content was not a risk factor for T2D progression in the offspring of patients with early onset T2D [37]. In this report, we observed such significance correlations between reduced mtDNA content in lymphocytes and the high level of fasting glucose and HOMA-IR after confounding factors adjustment. This phenotype of reduced mtDNA content was found to be increasing in those individuals with hypertension. In agreement to these, the present study appears to clarify the positive relationship between mtDNA content in lymphocytes and IFG complications. What is more, our study provides a new layer of evidence implicating the role of mtDNA content in lymphocytes of IFG participants with insulin resistance.
HDL-cholesterol is one of the useful clinical markers to predict the development of numerous metabolic disorders. A number of epidemiological studies revealed that low levels of HDL-cholesterol were consistently linked to the increased risk of T2D and cardiovascular-related complications [38, 39]. However, whether the increased plasma HDL-cholesterol level is causally predisposed to the risk of T2D and its comorbidities is yet to be determined. The association between mtDNA content and HDL-cholesterol level in diseased groups was previously investigated where reduced mtDNA content in peripheral blood monocytes was found in patients at an earlier age of type 2 diabetes onset with lower HDL level [31]. Furthermore, decreased level of HDL-cholesterol in individuals with hyperlipidemia and metabolic syndromes has been linked to the depleted mtDNA content in leukocytes [40]. In light of these previous observations, along with the current observation here, a significant negative correlation between mtDNA content in lymphocytes and plasma HDL-cholesterol in IFG patients was observed. For that reason, we are suggesting that reduced mtDNA content in lymphocytes combined with HDL-cholesterol measurement contributes to the improved investigation in classifying IFG individuals at risk for developing insulin resistance and T2D.
A causative link has been postulated between inflammation and insulin resistance. It was proved that the activation of induced NF-κB activity in diabetic animals with microvascular complications was partly regulated by mitochondrial genomes in response to the mitochondrial-mediated cell death pathways [41]. This inflammatory activation possibly includes different mechanism involving N-terminal kinase (JNK) pathways, pro-inflammatory cytokines release and macrophage recruitment [12, 42]. Of note, prolonged mtDNA damage may lead to impaired mitophagy process, leading to activation of inflammatory signaling [43]. This effect seems to predominantly result from altered mtDNA content and other deleterious effects exhibited by increasing DNA oxidative damage in numerous peripheral tissues [12]. In this context, it has been shown that the development of insulin resistance by decreased tyrosine kinase activity of the insulin receptor was likely mediated by excess release of TNF-α [44]. High level of circulating IL-6 may lead to insulin resistance, T2D and other metabolic dysregulations [44]; however, IL-6 displays pleiotropic functions in a tissue‐specific and physiological context‐dependent manner [45]. Increased level of circulating IL-10 was proved to enhance skeletal muscle insulin sensitivity by attenuating obesity-associated macrophage infiltration, inflammation and aging mediated-insulin resistance [46, 47]. In the present study, we observed amplified levels of TNF-α and IL-6 while decreased levels of IL-10 that were coincided with reduced mtDNA content and higher degree of HOMA-IR index among IFG patients. Our data indicated that increased levels of TNF-α and IL-6 substantially contribute to insulin resistance and early onset of IFG stages. Collectively, these data u support the contention that decreased mtDNA content in the IFG group is associated with the impaired glucose metabolism, insulin resistance and inflammation.
Our study has some limitations. We only assessed the association between mtDNA content in lymphocytes and other clinical risk factors at a single time point rather than multiple time points. The measurement of samples at multiple time points allows any prognostic value associated with the disease progression to be determined. Moreover, the current study adopted the cross-sectional design that did not allow the evaluation of such causality-involved associations. In addition, a larger sample size of populations is imperatively needed to verify and confirm our findings on the cutoff value for mtDNA content in lymphocytes to further predict the development of IFG and insulin resistance with other potential confounders. An experimental design with a large prospective cohort study would be of value to casually indicate the incidence of mtDNA damage in lymphocytes of IFG participants. Nevertheless, we have determined the possible disease risk factors that can be linked with altered mtDNA content in lymphocytes, thus providing statistical control for important confounding factors in our study population of interest.
In summary, we show evidence of multiple associations between reduced mtDNA content in lymphocytes and other clinical risk factors including inflammation and insulin resistance in IFG. We demonstrated that altered mtDNA content in lymphocytes was observed in IFG individuals with microvascular complications compared to those without complications. It is of importance to recognize that decreased mtDNA content was comparatively due to the prolonged inflammatory signaling activation induced by hyperglycemia in IFG participants. This finding is consistent with our previous report where impaired mitochondrial activities lead to the activation of subclinical inflammation [15], leading to insulin resistance and type 2 diabetes. The current study revealed that individuals with IFG exhibit a cascade of events that include depleted level of mtDNA content in lymphocytes with other increasing metabolic dysregulations, signifying such impairment in mitochondrial genomes precedes T2D rather than vice versa. Despite the potential pathophysiological and clinical significance to IFG, to date, there are no reported studies for measuring perturbations in mtDNA content in easily accessible human lymphocytes of individuals with IFG. To the best of our knowledge, this is the first cross-sectional study addressing the relationship between reduced mtDNA content in lymphocytes, inflammation, insulin resistance and other established clinical risk factors in individuals with IFG. Therefore, an understanding of preventive and therapeutic intervention for prediabetes patients must take into account the expression and activation of these retrograde signaling pathways in lymphocytes in order to strategize the best complementary approach against the development of such metabolic perturbations.
References
International Diabetes Federation. IDF Diabetes Atlas. 7th ed. Federation ID, editor. Brussels, Belgium: International Diabetes Federation; 2017.
Tabák AG, Herder C, Rathmann W, Brunner EJ, Kivimäki M. Prediabetes: a high-risk state for diabetes development. Lancet. 2012;379:2279–90.
Lu H, Koshkin V, Allister EM, Gyulkhandanyan AV, Wheeler MB. Molecular and metabolic evidence for mitochondrial defects associated with beta-cell dysfunction in a mouse model of type 2 diabetes. Diabetes. 2010;59:448–59.
Petersen KF, Dufour S, Befroy D, Garcia R, Shulman GI. Impaired mitochondrial activity in the insulin-resistant offspring of patients with type 2 diabetes. N Engl J Med. 2004;350:664–71.
Serradas P, Giroix M-H, Saulnier C, Gangnerau M-N, Borg LA, Welsh M, et al. Mitochondrial deoxyribonucleic acid content is specifically decreased in adult, but not fetal, pancreatic islets of the Goto-Kakizaki rat, a genetic model of noninsulin-dependent diabetes. Endocrinology. 1995;136:5623–31.
Antonetti DA, Reynet C, Kahn CR. Increased expression of mitochondrial-encoded genes in skeletal muscle of humans with diabetes mellitus. J Clin Invest. 1995;95:1383.
Widlansky ME, Wang J, Shenouda SM, Hagen TM, Smith AR, Kizhakekuttu TJ, et al. Altered mitochondrial membrane potential, mass, and morphology in the mononuclear cells of humans with type 2 diabetes. Transl Res. 2010;156:15–25.
Shenouda SM, Widlansky ME, Chen K, Xu G, Holbrook M, Tabit CE, et al. Altered mitochondrial dynamics contributes to endothelial dysfunction in diabetes mellitus. Circulation. 2011;124:444–53.
Perl A, Gergely P, Nagy G, Koncz A, Banki K. Mitochondrial hyperpolarization: a checkpoint of T-cell life, death and autoimmunity. Trends Immunol Elsevier. 2004;25:360–7.
Kim H-S, Park K-G, Koo TB, Huh S, Lee I-K. The modulating effects of the overexpression of uncoupling protein 2 on the formation of reactive oxygen species in vascular cells. Diabetes Res Clin Pract. 2007;77:S46–8.
Khan S, Raghuram GV, Bhargava A, Pathak N, Chandra DH, Jain SK, et al. Role and clinical significance of lymphocyte mitochondrial dysfunction in type 2 diabetes mellitus. Transl Res. 2011;158:344–59.
Bakar MHA, Kian Kai C, Wan Hassan WN, Sarmidi MR, Yaakob H, Zaman Huri H. Mitochondrial dysfunction as a central event for mechanisms underlying insulin resistance: the roles of long chain fatty acids. Diabetes Metab Res Rev. 2015;31:453–75.
Malik AN, Czajka A. Is mitochondrial DNA content a potential biomarker of mitochondrial dysfunction? Mitochondrion. 2013;13:481–92.
American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care. 2009;33:S62.
Abu Bakar MH, Sarmidi MR. Association of cultured myotubes and fasting plasma metabolite profiles with mitochondrial dysfunction in type 2 diabetes subjects. Mol BioSyst. 2017;13:1838–53.
Association AD. Standards of medical care in diabetes–2014. Diabetes Care. 2014;37:S14–80.
Piątkiewicz P, Czech A, Tatoń J. Glucose transport in human peripheral blood lymphocytes influenced by type 2 diabetes mellitus. Arch Immunol Ther Exp (Warsz). 2007;55:119–26.
Kramer PA, Ravi S, Chacko B, Johnson MS, Darley-Usmar VM. A review of the mitochondrial and glycolytic metabolism in human platelets and leukocytes: implications for their use as bioenergetic biomarkers. Redox Biol. 2014;2:206–10.
Hische M, Larhlimi A, Schwarz F, Fischer-Rosinský A, Bobbert T, Assmann A, et al. A distinct metabolic signature predicts development of fasting plasma glucose. J Clin Bioinforma. 2012;2:3.
Dagenais GR, Gerstein HC, Holman R, Budaj A, Escalante A, Hedner T, et al. Effects of ramipril and rosiglitazone on cardiovascular and renal outcomes in people with impaired glucose tolerance or impaired fasting glucose: results of the Diabetes REduction Assessment with ramipril and rosiglitazone Medication (DREAM) trial. Diabetes Care. 2008;31:1007–14.
Garber AJ, Abrahamson MJ, Barzilay JI, Blonde L, Bloomgarden ZT, Bush MA, et al. Consensus statement by the American Association of Clinical Endocrinologists and American College of Endocrinology on the comprehensive type 2 diabetes management algorithm–2016 executive summary. Endocr Pract. 2016;22:84–113.
Handelsman Y, Bloomgarden ZT, Grunberger G, Umpierrez G, Zimmerman RS, Bailey TS, et al. American Association of Clinical Endocrinologists and American College of Endocrinology–clinical practice guidelines for developing a diabetes mellitus comprehensive care plan–2015. Endocr Pract. 2015;21:1–87.
Lee JE, Park H, Ju YS, Kwak M, Kim J-I, Oh HY, et al. Higher mitochondrial DNA copy number is associated with lower prevalence of microalbuminuria. Exp Mol Med. 2009;41:253–8.
Malik AN, Shahni R, Iqbal MM. Increased peripheral blood mitochondrial DNA in type 2 diabetic patients with nephropathy. Diabetes Res Clin Pract. 2009;86:e22–4.
Madsen-Bouterse SA, Mohammad G, Kanwar M, Kowluru RA. Role of mitochondrial DNA damage in the development of diabetic retinopathy, and the metabolic memory phenomenon associated with Its progression. Antioxid Redox Signal. 2010;13:797–805.
Santos JM, Tewari S, Goldberg AFX, Kowluru RA. Mitochondrial biogenesis and the development of diabetic retinopathy. Free Radic Biol Med. 2011;51:1849–60.
Vincent AM, Calabek B, Roberts L, Feldman EL. Biology of diabetic neuropathy. Handb Clin Neurol. 2012;115:591–606.
Yagihashi S, Mizukami H, Sugimoto K. Mechanism of diabetic neuropathy: where are we now and where to go? J Diabetes Investig. 2011;2:18–32.
Fernyhough P, Roy Chowdhury SK, Schmidt RE. Mitochondrial stress and the pathogenesis of diabetic neuropathy. Expert Rev Endocrinol Metab. 2010;5:39–49.
Bogacka I, Xie H, Bray GA, Smith SR. Pioglitazone induces mitochondrial biogenesis in human subcutaneous adipose tissue in vivo. Diabetes. 2005;54:1392–9.
Wong J, McLennan SV, Molyneaux L, Min D, Twigg SM, Yue DK. Mitochondrial DNA content in peripheral blood monocytes: relationship with age of diabetes onsetand diabetic complications. Diabetologia. 2009;52:1953–61.
Lee HK, Song JH, Shin CS, Park DJ, Park KS, Lee KU, et al. Decreased mitochondrial DNA content in peripheral blood precedes the development of non-insulin-dependent diabetes mellitus. Diabetes Res Clin Pract. 1998;42:161–7.
Madsen-Bouterse SA, Zhong Q, Mohammad G, Ho Y-S, Kowluru RA. Oxidative damage of mitochondrial DNA in diabetes and its protection by manganese superoxide dismutase. Free Radic Res. 2010;44:313–21.
Weng S-W, Lin T-K, Liou C-W, Chen S-D, Wei Y-H, Lee H-C, et al. Peripheral blood mitochondrial DNA content and dysregulation of glucose metabolism. Diabetes Res Clin Pract. 2009;83:94–9.
Lu Y, Xiao T, Zhang F, Chen Y, Liu Y, Li Y, et al. Effect of mitochondrial tRNA Lys mutation on the clinical and biochemical characteristics of Chinese essential hypertensive subjects. Biochem Biophys Res Commun. 2014;454:500–4.
Zhu H-Y, Wang S-W, Martin LJ, Liu L, Li Y-H, Chen R, et al. The role of mitochondrial genome in essential hypertension in a Chinese Han population. Eur J Hum Genet. 2009;17:1501–6.
Singh R, Hattersley AT, Harries LW. Reduced peripheral blood mitochondrial DNA content is not a risk factor for Type 2 diabetes. Diabet Med. 2007;24:784–7.
Wilson PWF, Meigs JB, Sullivan L, Fox CS, Nathan DM, D’Agostino RB. Prediction of incident diabetes mellitus in middle-aged adults: the Framingham Offspring Study. Arch Intern Med. 2007;167:1068–74.
Schmidt MI, Duncan BB, Bang H, Pankow JS, Ballantyne CM, Golden SH, et al. Identifying individuals at high risk for diabetes. Diabetes Care. 2005;28:2013–8.
Liu C-S, Kuo C-L, Cheng W-L, Huang C-S, Lee C-F, Wei Y-H. Alteration of the copy number of mitochondrial DNA in leukocytes of patients with hyperlipidemia. Ann N Y Acad Sci. 2005;1042:70–5.
Romeo G, Liu W-H, Asnaghi V, Kern TS, Lorenzi M. Activation of nuclear factor-κB induced by diabetes and high glucose regulates a proapoptotic program in retinal pericytes. Diabetes. 2002;51:2241–8.
Esser N, Legrand-Poels S, Piette J, Scheen AJ, Paquot N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res Clin Pract. 2014;105:141–50.
Malik AN, Parsade CK, Ajaz S, Crosby-Nwaobi R, Gnudi L, Czajka A, et al. Altered circulating mitochondrial DNA and increased inflammation in patients with diabetic retinopathy. Diabetes Res Clin Pract. 2015;110:257–65.
Emanuela F, Grazia M, de Marco R, Maria Paola L, Giorgio F, Marco B. Inflammation as a link between obesity and metabolic syndrome. J Nutr Metab. 2012;2012:476380.
Kim J, Bachmann RA, Chen J. Interleukin-6 and insulin resistance. Vitam Horm. 2009;80:613–33.
Hong E-G, Ko HJ, Cho Y-R, Kim H-J, Ma Z, Yu TY, et al. Interleukin-10 prevents diet-induced insulin resistance by attenuating macrophage and cytokine response in skeletal muscle. Diabetes. 2009;58:2525–35.
Dagdeviren S, Jung DY, Friedline RH, Noh HL, Kim JH, Patel PR, et al. IL-10 prevents aging-associated inflammation and insulin resistance in skeletal muscle. FASEB J. 2016;31:701–10.
Acknowledgements
The authors would like to thank the Ministry of Higher Education, Malaysia, under the Fundamental Research Grant Scheme (Vot. No: R.J130000.7809.4F284), Universiti Teknologi Malaysia, University of Malaya Medical Centre and Universiti Sains Malaysia Short-Term Grant (Ref. No: 034/PTEKIND/6313329) for financial and technical support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval statement
The study was approved by the institutional medical ethics review board of the University Malaya Medical Centre (UMMC), Malaysia (reference number: 201495.95).
Human and animal rights
All procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (institutional and national) in compliance with the 1975 Declaration of Helsinki of the World Medical Association. This article does not contain any studies with animals performed by any of the authors.
Informed consent
All participants underwent the informed consent process.
Rights and permissions
About this article

Cite this article
Abu Bakar, M.H., Hairunisa, N. & Zaman Huri, H. Reduced mitochondrial DNA content in lymphocytes is associated with insulin resistance and inflammation in patients with impaired fasting glucose. Clin Exp Med 18, 373–382 (2018). https://doi.org/10.1007/s10238-018-0495-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10238-018-0495-4